
Abstract
The bone morphogenetic proteins (BMPs), which are involved in bone formation and repair, play an important role in tissue engineering. For example, BMP-9 and BMP-2, which are members of different BMP subfamilies, are osteoinductive factors. However, several studies have recently shown that BMP-9 is more osteogenic than BMP-2. We have previously shown that fetal bovine serum (FBS) strongly enhances the osteoblast differentiation of murine preosteoblasts (MC3T3-E1) to BMP-9 but not to BMP-2. This effect is mimicked by IGF-2, which primarily activates the PI3K/Akt pathway, but how Akt phosphorylation sites are implicated in such differentiation is unclear. The effects of BMP-9 and BMP-2 with or without FBS or IGF-2 on Akt phosphorylation sites and subsequent osteoblastic differentiation were determined, respectively, by western blot analysis and alkaline phosphatase activity measurements. The involvement of phosphorylated Akt at Thr308 and/or Ser473 on BMP-mediated osteoblast differentiation was further studied using specific inhibitors. In MC3T3-E1 incubated with or without FBS, BMP-9 and BMP-2 activate Akt on Ser473 and Thr308 very differently in a time and dose-dependent manner. Using inhibitors specific to each Akt phosphorylation site, we showed that both Ser473 and Thr308 must be phosphorylated for BMP-9 and/or IGF-2-induced osteoblast differentiation, whereas BMP-2 requires phosphorylation of only Ser473. Furthermore, cells stimulated with BMP-2 in the presence of FBS require the phosphorylation of Akt at Ser473 and the dephosphorylation of Akt at Thr308 to increase the osteoblast differentiation with alkaline phosphatase activity similar to that of BMP-9 plus FBS. These results provide a better understanding into how BMP-9 induces osteoblast differentiation and its synergy with IGF-2 at the signaling level. This knowledge is essential for preparing the serum-free osteogenic media required for bone tissue engineering or developing growth factor delivery systems to improve bone formation.
Introduction
T
BMPs are classified in subfamilies such as BMP-2/BMP-4 and BMP-9/BMP-10, which depend on their sequence homology.4,7 BMPs act on cells through their binding to type I (ALK1 to ALK-7) and type II (BMPRI, ActRIA, ActRIIA) Ser/Thr kinase receptors. For example, BMP-2 is known to mainly interact with ALK3 and ALK6 unlike BMP-9, whose highest binding affinity is for type I ALK1 followed by BMPRII receptors.8–10 The interaction of BMPs with their receptor can then trigger several signaling cascades such as Smad1/5/8 and MAPK leading to osteoblastic differentiation.4,5,11–13 As BMPs are expensive to produce and purify, using a more osteogenic protein like BMP-9 will be helpful to develop defined medium or delivery systems while minimizing their costs.
As part of our ongoing studies on the response of bone-forming cells to BMP-9, we have demonstrated that adding fetal bovine serum (FBS) dramatically potentiates the effect of BMP-9 on MC3T3-E1 murine preosteoblasts differentiation by reducing the half-maximal effective concentration (EC50) four to sixfold as shown by alkaline phosphatase (ALP) activity, while having no influence on the EC50 of BMP-2. 5 We have also identified a growth factor, IGF-2, that mimics the effect of FBS on BMP-9-induced ALP activity. IGF-2 at 100 ng/mL can activate Smad1/5/8 and p38 MAPK, while it reduces the concentration of phosphorylated ERK1/2.
However, the way in which BMP-9-mediated bone cell response is increased by IGF-2 is poorly known. Chen et al. 14 also observed the synergy between IGF-2 and BMP-9 on murine mesenchymal stromal cells (C3H10T1/2) using adenovirus. They used a specific phosphoinositide-3-kinase (PI3K) inhibitor (LY294002) to show that the PI3K/Akt pathway seems to be essential for the BMP-9-mediated osteoblast differentiation with and without IGF-2. However, there is not yet any evidence supporting that Akt is directly implicated. Akt is usually maintained in its inactivated form and is activated by PI3K, which is itself activated by many external stimuli. 15 Akt and the pyruvate dehydrogenase lipoamide kinase isozyme 1 (PDK1) then migrate together to the plasma membrane where they interact with phosphatidylinositol. PDK1 phosphorylates Akt at its catalytic site (Thr308), triggering a conformational change in the PH domain, which dramatically increases its activity. 15 However, the Ser473 in the hydrophobic motif of Akt must also be phosphorylated for Akt to be fully active. Studies on many cell types (HEK293T, HeLa, HT29, A549, and 3T3-L1) indicate that this phosphorylation is catalyzed by mTORC2.16–18 Akt also promotes the BMP-2-mediated Smad-dependent differentiation of murine stromal cells toward an osteoblast lineage.19,20 Activation of the BMP-mediated MAPK (ERK1/2) and Smad1/5/8 pathways also seems to be upregulated by PI3K/Akt. 21 The synthesis of BMP-2 and ALP proteins requires the activation of PI3K and ERK1/2.14,20–23 However, no reports on the involvement of phosphorylated Akt on Thr308 (pAkt-Thr308) and on Ser473 (pAkt-Ser473) in osteoblast differentiation induced by equimolar concentrations of BMP-9 and BMP-2 have yet been published.
This study investigates the effects of FBS and IGF-2 on the activation of specific phosphorylation sites (catalytic domain Thr308 and regulatory domain Ser473) in the Akt of murine MC3T3-E1 preosteoblasts stimulated with BMP-9 or BMP-2. It was designed to show how Akt is implicated in BMP-mediated osteoblast differentiation by examining its phosphorylation state (pAkt-Thr308 and pAkt-Ser473) and ALP activity using inhibitors to block the overall activation of Akt (MK-2206) or target separate Akt phosphorylation sites. We used the PDK1 inhibitor BX-795 for pAkt-Thr308 and mTORC inhibitors (rapamycin and KU-0063794) for pAkt-Ser473.24–26 The results obtained show new light on the role of the Akt pathway in BMP-mediated osteoblast differentiation and will be helpful to design defined medium containing BMP-2 or BMP-9 without serum.
Materials and Methods
Materials
Recombinant carrier-free human BMP-9 and BMP-2 (from Chinese hamster ovarian cells) were purchased from R&D Systems. Lyophilized IGF-2 was recovered as specified by the manufacturer's instructions (PeproTech). The inhibitors of AKT-1/2/3 (MK-2206, S1078), mTORC1/2 (rapamycin, S1039), mTORC2 (KU00063794), and PDK-1 (BX-795, S1274) were purchased from Selleckchem. Primary antibodies directed against pAkt-Thr308 and pAkt-Ser473 were purchased from Cell Signaling (New England Biolabs), and primary antibodies directed against β-actin were from Sigma (Sigma).
Cell culture
Murine calvarial MC3T3-E1 preosteoblasts, subclone 14 (CRL-2594™; ATCC®) were cultivated between passages 4 and 11 as previously described. 5 A single lot of serum was used for the entire study.
Western blot and spot densitometric analysis
MC3T3-E1 preosteoblasts were cultured to 80% confluence and then starved of serum for 24 h. For inhibitor assays, these cells were incubated for 1 h with Akt-1/2 inhibitor (MK-2206, 2.5 μM), PDK1 inhibitor (BX-795, 2.5 μM), or mTORC2 inhibitor (rapamycin, 100 nM, or KU-0063794, 5 μM), followed by stimulation with BMPs (1 nM) ± FBS (10% v/v) or ± IGF-2 (100 ng/mL) for 1 h. They were then lyzed as previously described 5 and equal amount of proteins for each condition was subjected to western blot analyses using rabbit antibodies against pAkt-Thr308 (diluted 1:1000) and pAkt-Ser473 (diluted 1:1000) and mouse antibodies against β-actin (diluted 1:1000). Bands were revealed using secondary antibodies (goat anti-rabbit IgG or goat anti-mouse IgG) coupled to HRP (diluted 1/10,000) and enhanced chemiluminescence (Amersham ECL Prime Western Blotting detection Reagent; GE HealthCare Ca), with β-actin as control. The resulting films were scanned and the intensities of the bands were analyzed by densitometry using image analysis software (AlphaEase® v4.0 Image Analysis software). The integrated densities of pAkt-Thr308 and pAkt-Ser473 bands were normalized to those of the appropriate β-actin bands. Each experiment was performed independently at least twice.
Cytotoxicity assay and morphology
MC3T3-E1 preosteoblasts were cultured to 80% confluence on 24-well plates 5 and then placed in serum-free medium for 24 h. The resulting cells were incubated for 72 h with various concentrations of Akt-1/2 specific inhibitors in DMEM without phenol red: MK-2206 (0, 1, 2.5, 5, 10, 25 μM), BX-795 (0, 1, 2.5, 5, 10, 25 μM), KU-0063794 (0, 1, 2.5, 5, 10, 25 μM), and rapamycin (0, 1, 10, 25, 50, 100 nM). The treated cells were incubated with MTS reagent (CellTiter 96® aqueous Solution; Promega) for 2 h at 37°C according to the manufacturer's instructions. Supernatants were collected and the absorbance at 490 nm was measured using a filter-based multimode Synergy spectrophotometer (Biotek). Cells were then fixed by incubation in 3% (m/v) paraformaldehyde for 15 min, permeabilized with Triton X-100® (0.5% v/v) for 5 min, and saturated with bovine serum albumin (1% m/v) for 30 min at 37°C. Cell nucleus was stained for 30 min by Hoechst (Life Technologies) (5 μg/mL) and actin cytoskeleton for 30 min with rhodamine-phalloidin (Probes by Life Technologies) (1/200). Four representative pictures of the cell density (Hoechst stained) per well were taken at a magnification of 10× using an Eclipse TE200s equipped with a Retiga 1300R camera (Nikon). Cells were counted using a MATLAB application developed by our group (MATLAB 2007B; MathWorks). Briefly, nucleus stained images, which have a high contrast, were binarized. Each nucleus was then labelled using a well-known MATLAB built-in function (“bwlabel,” Image Processing Toolbox), which assigns labels according to a defined pixel interconnectivity. Giving a normal average area per nucleus to avoid artifacts, the amount of labels within the average range gives the number of cells present in the picture. Cell density was then estimated as the average number of cells divided by the area of the picture analyzed (0.02 cm2/picture). Cell morphology was examined using representative randomly selected pictures of each condition using an EVOS FL auto® microscope (Life Technologies). These images were treated and merged using an image analysis MATLAB program (MATLAB 2007B; MathWorks). Each experiment was performed at least thrice in duplicate.
ALP activity
MC3T3-E1 preosteoblasts were cultured as described above, starved of serum for 24 h, and incubated with inhibitors of Akt-1/2 (MK-2206, 2.5 μM), PDK1 (BX-795, 2.5 μM), or mTORC2 (rapamycin, 100 nM and KU-0063794, 5 μM) for 1 h and then stimulated with BMPs (1 nM) ± FBS (10% v/v) or ± IGF-2 (100 ng/mL) for 72 h. ALP activity was assayed as previously described 5 using SensoLyte® pNPP ALP Assay Kit (AnaSpec) according to the manufacturer's instructions. ALP results were normalized to the cell density, then to the control. Briefly, cell nucleus was stained with Hoechst for 25 min (5 μg/mL) (Life Technologies) and four pictures were taken for each condition (0.02 cm2/picture) using an Eclipse TE200-S microscope coupled to a 10× objective and a CCD camera (Retiga 1300R; Nikon). The mean cell density (cell/cm2) was determined using an image analysis program developed with MATLAB software R2007b. 5 Each experiment was performed thrice in duplicate.
Statistical analyses
Statistical analyses were performed using Excel (Excel 2010®). Analysis of variance was performed using a 95% confidence limit followed by a Tukey–Kramer post hoc test to determine significant differences between treatments. Only differences with a p < 0.05 were considered significant.
Results and Discussion
Several studies highlight the importance of using defined medium containing growth factors for bone and cartilage tissue applications.1,2 For example, Mendes et al. have performed a factorial design with five growth factors (TGFbeta1, FGF2, BMP-2, BMP-6, and GDF-5) to identify the best combination that induced the chondrogenic differentiation of human periosteum-derived mesenchymal stem cells. 2 However, unless such factorial design gives many insights about the best growth factor combination in a given situation, a better understanding of the signaling pathway induced by these growth factors can also help to select them by preventing any antagonist signal transduction. We have previously shown that IGF-2 can mimic the effect of FBS on BMP-9 cell response. Indeed, FBS strongly reduced the EC50 of BMP-9, as shown by the gene expression of osteogenic markers and ALP activity, while it had no influence on EC50 of BMP-2. 5 Since IGF-2 mainly signals through the Akt pathway, 27 we first investigated the effect of BMP-9 and BMP-2 on Akt phosphorylation sites with or without FBS.
BMP-9 and BMP-2 activate the Akt signaling pathway differently
The dose–response effect of BMP-9 and BMP-2 on Akt activation at both phosphorylation sites in bone cells is unknown. Therefore, we first studied the effects of several concentrations of BMP-9 or BMP-2 on the activation of Akt at its two phosphorylation sites (Thr308 and Ser473) after incubation for 1 h (Fig. 1). A thick band of pAkt-Ser473 was detected in lysates from cells incubated for 1 h in serum-free medium supplemented with BMP-9 at 0.1 nM. By contrast, the band was thin when the concentration of BMP-9 reached 8 nM. A reverse effect of BMP-9 was noticed for pAkt at Thr308. At 8 nM of BMP-9, a thick band of pAkt-Thr308 was detected, while it was very thin at 0.1 nM (Fig. 1A–C). Interestingly, the exact opposite pattern of Akt phosphorylation occurred when cells were stimulated with BMP-2 in serum-free medium (Fig. 1A–C).
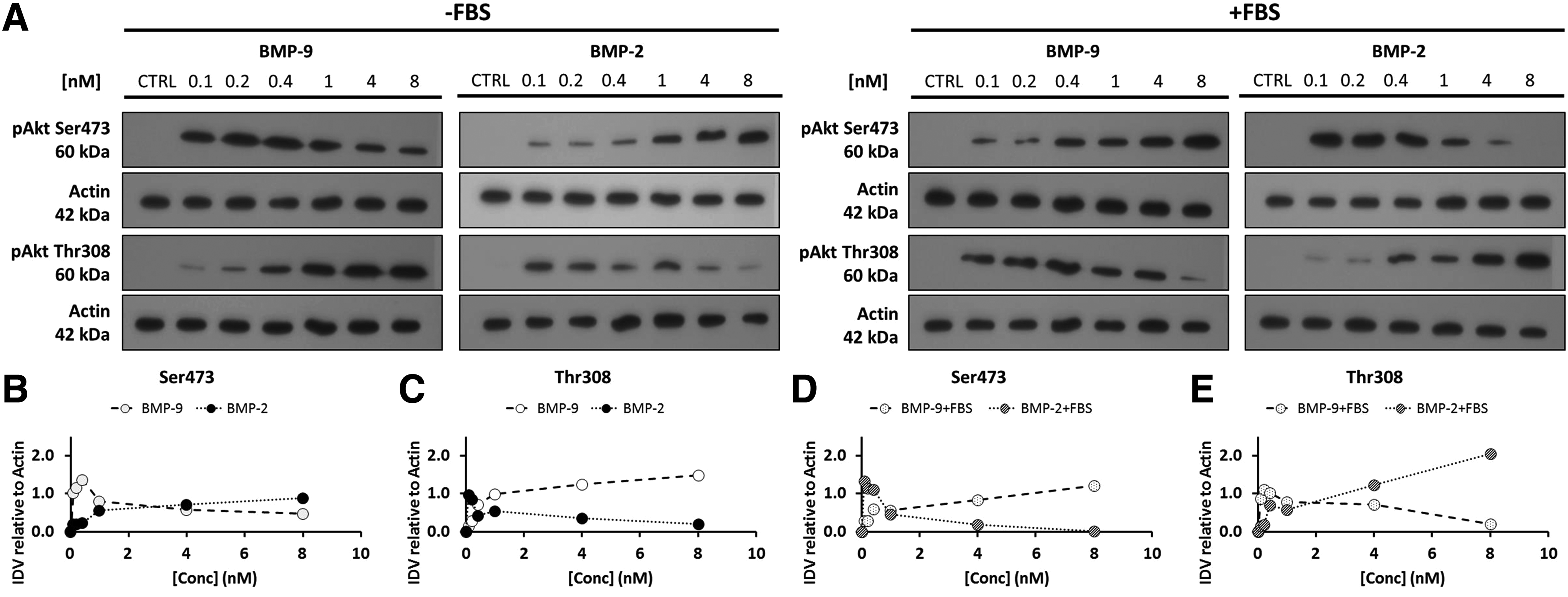
Effect of BMP-9 and BMP-2 on the phosphorylation of Akt (Ser473 and Thr308) with or without FBS.
In addition, cells stimulated with BMP-9 or BMP-2 in the presence of FBS responded exactly opposite to cells stimulated without FBS (Fig. 1A, D, E). Incubation with BMP-9 for 1 h appeared to increase pAkt at Ser473 and decrease pAkt at Thr308, while incubation with BMP-2 decreased pAkt at Ser473 and increased pAkt at Thr308. Indeed, pAkt-Ser473 was not detected in cells incubated with 8 nM BMP-2 plus FBS, while the band of pAkt-Thr308 was very intense as confirmed by the densitometric analyses (Fig 1D, E). These results clearly indicate that after incubation for 1 h BMP-9 and BMP-2 act differently on Akt signaling in murine preosteoblasts and that FBS also dramatically affects this activation. Mukherjee and Rotwein reported that multipotent murine C3H10T1/2 cells in 10% (v/v) fetal calf serum and stimulated with BMP-2 (8 nM) for 1–10 days showed no phosphorylation of Akt on Ser473. 19 However, other research groups indicate that the PI3K/Akt pathway is activated by BMP-2 and that this activation is necessary for osteoblast differentiation.20,21 Transient Akt pathway activation could also contribute to the observed effect. For instance, Ghosh-Choudhury et al. have shown that C2C12 myoblasts cultivated in serum-free medium supplemented with ∼15 nM of BMP-2 induced a time-dependent effect on Akt pathway. 20
We examined the time dependence of phosphorylation on both Akt phosphorylation sites by stimulating MC3T3-E1 preosteoblasts with equimolar concentrations of BMP-9 or BMP-2 (0, 0.1, 1, and 8 nM) with or without FBS for up to 240 min (Fig. 2A, B). The lower concentrations took more time to increase or decrease the phosphorylation of Akt than did the higher concentrations. The amount of pAkt-Thr308 in preosteoblasts stimulated with 0.1 nM BMP-9 without FBS increased with time from 30 min, while the amount of this phosphorylated Akt was higher within 15 min in cells stimulated with 1 or 8 nM (Fig. 2A). The general tendency was the same as that observed for the dose–response. For instance, BMP-9 increased pAkt-Thr308 in time with a faster effect with high concentration, while decreasing the activation of pAkt-Ser473. Again, the opposite effect was also observed for cells stimulated in the presence of FBS in comparison to serum-free results (Fig. 2B).
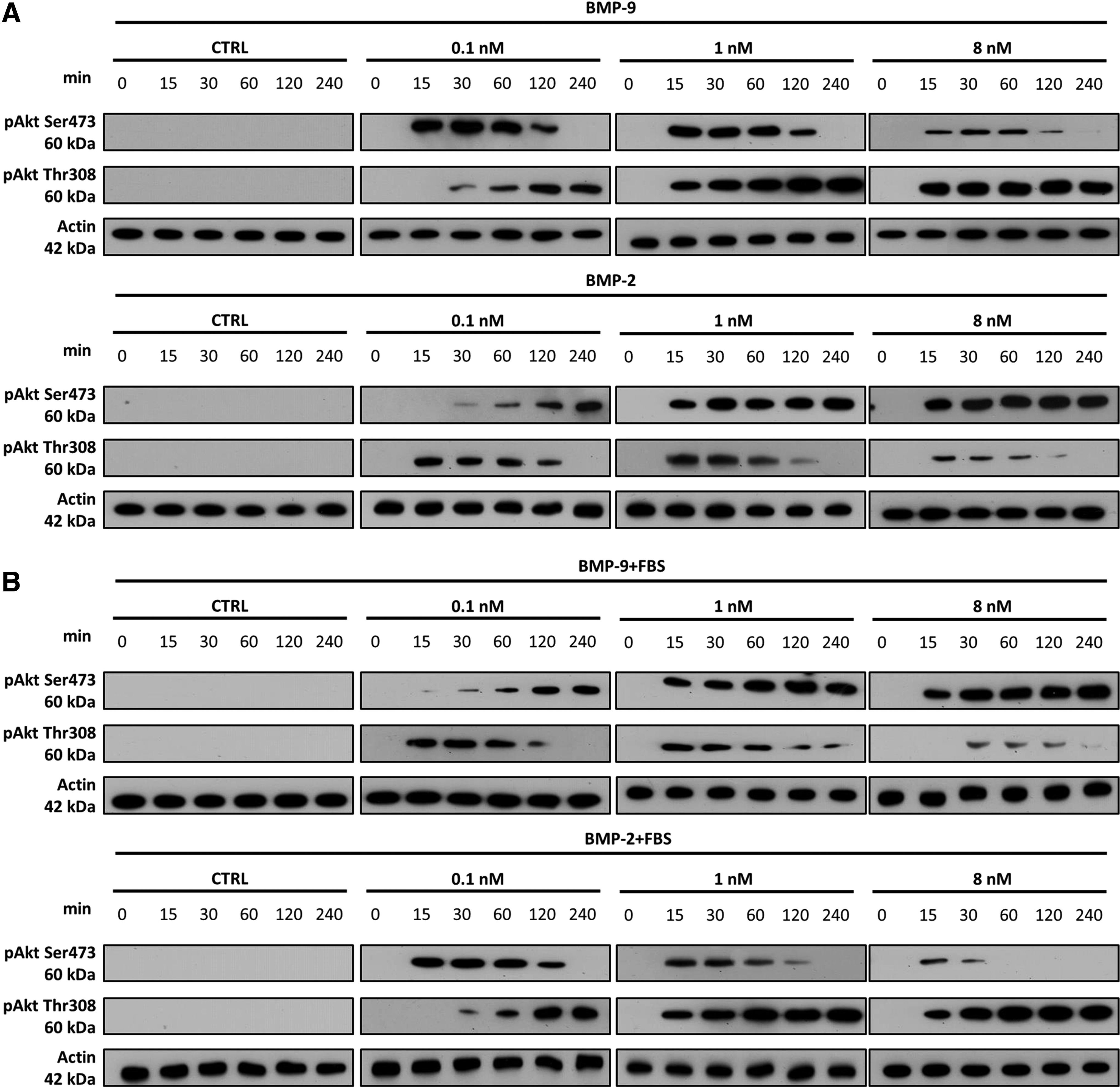
Time course and dose–response studies of Akt phosphorylation induced by BMP-9 or BMP-2 with or without FBS. Western blots showing the activation of Akt by phosphorylation of Thr308 and Ser473 and β-actin in MC3T3-E1 preosteoblasts incubated with 0.1, 1, and 8 nM BMP-9 or BMP-2 for 0, 0.25, 0.5, 1, 2, and 4 h
Phosphorylation kinetics on both sites differed depending on the experimental conditions (type of BMPs and the presence of serum). They also confirmed that the BMP concentration is important for the activation of Akt at both phosphorylation sites. Since Thr308 is the catalytic site of Akt, these results also suggest that high doses of BMP-9 without FBS upregulate the Akt pathway, whereas BMP-2 downregulates it. The results of our previous studies have shown that BMP-9 and BMP-2 activate Smad1/5/8 and MAPK (ERK, P38, and JNK) pathways similarly, in a dose-dependent manner, in the absence of FBS. 5 However, BMP-9 and BMP-2 seem to influence the Akt pathway differently, with or without FBS.
Effect of BMP-9 or BMP-2 plus IGF-2 on Akt phosphorylation
Since IGF-2 mimics the synergistic effect of FBS on BMP-9-mediated osteoblast differentiation, we evaluated the effect of IGF-2 combined with BMP-9 or BMP-2 on the activation of Akt at both phosphorylation sites by western blots (Fig. 3A, B) and their densitometric analysis (Fig. 3C, D). Cells were stimulated for 1 h with equimolar concentration (1 nM) of BMP-9 or BMP-2 plus IGF-2 (100 ng/mL) or FBS (10% v/v). We used this concentration of BMP because the dose–response experiments showed that Akt is activated similarly at both phosphorylation sites at this concentration regardless of the experimental conditions.

Effect of FBS and IGF-2 on Akt phosphorylation with or without BMP. Representative western blots showing the phosphorylation of Akt at
IGF-2 alone mostly activated Akt by phosphorylating both Ser473 and Thr308 (Fig. 3A, B). Alvino et al. obtained similar results using mouse fibroblasts (3T3 cells) where IGF-2 increased the phosphorylation of Akt on Ser473. 28 We found that pAkt-Ser473 was intense in cells treated with FBS, while FBS drastically reduced the phosphorylation of Akt on Thr308 with or without BMP-9 or BMP-2 (Fig. 3C, D). Cells incubated with BMP-2 plus IGF-2 contained significantly less pAkt-Ser473 than BMP-2 plus FBS (p < 0.05) (Fig. 3C). BMP-9 and BMP-2 with FBS gave pAkt-Thr308 bands with similar intensities. However, the presence of IGF-2 with or without BMPs induced a strong phosphorylation of Akt at Thr308 compared to FBS (p < 0.05). Thus, the effects of IGF-2 on cells stimulated with BMP-9 and BMP-2 are different. FBS increased Akt phosphorylation only on Ser473 in these cells, whereas IGF-2 increased phosphorylation on both sites. Clearly, FBS and IGF-2 influence the Akt signaling pathway differently in cells stimulated with BMP-9 and BMP-2.
Effectiveness of specific Akt pathway inhibitors and their effects on cell survival
To further determine the effect of each Akt phosphorylation site on BMP-mediated osteoblast differentiation, we then used specific Akt pathway inhibitors. We first assessed the effects of global Akt inhibitors (MK-2206) and then the actions of specific inhibitors that target PDK1 (BX-795) or mTORC (rapamycin and KU-0063794). We monitored cell survival directly by counting labelled nucleus and indirectly with an enzymatic assay (MTS survival assay) (Fig. 4). Incubation for 72 h with MK-2206 at 1 and 2.5 μM did not affect the metabolic activity (Fig. 4B) and the number of attached cells (Fig. 4A, F). By contrast at higher concentrations (5, 10, and 25 μM), MK-2206 decreased the metabolic activity of MC3T3-E1 cells in a dose-dependent manner (2.5 μM vs. 5 μM, p < 0.05; 5 μM vs. 10 μM, p < 0.05 [Fig. 4B]). The number of attached cells is also drastically decreased (2.5 μM vs. 10 μM and 25 μM, p < 0.05 [Fig. 4A, F]). Bao et al. obtained similar results after incubating MC3T3-E1 preosteoblasts with MK-2206 (1–10 μM). 25 Another study showed that inhibiting the PI3K in primary osteoblasts derived from newborn rat calvaria with LY294002 prevented the translocation of Akt to their nucleus, which then increased apoptosis time dependently. 29

Dose–response curves and cytotoxic effect of Akt pathway inhibitors (MK-2206, BX-795, KU-0063794, and rapamycin).
The specific PDK1 inhibitor, BX-795, at concentration higher than 5 μM significantly decreased the metabolic activity (2.5 μM vs. 10 μM, p < 0.01 [Fig. 4C]) and the cell number (2.5 μM vs. 10 μM, p < 0.05 [Fig. 4A, G]). In contrast, specific inhibitors of pAkt-Ser473 (rapamycin and KU-0063794) had little effect on cell survival. Incubation for 72 h with KU-0063794 (0–25 μM) or rapamycin (0–100 nM) produced no significant decrease in the cell viability. This, again, was confirmed using the MTS assay (Fig. 4D, E) and direct cell counting (Fig. 4H, I). These results are consistent with published data. 25 A survey of the actions of 44 kinase inhibitors on MC3T3-E1 preosteoblasts showed that inhibitors targeting mTOR (Ser473) mostly inhibited cell proliferation without significantly affecting cell viability. 25
We verified the effectiveness of these Akt inhibitors before studying how the two phosphorylation sites influenced the differentiation of preosteoblasts toward the osteoblast phenotype. We first evaluated how they inhibited their specific phosphorylation sites in cells incubated for 24 h with equimolar concentrations (1 nM) of BMP-9 or BMP-2 with or without FBS (Fig. 5A, B). Western blots showed that 2.5 μM MK-2206 inhibited pAkt at both phosphorylation sites in cells incubated with BMP-9 or BMP-2 either with or without FBS. Similar results were obtained when L6 skeletal muscle cells were stimulated with insulin. The inhibitor MK-2206 (10 μM) prevented Akt phosphorylation at both Ser473 and Th308. 30 BX-795 (2.5 μM) also prevented the phosphorylation of Akt-Thr308 in cells stimulated with BMP-9 or BMP-2 with and without FBS, all without affecting Akt phosphorylation at Ser473. Feldman et al. and Dangelmaier et al. used PC-3 and human platelet cells to test a range of BX-795 concentrations (0–10 μM) and obtained similar results.31,32 KU-0063794 (2.5–5 μM) and rapamycin (10–50 nM) also dose dependently affected the phosphorylation of Akt-Ser473, without any effect on pAkt-Thr308. These results also agree with published reports. Singha et al. showed that 0.01 nM rapamycin blocked the activation of p70S6K, which is activated by mTOR, and that inhibition with 10 nM rapamycin lasted for >72 h. 24

Dose–response effect of Akt pathway inhibitors on Akt phosphorylation induced by BMP-9 and BMP-2 with or without FBS. Representative western blots of two independent experiments showing the effects of inhibitors of Akt inhibitor (MK-2206, 0–5 μM), PDK1 (BX-795, 0–5 μM), mTORC2 (KU-0063794, 0–5 μM), and rapamycin (0–50 nM) on the phosphorylation of Thr308 and Ser473 of Akt in MC3T3-E1 preosteoblasts stimulated by incubation for 1 h with
Thus the Thr308 phosphorylation site seems to be important for cell survival, while Ser473 does not. Recent studies have also shown that depleting the PDK1 in DT40 cells severely impairs their survival. 33 We find the same differences between the two Akt sites in MC3T3-E1 preosteoblasts incubated with FBS (data not shown). Our western blot studies also showed that 2.5 μM MK-2206 and BX-795 inhibited only their respective phosphorylation sites without having any great effect on cell survival. The selected concentrations used for experiments on cell differentiation were therefore 2.5 μM for MK-2206 and BX-795, 5 μM for KU-0063794, and 100 nM for rapamycin.
Effect of Akt-specific inhibitors on ALP activity
We evaluated the effect of Akt-specific inhibitors on the activity of ALP in cells incubated for 72 h with BMP-9 or BMP-2 (1 nM) with or without FBS (10% v/v) or IGF-2 (100 ng/mL) since these inhibitors blocked the phosphorylation of Akt at Thr308 and Ser473 (Fig. 6A, B). We have shown previously that ALP activity is a good indicator of the BMP-9- and BMP-2-induced osteoblast differentiation of MC3T3-E1 preosteoblasts; its activity varies with cell signaling and the expression of late osteogenic genes such as that encoding osteocalcin. 5

Effect of Akt pathway inhibitors on ALP activity induced by BMP-9 and BMP-2 with or without FBS or IGF-2. ALP activities and cell densities ± SD (two to three independent experiments performed at least twice in duplicate) showing the effects of inhibitors of Akt1/2 (MK-2206, 2.5 μM), PDK1 (BX-795, 2.5 μM), and mTORC2 (rapamycin, 100 nM and Ku-0063794, 5 μM) on MC3T3-E1 preosteoblasts stimulated by incubation for 72 h with BMP-9
The BMP-9-induced ALP activity is enhanced by both FBS and IGF-2, but these factors have only a small influence on the ALP activity in BMP-2-stimulated cells. 5 The Akt inhibitor MK-2206 (2.5 μM), which inhibits Akt at both sites, completely inhibited the ALP activity in cells under all the experimental conditions tested involving BMP-9 or BMP-2 (Fig. 6A, B). BMP-2-induced osteoblast differentiation is known to require activation of the PI3K/Akt pathway. Both Mukherjee and Rotwein, using murine mesenchymal stromal cells (C3H10T1/2), and Fujita et al., using MC3T3-E1 preosteoblasts, have shown that inhibiting PI3K with LY294002 prevents Runx2 synthesis and severely impairs ALP activity and matrix mineralization.19,34 Activation of the PI3K/Akt pathway seems necessary for the synthesis of Runx2 and its binding to DNA. 34 Chen et al. 14 have also treated the murine mesenchymal stromal cells (C3H10T1/2) with LY294002 to show that the PI3K is required for the BMP-9-mediated osteoblast differentiation in terms of ALP activity, osteocalcin, and osteopontin expression, as well as mineralization. Mandal et al. found that inhibiting PI3K prevents the expression of the transcription factor osterix in the mouse myoblast cell line C2C12. It also inhibits the expression of Smad5. 35 Smad1/5/8 are usually activated in MC3T3-E1 preosteoblasts following stimulation with BMP-9 or BMP-2.4,36 Choi et al. also showed that Smurf2 (a ubiquitin-ligase that regulates Smad protein degradation 37 ) is downregulated by activation of Akt in BMP-2-stimulated HEK293 cells. 38 PI3K/Akt seems therefore involved in the regulation of the canonical BMP pathway.
However, the role of pAkt-Thr308 in MC3T3-E1 osteoblast differentiation appears to depend on the type of BMP used to stimulate them. Inhibiting the pAkt-Thr308 in MC3T3-E1 preosteoblasts stimulated with BMP-2 with or without IGF-2 did not significantly influence ALP activity. In contrast, inhibiting pAkt-Thr308 in BMP-9-stimulated cells (with or without IGF-2) completely shut down ALP activity. The pAkt-Thr308 inhibitor had no action on BMP-9-induced ALP activity in cells incubated with serum, while it doubled the ALP activity in BMP-2-induced cells, so that it reached values similar to those produced in response to BMP-9. Inhibiting pAkt-Thr308 also increased (fourfold) the ALP activity in cells stimulated with FBS alone. In addition, Thr308 appears to play an important role on cell survival and proliferation as the cell density for MC3T3-E1 cells stimulated with BMP-9 or BMP-2 ± IGF-2 or FBS in combination with PDK1 inhibitor was lower than the control without the inhibitor. These results indicate that activating Akt at Thr308 is essential for the BMP-9-induced differentiation of MC3T3-E1 preosteoblasts, with or without IGF-2. The results gained with PDK1 inhibitor in cells stimulated by BMP-9 plus FBS might be explained by the fact that other components in FBS can bypass the lack of Akt phosphorylation at Thr308. In contrast, activating Akt at Thr308 does not seem to be essential for BMP-2-induced ALP activity with or without IGF-2. FBS may have a greater effect on BMP-2-stimulated cells if the pAkt-Thr308 is decreased. Thus preventing the phosphorylation of Akt at Thr308 in this particular case mimics the effect of FBS on the cell response to BMP-9. This highlights the fact that Akt activity must be modulated for BMP-induced differentiation and that this also dramatically increases BMP effect in some cases.
Finally, our results also suggest that pAkt-Ser473 plays an important role in osteoblast differentiation (Fig. 6A, B). Both rapamycin and KU-0063794 inhibited ALP activity under all the experimental conditions involving cells incubated with BMP-9 or BMP-2. However, rapamycin (100 nM) did not completely inhibit the differentiation driven by either BMP-9 or BMP-2, unlike KU-0063794 (5 μM). These results correlate well with published data, since mTOR downstream signaling through S6K1 is essential for the differentiation of mesenchymal stem cells toward the bone phenotype. 39 For instance, rapamycin inhibits the differentiation of MC3T3-E1 preosteoblasts induced by ascorbic acid and inorganic phosphate by preventing the synthesis of Runx2, osterix, bone sialoprotein, and osteocalcin. 24 Suzuki et al. also demonstrated that the constitutively active Akt (phosphorylated on Ser473) of MC3T3-E1 preosteoblasts greatly increased their ALP activity and matrix mineralization in osteoblast differentiation medium. 40 The effect of constitutively active Akt could also be increased by the presence of TGF-β1 (0.1 ng/mL). 40 We also found that activating Akt by phosphorylating Ser473 also seems to be very important for cells incubated with IGF-2, since preventing this activation with KU-0063794 completely blocks ALP activity plus BMP-9. Shoba and Lee studied primary cultures of osteoblasts from rat calvaria that had been stimulated with BMP-7 (200 ng/mL) plus IGF-1 (25 ng/mL). 41 They inhibited PI3K with LY294002 and Akt with 109 nM rapamycin and obtained similar results between both inhibitors. 41
Our results using rapamycin and KU-0063794 confirmed that inhibiting pAkt-Ser473 decreased the proliferation. Garcia-Martinez et al. reported that KU-0063794 suppresses the growth of mouse embryonic fibroblasts by stopping the cell cycle in the G1 phase. 42 Similar results were obtained by Singha et al., who demonstrated that 0.1 nM rapamycin greatly slowed the proliferation of MC3T3-E1 (clone 4) preosteoblasts and decreased the concentrations of cyclins A and D1, which are involved in the transition from the G1 to S phases of the cell cycle. 24 These findings indicate that the phosphorylation of Ser473 is important for the proliferation of MC3T3-E1 preosteoblasts, but does not affect their viability, unlike that of Thr308 as shown by cytotoxicity assays.
Figure 7 summarizes how the two Akt phosphorylation sites are implicated very differently in the osteoblast differentiation induced by BMP-9 (Fig. 7A) and BMP-2 (Fig. 7B). The osteoblast differentiation driven by BMP-9 alone appears to require the phosphorylation of Akt on both sites. In contrast, only pAkt-Ser473 in cells stimulated by BMP-9 plus FBS is required, and blocking the phosphorylation of Thr308 produced no drastic change, probably due to the complex composition of FBS. In addition, the increased phosphorylation on both sites that is induced by IGF-2 in BMP-9-stimulated cells is necessary for their synergistic action. Ser473 seems also to be the only Akt site requiring phosphorylation for the increase in BMP-2-dependent ALP activity. Cells stimulated with BMP-2 in the presence of FBS require the phosphorylation of Akt at Ser473 and the dephosphorylation of Akt at Thr308 to increase the osteoblast differentiation with ALP activity similar to that of BMP-9 plus FBS. In the presence of IGF-2, inhibiting Ser473 had a greater impact on BMP-2-induced ALP activity. Our inhibitor and signaling results therefore indicate that Akt must be phosphorylated at Ser473 for osteoblast differentiation to occur, while the involvement of Thr308 site appears to be BMP-type dependent.
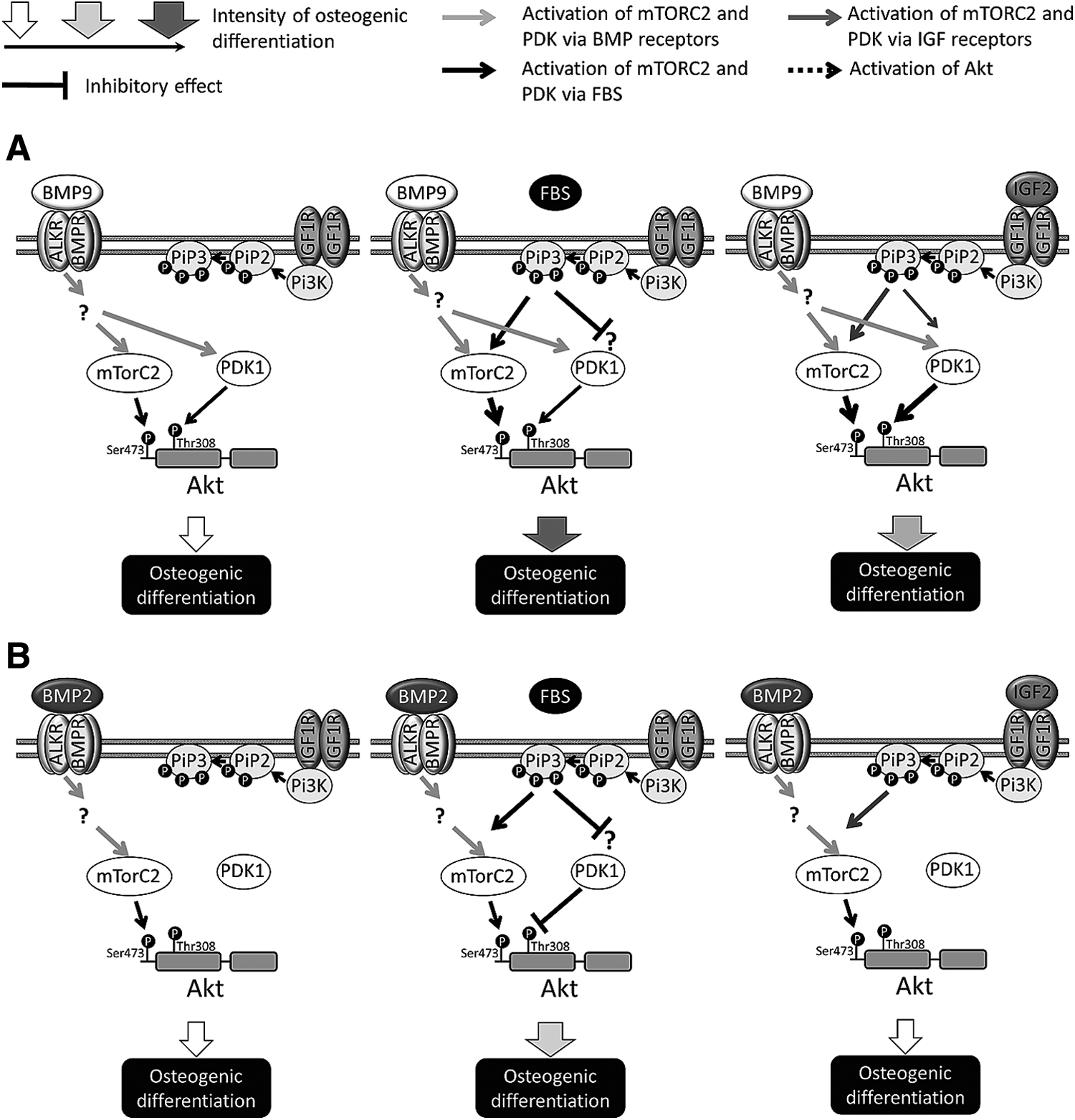
Involvement of Akt phosphorylation sites in osteoblast differentiation induced by BMP-9 and BMP-2 with or without FBS or IGF-2. Diagram showing how the Akt phosphorylation sites (Ser473 and Thr308) are implicated in the osteoblast differentiation induced by
Conclusion
The Ser473 site located in the hydrophobic motif appears to be essential for osteoblast differentiation for cells stimulated with BMP-9 or BMP-2. However, stimulation with BMP-9 and BMP-2 involves the catalytic site Thr308 differently. BMP-9-mediated osteoblast differentiation requires its activation, while BMP-2-mediated differentiation does not. We have also found an explanation for the synergistic effect of IGF-2 on cell response to BMP-9: the phosphorylation of Akt on both activation sites induced by IGF-2 favors the action of BMP-9, but not that of BMP-2. These results provide a clearer picture of the way signaling pathways are implicated in BMP-induced osteoblast differentiation. The patterns of Akt phosphorylation during osteoblast differentiation seem to be more complex than previously believed. Knowing these activation patterns and the signaling pathways that are essential will facilitate more accurate selection of growth factors for developing the defined, potent osteogenic media needed for tissue engineering. However, we have still to understand how these signaling pathways are interrelated and so obtain a clearer picture of the dynamics of this differentiation.
Authors’ Contributions
M.-A.L. took part in designing the study and carried out the cell viability, ALP activity assays, and the densitometric analyses. M.-A.L. also contributed to the western blots analyses, performed the statistical analyses, and wrote the article under the supervision of N.F. A.D. carried out most of the western blot analyses and helped design the study. O.D. helped with the western blot analysis and writing of the article. N.F. supervised the work, took part in the study design, coordinated the work, and helped write the draft article. All the authors have read and approved the final article.
Footnotes
Acknowledgments
The authors thank Dr. Owen Parkes for editing the English text. This research was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) program (grant number 298359). M.-A.L. was supported by an Alexander Graham Bell doctoral scholarship (NSERC).
Disclosure Statement
No competing financial interests exist.