
Abstract
In this study, we investigated the periodontal regenerative potential of dental follicle cell (DFC) sheets and periodontal ligament cell (PDLC) sheets in the simulating inflammatory microenvironment of periodontitis, to confirm their regenerative potential for clinical application and explain the possible mechanism. The biological characteristics of DFC sheets and PDLC sheets were explored in lipopolysaccharide from Porphyromonas gingivalis (P. gingivalis LPS)-induced inflammation microenvironment in vitro, then cell sheets were transplanted into canine periodontal defects with experimental periodontitis in situ for 3 months. The results showed that P. gingivalis LPS greatly impaired the differentiation of PDLC sheets, whereas promoted gene expression of bone sialoprotein (BSP), osteopontin (OPN), and periostin (POSTN) in DFC sheets. LPS activated toll-like receptor 4 and NF-κB p65 phosphorylation in PDLC sheets. In experimental periodontitis, new periodontal attachment could be obtained in both PDLC sheets and DFC sheets. However, the complete periodontal regeneration, including periodontal ligament–cementum complex structure was observed only in DFC sheet groups, which also showed more alveolar bone formation than PDLC sheets. These results suggest that DFC sheets were more effective for periodontal regeneration in chronic inflammatory microenvironment of periodontitis. It is probably because of their ability to adapt the inflammatory environment and strong capacity to promote periodontal regeneration. This approach provides a tangible pathway toward clinical translation.
Introduction
P
The ultimate goal of periodontal therapy is to achieve periodontal regeneration and restore the function of damaged periodontium. Stem cell therapy has become a promising approach. 6 However, it has always been a challenge to obtain the complete regeneration of periodontium, including cementum, alveolar bone, and functionally oriented periodontal ligament fibers inserting into them. 6 It has been reported that inflammatory microenvironment decreases the osteogenic potential of periodontal ligament cells (PDLCs), leading to impaired tissue regeneration. 7 Therefore, the key point of periodontal tissue regeneration lies in changing the inflammatory microenvironment and supplying a beneficial microenvironment for stem cells.
Cell sheet engineering ensures the delivery of robust cells connected through intact cell–cell interactions and their extracellular matrix (ECM) proteins, and has become the dominant strategy utilized in periodontal cell therapy.8,9 PDLC sheets have recently been assessed for periodontium formation in experimental models. 10 Besides PDLCs, dental follicle cells (DFCs) have attracted considerable attention. DFCs are a type of young mesenchymal stem cells (MSCs) found in developing periodontal tissues, which build up the periodontal structures, including cementum, PDL, and alveolar bone. 11 A study 7 showed that the function of PDLCs from a periodontitis patient could be damaged in the periodontitis microenvironment. Interestingly, DFCs enhance the self-renewal and multidifferentiation capacity of these PDLCs in vitro and Sharpey's fiber-like tissue formed without inflammatory cells when transplanting these PDLCs in vivo, indicates that DFCs could provide a beneficial microenvironment for periodontal regeneration. We previously reported that DFC sheet transplantation in vivo resulted in successful periodontal regeneration. 12 These findings suggest that DFC sheets may be an alternative and more easily an accessible source of multipotent stem cells for therapeutic and research applications compared with traditional cell sources.
Cell sheets provided a living microenvironment for the seeding cells, which increase the application efficiency. However, the regeneration ability of DFC and PDLC sheets in inflammatory microenvironment has not been explored in a large animal model. It is necessary to explore the biological characteristics of DFC and PDLC sheets in inflammatory microenvironment before their translation for therapeutic applications. Whether the inflammatory microenvironment can modulate the fate of cell sheets, such as differentiation and protein expression is unknown.
In this study we investigated the biological characteristics of DFC sheets and PDLC sheets in lipopolysaccharide from Porphyromonas gingivalis (P. gingivalis LPS)-induced inflammation microenvironment in vitro, and then access the periodontal regeneration ability of DFC sheets and PDLC sheets in canine periodontal defects with experimental periodontitis, to evaluate the potential usage of DFC sheets and PDLC sheets for periodontal regeneration in inflammation microenvironment.
Materials and Methods
The ethics committee of Sichuan University approved all human cell culture and animal experiments. Informed consents from patients who provided teeth for subsequent research were obtained.
Cell and cell sheet culture
Human DFCs were obtained from mandibular embedded wisdom teeth, and PDLCs were isolated from premolars extracted for orthodontic treatment. Canine DFCs were from unerupted dental germ of first mandibular molar, and canine PDLCs were from premolars. Primary cell culture protocols had been described in detail. 12
Human cell sheets were cultured for 4 weeks in 10% fetal bovine serum (FBS) α-MEM culture medium containing 50 μg/mL ascorbic acid 2-phosphate (AA, Sigma). Canine cell sheets were cultured for 2 weeks.
Immunofluorescence microscopy
Cells and cell sheets were fixed, permeabilized, and blocked. Antibody staining was performed according to the manufacturer's recommended protocol, including anti-Toll-like receptor 2 (TLR2, ab24192, 1:100; Abcam, United Kingdom), and anti-TLR4 (ab22048, 1:100; Abcam, United Kingdom). All samples were examined under the fluorescence microscope (Olympus, IX73P2F, Japan).
LPS induced inflammatory microenvironment
P. gingivalis LPS (Invitrogen, USA) were diluted by 10% FBS α-MEM into gradient concentrations (10, 100, 250, 500, 1000 ng/mL), then added to DFCs and PDLCs and cultured for 24 h. After cell viability and cytokine tests, the optimal concentration of LPS was chosen for the following experiments.
CCK-8 cell viability assay
Cell viability was investigated by the CCK-8 Kit (Dojindo, Japan). Assays were carried out according to the manufacturer's instruction. Human DFCs and PDLCs were seeded into 96-well plates (5 × 103 cells/well). After LPS induction for 24 h, 10 μL of CCK-8 solution was added to each well and incubated for 2 h. The absorbance was measured at 490 nm through a plate reader (Thermo, German). Each experiment was performed using three wells per experimental condition.
Quantitative real-time polymerase chain reaction
The total RNA of cells and cell sheets was extracted using RNAiso Plus (TaKaRa, Japan). First-strand cDNA was synthesized from 1 μg total RNA using a Reverse Transcriptase PCR Kit (Thermo, Germany). The real-time polymerase chain reaction system contained 50 ng reverse-transcribed total RNA, 400 nM primers, and SYBR Green Buffer (TaKaRa). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using Applied Biosystems Prism 7900HT Sequence Detection System (Thermo, German). The primers are listed in Table 1. Relative expression levels were calculated using the 2−ΔΔCT method and normalized by glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
Western blot
Cells or cell sheets were lysed in RIPA buffer (KeyGEN Biotech, China) with added 1 mM proteinase inhibitor phenylmethanesulfonyl fluoride (PMSF) and protein was extracted. The membranes were blotted with antibodies, which are listed in Table 2. After primary antibody incubation, the membranes were washed and incubated for 1 h in the peroxidase-conjugated anti-mouse or rabbit secondary antibody (ZSGB-BIO, China, 1:5000). The relative intensity of the tested protein was quantitatively analyzed by the ratio of the gray value between the target protein and GAPDH in the same sample.
TLR, toll-like receptor.
Canine experiment periodontitis model and in situ transplantation of cell sheets
Five 1-year-old healthy beagle dogs (10 kg, male) were used in this study. All surgical procedures were performed under general and local anesthesia. Canine periodontal two-wall intrabony defects (3 × 3 × 6 mm) were surgically created on the mesial side of mandibular third and fourth premolars bilaterally. The cementum was removed by Gracey curettes and the notch was made in the root extension of defect. Subsequently, a bacterial plaque retentive silk ligature (3–0) was placed in the gingival sulcus for 1 month to induce experimental periodontitis. Then dental plaque and calculus were removed by supragingival and subgingival debridement with hand instruments. The defects were filled with autologous DFC sheets or PDLC sheets. Nongrafted defects served as controls. Blood samples from a vein of the lower limb of dogs that were transplanted with cell sheets were obtained at the time of normal condition, pretransplantation surgery (1 month after experimental periodontitis induction), and posttransplantation (1, 2, 3, and 4 weeks after cell sheet transplantation) for turbidimetric inhibition immunoassay to evaluate serum complements (C3, C4), immunoglobins (IgA, IgG, IgM), and C-reactive protein.
Histological analysis
Cell sheets were fixed with 4% polyoxymethylene for 24 h, then dehydrated through a graded ethanol series, and embedded in paraffin. The paraffin blocks were sectioned into 5 μm thickness slides before histological staining.
Beagle dogs were sacrificed 3 months after surgery. The samples were scanned with a micro-CT system (ANALYSIS, Y. Cheetah, German). The overall topography of the defect sites with or without cell sheet transplantation was visualized in a three-dimensionally reconstructed image, which allowed the new bone height to be calculated, decalcified in 10% EDTA for 6 months, and embedded in paraffin. Histological specimens of 5 μm thickness were prepared. Periodontal defect healing was examined histologically by Hematoxylin and Eosin (H&E) staining, Masson's Trichrome staining and immunohistochemistry staining (COL I, ab90395, 1:200; Abcam, United Kingdom, and Periostin; sc-67233, 1:500, Santa cruz).
The PC-based image analysis system (Image-Pro Plus, Media Cybernetics, Inc., Silver Spring, MD) was used for histometric analysis. The following linear measurements were made at a magnification of × 40. (1) New bone height: distance from the base of the apical extension of root planing to the coronal extension of newly formed bone along the root surface; (2) New cementum height: distance from the base of the apical extension of root planing to the coronal extension of newly formed cementum or a cementum-like substance on the root surface; (3) Epithelial attachment: distance from cemento-enamel junction (CEJ) to the apical extension of an epithelial attachment on the root surface.
Quality control
All samples were done in triplicate and the results were confirmed by three independent experiments to duplicate the experimental conditions.
Statistical analysis
Data were expressed as mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was used to compare differences between groups using SPSS20.0 statistical software. Two-tailed t-tests were used to compare the two groups. Statistical significance was assumed for p-values less than 0.05.
Results
Inflammatory cytokine secretion and cell viability
LPS evoked inflammatory cytokine secretion of both cells after incubation for 24 h. However, the reaction to LPS of DFCs was more sensitive. For DFCs, IL-1β, IL-6, and TNF-α gene expressions increased to the maximum levels of 1.39-, 38.87-, and 2.48-folds, respectively, when the LPS concentration was 250 ng/mL; whereas the gene expression of IFN-γ upregulated to the top of 15.68-folds at 100 ng/mL LPS.
For PDLCs, IL-6 and TNF-α gene expressions increased to a maximum of 3.50- and 1.39-folds at 250 ng/mL LPS, and IL-1β gene expression reached the highest of 2.75-folds when stimulated with 500 ng/mL LPS. Like DFCs, the peak expression of IFN-γ for PDLCs was 2.00-folds at 100 ng/mL LPS (Fig. 1A).
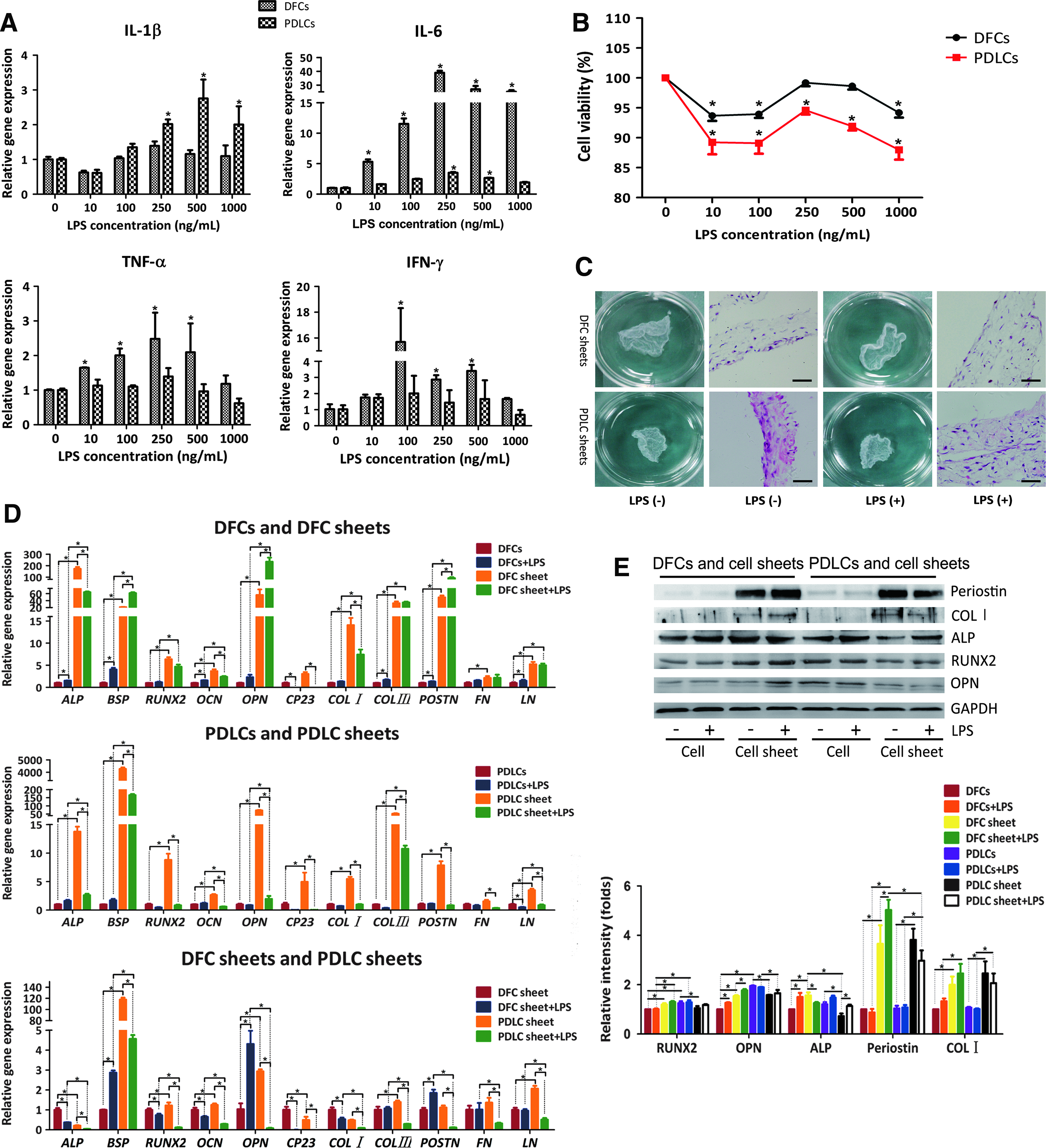
Lipopolysaccharide from Porphyromonas gingivalis (P. gingivalis LPS) induced inflammatory cytokine expressions and changes of cell viability, and the effect of P. gingivalis LPS on biological characteristics of cell sheets in vitro.
Cell viability of DFCs and PDLCs decreased independent of the LPS dosage after LPS incubation for 24 h. However, the influence of LPS on PDLCs was more obvious. When LPS concentration was 250 ng/mL, DFCs and PDLCs viability were relatively similar and close to the untreated group (Fig. 1B).
Overall, since 250 ng/mL LPS provoked remarkable inflammatory cytokine expressions and exerted less cytotoxicity effects on cells, it was chosen as a proper concentration to mimic the inflammatory microenvironment in vitro for the following experiments.
The effect of P. gingivalis LPS on biological characteristics of cell sheets in vitro
After culturing for 4 weeks, DFC sheets and PDLC sheets could be easily harvested as intact membrane from culture dishes by smooth mechanical stripping. H&E staining revealed that both DFC sheets and PDLC sheets were five to six cell layers and rich in ECM. LPS seemed to have no effect on the morphological structure of DFC sheets and PDLC sheets (Fig. 1C).
The osteogenic potential of PDLC sheets had been greatly impaired after LPS stimulation. In particular, the gene expressions of bone sialoprotein (BSP), RUNX2, OCN, and OPN significantly decreased (p < 0.05). Besides, the levels of cementum protein-23 (CP23), COL I, COL III, POSTN, LN, and FN of PDLC sheets were downregulated. In contrast, in DFC sheets, LPS repressed the gene expressions of ALP, RUNX2, OCN, CP23, and COL I (p < 0.05), whereas evoked expressions of BSP, OPN, and POSTN. Therefore, cell differentiation-associated genes and cell adhesion and migration-related genes of DFC sheets showed a clear advantage to PDLC sheets after being stimulated by LPS (Fig. 1D).
Protein expressions of DFC sheets and PDLC sheets were detected by western blot (Fig. 1E). The expressions of periostin and COL I were hardly detected in DFCs and PDLCs, but greatly increased in cell sheets. After being treated with LPS, they were upregulated in DFC sheets, whereas downregulated in PDLC sheets. The changes of osteogenic-related protein expressions were not so distinct. In DFC sheets, when stimulated with LPS, OPN increased, RUNX2 remained unchanged, whereas ALP decreased. In PDLC sheets, after inducing LPS, ALP expression was increased, whereas RUNX2 and OPN remained unchanged.
The influences of P. gingivalis LPS on TLRs signaling pathway of cell sheets
To demonstrate the role of TLRs in immunoregulation of cell sheets in inflammatory microenvironment, the protein expressions of TLRs and signal transduction molecules of cell sheets were evaluated. For immunostaining, TLR2 and TLR4 were positively expressed on the cell surface of DFCs and PDLCs, as well as cell surface in cell sheets (Fig. 2A).
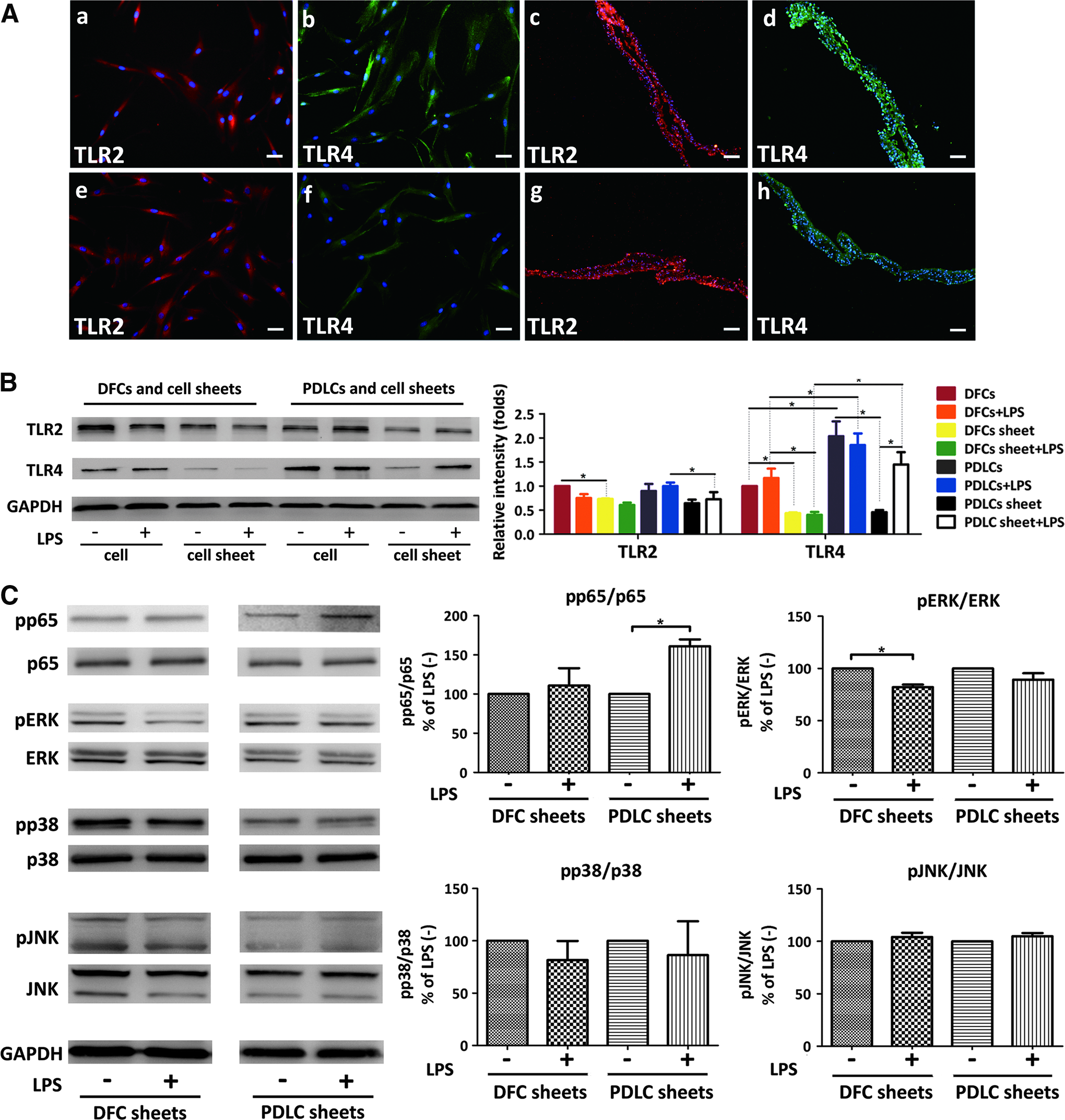
The influences of P. gingivalis LPS on toll-like receptor (TLR) signaling pathway of cell sheets.
The results of western blot showed that cell sheets expressed lower levels of TLR2 and TLR4 compared with cells, especially TLR4. After induction by LPS, TLR4 significantly increased in PDLC sheets. TLR2 decreased in DFC sheets when treated with LPS, whereas slightly raised in PDLC sheets (Fig. 2B).
It exhibited LPS-activated phosphorylation of p65 in PDLC sheets (p < 0.05), whereas phosphorylation of p65 also slightly increased in DFC sheets by LPS stimulation (p > 0.05). LPS significantly inhibited ERK phosphorylation in DFC sheets (p < 0.05). Phosphorylation of p38 seemed to decrease in both cell sheets (p > 0.05). LPS did not affect JNK phosphorylation in DFC sheets and PDLC sheets (p > 0.05) (Fig. 2C).
Periodontal regeneration of cell sheets in experimental periodontitis in situ
In canine experimental periodontitis, the marginal gingiva was bluish red and thickened after 1 month. Clinical attachment loss was found (3 mm) and the percentage of bleeding on probing reached to 54.2% (Fig. 3A). Canine DFC sheets and PDLC sheets were obtained after culturing for 2 weeks in cell sheet induction medium (Fig. 3B). Then the homologous cell sheets were transplanted into canine periodontal defects (Fig. 3C).
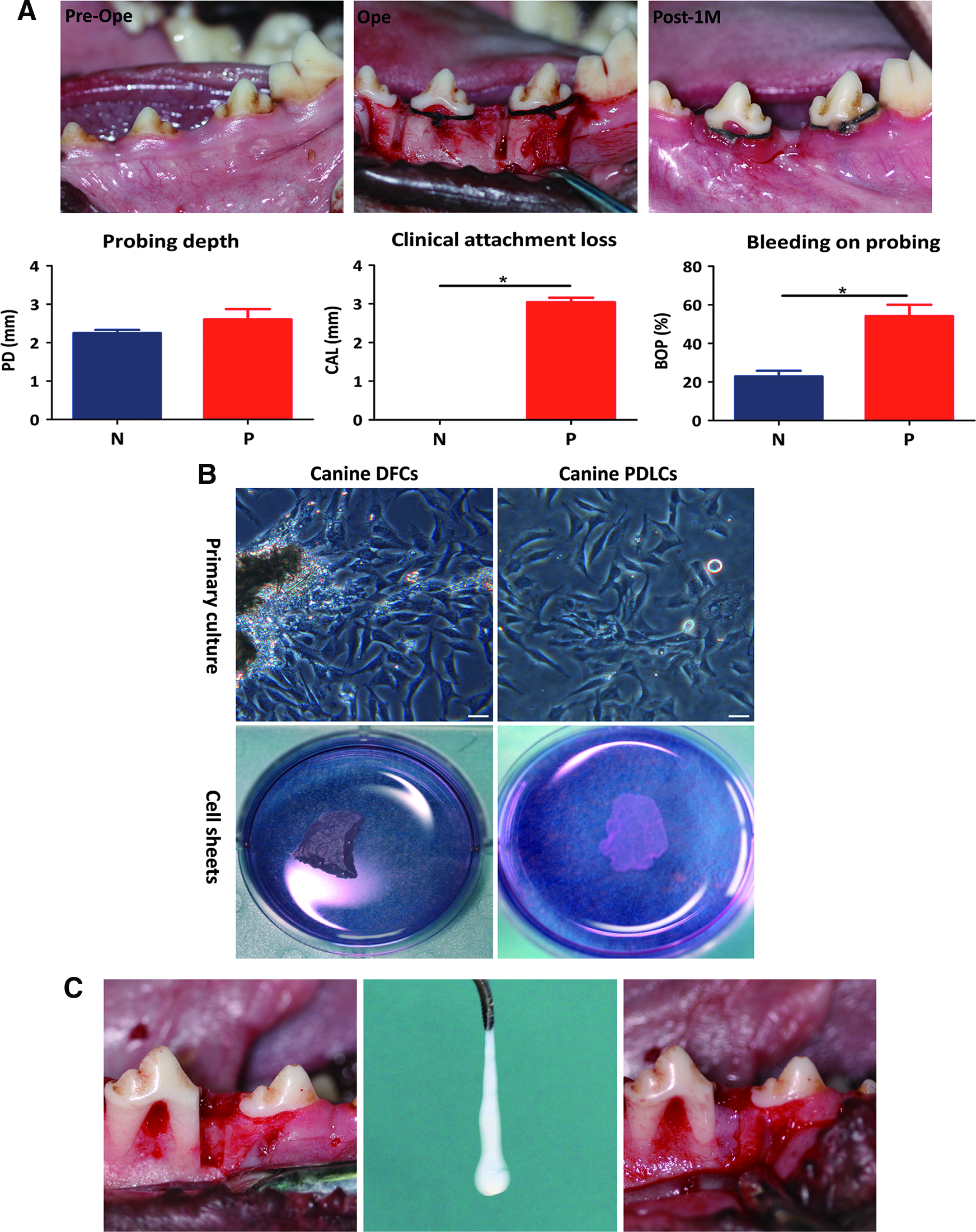
In situ transplantation of cell sheets in experimental periodontitis.
Micro-CT analysis illustrated that DFC sheet group had better newly formed alveolar bone than PDLC sheet group, which was close to the healthy periodontal tissue (Fig. 4A and Table 3). H&E staining sections confirmed the new alveolar bone formation in DFC and PDLC sheet groups. The denuded root surface was covered with a thin layer of cellular cementum and periodontal ligament aligned obliquely from new cementum to alveolar bone in DFC and PDLC sheet groups (Fig. 4B). Histometric analysis revealed that the new bone height was 4.67 ± 0.35 and 3.42 ± 0.26 mm in DFC and PDLC groups, respectively, whereas new cementum height was 5.16 ± 0.23 and 3.84 ± 0.30 mm. The epithelial attachment was 0.51 ± 0.06 mm in DFC and 0.62 ± 0.08 mm in the PDLC group, which did not differ significantly between the two groups (Table 3).
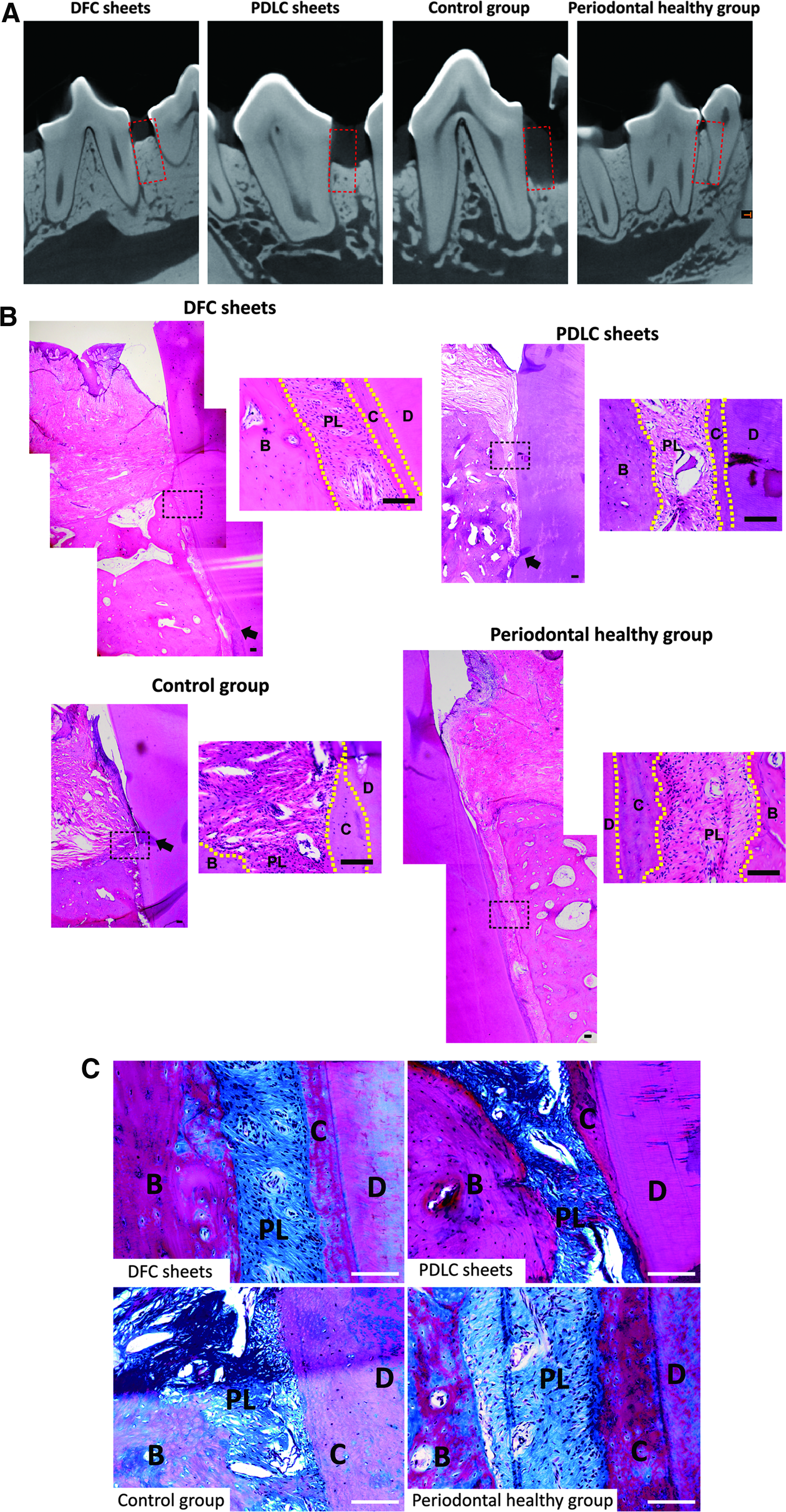
Representative micro-CT images and histological observation of regenerated tissue derived from cell sheets.
Statistically significant difference from control (p < 0.05).
Statistically significant difference from periodontal healthy group (p < 0.05).
Statistically significant difference from PDLC group (p < 0.05).
Statistically significant difference from DFC group (p < 0.05).
DFC, dental follicle cell; PDLC, periodontal ligament cells.
Masson staining exhibited much clearly the functionally oriented periodontal ligament fibers in both groups (Fig. 4C). The complete periodontal regeneration, including periodontal ligament–cementum/bone complex structure was observed in DFC sheet group, whereas partial periodontal regeneration with junctional epithelium extending apically was observed in PDLC sheet group. In the control groups, the alveolar crest was almost at the edge of the defect and junctional epithelium migrated apically (Fig. 4B, C).
Immunohistochemistry observed periostin expression in the regenerated periodontal ligament in both groups as that in periodontal healthy group. In control groups, periostin was weakly expressed in the periodontal ligament. COL I was detected in alveolar bone, periodontal ligament, cementum, and dentin in all the groups (Fig. 5A).
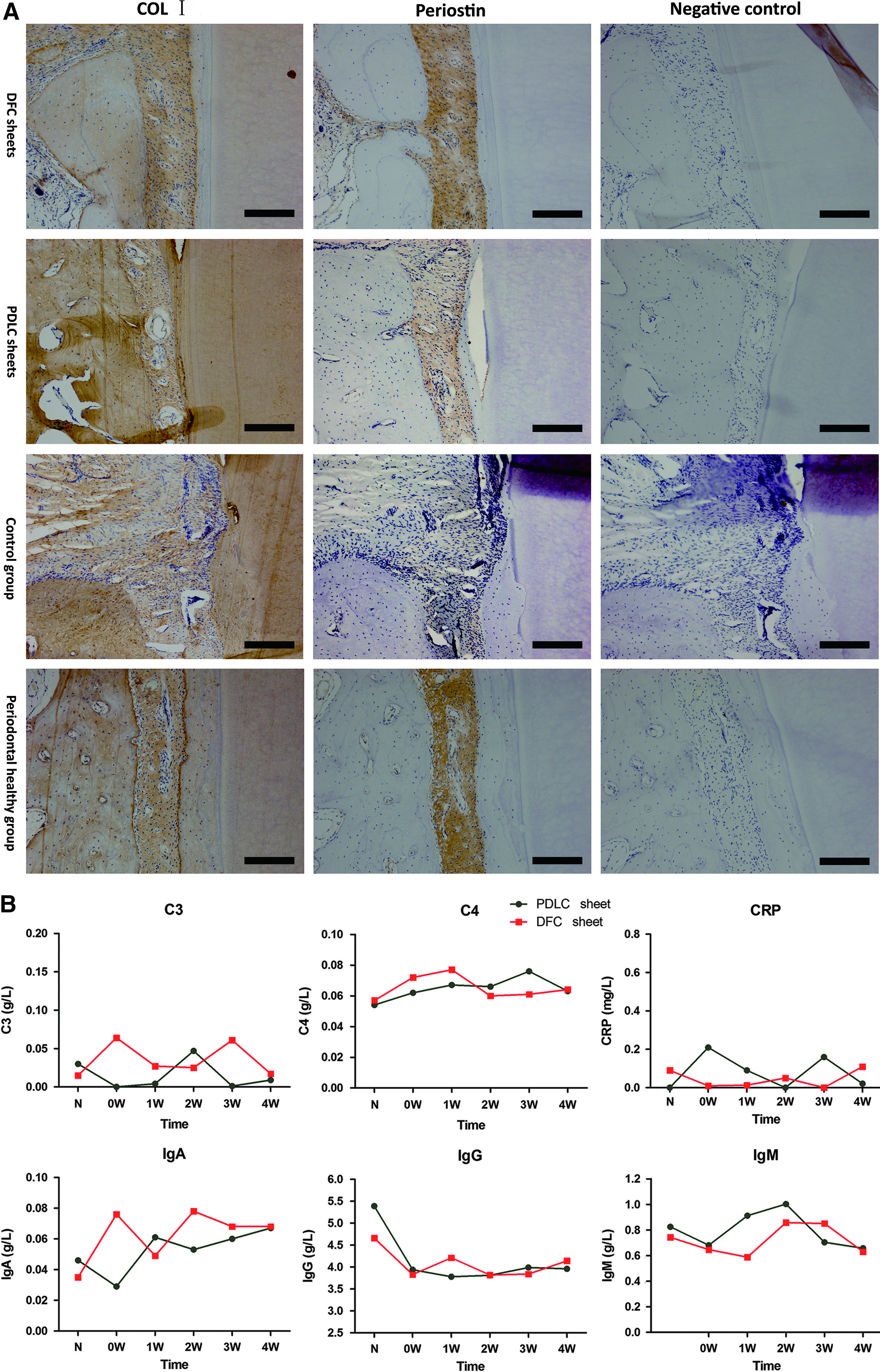
Immunohistochemistry staining for regenerated periodontal tissue and host immune response in experimental periodontitis.
Serum complements (C3, C4), immunoglobins (IgA, IgG, IgM), and CRP fluctuated slightly around normal levels in the following month after surgery. In addition, no lymphocytic infiltration occurred in the defects histologically 3 months after surgery (Fig. 5B).
Discussion
The results of this study indicated that DFC and PDLC sheets, although of the similar mesenchymal origin, have different characteristics under inflammation microenvironment. DFC sheets were more efficient for periodontal regeneration in chronic inflammatory microenvironment of periodontitis, in which the complete periodontal regeneration, including periodontal ligament–cementum complex structure was observed.
A key requirement for successful utilization of cell-sheet technology is the production of functional cell sheets with ECM protein to construct an appropriate extracellular microenvironment. Acting as a stimulator of proinflammatory response and bone resorption, LPS, the crucial virulence factor of P. gingivalis, was strongly involved in the initiation and development of periodontitis.13,14 H&E staining showed that after incubation with LPS, functional DFC and PDLC sheets were obtained possessing five to six cell layers and rich in ECM, indicating that LPS have little impact on morphological structure of DFC and PDLC sheets (Fig. 1C). However, LPS have evident influences on cell sheets in terms of multiple differentiation potential and matrix formation.
The differentiation of transplanted cells into the targeted cell types is vital for regeneration of functional tissues. In DFC sheets, after the stimulation of LPS, ALP, RUNX2, and OCN were downregulated. Interestingly, BSP, OPN, and POSTN were promoted. BSP and OPN are the essential components of the organic matrix of cemetum and bone. Periostin plays a key role in keeping the functional and structural integrity of the periodontium. It significantly decreased under chronic inflammation and correlated with alveolar bone loss over time.15,16 However, elevated gene and protein expressions of periostin in DFC sheets were observed after inducing LPS, suggesting that DFC sheets could encourage periodontal tissue repair through periostin, involving in angiogenic activity and osteoblast adhesion, differentiation, and survival.17,18
Col I is the chief element of periodontal ligament. 19 CP23 is a cementum-specific protein playing a key role in cementoblastic differentiation.20,21 Both of them decreased significantly in DFC sheets, which are harmful for periodontal regeneration. In contrast, gene expressions related to periodontal differentiation (namely osteoblastic differentiation, cementoblastic differentiation, and fibroblastic differentiation) of PDLC sheets were inhibited after inducing LPS without exception. PDLC sheets had advantageous osteogenic gene expressions over DFC sheets, but the superiority was abolished by P. gingivalis LPS. The simultaneous impaired osteogenic advantage of PDLC sheets and proinflammatory cytokine expressions stimulated by P. gingivalis LPS might challenge the clinical periodontal regeneration.
P. gingivalis LPS is mainly recognized by host cells via membrane receptors-TLR2/TLR4.22,23 Our results showed that P. gingivalis LPS increased TLR4 protein expression in PDLC sheets and led to phosphorylation of NF-κB p65. NF-κB activation negatively affects osteoblast function and bone formation by promoting β-catenin ubiquitination and degradation. 24 Blocking TLR4 or NF-κB signaling partially reversed the impaired osteogenic potential of PDLCs and were resistant to alveolar bone loss following LPS-induced experimental periodontitis in rats. 25 Therefore, TLR4 took part in LPS-induced NF-κB pathway activation, which decreased the osteogenic potential of human PDLCs. In this study, NF-κB p65 phosphorylation of DFC sheets remained unchanged after P. gingivalis LPS incubation, which relieved the DFC sheets from negative pluripotent regulation. Besides, P. gingivalis LPS inhibited ERK phosphorylation in DFC sheets. Previous studies have shown that ERK phosphorylation leads to the inhibition of osteogenisis.26,27 However, some studies demonstrated that activation of ERK signaling pathway participates in the process of MSCs differentiating into osteoblasts.28,29 Further study is needed to clarify the roles of TLRs and their signaling pathways in function modulation of cells and cell sheets.
LN and FN are the major component of ECM, which helps in directing the migration, growth, differentiation, and synthetic activity of both inflammatory cells and structural cells in injured tissue.30,31 FN has already been applied in the clinical treatment of periodontal defects to promote the new attachment formation. In our study, LPS significantly inhibited the expression of LN and FN genes in PDLC sheets, but had little effect on those in DFC sheets, which suggested that DFCs migrating from cell sheets could easilly attach to root surface and contribute to the new attachment formation.
In vitro results showed that DFC sheets have advantages to PDLC sheets on cell differentiation, inflammatory reactions, and ECM formation, which indicated that DFCs sheet is promising in periodontitis therapy. Thus, we verified this idea in beagle dog as an orthotropic transplantation animal model. In situ histological analysis revealed formation of new bone and cementum with embedded Sharpey's fibers in DFC and PDLC sheet groups in experimental periodontitis, but DFC sheets group gained more bone formation that was almost close to the healthy periodontal tissue.
The complete periodontal regeneration, including periodontal ligament–cementum–bone complex structure was observed in DFC sheet groups, whereas partial periodontal regeneration with junctional epithelium extending apically were observed in PDLC sheet groups. This was partly because of the comprehensive periodontal tissue differentiation potential of DFCs sheets in inflammatory environment, and partly because of a favorable stem cell niche for periodontal regeneration provided by embryonic characteristics of DFCs. It has been proven that coculturing with DFCs appeared to enhance the self-renewal and multidifferentiation capacity of PDLCs derived from inflamed periodontium (7). It implied that cells from DFC sheets not only devoted themselves to tissue repair, but also improved the potential of residing PDLCs.
The systemic response to allogeneic DFC or PDLC sheets in periodontal defects was examined at the level of circulating complements and antibodies. Both DFC and PDLC sheets did not elicit strong host immune responses. The histological results demonstrated effectual periodontal regeneration without significant host inflammatory response. Lack of MHC class II antigen and costimulatory molecules expression on DFCs and PDLCs hinted less immunogenicity like bone marrow MSCs. 32 DFCs and PDLCs also possess immunosuppressive properties to inhibit peripheral blood mononuclear cell proliferation.33–35 PDLCs were shown to induce CD4+ CD25+ Foxp3+ regulatory T cells and suppress Th17 differentiation, 35 which then promote regeneration over inflammation. But considering the outstanding periodontal regeneration of DFC sheets over PDLC sheets in inflammatory environment, DFCs offer the possibility to simultaneously target the inflammatory response and to promote the regeneration of periodontal structure, which would enable DFCs a better cell source for periodontal tissue engineering.
Collectively, this study revealed that DFC and PDLC sheets are promising approaches for periodontal regeneration under inflammation microenvironment. DFC sheets were more effective for periodontal regeneration in chronic inflammatory microenvironment of periodontitis. It is probably because of their ability to adapt to the inflammatory environment and strong regenerative capacity. The successful approach applied in this study might accelerate the clinical translation.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (Grant No. 81200792, Beijing, China), and Sichuan Province Science and Technology Support Program (Grant No. 2013FZ0033, Chengdu, China).
Disclosure Statement
No competing financial interests exist.