
Abstract
Recent studies in the field of neuro-tissue engineering have demonstrated the promising effects of aligned contact guidance cue to scaffolds of enhancement and direction of neuronal growth. In vivo, neurons grow and develop neurites in a complex three-dimensional (3D) extracellular matrix (ECM) surrounding. Studies have utilized hydrogel scaffolds derived from ECM molecules to better simulate natural growth. While many efforts have been made to control neuronal growth on 2D surfaces, the development of 3D scaffolds with an elaborate oriented topography to direct neuronal growth still remains a challenge. In this study, we designed a method for growing neurons in an aligned and oriented 3D collagen hydrogel. We aligned collagen fibers by inducing controlled uniaxial strain on gels. To examine the collagen hydrogel as a suitable scaffold for neuronal growth, we evaluated the physical properties of the hydrogel and measured collagen fiber properties. By combining the neuronal culture in 3D collagen hydrogels with strain-induced alignment, we were able to direct neuronal growth in the direction of the aligned collagen matrix. Quantitative evaluation of neurite extension and directionality within aligned gels was performed. The analysis showed neurite growth aligned with collagen matrix orientation, while maintaining the advantageous 3D growth.
Introduction
T
During neuronal pathfinding, growth cones are guided by multiple factors in their vicinity, including topography. 4 Interactions of neurons with 2D topographically patterned substrates have been studied extensively.5,6 We have also shown how surfaces decorated with line-patterned ridges of nanometer height direct neurite growth in a culture system.7,8 Electrospinning, for example, has been used to create fibrous aligned surfaces for directing neuronal growth.9,10 Decorating electrospun scaffolds with nanoparticles further enhanced growth. 11 Since 2D substrates limit neuronal growth to a planar surface, there is a growing need to provide a more complex 3D architecture through which neurons can navigate. Several studies have focused on neuronal growth in 3D constructs, showing a preference toward a 3D environment.12–15 Methods for manipulating neuronal growth in 3D environments include electrospinning,16,17 3D printing, 18 microfabrication of cues,19–21 and self-assembled gels. 22 Hydrogel systems have been employed as 3D materials for TE applications as they possess mechanical and structural properties similar to soft tissues.15,23–25 Hydrogels comprising extracellular matrix (ECM) proteins, such as collagen, have been suggested as preferable platforms for neuronal growth due to their natural origin, biodegradability, weak antigenicity, and good biocompatibility.26,27
When dealing with collagen, a fiber-based alignment is of particular interest since the microscale fibrillary organization of collagen profoundly influences tissue mechanical properties and cell organization.28,29 Furthermore, collagen exhibits an oriented fibrillary organization in nerve tissue and may be important in guiding regeneration.30,31 Alignment of collagen fibers has been shown to direct neuronal growth, including human stem cell-derived neurons.32,33
Several methods for directing and orienting collagen gel structures that can affect cellular organization have been studied, including 3D microfabrication, 34 magnetic flow-induced alignment, 35 and electrochemical fabrication. 36 Strong magnetic fields have also been employed to align collagen gels, affecting neurite growth in vitro and axonal migration within a nerve guide in vivo.37,38 We have previously oriented collagen gels through the integration of magnetic nanoparticles and magnetic fields during the gelation period and showed consequent alignment of neuronal growth. 39 Continuous or cyclic stretching or tethering throughout the cellular growth has also affected the remodeling of collagen fibers,40–42 proven to be beneficial for TE applications43–45 and further harnessed for in vivo TE. 46 Many of these methods often require specific setups or long preparations, raising the need for accessible strategies to align 3D gels for potential future therapeutics. In this study, we used a simple mechanical method to orient collagen gel fibers as a 3D template to direct neuronal regeneration. Before gelation, we mixed whole dorsal root ganglia (DRG) mouse explants, primary single neurons, as well as neuron-like PC12 cells, in the gel. Immediately after the gelation, we oriented the fibers using a mechanical strain method by applying a single short-range stimulation based on the analysis of collagen organization of Vader et al. 47 For the single neuron assays, we used neurons of the medicinal leech (Hirudo medicinalis), which develops low-density neuronal cultures, useful for studying regeneration and morphological analysis. 48 This approach provided a simple and reproducible method for orienting 3D collagen gels. As collagen is a natural component of the nervous system, implants of collagen with organized and oriented nanofibers can support directed growth for effective neuronal regeneration, particularly as conduits for bridging nerve gaps following injuries in vivo.
Materials and Methods
Collagen hydrogel preparation
The method used for preparing the 3D gels was based on that of Blackshaw et al. 49 The following collagen mixture was prepared under sterile conditions and held on ice to prevent initiation of gelation. In a sterile 1.5 mL reaction tube, collagen (type-I rat tail extract, in 0.02 M acetic acid; BD-Biosciences) was diluted by the addition of 10× appropriate media, 10× Leibovitz-15 (L-15) for leech neuronal culture or 10× DMEM for PC12 or DRG culture (Sigma Aldrich Co.) (filter sterilized), and 7.5% sodium carbonate (filter sterilized). Following the addition of cells, typically in 5–10 μL of appropriate media, collagen concentration was brought to 3 mg/mL and physiological pH was indicated by phenol red within the mixture.
The mixture was then mixed carefully by pipettation, taking care not to introduce air bubbles. One hundred microliters of the mixture was transferred and spread upon a 13-mm glass cover slip placed in a 35-mm Petri dish (both UV sterilized). For examining the alignment effect on larger gels, 200 μL of the mixture were spread (2 cm length) within a 35-mm Petri dish. The gel was allowed to solidify and stabilize in a 25°C humidified chamber for 40 min, and then was immersed in 3–4 mL appropriate media. Several collagen gels were subjected to strain-induced alignment following the gelation period.
Gel macrodimensions were measured using confocal microscopy.
Primary neuronal culture
Neurons were isolated from the central nervous system of adult medicinal leeches Hirudo medicinalis as described previously.8,50 Briefly, leeches were anaesthetized in ice for 20 min before dissection. Each animal was then stretched longitudinally and pinned dorsal side up on a Sylgard ice-cooled surface. The full length of the animal dorsal skin was cut and access blood washed with leech ringer solution (115 mM NaCl2, 1.8 mM CaCl2, 4 mM KCl, and 10 mM Tris maleate, pH 7.4). Nerve cord and ganglia were then exposed and dissected with fine scissor tips taking care not to cut nerve roots or connectives. Ganglia were pinned down on a Sylgard-184 Petri dish using 0.002 tungsten cord and immersed in an L-15 complete medium (L-15 supplemented with 6 mg/mL glucose, 0.1 mg/mL gentamicin, 2 mM/mL glutamine, and 2% fetal bovine serum [FBS]). Next, ganglia were treated enzymatically with 2 mg/mL collagenase/dispase enzyme solution (Roche) within the L-15 complete medium for 1 h at room temperature. Finally, ganglia capsules were opened carefully using fine scissors to expose cells. Cells were collected by micropipette to a 1-mL test tube containing the L-15 medium. Following three successive washes by spin-down (1500 rpm), cells were suspended in 20–30 μL L-15 and immediately seeded on either a 2D Concanavalin-A (Con-A) substrate or within 3D collagen gel, as described above. Gels were either subjected to mechanical strain or maintained as control gels. Control 2D cultures were prepared by plating cells in 80 μL L-15 on a 35 Petri dish precoated with Con-A (Sigma-Aldrich Co.; 0.5 mg/mL). Typically, cultures were kept in the dark at 25°C in a humidified chamber for 7 days, and the L-15 complete medium was replaced every 2 days. Neurons out of five to six ganglia were plated per gel, or 2D cell culture (typical yield of 300–400 cells per gel). Results of three separate experiments are shown.
PC12 cell culture
Rat pheochromocytoma PC12 cells (ATCC) were grown in suspension in the RPMI medium supplemented with 10% horse serum (HS), 5% FBS, 1% L-glutamine, 1% penicillin–streptomycin, and 0.2% amphotericin, in a humidified incubator at 37°C containing 5% CO2 (medium and supplements were purchased from Biological Industries). Cells were passaged as previously described.51,52 Fifteen thousand cells were collected and either plated on a 2D 35-mm culture dish coated with collagen at 6 μg/cm2, or within 3D collagen hydrogels, as described above. Gels were then allowed to solidify for 40 min in a 37°C incubator, and then gels were either subjected to mechanical strain to induce alignment or maintained as control gels (n = 2 gels or 2D plates for each group). Two-dimensional samples and gels were immersed in 2 mL reduced serum media (1% HS). To induce PC12 differentiation, murine β-nerve growth factor (NGF; Peprotech) was added to the medium at a final concentration of 50 ng/mL. The medium containing murine β-NGF was replaced every 2 days and kept for 7 days in culture. Results of three separate experiments are shown.
Mouse DRG explant
All animal studies were conducted in accordance with animal care and protection regulations as approved by the Bar-Ilan University ethics committee (24042015 license number). Adult C57b1 mice, 6–9 week old, were euthanized. An incision was made along the back with a scalpel to expose the spinal cord. Connective tissue was removed using small sharp scissors, and then the cleaned spine column was transferred to a Petri dish at room temperature. The spinal cord was removed under a dissecting microscope using fine forceps, and DRG were located. Whole DRG were transferred to a clean Petri dish containing F-12 (Thermo Fisher Scientific) media supplemented with 10% fetal calf serum and 1% penicillin–streptomycin. DRG were immediately transferred using fine tweezers to the collagen gel solution. Following 40 min of gelation at 37°C, gels were immediately stretched, or maintained as control gels without treatment (three gels in each group, two ganglia per gel). Media were replaced daily, with the addition of NGF at a final concentration of 50 ng/mL.
A schematic representation of the experimental setup is shown in Figure 1.

Experimental setup. Cells or whole ganglia were extracted from animals and
Turbidity
Rate of fibrillogenesis was measured using turbidity. The collagen solutions were prepared on ice as described above, and 100-μL aliquots were pipetted into a cooled 96-well plate, in triplicates. The absorbance of the solution was measured at 405 nm every 30 s at temperatures of 25°C in a synergy4 plate reader (BioTek). Three separate experiments were analyzed.
Rheometry
Rheological analysis was performed on a rheometer TA instruments HR-3 using 8 mm parallel-plate geometry. Collagen gels were prepared as described and samples were placed on the rheometer stage. The rheometer plate was lowered and the stage was adjusted so that the gel sample was placed directly under the plate. A solvent trap was employed to reduce effects of dehydration. Settings were held at 25°C and frequency sweeps from 1 to 100 Rad/s were performed using a controlled stress of 0.5 Pa and 5% strain. Storage, loss, and complex moduli were recorded and values found to be within the linear viscoelastic range of the system are presented. Three experiments in triplicates were recorded, results are presented ± standard deviation (STD).
Confocal reflectance microscopy
Polymerized 3D collagen gels were imaged using a Leica TCS SP5 confocal microscope by a 63, 0.9 NA water immersion lens. Samples were illuminated with 488 nm Argon laser light and the Meta channel of the microscope was set to detect wavelengths between 474 and 494 nm to allow reflectance mode.
High resolution scanning electron microscopy
Seven days after seeding, neurons growing in collagen gels were fixed using 2.5% paraformaldehyde/2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer for 1 h at room temperature. After fixation, cultures were repeatedly rinsed with phosphate-buffered saline (PBS; without Ca+2 or Mg+2, pH 7.4) and then treated with a guanidine HCl/tannic acid (4:5) solution (2%) for 1 h at room temperature. Cultures were repeatedly rinsed with PBS and then dehydrated in a graded series of 50%, 70%, 80%, 90%, and 100% ethanol/water (v/v) for 10 min each. The residual ethanol was then removed using a series of 50%, 75%, and 100% ( × 3) Freon solutions in ethanol, for 10 min each. Finally, the samples were left for a few seconds to air dry. Samples were either of neurons embedded in thin collagen films (∼0.1 mm height), to allow for top view of the cells within the collagen matrix, or alternatively, to view the cells inside the scaffolds, the samples were broken into pieces using fine tips. The dried samples were mounted on aluminum stubs, sputter coated with carbon, and viewed with the scanning electron microscope (SEM; Model FEI Quanta250 FEG).
Electrophysiological recording
Microelectrodes for intracellular recordings were made with borosilicate glass of 1 mm exterior diameter and 0.75 mm internal diameter pulled in a P97 puller (Sutter Instruments) to create a tip diameter of 0.7–0.9 μm. The microelectrodes had resistances of 18–23 MΩ when filled with 3 M potassium acetate. We used a standard single-electrode current-clamp intracellular recording technique to monitor the spike activity. Neurons were embedded in thin collagen gels and maintained in culture for 7 days. Using thin collagen films compensated for electrode tendency to stick and detach collagen gel. Signals were amplified (Molecular devices multi clamp 700B), filtered, and digitized by an analog-to-digital board Digidata 1400A (Axon Instruments).
Live/dead assay
To examine neuronal survival in collagen gels, live/dead cells were labeled after 7 days in culture (Live/Dead imaging kit R37601; Life Technologies). Following 15 min of incubation with the live/dead stain, the gels were imaged using confocal microscopy. Cells were manually counted and the death rate was quantified. The average of three separate experiments is shown.
Immunocytochemistry
The L-15 medium was removed from leech primary neuronal collagen gel cultures and each culture well was rinsed with 2–3 mL PBS, followed by fixation with 4% paraformaldehyde for 45 min at room temperature. The cultures were again washed with PBS and permeabilized with 0.5% Triton X-100 in PBS (PBT) for 10 min. Gels were incubated in a blocking solution (containing 1% bovine serum albumin [BSA] and 1% normal donkey serum in 0.25% PBT) for 1 h. Next, leech and PC12 cultures were treated with 1:500 rabbit α-tubulin antibody (Santa Cruz Biotechnology, Inc.) in blocking solution overnight at 4°C. The cultures were rinsed thrice with PBS and incubated for 45 min at room temperature with 1:100 Cy3 or Cy2-conjugated AffiniPure Donkey Anti rabbit secondary antibody in blocking solution (Jackson ImmunoResearch Laboratories, Inc.). For DRG labeling, gels were incubated with a mouse anti- Neurofilament H (NF-H) primary antibody overnight at 4°C (BioLegend). Following the incubation, gels were rinsed with PBS and incubated for 45 min at room temperature with Donkey anti mouse IgG H&L conjugated Alexa Fluor 488 for secondary antibody (Thermo Fisher Scientific).
Cultures were rinsed with PBS and gels were removed and placed in a porcelain-glazed cavity slide. Gels were mounted in an aqueous mounting medium (Immu-mount, Thermo Fisher Scientific) and covered with cover slips (diameter 20 mm, depth 3 mm). Confocal imaging was performed using Leica TCS SP5 to acquire fluorescent and bright-field images. To obtain confocal reflectance microscopy (CRM) images simultaneously with labeled cells, gels were maintained in PBS and imaged by a water immersion lens.
Collagen gel orientation setup
The method for strain-induced orientation in collagen gels was based on Vader et al. 47
Collagen gel samples, atop glass cover slips, were perforated with two 1-mm-diameter glass cylinders (A-M Systems) (capillaries). Glass capillaries were attached to two linear transducers (Newport) controlled by 562F-XYZ controllers (Newport). One capillary remained stationary, while the movement of the second capillary resulted in the local stretching of the gel. The tips, initially 6 mm apart were moved at 0.8 μm/s for 25 min to induce 16% stretch (final distance of capillaries 7.2 mm). This movement induced a local stretching of the gel matrix between the two tips. The glass capillaries were then removed from the gel permanently.
Morphometric analysis
We measured the morphometric parameters of neurons in 3D collagen gels and compared them to neurons plated on 2D Con-A substrates. A minimum of three samples for each category were considered for the purpose of analysis. Light microscopy images of 2D cultures and confocal 3D stacks of immunolabeled neurons in 3D gels were acquired for further analysis. To analyze 3D growth, data from confocal z-stacks were extracted manually and analyzed. XYZ coordinates of neurite endpoints, as well as soma coordinates, were imputed in a specialized MATLAB script, which plotted a representation of the spatial arrangement of each cell was in the form of a cuboid. Figure 5C shows an example of such a cuboid, with small circles representing neurite endpoints and a large circle indicating soma position. Measurements of cuboid axes were marked as growth in XYZ dimensions. The dimensions of growth are represented as cuboid axes ± standard error (SE).
The morphometric parameters measured were average neurite length per cell and neurites total branching tree length. To measure neurite length on 2D substrates we used NeuronJ, 53 and to measure neurite length in 3D collagen gels, we used Simple neurite tracer; 54 both are ImageJ plugins (US National Institutes of Health), which enable semiautomatic tracing of neurites and length measurements. Measurements of the morphometric parameters are summarized in bar charts. Error bars represent SEs, and p-value is shown as calculated by t-test.
Analysis of aligned collagen gels
To examine gel and fiber properties, SEM and CRM images were analyzed using ImageJ. The average fiber diameter was measured from SEM images and average pore size was measured from CRM images, results are presented as ± STD. To evaluate the efficiency of strain-induced alignment of collagen gels, light microscopy images of aligned gels were examined for matrix orientation. To maintain uniformity, images were acquired from the center of the stretched region. We used 2D Fast Fourier transform (FFT) of aligned gel images and compared them to control gels. Angular analysis of the FFT data was performed in accordance with the method described in Ayres et al. 55
As in control gels, confocal 3D stacks of immunolabeled neurons in aligned gels were analyzed using a MATLAB script and represented in the form of a cuboid (as in Fig. 5D). Average cuboid dimensions in x, y, and z axis are presented as ± SE. The cuboid's longest axis, the x axis, was marked as the growth direction and the orientation vector and angle (in the XY plane) were calculated from it. To compare neuronal alignment with collagen fiber orientation, we extracted individual fiber coordinates from CRM stack images and calculated the average vector and orientation angle (in the XY plane). The difference between the orientation of cell growth and collagen matrix alignment was designated as Δθ, and data are shown as a histogram (bins of 10°) (Figs. 5F and 6E).
Results and Discussion
Design and characterization of collagen gel for neuronal culture
We based our collagen gel on a known procedure. 49 The collagen mixture was prepared as described in the Materials and Methods section and 100 μL (3 mg/mL) was plated on top of a cover glass. The collagen drop was then allowed to solidify at 25°C in a humidified chamber for 40 min obtaining drop-shaped stable gels (Fig. 1) with a maximum height of 1.08 ± 0.04 mm (n = 3). We examined the gelation kinetics using turbidity analysis (Fig. 2A). The gelation process was recorded as a smooth sigmoidal shape showing a stable process. Gelation started 10 min after the mixture was removed from ice, reached 50% gelation after 12 min, and a complete gelation within 20 min. We examined the mechanical properties of the collagen gels by rheometry (Fig. 2B). The storage modulus (G′) measured was greater than the loss modulus (G′′), indicating a gel-like behavior. Rheometry measurements revealed soft gels with an average storage modulus of 141.32 ± 30 Pa as measured at 10rads/sec. These measurements correlate with elastic moduli of the brain and nervous tissue (0.1–1 kPa).56,57 To note, neurons show bias for growth and branching in softer gels (specifically within the Pa range of 50–500 Pa).26,58

Collagen gel characteristics.
To characterize the microarchitecture and fiber organization, collagen gels were imaged by SEM and CRM (Fig. 2C, D respectively). Collagen gels exhibited long, tangled, and nonoriented fibers. An average fiber diameter of 159.4 ± 80.8 nm was measured using SEM imaging and an average pore size of 3.7 ± 2.3 μm was measured by CRM images. Both fiber diameter and pore size are suitable for neurite outgrowth.10,26,59,60 Moreover, neuronal growth cones growing atop line-shape nanotopographical elements have been shown to exhibit maximal alignment and change in response to 150 nm high ridges. 8 We chose collagen concentration to be relatively high since high-density collagen fibers have been shown as better orienting cues for neuronal growth than low-density fibers. 32
Strain-induced alignment of collagen gels
To align the collagen gel fibers, we applied mechanical stimulation that induced an axial strain (as described in the Materials and Methods section). The level of strain induced on collagen gels was controlled using a semiautomated method. Collagen gels were perforated with 1-mm glass capillaries that were connected to transducers. One capillary remained stationary, while the second capillary was moved with programmed time and distance to induce local stretching of the gel matrix. We examined a range of stretching parameters and chose to work with a slow stretching rate that induced 16% stretch and resulted in significantly aligned gels, in accordance with the stretch previously described. 47
This stretch resulted in a local deformation of the collagen matrix between the placements of the two tips. Gel diameter was elongated by 1.2 mm from an initial 6-mm distance, and maximal height decreased to 0.69 ± 0.1 mm (n = 3). Increasing gel diameter and the initial distance between capillaries led to longer gel structures. By this simple single stretching method, we were able to align fibers of gels up to 1.2 cm long (1 cm before stretching) (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Fiber alignment at the centimeter scale is important for demonstrating feasibility of the method for bridging longer nerve gaps.
The gels maintained the deformation throughout all experiments. To evaluate the effect of strain on fiber orientation, light microscopy images of stretched and control gels were analyzed using 2D FFT (Fig. 3A, B). A normalized FFT plot of the stretched versus control images is shown in Figure 3C, demonstrating a clear spatial frequency of the stretched gel. The alignment of individual collagen fibers can be detected on the microscale as can be seen in high-resolution SEM and CRM microscopy images (Fig. 3D, E).
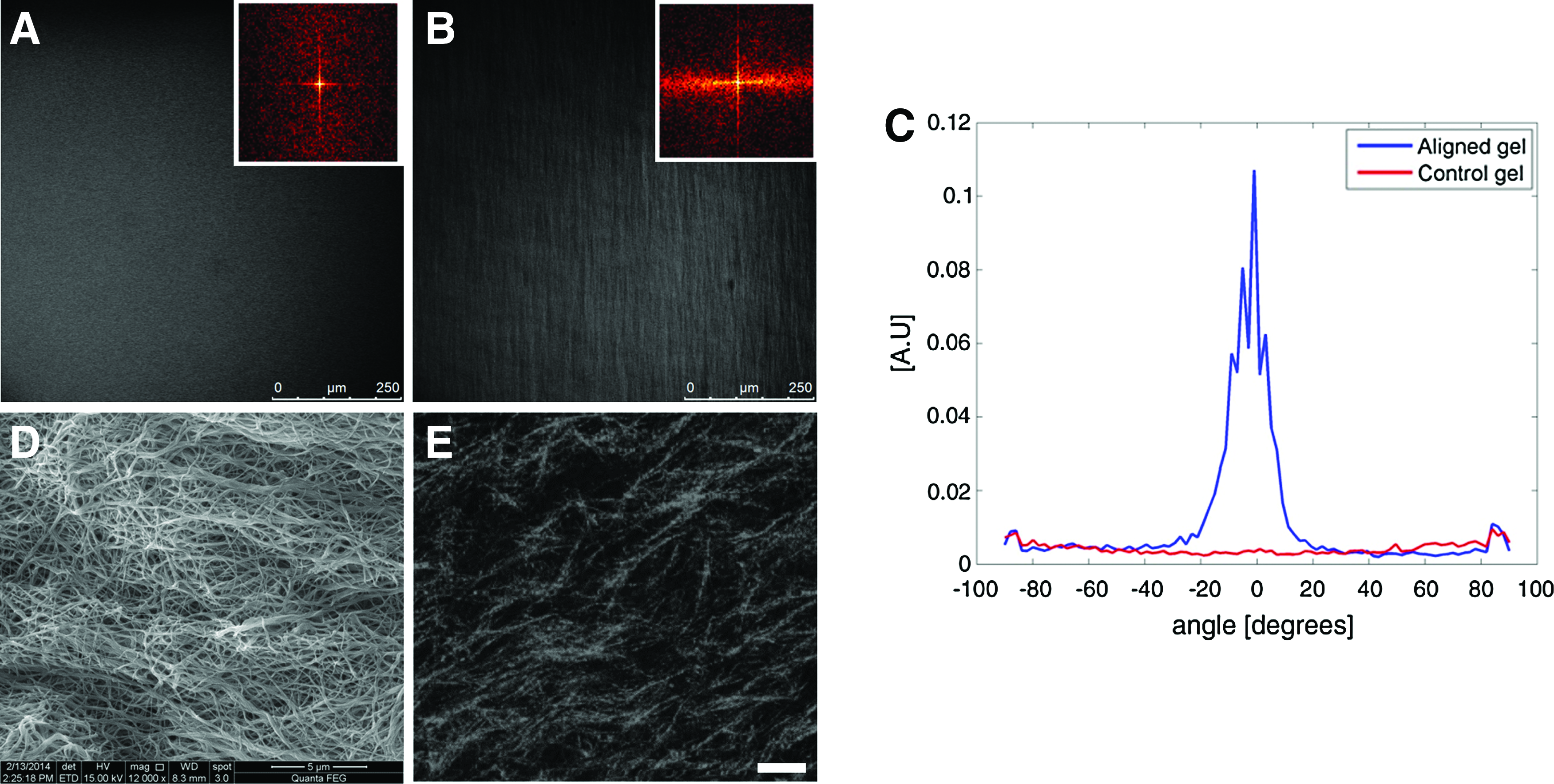
Effects of strain-induced alignment on collagen gels. Confocal microscope images of
Primary neuronal culture within 3D collagen gels
Isolated neurons were embedded in collagen gels before gelation and their growth was followed for a week (as described in the Materials and Methods section). It can be seen that 2 days after seeding, neurites already initiated from the cell soma (Fig. 4A). Within several days, neurites developed into a 3D complex dendritic tree (Supplementary Movie S1). To evaluate neuronal survival, we used a live/dead assay that indicated high viability of 85% of the cells. To further examine neuronal vitality, the electrophysiological activity of neurons embedded in collagen gels was measured with a standard single-electrode current-clamp intracellular recording technique. An example of an action potential of a leech neuron grown in 3D collagen gel films is presented in Figure 4B. Spontaneous, as well as current-responsive action potentials were recorded, exhibiting similar shapes and amplitudes to action potentials recorded from an intact leech ganglia chain (Supplementary Fig. S2). 61 To examine the neuronal pathfinding through the gel pores, neurons embedded within the gels were scanned with high-resolution microscopy. As can be seen in Figure 4C–E, the neurites intertwined with the collagen fibers as they grew, navigating through the pores.
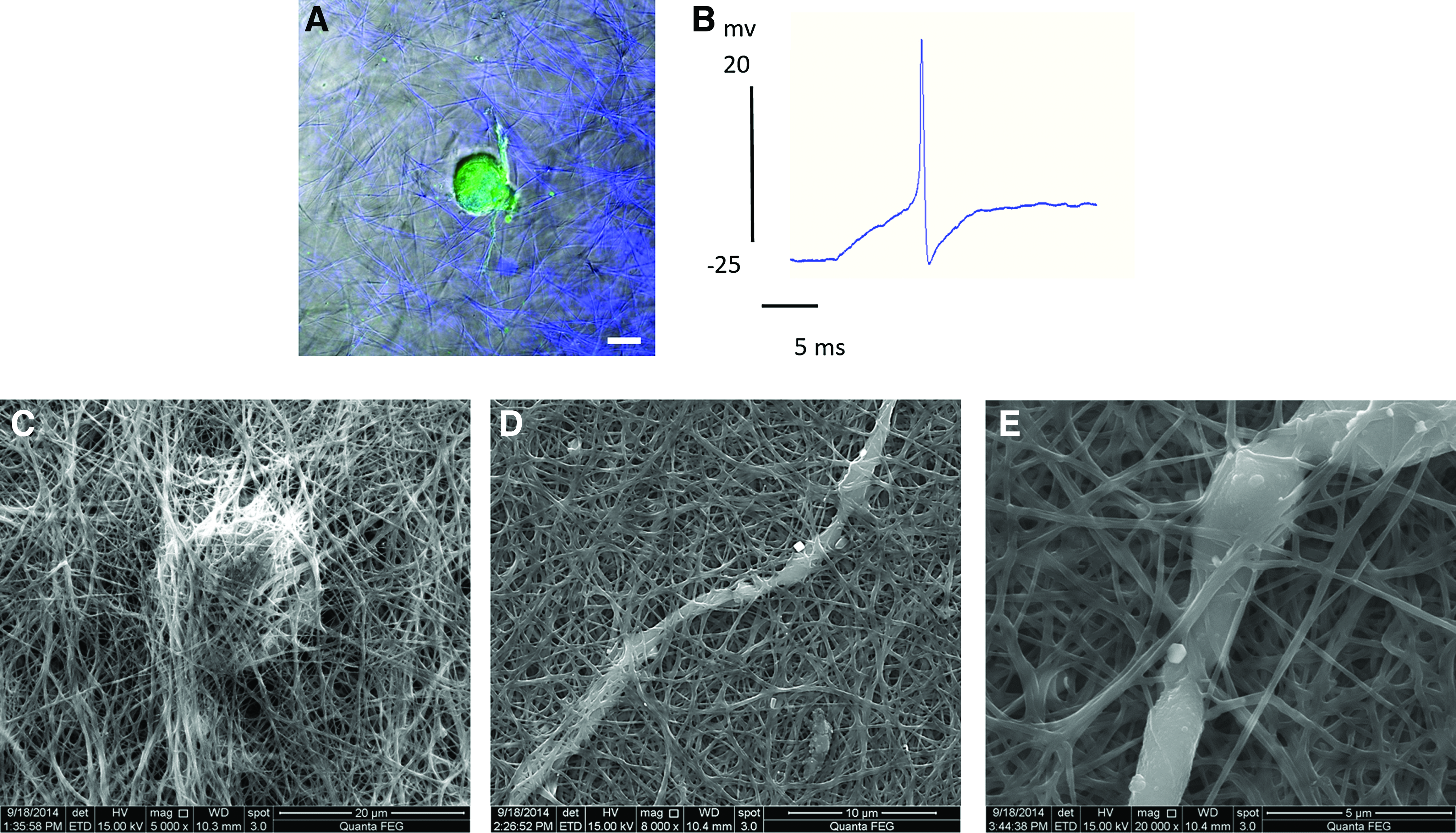
Leech neurons in 3D culture.
Neuronal growth in 3D aligned gel
Neurons were embedded within collagen gels that were then strained to induce maximal fiber alignment. Neurons grown in aligned gels show similar viability percentages as in control gels (90%). Following 7 days in culture, cells were immunostained and imaged, to evaluate the effect of collagen fiber orientation on neuronal growth. An elongated growth pattern was observed in neurons within the stretched gel, but not in cells growing in control gels (Fig. 5A, B). To analyze and quantify 3D growth and cell orientation, data from confocal z-stacks were extracted manually. Neurite endpoints as well as cell soma coordinates were detected and a representation of the spatial arrangement of each cell was plotted in the form of a cuboid. This method allowed us to analyze spatial parameters such as the dimensions of growth per axis and level of neuronal elongation. An example of cuboids for a control cell and a cell within a stretched gel is presented in Figure 5C and D. Cuboid dimensions were measured to compare the shape of growth and elongation of cells grown in aligned gels to cells grown in control gels. Since the x and y axes are interchangeable, we designated the x axis to the longest axis of growth and y to the second axis of growth. Neurons grown in control gels x axis (275 ± 21 μm), y axis (170 ± 12 μm), and z axis (87 ± 9 μm) and within the aligned matrix were measured in x axis (285 ± 28 μm), y axis (145 ± 17 μm), and z axis (27 ± 5 μm).
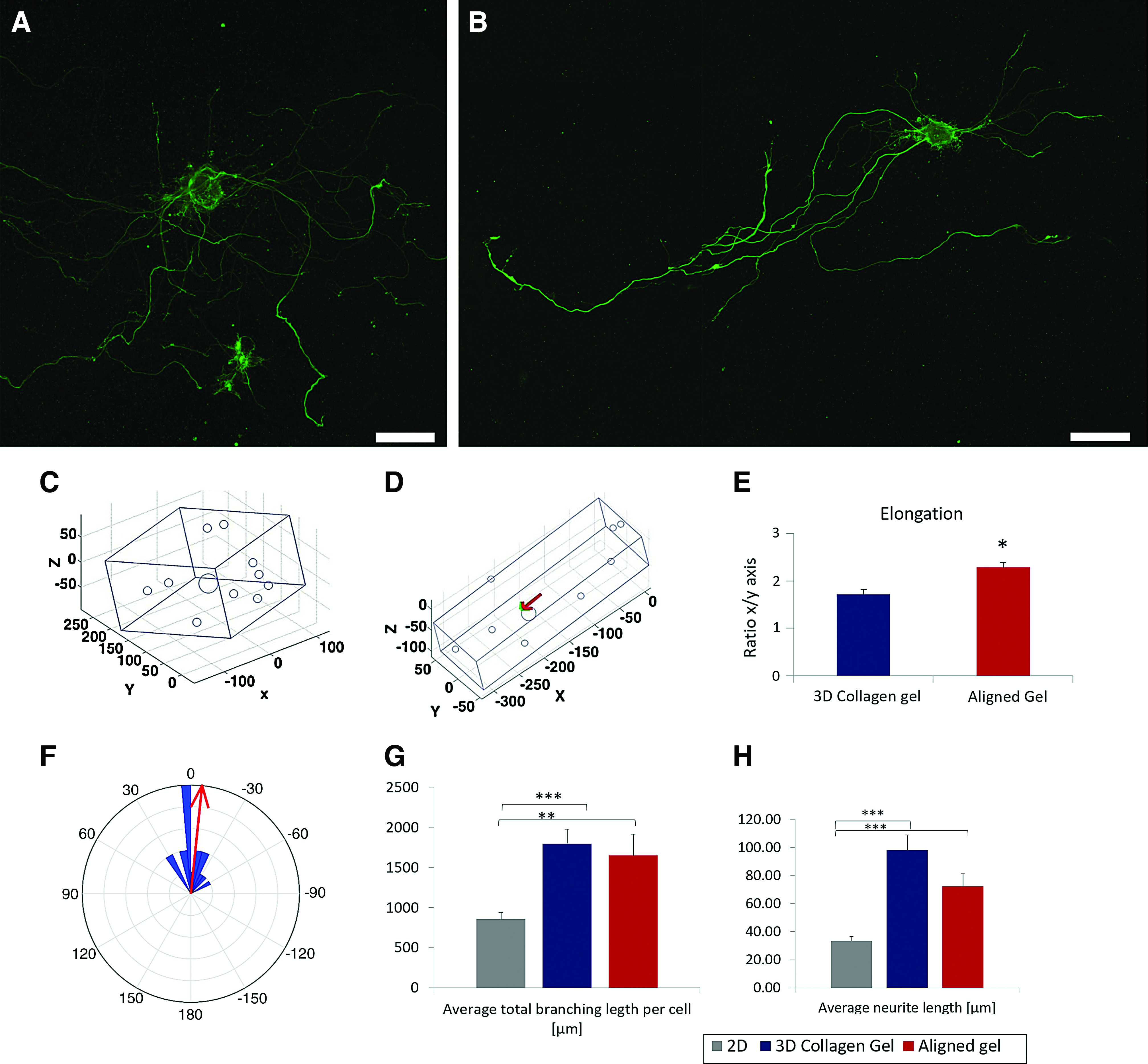
Neuronal orientation and morphometric parameters of growth within aligned gels. Confocal images of α-tubulin-stained leech neurons 7 days after seeding
The ratio between the x axis (longest axis) of growth and the y axis gave an elongation measurement. Cells grown in aligned collagen matrix showed elongated growth patterns compared to cells grown in 3D collagen control gels (Fig. 5E). To examine the direction of neuronal growth in comparison to collagen alignment, measurements of the longest axis direction were marked as the direction of growth. Collagen matrix orientation was measured using vectors extracted from collagen gel images. The angle between neuronal orientation and collagen gel orientation in the XY plane (Δθ) was measured and deviation from orientation was quantified (Fig. 5B). Average deviation was measured as −5.9° ± 5.3°. Fifty-five percent of the cells grow within Δθ 15° and 22%, additionally, within a margin of Δθ 30°. A histogram of Δθ results is shown in Figure 5F.
Low-density leech neuronal cultures allow single-cell measurements of neurite branching parameters. Morphometric parameters of neurons grown in aligned gels were measured and compared to growth in control 3D gels and commonly used 2D cultures (Con-A substrates as described in Materials and Methods section). Neurons grown in aligned gels exhibited average total branching length per cell (1650 ± 265 μm), similar to cells grown in 3D control gels (1799 ± 180 μm, p = 0.584) and which were significantly longer then neurons grown on 2D substrates (856 ± 81 μm, p < 0.01 and p < 0.001, respectively) (Fig. 5G) Average neurite length of neurons grown in aligned gels (72.4 ± 8.8 μm) showed similar results to 3D growth (98.1 ± 10.7 μm, p = 0.13) and were also significantly longer compared to neurons grown on a 2D Con-A substrate (33.5 ± 3.0 μm, p < 0.001) (Fig. 5H). To note, neurons grown on Con-A substrates present better growth than on other substrates, including collagen-coated plates (not shown).
PC12 growth within 3D aligned gel
To further validate the mechanically aligned collagen gel system for directing 3D neuronal cultures, we used neuron-like rat pheochromocytoma PC12 cells as a mammalian neuronal analog. PC12 cells differentiate when treated with NGF and exhibit neuronal properties, such as extruding long processes similar to neurites. Similar to leech neuronal cultures, PC12 cells were embedded in collagen solution before gelation, and following solidification, gels were either subjected to stretching or were maintained as control gels. PC12 cells grown on the 2D substrates for a week have shown no preference in the direction of neurite growth (Supplementary Fig. S3). Immunostained PC12 cells grown for 1 week within a control 3D collagen gel (Fig. 6A and Supplementary Movie S2) exhibited isotropic neurite branching, while within mechanically aligned collagen gels, neurite outgrowth displayed an orientation of growth that coincided with mechanical stretching and fiber orientation (Fig. 6B and Supplementary Movie S3). Furthermore, high-magnification SEM images of PC12 cells grown on top of collagen gels (Fig. 6C) and mechanically aligned collagen gels (Fig. 6D) show neurite branching when attached to the collagen fibers. To quantify dimensions of growth within 3D collagen gels, data from confocal z-stacks were analyzed using the MATLAB script used for primary neuronal culture analysis. Cuboid dimensions of PC12 cells grown within control 3D collagen gels were measured in PC12 growth within the mechanically aligned matrix, measured in x axis (324 ± 18.9 μm), y axis (59 ± 4.53 μm), and z axis (32 ± 3.19 μm). These yielded significant elongation of cellular growth within stretched gels, compared to control gels (Fig. 6E). To verify that the orientation of neurite growth corresponded to the stretching alignment direction, Δθ was measured and deviation from orientation was quantified. Average deviation was measured as −5.39 ± 1.6. 80.7% of the cells have grown within Δθ of 15°, and 15% additionally within a margin of Δθ of 30°. A histogram of Δθ results is shown in Figure 6F.

PC12 neurite orientation within aligned collagen gels. Confocal images of α-tubulin-stained PC12 cells 7 days after seeding
DRG explants
To demonstrate the orienting effect of mechanically aligned collagen gels on neurite extension and outgrowth of a population of neurons, we encapsulated whole mouse DRG within aligned collagen gels. Mouse DRG neurons normally show radial outgrowth from their explant and are known to follow nanotopographic cues, such as fibers, on 2D substrates or within 3D scaffolds.62–64 In this study, whole DRG explants were embedded within collagen solution before gelation, and following solidification, gels were either subjected to stretching or maintained as control gels. To validate the guidance of neurite extensions within the mechanically aligned gels, we stained gel cultures with an antineurofilament and simultaneously imaged gel fibers. After 1 week of growth within control collagen gels, DRG explants showed extensive neuronal outgrowth with no preferred direction (Fig. 7A). Within aligned gels, we observed directed neurite extension guided by fiber alignment (Fig. 7B and Supplementary Movie S4). Quantification of neurite orientation revealed that within aligned gels, 87.8% of neuronal extensions grew within Δθ of 30°.We further measured the average angle of neurite orientation (between −90° and 90°) and calculated the standard deviation (σ). Neurites within aligned gels exhibit low σ of ±15.9°, while within control gels, a high σ of ±48.7° was measured. This result indicates no preferred direction of growth without aligned collagen fiber cues and shows a high level of neuronal alignment with narrow distribution relative to collagen fiber direction. Similar level of directionality was presented in recent articles presenting nanotopographical fiber cues, other than collagen, for directing neuronal growth. For example, Berns et al. developed aligned peptide amphiphile gels presenting ECM epitopes and Kim et al., designed synthetic polymer-based aligned constructs.22,62
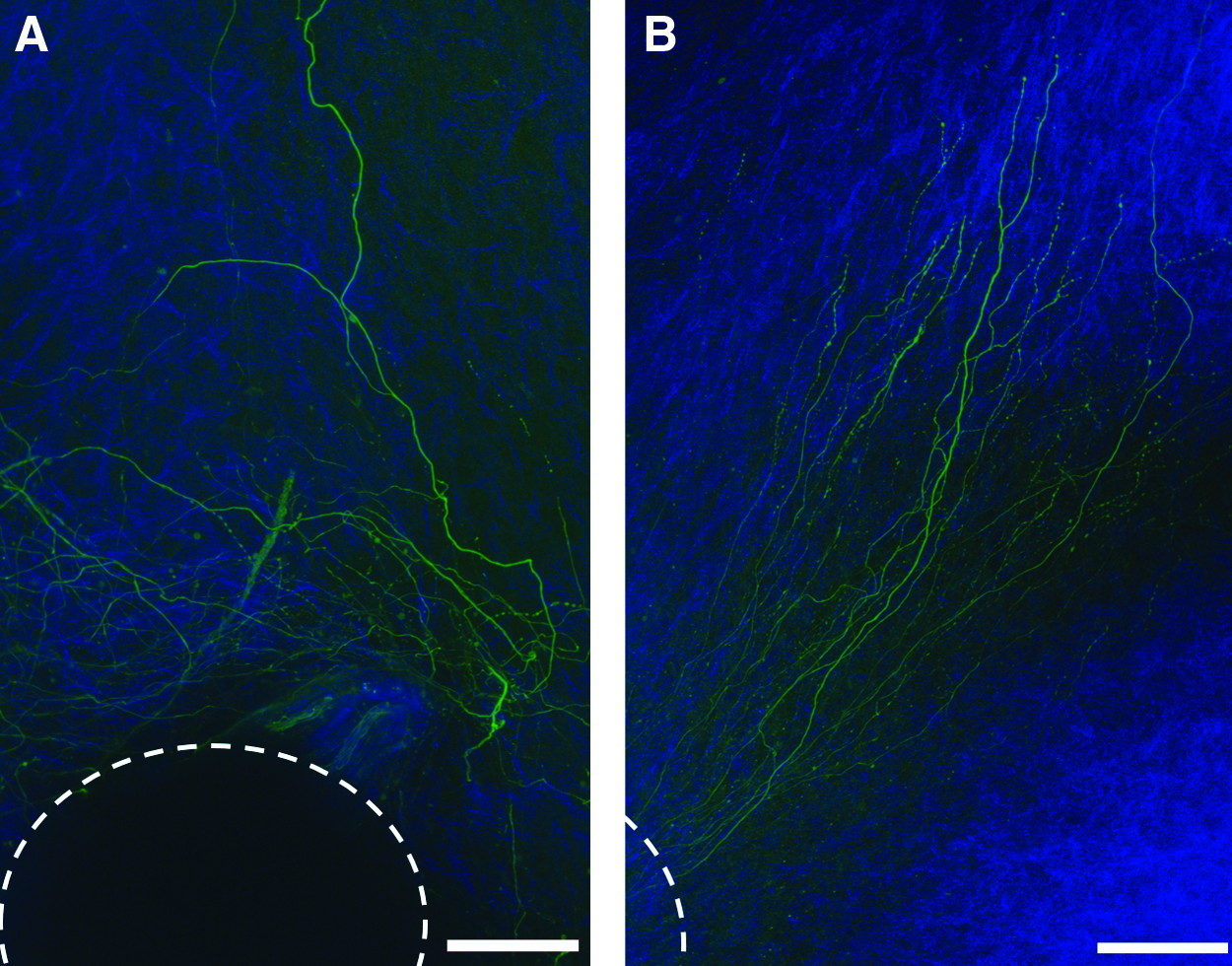
Confocal images of neurofilament-stained DRG explants grown for 1 week in
Conclusions
There is a growing need for fabrication of 3D scaffolds that can direct neuronal growth for effective regeneration posttrauma. Specifically, directed growth is required following peripheral nerve injuries where relatively long gaps may occur. Aligned nanotopographical features have been suggested as useful guidance cues for neuronal outgrowth. Oriented collagen gels have previously been designed as nerve guide fillings showing potential therapeutic applications for both peripheral and central nervous system.38,46,65–68 In this study, we propose a mechanically strained collagen gel with oriented fibers as a 3D platform for directing neuronal growth. First, we showed that neurons in 3D gels exhibit enhanced growth compared to 2D substrates. Then, to achieve an oriented 3D topography, we aligned the collagen matrix using uniaxial strain on the gels. We used a single short-range mechanical strain stimulation on gels that resulted in aligned 3D collagen matrices even at the fiber level. We evaluated leech single-cell neuronal growth patterns, as well as PC12 cells and whole DRG explants, in aligned gels and presented elongated and directed morphology. Our results, as demonstrated in all three model systems, show high level of neuronal alignment relative to collagen fiber direction.
As this method is easily reproducible and the effects on neuronal regeneration patterns are significant, it offers a simple method for designing oriented implants for neuronal repair therapeutics.
Footnotes
Acknowledgments
We thank Smadar Klugman from Tel-Aviv University for her help and advice with the rheological experiments. We thank Yakov Langsam from the Bar Ilan Institute of Nanotechnologies and Advanced Materials for valuable help with the SEM imaging. We thank Itamar Greenberg and Ziv Shemesh for assistance with morphometric measurements. This work was supported, in part, by the Israel Science Foundation for Individual Research Grants (1053/15).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.