
Abstract
Tendon and ligament injuries are a persistent orthopedic challenge given their poor innate healing capacity. Nonwoven electrospun nanofibrous scaffolds composed of polyesters have been used to mimic the mechanics and topographical cues of native tendons and ligaments. However, nonwoven nanofibers have several limitations that prevent broader clinical application, including poor cell infiltration, as well as tensile and suture-retention strengths that are inferior to native tissues. In this study, multilayered scaffolds of aligned electrospun nanofibers of two designs–stacked or braided–were fabricated. Mechanical properties, including structural and mechanical properties and suture-retention strength, were determined using acellular scaffolds. Human bone marrow-derived mesenchymal stem cells (MSCs) were seeded on scaffolds for up to 28 days, and assays for tenogenic differentiation, histology, and biochemical composition were performed. Braided scaffolds exhibited improved tensile and suture-retention strengths, but reduced moduli. Both scaffold designs supported expression of tenogenic markers, although the effect was greater on braided scaffolds. Conversely, cell infiltration was superior in stacked constructs, resulting in enhanced cell number, total collagen content, and total sulfated glycosaminoglycan content. However, when normalized against cell number, both designs modulated extracellular matrix protein deposition to a similar degree. Taken together, this study demonstrates that multilayered scaffolds of aligned electrospun nanofibers supported tenogenic differentiation of seeded MSCs, but the macroarchitecture is an important consideration for applications of tendon and ligament tissue engineering.
Introduction
T
Surgical reconstructions, such as those performed for anterior cruciate ligament (ACL) tears, typically utilize autografts and allografts. The former is limited by availability and donor site morbidity, while the latter carries the risk of disease transmission and impaired host integration. As a result, there is continued interest in developing novel biomaterials that can serve as engineered grafts for surgical reconstruction or scaffolds capable of augmenting primary repairs. In both cases, the ideal tissue-engineered construct would possess the following characteristics: (1) biodegradable and biocompatible, (2) promote a fibroblastic phenotype of exogenously seeded or endogenously recruited cells, (3) possess topographical or chemical cues that recapitulate native anatomy, structure, and biochemical composition, (4) possess material properties comparable to native tissue, and (5) possess a suture-retention strength equal to, or exceeding, that of the surgical repair.
Tissue engineers have utilized electrospinning to fabricate scaffold sheets comprising aligned nano/microfibers, mimicking the aligned collagen fibrils of native tendon and ligament.3–5 Modulation of fiber alignment and diameter affects cell proliferation, differentiation, and matrix deposition,6–8 while polymer composition predictably controls degradation rate and mechanical properties of the scaffold. 9 While the material properties of electrospun fibrous sheets have been shown to approach those of native tendon and ligament,4,5 adapting scaffold sheets for clinical application presents several challenges.
During electrospinning, as fibers collect on a grounded mandrel, there is a reduction in the strength of the electrostatic field, ultimately limiting sheet thickness to several millimeters. The resulting scaffold possesses high porosity but small pore sizes, especially when fibers are aligned, limiting cell infiltration. 10 Several methods, such as inclusion of sacrificial fibers, 11 ultrasonication, 12 or laser irradiation, 13 have been shown to increase pore size and cell infiltration, but often compromise fiber architecture or mechanical strength. Conversely, individual sheets of cell-seeded electrospun scaffold can be combined as multilayered constructs to produce a thicker scaffold with more homogeneous cell distribution while still preserving the original fiber topography. 14 Nevertheless, multilayered constructs of nonwoven electrospun fibers remain limited in size and often possess inferior material properties to native tendon and ligament. In addition, electrospun scaffold sheets possess poor suture-retention strength, limiting their use as reconstructive grafts or devices capable of providing mechanical support to primary repairs. 15
The application of textile patterns to biomaterial fabrication has been recently explored in an effort to overcome these limitations of nonwoven electrospun scaffolds. 16 Braided microfibrous grafts designed for ACL reconstruction possessed good mechanical properties and cytocompatibility while promoting tissue regeneration when implanted in vivo.17–19 Braided scaffolds of aligned poly-L-lactic acid (PLLA) nanofibers were reported to support tenogenic differentiation of human mesenchymal stem cells (MSCs) and possessed a toe region in the stress–stress curve reminiscent of native tendons. 20 More recently it was demonstrated that scaffold properties and cell behavior were affected by braiding angle and polymer composition (i.e., differing ratios of PLLA and poly-ɛ-caprolactone [PCL]). 21 While the effect of fiber diameter (i.e., nano vs. micro) on cell behavior has been investigated,6,22 the effect of macroscale architecture of multilayered nanofibrous scaffolds remains unknown, despite their intended use for similar tendon and ligament injuries. Using a microphotopatterning model system to precisely control substrate architecture, Gilchrist et al. 23 found that micro- and mesoscale topographical cues could be combined to promote synergistic or antagonistic effects on tenogenic differentiation.
Stacked and braided scaffolds ostensibly possess similar nanoscale cues (i.e., aligned nanofibrous polymers), but with different macroscale architecture. A better understanding of the effect of scaffold design (stacked vs. braided) and polymer composition (PCL vs. PLLA) of aligned nanofibers on human MSC behavior is clearly needed to optimize functional tissue engineering scaffolds. In this study, we tested the hypothesis that (1) braided scaffolds would possess greater tensile and suture-retention strength than stacked scaffolds and that (2) stacked scaffolds, by virtue of possessing greater surface area, would permit a greater degree of cell proliferation, cell infiltration, and matrix deposition, while gene expression would not differ between groups.
Materials and Methods
Study design
To examine the effect of scaffold design and polymer type on mechanical properties and tenogenic differentiation of seeded MSCs, four scaffolds were fabricated as follows: (1) braided PCL, (2) braided PLLA, (3) stacked PCL, and (4) stacked PLLA. First, aligned nanofibrous sheets of PCL and PLLA were electrospun and characterized. Second, mechanical properties and suture-retention strengths of acellular scaffolds were determined. Finally, scaffolds were seeded with human MSCs, which attached and underwent tenogenic differentiation, as confirmed by assays for gene expression, histological staining, and biochemical composition.
Electrospun nanofibers
Electrospinning aligned nanofibers
Sheets of aligned nanofibers were fabricated from a solution of PCL (MW = 70k–90k, Sigma-Aldrich, St. Louis, MO) prepared at 15% w/v in 1:1 (v/v) tetrahydrofuran (THF, Sigma-Aldrich):dimethylformamide (DMF, Fisher Scientific, Pittsburgh, PA) or PLLA (MW = 40k–70k, Polysciences, Warrington, PA) prepared at 10% w/v in 4:1 (v/v) dichloromethane (DCM, Fisher):DMF. 9 Polymer solutions were loaded into 10 mL syringes and extruded through an 18-gauge blunt tip needle at 3.0 mL/h using a syringe pump (PY2 70,2209, Harvard Apparatus, Holliston, MA) (Fig. 1A). The needle tip was placed 10 cm from a custom-designed cylindrical mandrel, which rotated at a surface velocity of 10 m/s. 10–18 kV DC potential (Gamma High Voltage, Ormond Beach, FL) was applied to the polymer solution, while an 8 kV potential was applied to two aluminum shields placed perpendicular to the mandrel axis but parallel to the needle axis, narrowing the width of the aligned nanofibrous sheet collected on the grounded mandrel.
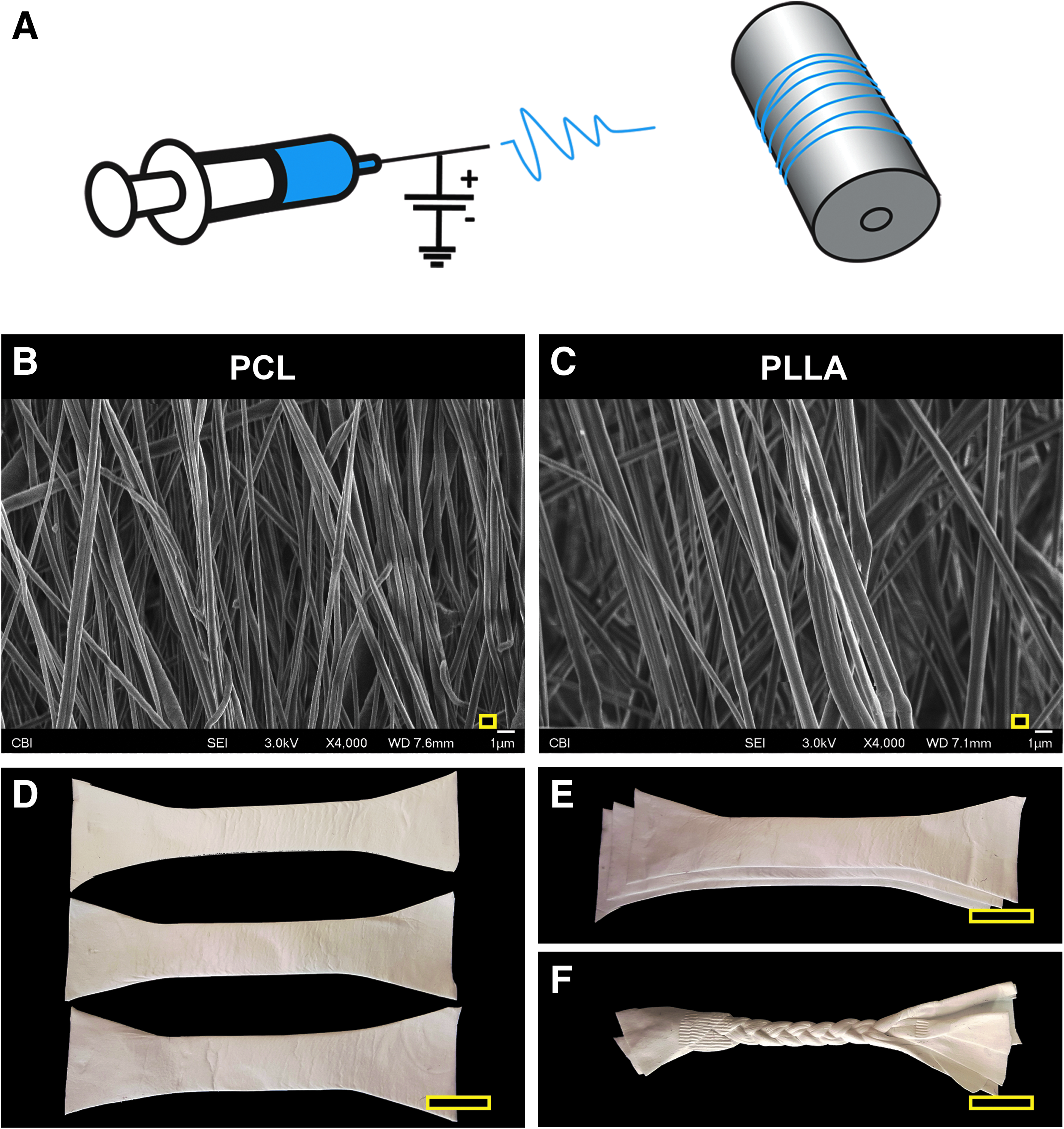
Fabrication of multilayered nanofibrous scaffolds.
Fiber characterization
Acellular scaffolds were dried under vacuum and mounted on aluminum stubs, sputter-coated with 4.5 nm of gold, and imaged by a scanning electron microscope (SEM, field emission, JEOL JSM6335F, Peabody, MA) operated at 3 kV accelerating voltage and 8 mm working distance. Fiber diameter, fiber alignment, and pore size were determined using ImageJ (National Institutes of Health, Bethesda, MD) (Fig. 1B, C). More specifically, 100 total fibers were analyzed across four independent SEM images for a given scaffold type. The width of a fiber was measured at three locations spanning its length; the average value was taken as the diameter of the fiber. The angle of a fiber relative to a horizontal line was measured for 100 fibers; fiber angles were normalized against the mean angle (set at 0.0°) to determine the standard deviation of fiber orientation of scaffolds composed of different polymers. To determine pore size, the areas of polygonal regions encompassed by intersecting fibers were measured.
Braided and stacked multilayered scaffolds
Multilayered scaffold fabrication
Three individual nanofibrous sheets (∼500 μm thickness) were cut using a dog bone-shaped template with the central portion measuring 3.75 × 0.75 cm (aspect ratio 5:1) (Fig. 1D). The three sheets were either manually stacked (Fig. 1E) or rolled and braided (Fig. 1F), with the latter consisting of 6.3 ± 0.3 stitches/cm (16 ± 0.8 stitches/inch). 21 The stitch number per unit length was determined using digital calipers, with a range of 14–18 stitches/inch. There was no difference between PCL and PLLA scaffolds.
Tensile test
Cross-sectional area (CSA) was determined by a custom-made scanning micrometer and corroborated with digital calipers. CSA was determined at three positions (top, middle, and bottom), with the average used to determine material properties following tensile testing. The multilayered scaffolds were then clamped into a material testing machine (Instron, Model 4502, Norwood, MA) and loaded under tension in a direction parallel to nanofiber alignment. After preloading to 0.5 N, scaffolds were preconditioned from 0% to 2% strain for 15 cycles at 7.5 mm/min before undergoing load to failure at the same elongation rate. A custom digital motion tracking system (Spica Technology, Kihei, Maui, HI; 0.01 mm accuracy) was used to track the vertical displacement of the strain markers (black pen) using a single video camera aligned perpendicular to the plane. These data were inputted into ABAQUS software (ABAQUS/CAE Student Version 6.4; Simulia, Providence, RI) to determine strain. Both structural (ultimate load, ultimate elongation, stiffness, yield load, and yield elongation) and material properties (ultimate strength, ultimate strain, modulus, yield strength, and yield strain) were determined. Notably, stiffness (modulus) was determined from the slope of the linear region of the load-elongation (stress–strain) curve, while yield load (stress) and elongation (strain) were taken following 0.02% offset, as described previously. 21
Suture-retention test
Scaffolds were clamped on both ends, as in the tensile testing protocol described above, and preloaded to 0.5 N. At a distance 2 mm proximal and distal to the scaffold midpoint, 4–0 Vicryl suture was run horizontally back and forth through all layers of the stacked and braided scaffolds to bind each sheet (braid) with the others. The suture was secured with a knot, and the scaffold was then transected, providing two constructs for suture-retention testing. A single loop of 3–0 PROLENE suture was passed through the midline at a distance of 7 mm from cut edges (i.e., 5 mm from the horizontal bridge suture) and secured to an immovable cylindrical rod mounted on the material testing machine (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). A preload of 0.5 N was applied before loading to failure at 7.5 mm/min. The maximum load was recorded as the suture-retention strength.
Cell-seeded layered scaffolds
Human bone marrow-derived MSCs were obtained from femoral heads of three patients undergoing total hip arthroplasty, as approved by the Institutional Review Board (University of Washington and University of Pittsburgh) and described previously. 24 The colony-forming capacity and multipotency of the MSCs were confirmed for each patient. MSCs were expanded in basal medium (Dulbecco's modified Eagle's medium, 10% v/v fetal bovine serum, 1% v/v penicillin-streptomycin-fungizone [PSF]; Gibco, Grand Island, NY) through passage 3 before use in experiments.
Three sheets of aligned PCL or PLLA nanofibers each measuring 5 × 25 × 0.25 mm were braided, as described above, or left as individual sheets that would be stacked following cell seeding. The braided scaffolds or individual sheets were placed in 6-well plates previously coated with 3 mm of silicone (Sylgard 184, Dow Corning, Midland, MI) and exposed to 30 min of UV irradiation (100–290 nm) followed by serial washes in 95% ethanol and then phosphate-buffered saline (PBS) containing 1% PSF. Scaffold sheets were soaked overnight at 4°C in basal medium to promote protein adsorption and enhance cell attachment. After aspiration of medium, 1.0 × 106 MSCs (P3) suspended in 300 μL basal medium were added to each braided scaffold or individual sheets (3.33 × 105 MSCs in 100 μL medium per sheet) and incubated for 1 h at 37°C before the addition of 3 mL of differentiation medium (Basal medium + 50 μg/mL ascorbic acid 2-phosphate + 10 ng/mL TGF-β3; Peprotech, Rocky Hill, NJ). Individual scaffold sheets were stacked and pinned at each end. Braided and stacked scaffolds were cultured for up to 28 days with changes in differentiation medium every 2–3 days. Concurrently, 1.0 × 105 MSCs were seeded in 6-well tissue culture plates and grown in either basal or differentiation medium. This two-dimensional (2D) substrate served as a control for cell differentiation assays, described below.
Real-time polymerase chain reaction analysis of gene expression
At days 3, 7, and 14, total cellular RNA was isolated from MSCs grown on either 2D plastic or 3D scaffolds through the sequential use of TRIzol (Thermo Fisher) and an RNeasy Plus Mini Kit (Qiagen, Valencia, CA). One thousand micrograms of RNA was reverse transcribed into cDNA using the SuperScript III First-Strand Synthesis Kit (Thermo Fisher). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed for 40 cycles using SYBR Green Supermix in a StepOnePlus real-time PCR machine (Applied Biosystem, Life Technology, Grand Island, NY). Primers for tenogenic differentiation–Scleraxis (Scx), Mohawk (Mkx), Tenomodulin (Tnmd), Tenascin C (Tnc), Collagen type I (Col1a1), Collagen type III (Col3a1)–and osteogenic differentiation runt-related transcription factor 2 (Runx2)–were used. The targets and sequences of the primers are shown in Supplementary Table S1. Relative gene expression was normalized with GAPDH, and scaffold constructs were compared with MSCs grown on 2D plastic utilizing the ΔΔCT method. Primer specificity and product size were confirmed through melt curve analysis and agarose gel electrophoresis, respectively.
Histology
On days 14 and 28, stacked and braided scaffolds were washed in PBS and fixed overnight in 4% paraformaldehyde. After serial soaking in an increasing sucrose gradient (10%, 20%, and 30%, for 2 h each), scaffolds were embedded in Tissue-Tek® O.C.T. Compound (Fisher Healthcare, Houston, TX), and sections of 18 μm thickness were obtained using a cryotome (CM1850, Leica, Buffalo Grove, IL). Constructs were sectioned either parallel (i.e., longitudinal) or transverse to the direction of electrospun nanofibers. After rehydration in ddH2O, sections were stained either with hematoxylin and eosin (H&E) or 4′,6-diamidino-2-phenylindole (DAPI, Thermo Fisher) to visualize matrix deposition and cell infiltration, respectively. Cell morphology of cells grown on each scaffold type was determined in both the transverse and longitudinal directions. The aspect ratio (length/width) of 50 cells for a given scaffold type and direction was calculated from H&E stained sections imaged at high magnification and processed with ImageJ (NIH).
Biochemical composition
On days 1, 14, and 28, constructs were washed in PBS and dried overnight at 65°C. After obtaining measurements of dry weight, samples were digested overnight at 65°C in a papain digestion buffer (1:50 papain from papaya latex, Sigma-Aldrich; 0.1 M sodium acetate, 0.01 M cysteine HCl, 0.05 M EDTA, dissolved in ddH2O, pH 6.0 adjusted with HCl).
Total collagen content
Total collagen was determined using a modified hydroxyproline assay. Briefly, 200 μL of each sample was hydrolyzed with an equal volume of 4 N sodium hydroxide at 121°C for 75 min, neutralized with an equal volume of 4 N hydrochloric acid, and then titrated to an approximate pH of 7.0. The resulting solution was combined with 1.2 mL chloramine-T (14.1 g/L) in buffer (50 g/L citric acid, 120 g/L sodium acetate trihydrate, 34 g/L sodium hydroxide, and 12.5 g/L acetic acid) and allowed to stand at room temperature for 30 min. The solution was then combined with 1.2 mL of 1.17 mM p-dimethylaminobenzaldehyde in perchloric acid and placed in a 65°C water bath for 20 min. Two hundred microliters of each sample was added to a clear 96-well plate, in triplicate, and absorbance at 550 nm was read.
Sulfated glycosaminoglycan and dsDNA concentrations
Sulfated glycosaminoglycan (sGAG) concentrations were quantified with a Blyscan Assay, according to manufacturer's instructions (Biocolor, Carrickfergus, United Kingdom), while Quant-iT PicoGreen dsDNA assay (Invitrogen) was used to determine dsDNA concentrations.
Statistical analysis
Data are expressed as mean ± standard deviation. For all assays, ≥6 experimental samples were used for a given timepoint. Specific sample sizes for an assay are indicated in figure legends. One-way ANOVAs with post hoc Tukey tests were performed to compare across groups at a given timepoint, with significance considered at p < 0.05.
Results
Scaffold mechanical properties
Sheets of PCL and PLLA nanofibers were similar in fiber diameter (PCL: 898 ± 291 nm; PLLA: 869 ± 265 nm), pore size (14.3 ± 14.0 μm2; 21.0 ± 14.2 μm2), and fiber alignment (0 ± 15.4°; 0.0 ± 10.2°). Structural properties of scaffolds are shown in Table 1. Of note, braided scaffolds of both polymers demonstrated greater ultimate and yield elongations compared to their stacked counterparts, with a corresponding reduction in stiffness, respectively. Conversely, braiding increased both the ultimate and yield loads compared to stacked scaffolds, but only for PCL scaffolds. Regardless of scaffold design, PCL scaffolds bore significantly more load than PLLA scaffolds. A similar pattern is found when scaffolds were normalized as material properties (Table 2). Namely, PCL scaffolds of both designs possessed superior tensile strength to PLLA scaffolds (Fig. 2A). The stacked PLLA scaffolds exhibited a greater modulus than stacked PCL scaffolds, although this effect was lost when scaffolds were braided (Table 2). When looking at a physiological strain range of 0–12% (Fig. 2B), only the stacked PLLA scaffold reached the yield point. In terms of suture retention, braided scaffolds of both polymers possessed increased strength compared against stacked scaffolds (Fig. 2C). Similar to the mechanical properties of the scaffolds, PCL scaffolds exhibited higher suture-retention strength than PLLA scaffolds.
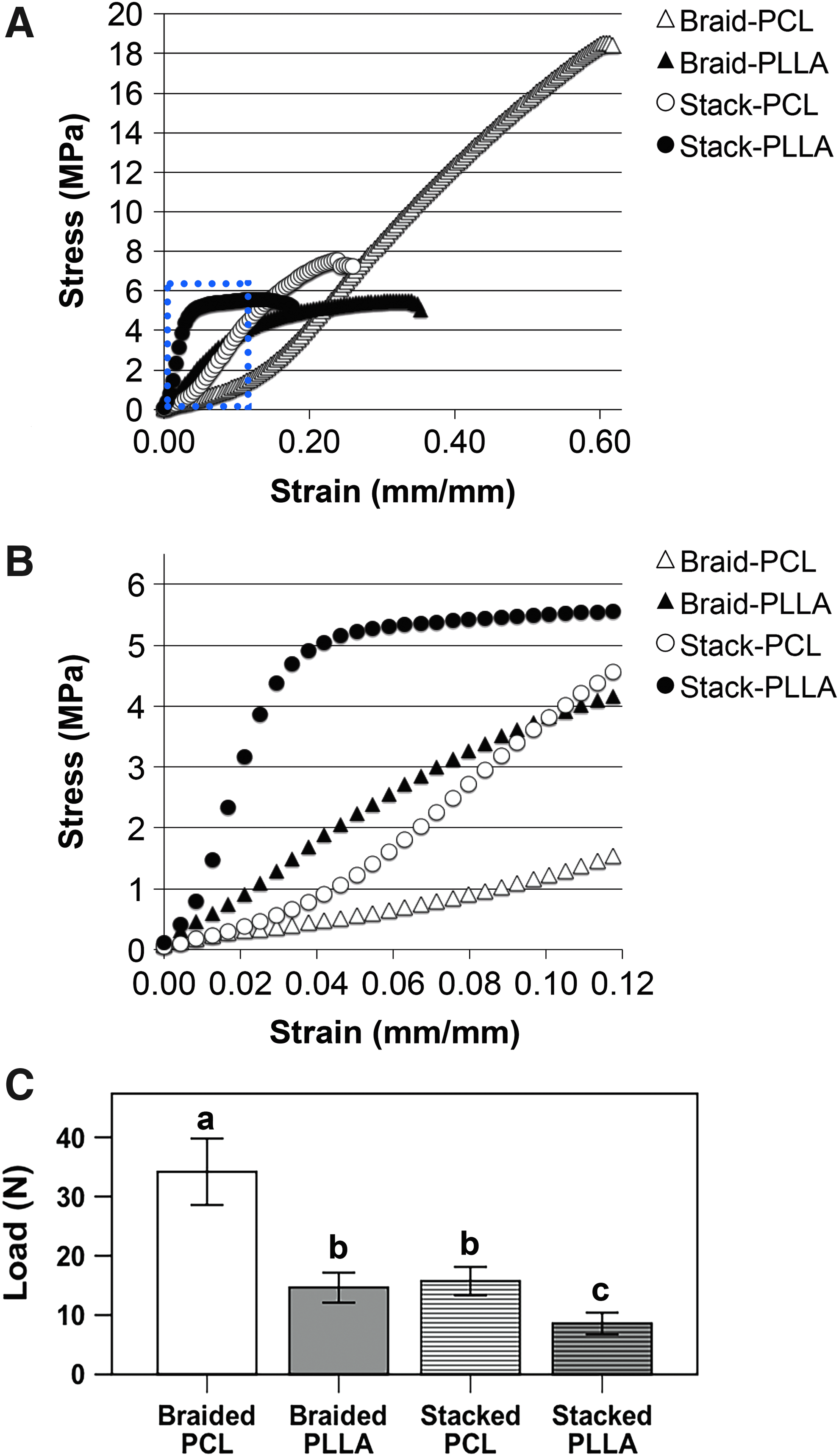
Mechanical properties of acellular scaffolds.
For a given scaffold design, significant difference between PCL versus PLLA, p < 0.05.
For a given polymer, significant difference between braided versus stacked, p < 0.05, n = 8 per group.
PCL, poly-ɛ-caprolactone; PLLA, poly-L-lactic acid.
For a given scaffold design, significant difference between PCL versus PLLA, p < 0.05.
For a given polymer, significant difference between braided versus stacked, p < 0.05; n = 8 per group.
CSA, cross-sectional area.
Gene expression profiles analyzed by qRT-PCR
Gene expression analysis is normalized against MSCs grown on 2D tissue culture plastic in basal growth medium to discern the effect of TGF-β3/ascorbic acid supplementation and scaffold architecture (Fig. 3). Compared against 2D controls (i.e., growth factor supplemented), all scaffolds upregulated Scx expression on day 3. By day 7, 2D controls demonstrated an ∼6.3-fold increase over the MSC control; braided scaffolds showed further upregulation, while stacked scaffolds expressed Scx comparable to MSC controls. By day 14, all groups began to normalize toward MSC controls, yet braided constructs remained elevated over 2D controls. In contrast, expression of tenogenic transcription factor Mkx and extracellular matrix (ECM) glycoprotein Tnc was not enhanced by scaffold topography at any of the measured timepoints. Rather, Mkx expression in the stacked scaffold was significantly less than 2D controls and braided PLLA scaffolds on days 7 and 14, respectively. A similar pattern was seen for Tnc expression, although all scaffolds were significantly inferior to 2D controls on day 14.
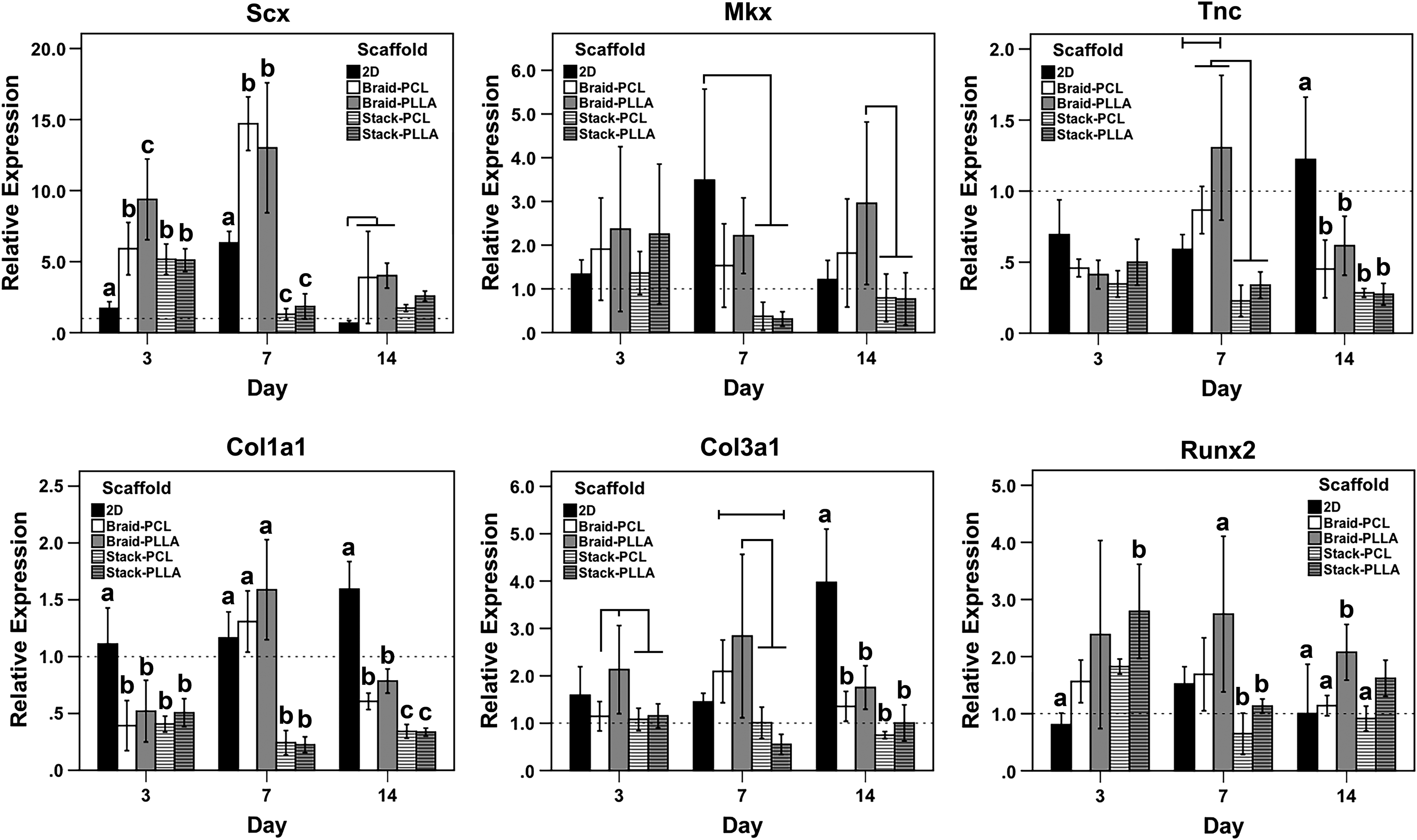
Gene expression analyzed by quantitative real-time polymerase chain reaction. Genes analyzed included Scx, Mkx, Tnc, Col1a1, Col3a1, and Runx2, from cultures harvested on days 3, 7, and 14. Letters correspond to statistically equivalent groups at a given timepoint. Statistical differences between particular scaffolds are further highlighted with horizontal lines above columns. n = 6 for each scaffold at each timepoint, p < 0.05.
Expression of Col1a1 and Col3a1 mRNA was subtly affected by growth factor supplementation, as 2D controls demonstrated an ∼1.5-fold and ∼4.0-fold increase in Col1a1 and Col3a1 expression, respectively, on day 14. Cells seeded on scaffolds tended to have comparable or reduced Col1a1 and Col3a1 expression at all timepoints, with stacked scaffolds promoting stronger downregulation than braided scaffolds. Tnmd expression (data not shown) was only discernible at day 14 for MSCs seeded on scaffolds (i.e., not MSC or 2D controls), with no differences across groups. Expression of osteogenic transcription factor, Runx2, was not significantly affected by TGF-β3/ascorbic acid supplementation with various scaffolds inducing very modest but significant upregulation over the culture period. By day 14, Runx2 expression was only upregulated in braided PLLA scaffolds (∼2.0-fold).
Histology
Stacked PLLA scaffolds demonstrated considerable swelling after 28 days of culture corresponding to a contracted length and width (Supplementary Fig. S2). At day 14, cells were predominantly located at the surface of individual nanofibrous sheets, regardless of design (Supplementary Fig. S3). However, by day 28, cells could be found through the entire depth of stacked scaffolds, with greater apparent cell number and homogeneity within stacked PLLA scaffolds (Fig. 4C, D, G, H). In contrast, cells were almost exclusively located on the surface of braided PCL scaffolds (Fig 4A, E), with braided PLLA scaffolds (Fig. 4B, F) exhibiting only modest cell infiltration compared to stacked constructs. H&E staining confirmed these observed trends in cell infiltration and demonstrated matrix deposition at sites of greatest cell density, namely, the surfaces of braided scaffolds and interface of layers in stacked constructs (Fig. 5). The effect of nanofiber direction on cell morphology was particularly evident when comparing longitudinal and transverse sections of stacked scaffolds. Cell nuclei were elongated in the direction of fiber orientation, as shown in the insets of Figure 5 and quantified as an aspect ratio (i.e., cell length/wide). Regardless of scaffold type, average cellular aspect ratios of ∼7 and ∼2 were found in the longitudinal and transverse sections, respectively (Supplementary Fig. S4).

Localization of cells in scaffolds on day 28 based on DAPI nuclear staining.
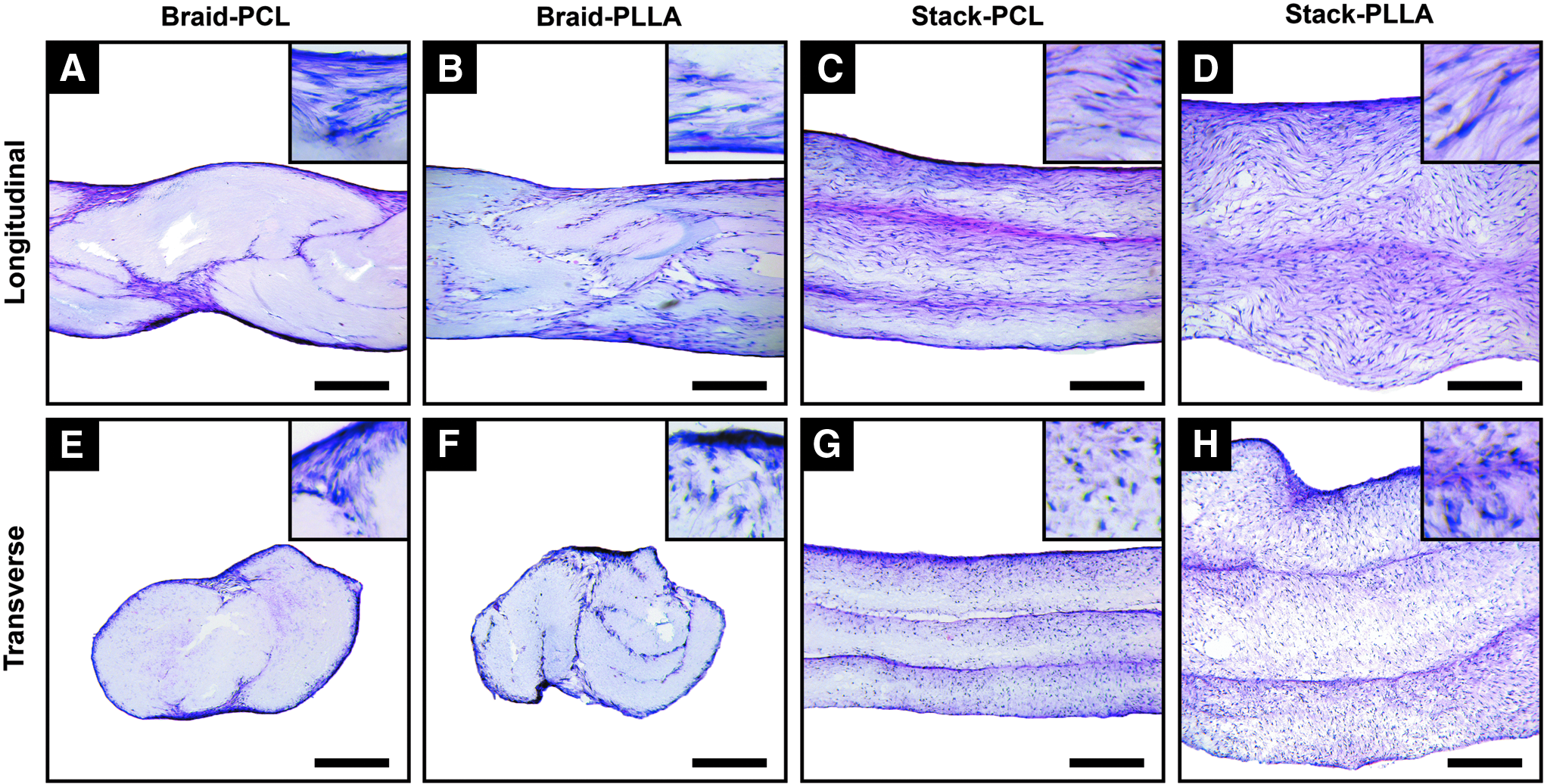
Histology of cell-seeded scaffolds on day 28 by hematoxylin and eosin staining.
Biochemical composition
Day 1 measurements of total dsDNA content suggested greater initial cell adhesion in stacked scaffolds (Fig. 6A). In general, greater cell proliferation was noted for scaffolds of stacked design and composed of PLLA (Fig. 6A). By day 28, stacked PLLA scaffolds possessed the greatest cell number, while braided PCL scaffolds held the least. Braided PLLA and stacked PCL scaffolds contained intermediate cell numbers and were equivalent when comparing the two groups. Similar trends were seen for total collagen content (Fig. 6B) and total sGAG content (Fig. 6C). Namely, stacked scaffolds supported the greatest matrix deposition by day 28, an effect further enhanced on scaffolds composed of PLLA. However, when collagen and sGAG contents were normalized against dsDNA contents, the effect of scaffold design was lost by day 28 (Supplementary Fig. S5). In particular, relative collagen deposition in PCL scaffolds was higher than in PLLA scaffolds, with no differences between scaffold designs (Supplementary Fig. S5A). By day 28, all groups showed equivalent relative sGAG content (Supplementary Fig. S5B).
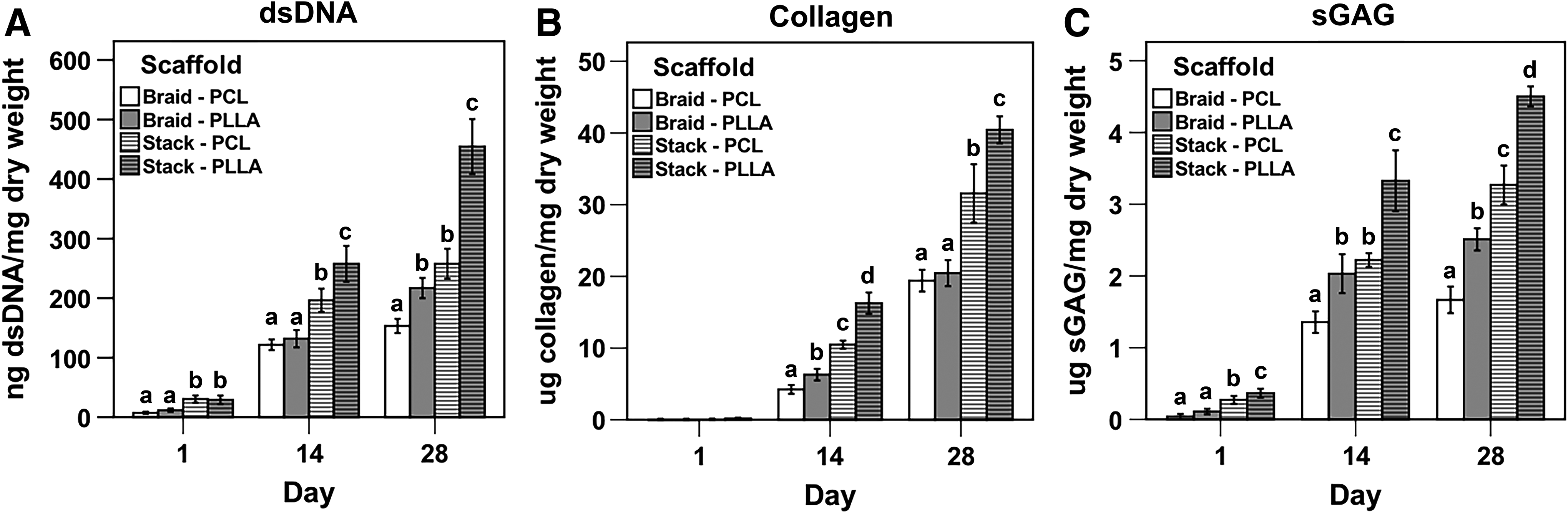
Biochemical composition of cell-seeded scaffolds over 28 days.
Discussion
This study compared two designs of aligned nanofibrous scaffolds–stacked versus braided–in terms of mechanical properties and effects on MSC behavior. In support of our hypothesis, braiding increased both suture-retention and tensile strength, yet decreased cell infiltration and proliferation compared to stacked constructs. Similarly, while stacked constructs permitted greater total collagen and sGAG deposition, this ostensible anabolic effect was lost when normalized against cell number (i.e., dsDNA content). The scaffold design had a greater effect on tenogenic differentiation of seeded MSCs, as measured by gene expression, than did polymer composition, with braided constructs upregulating tenogenesis to a greater degree than stacked constructs. Both stacked5,25–27 and braided18,20,21,28,29 scaffolds have been developed as engineered tendon or ligament grafts, yet a direct comparison has not been previously performed. While braided microfibrous scaffolds (i.e., fiber diameter 10–200 μm) have shown good cytocompatibility and mechanical properties both in vitro18,28 and in vivo, 19 how the bioactivity of these structures differs from structural proteins of the ECM (e.g., collagen fibrils, glycoproteins, etc.), which possess fiber diameters on the nanoscale, is not known. The fabrication technique of electrospinning can model these biophysical cues of the native ECM to permit mechanistic investigation of cell–matrix interactions and serve as biodegradable scaffolding for tissue-engineered constructs. Several groups have found that nanofiber orientation can control cell morphology, with aligned fibers upregulating tendon-related gene expression.7,8,30 Fiber diameter also affects cell behavior, as small diameter fibers (e.g., 320 nm) appear to preferentially promote proliferation and matrix deposition, while larger fibers (e.g., 1.80 μm) promote a stronger tenocytic phenotype.6,31 Interestingly, combining nanoscale cues resembling collagen fibrils with macroscale cues mimicking collagen fibers/fascicles of native tendon/ligament has been shown to promote synergistic effects in terms of tenogenic differentiation.23,32,33 This study examined whether higher order patterns–braiding versus stacking–affected tenogenic differentiation.
TGF-β3 and ascorbic acid were added as supplements to the culture medium to facilitate tenogenic differentiation of seeded MSCs. TGF-β3 is a pleotropic growth factor previously shown to be essential for tenogenic differentiation in both in vivo development34,35 and in vitro experiments,36–38 although other growth factor cocktails have also been shown to enhance in vitro tenogenic differentiation.39,40 At present, a standard tenogenic culture medium has not been established. That said, the differentiation medium upregulated tenogenic transcription factors Scx and Mkx in 2D controls on days 3 and 7 before returning to baseline at day 14. Upregulation of Scx was most strongly enhanced on braided scaffolds of both polymers, while stacked constructs showed early upregulation on day 3 before returning to baseline. This pattern is consistent with the in vivo role of Scleraxis, 41 which is the earliest marker of tenogenesis and drives subsequent expression of mature tenocytic markers tenomodulin 42 and collagen type I. 43 Despite the changes in Scx (and to a lesser extent, Mkx) expression, collagens type I and type III expression showed minimal changes in the braided and stacked constructs, with the former maintaining higher expression over the culture period. Other studies examining the effect of multilayered5,25 and braided20,21 nanofibrous scaffolds on tenogenic differentiation also reported modest changes in collagen expression. Conversely, discernible tenomodulin expression was only found on day 14 in scaffold conditions, similar to results reported in similar studies.21,25,44 The master osteogenic regulator, Runx2, was modestly upregulated in scaffold constructs (approximately two to threefold) early, before declining on day 14. Barber et al. 20 described comparable findings using similarly braided nanofibrous scaffolds.
In contrast to our hypothesis, stacked scaffolds exhibited decreased expression of tenogenic transcription factors and ECM proteins compared to braided scaffolds. This observation could be caused by several possible scenarios. Cells of the stacked scaffolds were more homogeneously distributed and exhibited an elongated morphology in the direction of aligned nanofibers, whereas cells seeded on braided scaffolds were overwhelmingly localized to scaffold surfaces and were more heterogeneous in their distribution. Cell morphology is an established mediator of gene expression; the differences in cell morphology seen between scaffold groups may have contributed to the noted differences in gene expression. In addition, gap junctions form a three-dimensional communication network among tenocytes in vivo.45,46 Scaffold-mediated differences in cell–cell communication could produce differences in gene expression. Finally, the greater cell number found on stacked scaffolds could have contributed to a lower relative effective dose (per cell) of the tenogenic supplements TGF-β3 and ascorbic acid. Additional factors likely contributed as well, but the mechanistic pathways of the scaffold-mediated differences are beyond the scope of the present study.
On a protein level, total collagen and sGAG deposition increased over time, as shown histologically and biochemically. Stacked scaffolds supported greater protein content, but this effect was due to proportional increases in cell number. As expected, PLLA scaffolds, presumably due to their greater hydrophilicity, supported greater cell numbers than their PCL counterparts, as also reported by Czaplewski et al. 21 On the other hand, cell distribution was more homogeneous in stacked scaffolds than braided scaffolds. The braiding process requires a modest degree of tension exerted on each braid, likely reducing the pore size by moving aligned fibers closer together. Bosworth et al. 47 similarly noted that MSCs were limited to the perimeter of dense electrospun yarns. Several methods have been described to increase pore size and cell infiltration into aligned nanofibrous constructs, but often at the expense of fiber architecture and scaffold strength. 10
Mechanical properties of the scaffold are of particular concern if the scaffold is to be used as an engineered graft or biomaterial providing mechanical support to a primary repair. In either case, the scaffold should have mechanical properties comparable to the native tissue and possess suture-retention strength of equal or greater strength than the repair alone.15,48 In this study, PCL constructs possessed a higher tensile strength (i.e., ultimate stress) than their PLLA counterparts, although failure occurred well beyond the range of physiological strain in tendons/ligaments (<12%). 49 Braiding had no effect on the ultimate stress of PLLA constructs, but produced superior values for PCL constructs. For both polymers, braiding reduced the modulus, as individual fibers were necessarily oriented away from the vector of tensile stress. Barber et al. 20 postulated that this offset of individual fibers away from the axis of tension accounted for the distinct toe region found in their braided PLLA scaffolds. Conversely, the braided PLLA scaffolds studied in this study did not exhibit the anticipated toe region. Such a discrepancy may be attributable to differences in the scaffold fabrication process or the increased strain range used for preconditioning; this study cycled scaffolds from 0% to 2%, whereas Barber et al. 20 did not exceed 0.5% strain as an upper limit for preconditioning. Nevertheless, the material properties found herein are similar to reported values for aligned PCL or PLLA nanofibrous scaffolds.20,21,50
Within a range of physiological strain (Fig. 2B), the stacked PLLA scaffold exhibited the highest ultimate stress (5.76 ± 0.53 MPa) and modulus (118.47 ± 21.81 MPa), values that approach those of the middle and posterior regions of native rotator cuff, 51 but inferior to other ligaments and tendons. For instance, the native human patella tendon has an ultimate tensile strength of ∼60 MPa and modulus of ∼600 MPa. 52 Aligned nanofibrous scaffolds composed of poly-lactide-co-glycolide (PLGA) or poly-glycolic acid (PGA) possess higher strength and stiffness, but may degrade too quickly to provide sustained postoperative mechanical support.4,9 Conversely, polymers other than the conventional polyesters (i.e., PCL, PLLA, PLGA, and PGA) have recently shown promise as scaffolds for tendon/ligament tissue engineering, possessing superior material properties, but still supporting cell viability and tenogenic differentiation of seeded stem cells.17,53,54
Few studies on engineered scaffolds measure suture-retention strength, despite its established importance for clinical implementation. 48 In this study, braided scaffolds possessed superior suture-retention strength compared to stacked scaffolds. In the absence of a running horizontal suture to bind the three stacked sheets/braids, the suture- retention strength was only ∼3 N (data not shown), but increased to ∼16 N for stacked PCL scaffolds and ∼34 N braided PCL scaffolds. Stacked scaffolds failed when the suture sheared through individual fibers while braided scaffolds failed when the suture cut through an individual braid transverse to the direction of nanofiber alignment. While braiding increased suture-retention strength, the values were far inferior to maximum loads of surgical repairs. For instance, cadaveric models of rotator cuff repairs commonly fail at ∼350 N,48,55 with a single suture bearing up to 225 N. 56 Several commercially available scaffolds possess suture-retention strengths approaching these values, including dermis-derived grafts (∼100 N),57,58 poly(urethane urea) scaffolds (∼80 N), 57 and woven microfibrous PLLA scaffolds (∼400 N for three simple sutures pulled to failure). 59 We are not aware of investigations reporting the suture-retention strength of nonwoven electrospun nanofibrous scaffolds, as fabricated in this study.
Although the braiding pattern used in this study increased tensile and suture- retention strength compared to stacked multilayered scaffolds, the mechanical properties of all investigated scaffold types are inferior to those of native tendon or ligament. Furthermore, braiding reduced cell infiltration, which would likely limit the homogeneous deposition of new matrix throughout the scaffold if applied clinically. Given these limitations, the independent use of nonwoven electrospun nanofibers as a biomaterial for tendon or ligament replacement is not supported. In contrast, it may be possible to exploit the tenogenic induction promoted by aligned nanofibers if combined with more mechanically robust biomaterials. Hakimi et al. 22 recently developed a novel scaffold of woven microfibers with overlying electrospun nanofibers that upregulated tenogenic markers in seeded cells to a greater extent than the woven microfibrous scaffold alone. However, as demonstrated herein, it is important to analyze the effect of the macroscale architecture of novel scaffolds on cell behavior, as focal distortions in the otherwise consistent nanoscale cues can affect cell phenotype. Moving forward, in vitro methods to predict in vivo responses to biomaterials should also be pursued,60,61 as substrate guided cell alignment and tenogenesis in vitro does not inevitably induce favorable in vivo healing. 62 Furthermore, validated criteria of successful in vivo tendon/ligament healing should be established so as to allow more direct comparison across different tissue-engineered constructs. 63
Conclusions
Aligned nanofibrous sheets of PCL or PLLA were combined to form multilayered braided or stacked scaffolds for tendon and ligament tissue engineering. Braiding can increase ultimate tensile and suture-retention strength, but concurrently reduces the modulus. While both scaffolds support expression of tenogenic markers, differences between scaffold designs confirm the importance of the macroarchitecture in modulating the bioactivity of nano/microarchitectural cues. Stacked scaffolds supported greater cell proliferation and migration than braided constructs, resulting in greater total collagen and sGAG deposition, but comparable relative deposition when normalized against dsDNA content. Design of future scaffolds may combine textile patterns, novel polymers, and region-specific biophysical cues to provide robust mechanical properties while promoting a tenogenic phenotype of seeded cells.
Footnotes
Acknowledgments
The authors thank Dr. Jian Tan for MSC isolation and Matthew Miller and Gerald Ferrer for their technical assistance in mechanical testing. This work was supported by the U.S. Department of Defense (W81XWH-13-2-0030). B.B.R. is a predoctoral trainee supported by an NIH Training Grant (5T32 EB001026).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.