
Abstract
A three-dimensional in vitro Matrigel plug was used as a model to explore delivery patterns of platelet-derived growth factor (PDGF) and bone morphogenetic protein-2 (BMP-2) to a coculture of human mesenchymal and endothelial cells. While BMP-2 is well recognized for its role in promoting fracture healing through proliferation and differentiation of osteoclast precursors, it is not a growth factor known to promote the process of angiogenesis, which is also critical for complete bone tissue repair. PDGF, in contrast, is a known regulator of angiogenesis, and also a powerful chemoattractant for osteoblast precursor cells. It has been suggested that presentation of PDGF followed by BMP may better promote vascularized bone tissue formation. Yet, it is unclear as to how cells would respond to various durations of delivery of each growth factor as well as to various amounts of overlap in presentation in terms of angiogenesis. Using a three-dimensional in vitro Matrigel plug model, we observed how various presentation schedules of PDGF and BMP-2 influenced tubule formation by human mesenchymal stem cells and human umbilical vascular endothelial cells. We observed that sequential presentation of PDGF to BMP-2 led to increased tubule formation over simultaneous delivery of these growth factors. Importantly, a 2–4 day overlap in the sequential presentation of PDGF and BMP-2 increased tubule formation as compared with groups with zero or complete growth factor overlap, suggesting that a moderate amount of angiogenic and osteogenic growth factor overlap may be beneficial for processes associated with angiogenesis.
Introduction
A
While the importance of angiogenesis in bone repair is apparent, the only current FDA-approved growth factor for addressing poor bone healing is bone morphogenetic protein-2 (BMP-2), an osteogenic growth factor.6–9 BMP-2 is widely recognized for its role in upregulating osteogenesis through the proliferation and differentiation of osteoblasts, 10 but less is known about the roles BMP-2 may have during angiogenesis. There is some evidence that BMP may enhance angiogenesis; for example, it has been shown that BMP-2 stimulates the secretion of vascular endothelial growth factor-A (VEGF-A) from osteoblasts. 11 However, it was also demonstrated that BMP-2 delivery alone has limited capacity to upregulate vascular formation at the fracture site. 12 Furthermore, BMPs have even been shown to inhibit angiogenesis in some situations, 13 and antagonisms are known to exist between angiogenic growth factors, such as platelet-derived growth factor (PDGF), and members of transforming growth factor-β (TGF-β) superfamily (including BMP-2). 14 This lack of understanding of the interplay between BMP-2 and angiogenic growth factors may explain the inconsistent results of those studying combined delivery of angiogenic growth factors (such as VEGF, PDGF, IGF, etc.) with BMP-2.15,16
These inconsistencies likely arise because bone fracture healing is widely understood to be a complex, multistage process of restorative steps, not simply mineralization.17–19 Specifically, during successful bone healing, revascularization of the wound space first occurs (angiogenesis), followed by either cartilaginous callus formation or primary bone healing (osteogenesis).17,20 While multiple growth factors may be appropriate to enhance multiple stages of healing, the potential for antagonistic effects may necessitate a distinct growth factor sequence. Accordingly, sequential presentation has been shown in previous studies to dramatically enhance proper tissue formation.21,22 One such study suggests that to properly stimulate ordered biological events, the early presence of a growth factor that upregulates initial activities may be just as important as its later absence. Specifically, it was determined that sphingosine-1-phosphate (S1P) and VEGF have distinct roles, and will only enhance vessel formation when presented in a biologically relevant sequence. 22 Notably, the role, if any, of additional parameters of growth factor presentation, including length of delivery and degree of overlap, has yet to be fully explored for angiogenesis during bone tissue regeneration.
The resulting effects of order and spacing of multiple growth factor timing and overlap may help guide the development of biomimetic therapeutics, including controlled release systems for temporal or spatiotemporal separation of growth factors.23–25 To gain information regarding the effects of variation in growth factor presentation for upregulating angiogenesis in bone repair, we developed a new in vitro Matrigel plug model assay to assess tubule formation in response to various growth factor delivery schemes. A coculture of human umbilical vascular endothelial cells (HUVECs) and human mesenchymal stem cells (hMSCs) allows for the exploration of both angiogenic (organization of HUVECs) and osteogenic (differentiation of hMSCs toward an osteogenic phenotype) responses to growth factor treatment groups. Additionally, hMSCs have the potential to differentiate into mural cells that support the maturation of newly formed tubules. 26 Using the HUVEC/hMSC coculture in this Matrigel plug assay, we are able to assess cellular responses to length, sequence, and degree of overlap in growth factor delivery. We have used this model to investigate our hypothesis that the sequence and degree of overlap in PDGF and BMP-2 delivery will impact tubule formation in the context of bone repair, as previous studies have shown the importance of growth factor sequence regarding angiogenesis alone.21,22
Materials and Methods
Tubule development assay
Well inserts of a 24-well transwell plate (Sigma-Aldrich) were filled with 250 μL of growth factor-reduced, phenol-free Matrigel (LDEV free; Fisher Scientific). Human umbilical vascular endothelial cells (HUVECs; Lonza) and human bone marrow-derived mesenchymal stem cells (hMSCs; Life Technologies) were seeded at a density of 1.5 × 105 cells per well, in a 1:1 (hMSC:HUVEC) ratio. The lower reservoirs of MSC-HUVEC cocultures we filled with 500 μL of a 1:1 ratio of endothelial basal medium and minimum essential medium (Life Technologies) supplemented with 1% penicillin–streptomycin and 1% antibiotic–antimycotic to provide hydration to Matrigel plugs.
Growth factor delivery to cell cultures
Growth factor-enriched medium was prepared by combining endothelial basal medium and minimum essential medium in a 1:1 ratio, and supplementing it with 1% penicillin–streptomycin and 1% antibiotic–antimycotic. For PDGF treatment groups, soluble PDGF-BB (Fisher Scientific) was added to the medium for a final concentration of 10 ng/mL PDGF. For BMP treatment groups, soluble BMP-2 (Fisher Scientific) was added to the medium for a final concentration of 100 ng/mL BMP, and for PDGF+BMP treatment groups, PDGF and BMP were added to the medium for a final concentration of 10 and 100 ng/mL, respectively. Growth factor concentrations were chosen based on dose-dependency studies found in literature references.27,28 Based on these studies, the lowest dose for which a cellular response was observed was chosen for use in this study. Prepared medium was delivered to corresponding groups by injecting 20 μL aliquots per well per day for the duration of the experiment. PDGF and BMP were each delivered for a minimum of 1 day, to a maximum of 10 days, with between 0 and 10 days of overlap, depending on the conditions studied.
Immunofluorescence
At the conclusion of the experimental timeframe (10 days), Matrigel plugs were removed from transwell inserts, embedded in Tissue-Tek (Fisher Scientific) and snap frozen in liquid nitrogen. Frozen sections (10 μm) were incubated with primary antibody mouse anti-human CD31/PECAM-1 (R&D Systems) and secondary antibody donkey anti-mouse IgG (R&D Systems) to identify endothelial cell activity and vessel formation. Images of CD31-stained sections were taken at 20× and 10× using a Nikon inverted light microscope, then analyzed using WimTube software (Wimasis Image Analysis, GmbH). For analysis, 10× images were used to gain a larger representative sample area. Wimasis software was used for analysis of four parameters for every image: total area covered by vessels, total length of vessels, number of branching points, and number of closed loops. These parameters were measured for each representative area imaged at 10× magnification, and are illustrated in more detail in Supplementary Figure S1; Supplementary Data are available online at www.liebertpub.com/tea.
To identify MSC differentiation toward a pericyte phenotype when exploring length of PDGF delivery, slides were incubated with primary antibody rabbit anti-alpha smooth muscle actin (ab5694; Abcam) and secondary antibody goat anti-rabbit IgG H&L (Alexa Fluor 488; Abcam). Images of slides were taken at 20× using a Nikon inverted light microscope. Alpha-smooth muscle actin (α-SMA) staining was quantified by analyzing fluorescent intensity of the resulting using ImageJ software. Image files were converted to 8-bit formatting, then automatically thresholded using the triangle thresholding method in ImageJ. Resulting binary images were quantified with the ImageJ “analyze particles” method with default settings.
Statistical analysis
Multivariate analysis of variance was performed using GraphPad 5.0 software to identify statistically significant differences (p < 0.05) across multiple experimental groups for each set of in vitro growth factor delivery data. Post hoc multiple comparison testing was performed to compare mean using Tukey's test. Each experimental growth factor delivery group (PDGF, BMP, PDGF+BMP, etc.) contained N = 3 samples. For every N, measured outcomes of n = 3 cryoslices were first averaged to better represent each sample.
Results
Formation of tubules in response to sequential treatment schedules
To explore the variable space related to the sequence of PDGF/BMP for induction of vascular structures, we explored the growth factor delivery of (1) PDGF alone, (2) BMP alone, (3) PDGF+BMP [10 days of PDGF and BMP together], (4) PDGF to BMP [5 days of PDGF alone, followed by 5 days of BMP alone], (5) BMP to PDGF [5 days of BMP alone followed by 5 days of PDGF alone], (6) “blank” medium with no added growth factors (Fig. 1). The “blank” experimental group, which was supplemented with neither PDGF nor BMP, served as a negative control. All growth factor delivery schedules were assayed using the HUVEC/hMSC cocultures seeded in Matrigel plugs. As a baseline, CD31 staining of Matrigel sections revealed at least some degree of tubule formation in each experimental group (Fig. 2A, B). The greatest amount of CD31 staining, as quantified by tubule area coverage and tubule length, was observed in samples treated with PDGF alone (Fig. 3). However, CD31 staining also showed that the sequential delivery of PDGF to BMP outperformed other treatment groups in terms of tubule density (quantified by area coverage and tubule length per frame) and tubule organization (quantified by number of branching points.)
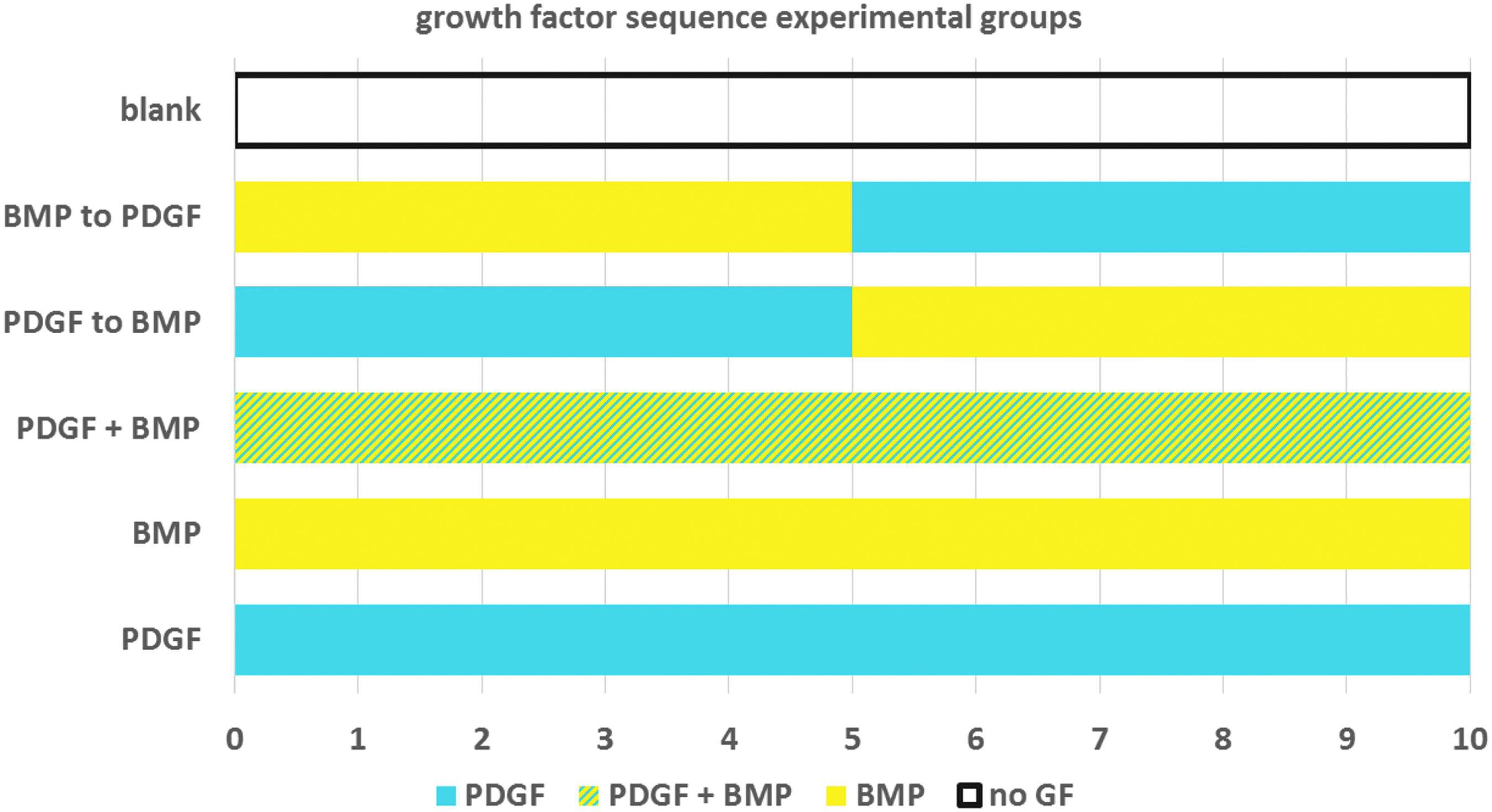
Sequential growth factor delivery regimens showing type of growth factor delivery per day over 10 days. Growth factors were delivered by pipetting 20 μL of medium with concentrations of 10 ng/mL PDGF and/or 100 ng/mL BMP. BMP, bone morphogenetic protein; PDGF, platelet-derived growth factor.
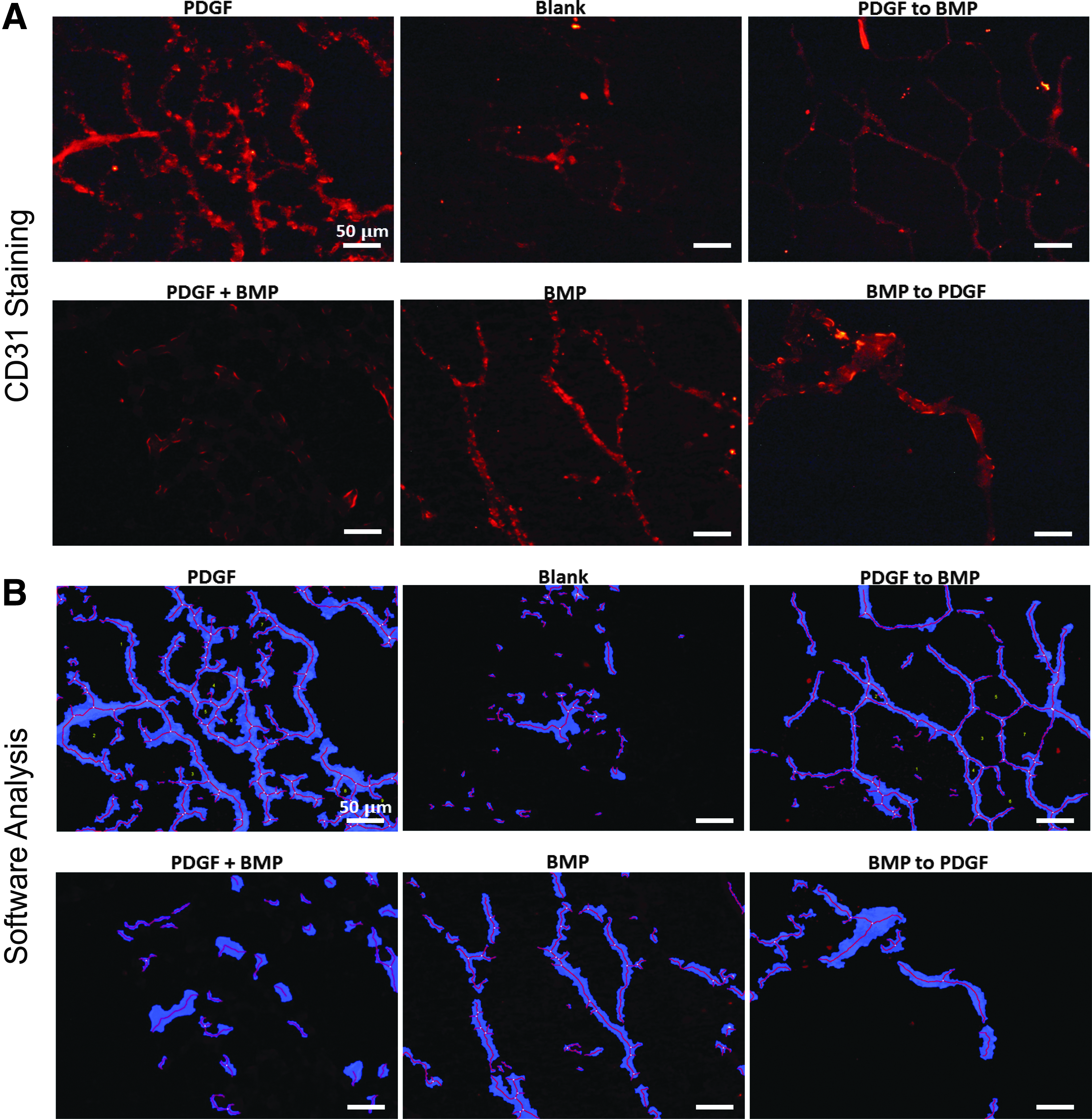
Sequential delivery of PDGF to BMP and delivery of PDGF alone results in organized tubule formation in vitro.
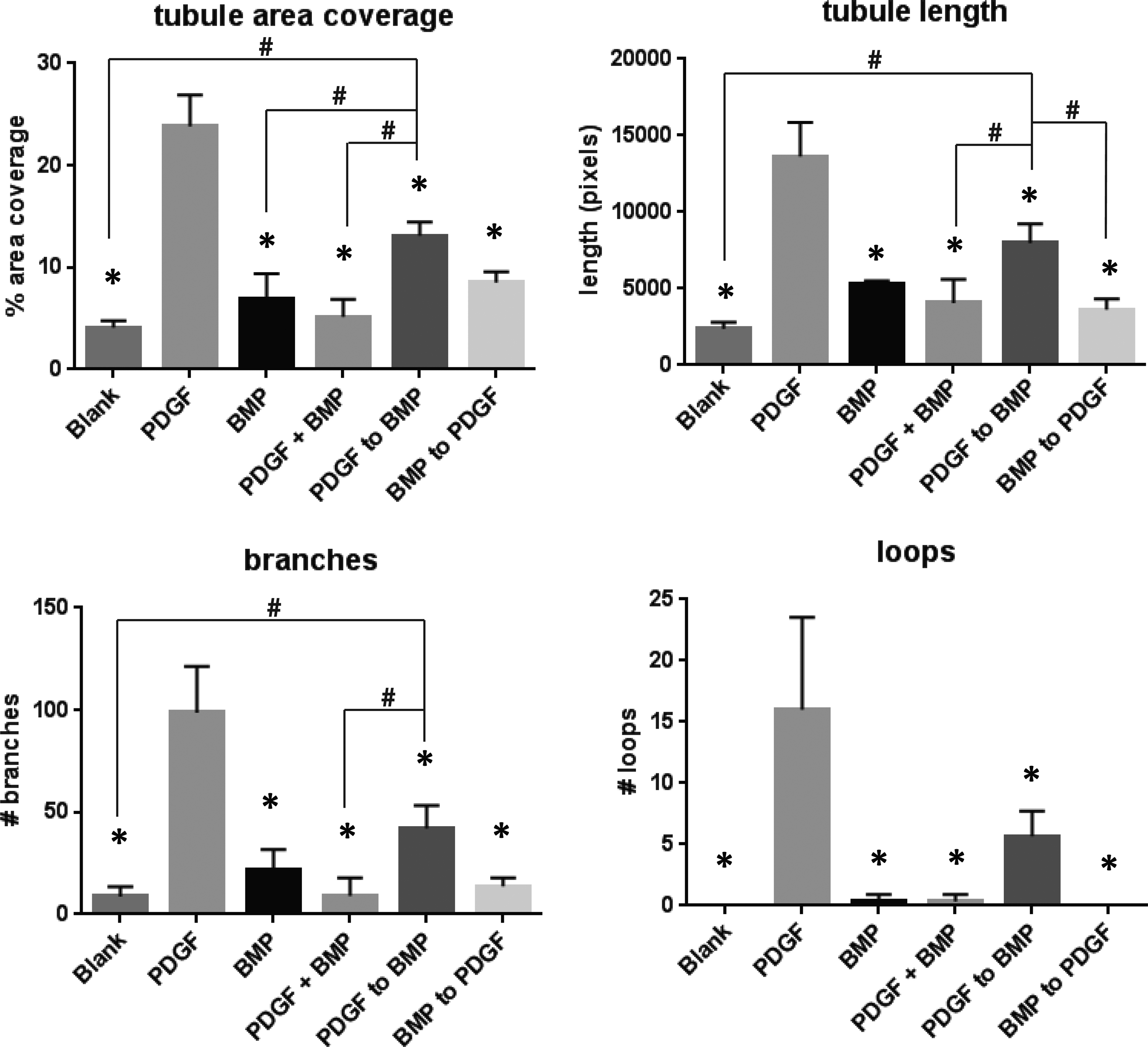
Quantification of tubule area, tubule length, branches, and loops for sequential growth factor delivery groups showing that PDGF delivery alone (positive control) results in significantly greater tubule formation when compared with all other treatment groups at each parameter. Sequential PDGF to BMP delivery results in significantly greater tubule area coverage, tubule length, and branching points when compared with several other treatment groups. #Indicates significant difference from PDGF to BMP treatment group, *indicates significant difference from PDGF treatment group. ANOVA followed by post hoc multiple comparisons testing using Tukey's test. α = 0.05. ANOVA, analysis of variance.
Formation of tubules in response to length of PDGF delivery
Three-dimensional Matrigel plugs in transwell plate inserts were seeded with a 1:1 coculture of hMSCs and HUVECs to evaluate tubule formation and α-SMA expression in response to various lengths of PDGF delivery when sequentially delivering PDGF and BMP-2. Within the 10-day experimental timeframe, we explored five variations in length of PDGF to BMP delivery, with the sequential switch occurring after 1, 3, 5, 7, or 9 days of PDGF delivery (Fig. 4). Matrigel sections treated with longer PDGF delivery before the switch to BMP-2 exhibited greater amounts of tubule formation (CD31 staining) and α-SMA expression (Fig. 4). As shown in Figure 4, when PDGF was delivered for 7 days before the switch to BMP-2, sections of Matrigel plugs displayed significantly more α-SMA expression as compared with 1, 3, and 5 days of PDGF delivery. When PDGF was delivered for 9 days before the switch to BMP-2, significantly more α-SMA staining was observed as compared with 1 and 3 days of PDGF delivery. Greater amounts of CD31 staining were also observed for 9 days of PDGF delivery as compared with 1, 3, and 5 days. Although tubule lumens were not specifically assayed for in this study, they can be longitudinally observed in “7 days PDGF/3 days BMP” and “9 days PDGF/1 day BMP” frames of Figure 4, and are similar in appearance to longitudinally oriented tubule lumens in similar 3D in vitro studies.29,30 Individual color channels for CD31, α-SMA and DAPI staining are found in Supplementary Figure S2.

Immunofluorescent staining of CD31 (red), α-SMA (green), and nuclei (blue) of Matrigel cross sections suggests that longer PDGF delivery before switch to BMP (7 days, 9 days) results in greater amounts of tubule formation and presence of pericytes * indicates significantly more area coverage versus days 1, 3, and 5. #Indicates significantly more area coverage versus days 1, 3 when compared using ANOVA and Tukey's test for post hoc comparisons, α = 0.05. All scale bars = 50 μm.
Formation of tubules in response to various amount of overlap in PDGF to BMP sequence
Three-dimensional plugs of Matrigel in transwell plate inserts were seeded with a 1:1 coculture of hMSCs and HUVECs to evaluate tubule formation in response to treatment groups representing various degrees of growth factor presentation overlap in vitro over the 10-day experimental timeframe as shown in Figure 5. CD31 staining of Matrigel sections revealed that the greatest amount of tubule formation occurred in groups with moderate growth factor overlap (2–4 days of overlap) as shown in Figure 6B. In sections representing 4 days of growth factor overlap, a significantly higher amount of tubules (tubule area, length) and greater level of tubule organization (loops, branching points) were observed than in any other experimental group (Fig. 6B). Sections representing 2 days of overlap were observed to have significantly more tubule area coverage and length than sections representing longer periods of growth factor overlap (8 and 10 days). For purposes of comparison, the 0-day overlap group in Figure 6 corresponds to the same treatment schedule as the “PDGF to BMP” group in Figure 2, and the 10-day overlap group corresponds to the same treatment schedule as the “PDGF+BMP” group in Figure 2. Merged color channels for CD31, α-SMA and DAPI staining are found in Supplementary Figure S3.
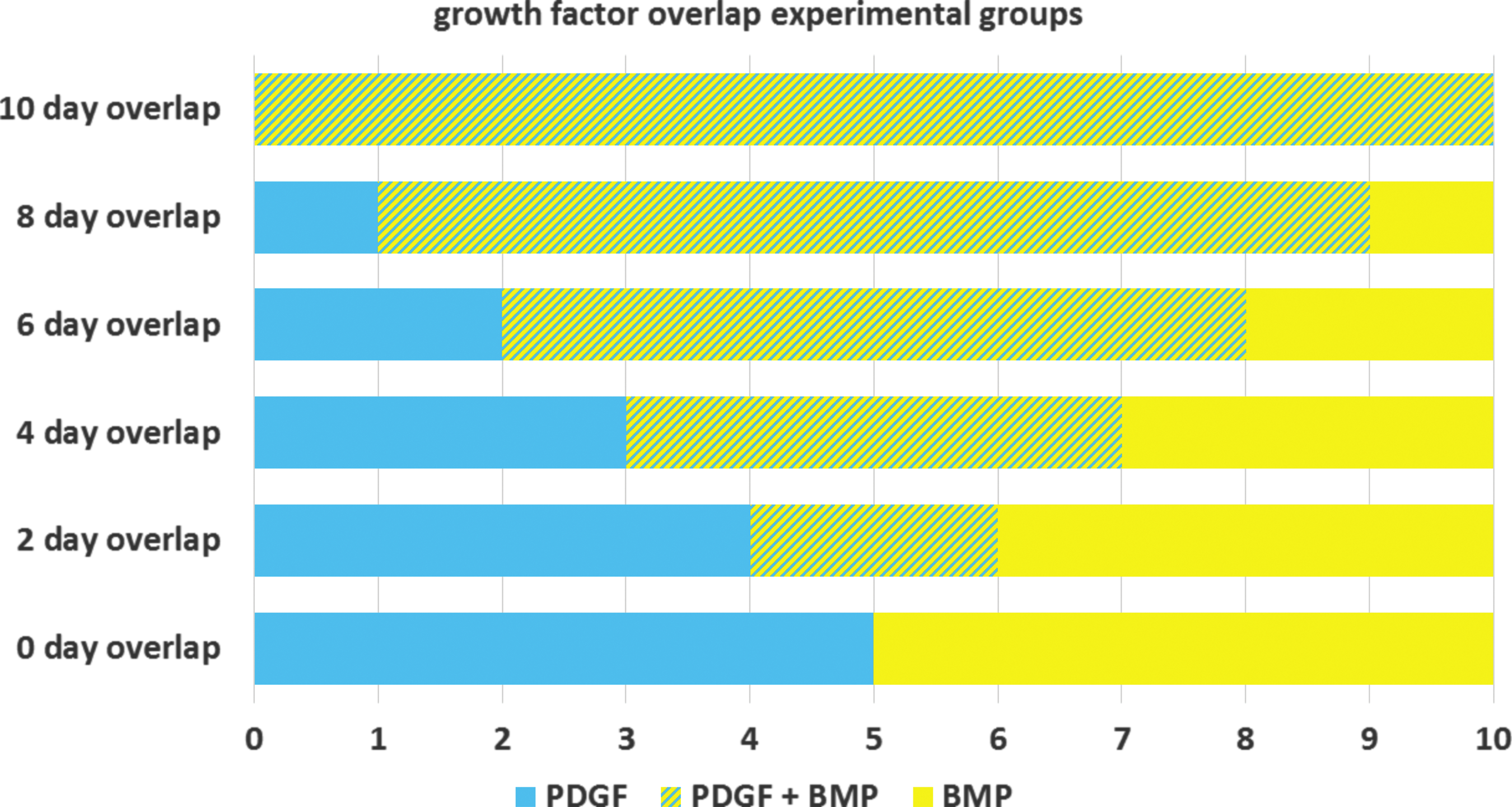
Overlapping growth factor delivery regimens showing type of growth factor delivery per day over 10 days. Growth factors were delivered by pipetting 20 μL of medium with concentrations of 10 ng/mL PDGF and/or 100 ng/mL BMP.

Discussion
Bone regeneration is a complex, stage-wise process, including phases of angiogenesis and osteogenesis. 31 To coordinate these steps, there are many influential factors, including biochemical cues from growth factors, 32 mechanical signaling, 33 and the physiological condition of the patient.34,35 To overcome lack of bone formation in the case of nonunions, supplemental BMP-2 is a well-established and clinically approved method for enhancing fracture healing by upregulating osteoblast proliferation and differentiation.10,36 However, BMP-2 alone has not been shown to be effective in upregulating angiogenesis in the bone regenerating niche, 12 and thus, the early stage of vascular infiltration may benefit from the administration of an angiogenic growth factor. As shown in previous studies, the sequence of growth factor presentation is critical for successful stage-wise tissue formation.21,22
The objective of this study was to determine how the pattern of presentation of angiogenic and osteogenic growth factors affect vascular formation in the context of bone regeneration. While some recent bone regeneration studies design controlled release systems for temporal separation of growth factors,23–25 cellular responses to multiple growth factor timing and overlap are otherwise largely uncharacterized. For the purposes of this study, we utilized a 3D Matrigel plug model to focus on the sequence, timing, and overlap of two growth factors involved in bone regeneration that influence both angiogenesis (PDGF) and osteogenesis (BMP-2) in coculture with endothelial cells and mesenchymal stem cells.
The known, potent effect of PDGF on angiogenesis was apparent in our study, which suggests that PDGF delivery alone (for the entire course of the study) results in the greatest amount of tubule formation (Fig. 2). Consistent with this, when delivered in sequence with BMP-2, longer PDGF delivery results in greater amount of tubule formation, as expected (Fig. 4). PDGF is known to modulate angiogenesis both directly (through endothelial cell PDGFβ receptors) 37 and indirectly (through stimulation of VEGF production from other cell types), 38 mechanisms which have both been observed in vitro.37,39 In addition to the role of PDGF in supporting angiogenesis, there is also evidence suggesting that PDGF is capable of acting as a chemoattractant for vasculature-stabilizing pericytes, or mural cells.40,41 Additionally, it has been shown that MSCs are capable of differentiating into this pericyte phenotype and supporting the maturation of vasculature. 26 In accordance with this information, we observed that when delivering a sequence of PDGF to BMP-2, with increasing length of PDGF delivery, a greater amount of α-SMA+ cells were present (Fig. 4), suggesting that some MSCs present in the cell cultures may be acting phenotypically as pericytes, supporting tubule integrity. 26 This is significant, given that the colocalization of α-SMA+ cells with CD31+ cells indicates mature tubule formation. 42 By coculturing HUVECs and MSCs in this model, we add a layer of complexity over single cell culture, including an additional cell type and providing a platform for cellular crosstalk to occur. The HUVEC/MSC coculture may also be useful in exploring cellular tendency toward bone formation at later time points, as MSCs may serve as osteoblast precursors. 43
Due to the stage-wise nature of bone repair, involving early steps of vascular infiltration and cell recruitment (coordinated by PDGF), followed by osteoblast proliferation and mineralization (upregulated by BMP-2), 31 we hypothesized that PDGF delivery followed by BMP-2 delivery would most effectively promote tubule formation. The results illustrated in Figure 3 demonstrate that the PDGF to BMP sequence did indeed result in greater tubule formation than either the reverse sequence (BMP to PDGF) or simultaneous delivery of PDGF and BMP. These results agree with existing studies that also suggest the importance of order in growth factor delivery for stage-wise regeneration.21,22,44 For example, it has been shown that to promote specific, ordered stages of angiogenic repair, certain growth factors must be delivered in a particular sequence (not in reverse or simultaneously) for optimal results.21,22 Importantly, it appears that an antagonistic effect may exist between PDGF and BMP signaling, as the simultaneous delivery of these growth factors results in significantly less tubule formation when compared with sequential growth factor delivery (Fig. 3). This is not surprising, given evidence that a molecular antagonism (regulation of microRNA-24 expression) has been observed between PDGF and BMP signaling that controls cellular phenotypes, particularly vascular smooth muscle cell phenotypes. 14 Specifically when considering bone repair, these results are consistent with a previous study, which suggests that temporal or spatiotemporal separation of angiogenic and osteogenic growth factors are beneficial for both the vascularization and mineralization stages of healing. 44
While complete overlap in PDGF and BMP-2 delivery resulted in poor tubule formation, we observed that some degree of overlap in the transition from PDGF to BMP-2 delivery may be beneficial for tubule formation (Fig. 6). Greater amounts of tubule formation were observed when there was a 2–4-day overlap in growth factor delivery as compared with a “hard transition” in growth factor delivery, with no overlap in the sequence of PDGF to BMP-2 (Fig. 6). This observation may be explained, in part, by how biological cascades of growth factors are regulated and expressed in situ. In the general wound healing model, there is evidence that growth factors are expressed in a cascading fashion, with overlapping waves of expression that may coincide for a few hours to days.45,46 Likewise, it has been shown that phases of bone healing and remodeling are not discrete stages, but instead, have some amount of overlap in transition.17,47 In addition to these observations, moderate overlap (2–4 days) in PDGF and BMP-2 delivery resulted in greater tubule formation than longer periods of overlap (6–10 days, Fig. 6), which agrees with literature suggesting that the prolonged expression of PDGF may be deleterious for mineralization stages bone formation,48–50 and therefore, may be most effective when restricted to earlier healing stages.
Importantly, a baseline amount of CD31+ staining was observed in each experimental group, including Matrigel constructs with no delivered PDGF or BMP-2. This may be explained by the minor amount of growth factors found in Matrigel, including TGF-β and insulin-like growth factor 1 (IGF-1). 51 However, these low levels of background growth factors are not predominantly responsible for tubule formation, as there were statistically significant differences observed across experimental groups based on the presence or absence of PDGF and BMP-2 (Fig. 2). Additionally, according to the manufacturer's analysis (Corning), concentrations of growth factors in the Matrigel used in these experiments range from 0.1 pg/mL to 5 ng/mL (one to six orders of magnitude less than the quantities of PDGF and BMP-2 used).
Our current investigation of growth factor presentation on tubule formation suggests that PDGF and BMP-2 presentation patterns do indeed play a role in angiogenesis, by affecting the quantity and organization of tubules. Due to the flexibility of our model, further studies to explore the effects of growth factor timing on a variety of cell types are possible. To assess the impact of growth factor delivery schemes on the expression of bone markers and later stages of mineralization, future studies will focus on later time points (14–28 days) when these effects become apparent. Importantly, the evaluation of later time points may provide additional information on how earlier growth factor timing patterns affect downstream healing activity, such as the eventual mineralization of bone. Additional cell types and growth factors can also be explored, along with in vivo validation, as there are many factors involved in fracture healing aside from those explored in this study. While this model provides a controllable in vitro system to evaluate cellular responses, the complexity of the system can be increased to better recapitulate an in vivo environment. Ultimately, in vivo validation studies must be performed in the future to confirm these results.
Conclusion
We have utilized a three-dimensional, in vitro model that permits the exploration of the effects of growth factor timing on vascularization in the context of bone repair with a large degree of variables. Specifically, with this system, we were able to evaluate resulting tubule formation from variations in sequence, length, and degree of overlap in growth factor delivery. We observed that the sequence of angiogenic (PDGF) to osteogenic (BMP-2) presentation resulted in greater tubule formation than the reverse presentation or codelivery of these growth factors. Additionally, moderate overlap (2–4 days) in the sequence of PDGF to BMP-2 delivery resulted in greater tubule formation in comparison to the absence of overlap as well as prolonged overlap. This approach may be further applied to investigate the effects of growth factor timing, overlap, and sequence on additional cell types, later time points to gather information that may lead to improved in vivo outcomes. The resulting information could serve as a valuable contribution to our understanding of the effects of growth factor timing and sequence in the bone regeneration niche, and ideally, advance the development of growth factor therapies for fracture repair.
Footnotes
Acknowledgments
The work of the authors was supported by the National Institutes of Health CATER (Cellular Approaches to Tissue Engineering and Regeneration) training program (T32EB001026) (to E.A.B.), and the Camille and Henry Dreyfus Foundation Teacher Scholar Award (to S.R.L.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.