
Abstract
Embryonic stem cells (ESCs) are an ideal source for chondrogenic progenitors for the repair of damaged cartilage tissue. It is currently difficult to induce uniform and scalable ESC differentiation in vitro, a process required for stem cell therapy. This is partly because stem cell fate is determined by complex interactions with the native microenvironment and mechanical properties of the extracellular matrix. Mechanical signaling is considered to be one of the major factors regulating the proliferation and differentiation of chondrogenic cells both in vitro and in vivo. We used biocompatible and elastic polydimethylsiloxane (PDMS) scaffolds, capable of transducing mechanical signals, including compressive stress in vitro. ESCs seeded into the PDMS scaffolds and subjected to mechanical loading resulted in induction of differentiation. Differentiated ESC derivatives in three-dimensional (3-D) PDMS scaffolds exhibited elongated single cell rather than round clonal ESC morphology. They expressed chondrogenic marker, Col2, with concomitant reduction in the expression of pluripotent marker, Oct4. Immunocytochemical analysis also showed that the expression of COL2 protein was significantly higher in ESCs in 3-D scaffolds subjected to compressive stress. Further analysis showed that compressive stress also resulted in expression of early chondrogenic makers, Sox9 and Acan, but not hypertrophic chondrogenic markers, Runx2, Col10, and Mmp13. Compressive stress induced differentiation caused a reduction in the expression of β-Catenin and an increase in the expression of genes, Rhoa, Yap, and Taz, which are known to be affected by mechanosignaling. The chondroinductive role of RhoA was confirmed by its downregulation with simultaneous decrease in the transcriptional and translational expression of early chondrogenic markers, SOX9, COL2, and ACAN, when ESCs in PDMS scaffolds were subjected to compressive stress and treated with RhoA inhibitor, CCG-1432. Based on these observations, a model for compression induced chondrogenic differentiation of ESCs in 3-D scaffolds was proposed.
Introduction
C
Nonoperative treatments can manage pain symptoms by nonsteroidal anti-inflammatory medications and intra-articular injections of corticosteroids or hyaluronic acid, but do little to halt the degeneration process. 5 Similarly, surgical procedures, including debridement, microfracture, autograft cartilage transplantation, or full joint replacement, often result in fibrocartilage formation, lacking in the mechanical properties of the native cartilage. 6 Use of cartilage autografts is also difficult to practice, due to limited tissue availability and donor site morbidity. 2 Autologous transplantation of chondrocytes has been used to reverse the symptoms and pathophysiology of osteoarthritis with limited success. 4 More recent approaches have been focused on using a combination of cell therapy and tissue engineering techniques for the regeneration of cartilage tissue. 7 Isolated chondrocytes have limited proliferative abilities and have been shown to dedifferentiate during expansion in vitro.8,9
For the above reason, embryonic stem cells (ESCs), with their unlimited ability to self-renew and differentiate into various cell lineages, are an ideal cell source for tissue engineering applications, including cartilage repair and regeneration. 10 However, most cell therapies require a high quality population of differentiated derivatives to avoid adverse complications, including teratoma formation. In vivo, ESC fate toward self-renewal or differentiation is controlled by complex interactions of cell–cell and growth factor signaling, organization, and composition of the extracellular matrix (ECM), and the three-dimensional (3-D) biomechanical microenvironment.11–13 Mimicking the native microenvironment in vitro has been shown to improve the maintenance and propagation of pluripotent ESCs. 14 For instance, ESC self-renewal was maintained during extended culture in soft hydrogel scaffolds, exhibiting upregulation of Oct4 and Nanog expression. 15 Furthermore, varying substrate stiffness has been shown to affect cell fate, with elastic substrates favoring chondrogenic and stiff matrices resulting in osteogenic differentiation.16,17
Consequently, mechanical stimulation is considered to be one of the major epigenetic factors regulating the proliferation, survival, and differentiation of cells. 18 Chondrocytes and chondroprogenitors are capable of transducing and responding to various mechanical stimuli, including hydrostatic pressure, flow-induced shear stress, tension, and compression, altering chondrogenic gene expression and ECM deposition.13,19,20 However, the influence of compressive stress on ESCs is not well understood. In this study, we investigated ESC fate following compression using 3-D scaffolds composed of polydimethylsiloxane (PDMS), which are considered to be nontoxic and biocompatible.21,22 In this study, we report that ESCs encapsulated in 3-D PDMS scaffolds and subjected to compressive stress differentiated and expressed chondrogenic markers, Sox9, collagen type 2 (Col2), and aggrecan (Acan). Based on the results of our study we proposed a mechanism of mechanostimulation of ESC differentiation into chondrogenic lineage.
Materials and Methods
Maintenance and culturing of ESCs in 2-D culture plates
Enhanced yellow fluorescent protein (EYFP/GFP) labeled mouse ESCs (7AC5; ATCC) were grown in the culture medium (CM) under two-dimensional (2-D) conditions as previously described. 15 The CM contained high-glucose Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% fetal bovine serum (FBS; Aleken Biologicals) and 1000 U/mL of leukemia inhibitory factor (LIF; Chemicon International) in a 5% CO2 incubator at 37°C.
Fabrication of PDMS scaffolds
A microsphere-templating method developed in our previous studies23–25 was used to fabricate the PDMS scaffolds (Fig. 1). Briefly, white Jojoba wax microspheres (Natural Sourcing, LLC) with a diameter ranging from 425 to 500 μm were sintered at 66°C for 1 h to form a porous wax template. Well mixed two-component Ecoflex 30 PDMS resin (Smooth-On, Inc.) was then cast into the porous template. After thoroughly degassed in vacuum, the cast samples were cured at 40°C for 72 h. The wax was then extracted in a stirring acetone bath at 50°C for 48 h. Finally, the acetone in the PDMS scaffolds was vaporized off in a fume hood for 48 h at room temperature. The resultant PDMS scaffolds had a porosity of approximately 56% and with this method pore size can be altered using either smaller or larger Jojoba wax microspheres. 23

Illustration of a microsphere-templating process for scaffold fabrication.
Characterization of scaffolds
The morphology of sintered templates was observed by an Olympus BX51 microscope under the reflective mode. A Hitachi SEM S800 scanning electron microscope was used to capture the morphology of the elastomeric porous structure. Sliced samples were sputtered with gold for 45 s before SEM imaging. An Instron 5667 universal testing machine (Instron Corporation) with a load cell of 100 N was used to measure the tensile properties of porous elastomer. The samples were cut into a dumbbell shape and tested according to ASTM D412-C.
Seeding and culturing of ESCs in 3-D PDMS scaffolds
ESCs grown to 70% confluency in 12-well culture plates were trypsinized and counted using a hemocytometer. PDMS scaffolds were sterilized with 70% ethanol, rinsed with PBS, and incubated in ESC growth medium overnight. PDMS scaffolds were then seeded with ESCs (2 × 104 cells/mm2) by diffusion and cultured in CM in a 5% CO2 incubator at 37°C. ESC growth, distribution, and differentiation in PDMS scaffolds were monitored using confocal microscopy.
Cell proliferation assay
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) assay was performed to determine the proliferation of ESCs grown in 3-D PDMS scaffolds cultured for 14 and 25 days. Cells were treated with 5 mg/mL MTT reagent (Sigma) at 37°C for 4 h to obtain insoluble formazan, which was then solubilized using isopropanol/HCL (15:1). The absorbance of the solubilized formazan was measured at 570 nm using an Epoch microplate reader (BioTek), and the background absorbance of the cells was subtracted from all measured values.
Application of mechanical stress to the 3-D PDMS scaffold seeded ESCs
ESCs were seeded in the 3-D PDMS scaffolds as stated above and cultured for 7 days in CM before subjecting them to static compressive stress of 0.05 MPa. After 24 h static stress, the cells were subjected to molecular and immunocytochemical analysis.
Treatment with RhoA inhibitor
ESCs were grown in 3-D PDMS scaffolds for 7 days and then treated with 10 μM RhoA inhibitor (RhoAi), CCG-1423 (Tocris), in the presence of 0.05 MPa compressive stress for 24 h. ESCs grown in 3-D PDMS scaffolds were also similarly treated with RhoAi in the absence of compressive stress.
Expression of genetic markers
Gene expression analysis of 2-D and 3-D grown cells was performed using quantitative real time PCR (qRT-PCR). RNA was isolated from cells using the RNeasy Mini Kit (Qiagen) and purified by treating with RNase-free DNase (Promega). cDNA was synthesized with the iScript Kit (Bio-Rad). PCR reactions were performed in a 10 μL reaction volume using the Bio-Rad CFX90 RT-PCR system and SsoAdvanced SYBR Green Supermix. The specific qRT-PCR conditions used were as follows: polymerase activation 3 min at 95°C, 40 cycles of denaturation, 15 s at 95°C; annealing, 20 s at 60°C; and melt curve, 5 s/step at 60–95°C. The primers (IDT Technologies) used in this study represent pluripotent and cell lineage markers as listed in Table 1. All reactions were prepared in triplicate and normalized to reference genes, Gapdh and β-Actin.
Immunocytochemistry
Cells grown under 2-D and 3-D conditions were fixed in 4% paraformaldehyde for 10 and 30 min, respectively. Cells were then rinsed with PBS, and 3-D PDMS scaffolds were embedded in O.C.T. and frozen for cryosectioning for immunostaining. To treat the cells and cryosections with primary antibody, they were permeabilized with 0.5% Triton X-100 (Sigma) for 10 min and then blocked with 2% BSA (Sigma) for 1 h at room temperature. Fixed samples were treated with primary antibodies, SOX9, COL2A1, and ACAN (sc-17341, sc-7764, and sc-25674, respectively; Santa Cruz Biotechnology), diluted 1:100 in blocking buffer overnight at 4°C. Primary antibody treated samples were washed thrice with PBS, stained with APC or TR-labeled secondary IgG (sc-3860 and sc-2780, respectively; Santa Cruz), and counterstained with DAPI (1 mg/mL; 4′,6-diamidino-2-phenylindole dihydrochloride; Molecular Probes). The stained samples were visualized using confocal microscopy.
Statistical analysis
All quantitative data were expressed as mean ± standard error. One-way ANOVA analysis was performed and analyzed for unequal variances using post hoc tests for multiple comparisons. Results with a p-value less than 0.05 were considered to be significant (*p < 0.05 and **p < 0.01). All analyses were performed using SPSS version 11.5 (SPSS, Inc.).
Results
Fabrication and characterization of PDMS scaffolds
Characterization of fabricated PDMS scaffolds revealed sintering necks formed between the adjacent wax microspheres after the sintering process (Fig. 2A). The necks were the templates for the microchannels, which connect the neighboring pores in the PDMS scaffolds. Structure of the scaffolds was further confirmed by the SEM analysis. Figure 2B demonstrates a representative morphology of pores and microchannels that interconnect the adjacent pores. Microchannels have gradually enlarging outlets that join the adjacent round micropores enabling a cocontinuous porous structure. SEM images were taken at random locations, and it was found that most of the pores are fully interconnected with neighboring pores. Since the porous structure is a negative replicate of a fused wax bead pattern, the connectivity is expected to inherit the original wax pattern. The pore size ranges from 400 to 500 μm, reflecting the size of the wax templating microspheres.

Analysis of PDMS scaffolds.
A representative tensile stress–strain curve of porous PDMS scaffolds fabricated from Ecoflex 30 elastomer is depicted in Figure 3. The test was conducted in air using dry scaffolds. No anisotropic properties were observed as the pores are homogeneously distributed in the deformed scaffold. 23 The porous elastomer has low stiffness and high elasticity with a tensile modulus of 0.238 ± 0.034 MPa. The tensile stress and strain of the PDMS scaffolds at break are 0.568 ± 0.052 MPa and 6.13 ± 0.35 mm/mm, respectively. Moreover, according to our previous study, 23 porous scaffolds with micropores connected by the microchannels follow a very unique nonaffine deformation under external stresses. For this specific type of porous elastomer, the compressive modulus is comparable to the tensile modulus. 23 Therefore, tensile properties were used as representative mechanical properties for the scaffolds.
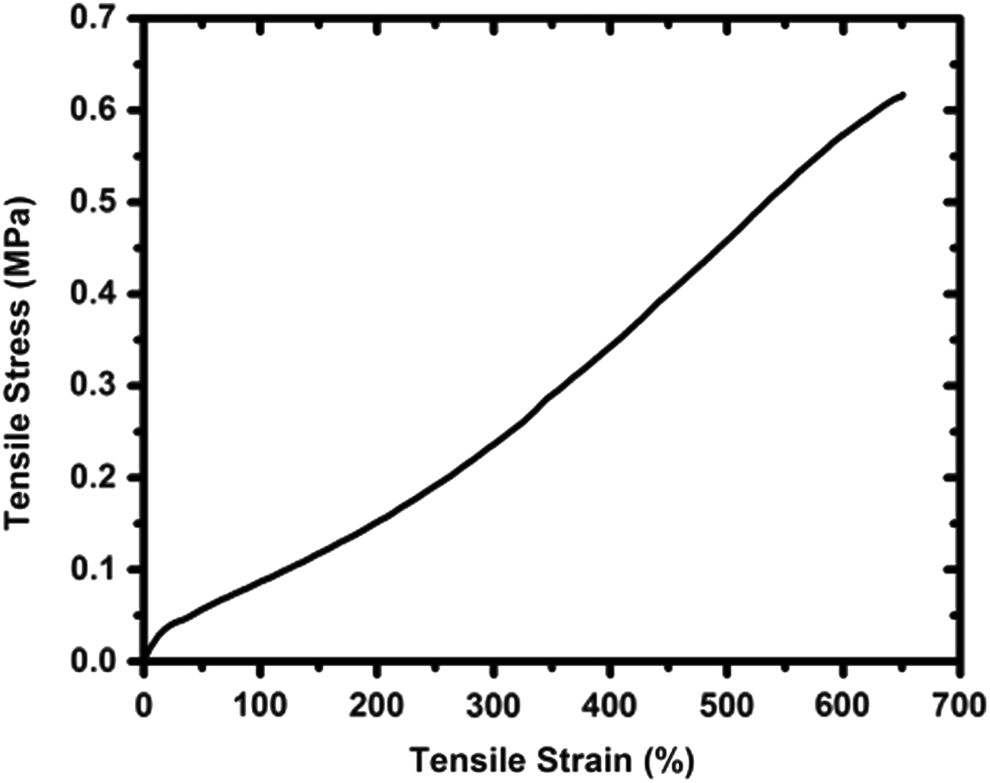
Representative stress–strain curve of porous PDMS fabricated using Ecoflex 30 elastomer.
Growth of ESCs seeded in 3-D PDMS scaffolds
Growth of EYFP/GFP labeled ESCs under 2-D and 3-D conditions is depicted in Figure 4. ESCs exhibited typical growth in 2-D culture, forming compact round colonies on day 2 cultures (Fig. 4B). However, the colonies flattened within 4 days of culture (Fig. 4C), exhibiting spontaneous differentiation. By day 7, cell colonies showed signs of extensive differentiation, reaching confluence in 2-D culture conditions (Fig. 4D). Unlike the round compact morphology of ESCs, differentiated cells displayed enlarged, flattened, or fibroblast-like morphology in 2-D culture when grown for 4 and 7 days. Whereas ESCs seeded into 3-D PDMS scaffolds showed almost uniform distribution within the scaffolds and compact colony morphology as shown by z-stack confocal microscopy (Fig. 4E–H). Furthermore, composite image analysis revealed that although 3-D grown ESCs grew undifferentiated longer than 2-D grown ESCs, they also showed differentiation as evident from the fibroblastoid cell morphology after 14 days of culture (Fig. 4I–L). Whether the ESCs grown in PDMS scaffolds were viable and proliferated during a long incubation period was investigated using MTT assay. The results shown in Figure 4M demonstrated that the cells survived and proliferated during the 25 days of culture. Differentiation of ESCs was further investigated by determining the expression of lineage specific markers, myogenin (Myog), osteopontin (Opn), Nestin, and Col2, for myogenic, osteogenic, neurogenic, and chondrogenic cells, respectively, using qRT-PCR. Results depicted in Figure 4N, O show that pluripotent marker expression decreased upon culture, indicating ESC differentiation in both 2-D and 3-D culture, respectively.
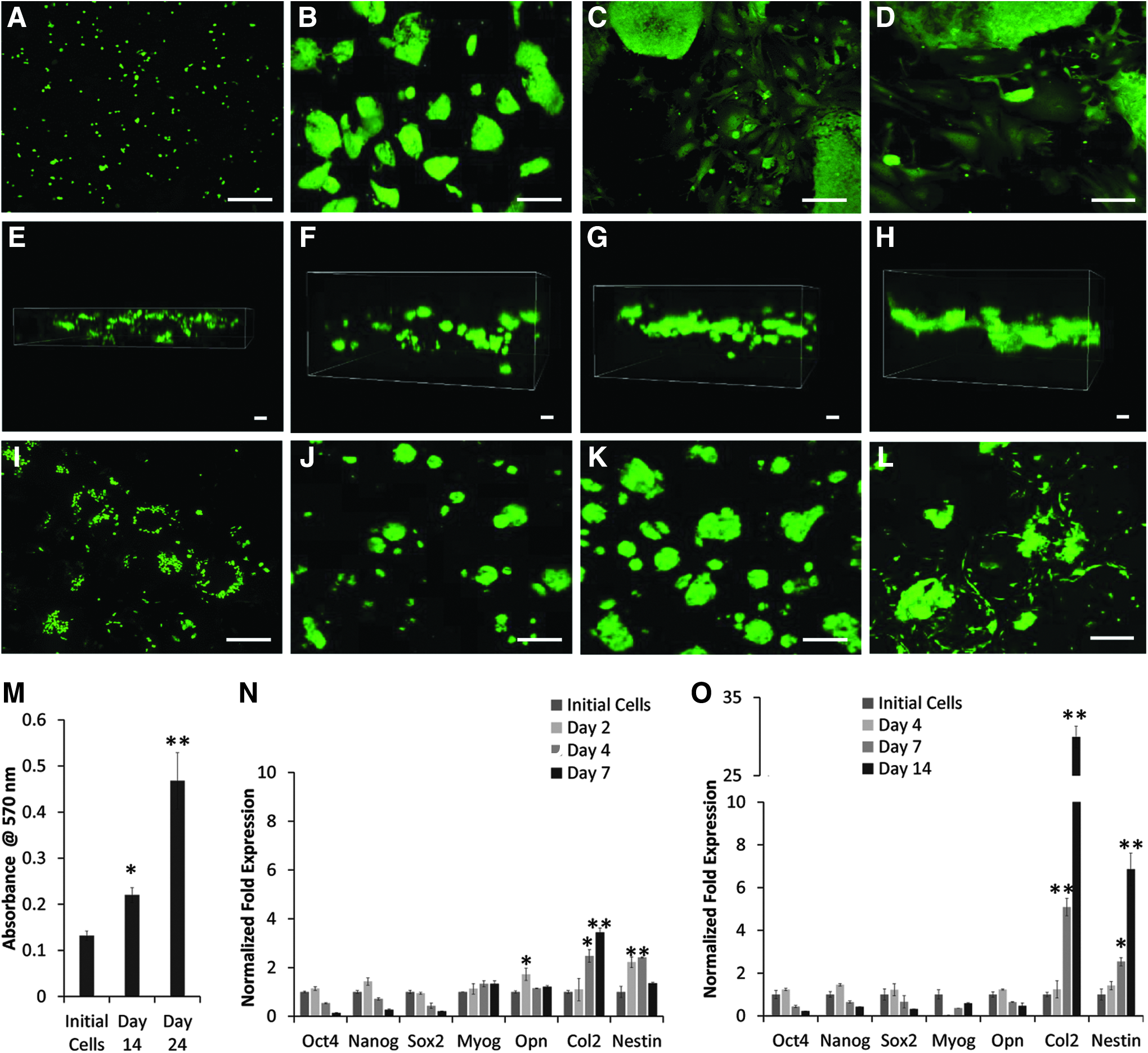
Comparison of EYFP labeled ESCs grown in 2-D culture and 3-D PDMS scaffolds. Growth of ESCs was visualized by confocal microscopy.
Two-dimensional culture leads to upregulation of lineage specific markers, Myog, Opn, Nestin, and Col2, signifying random ESC differentiation into numerous cell lineages. However, when ESCs were cultured in 3-D PDMS scaffolds, upregulation of predominately Col2 expression suggested selective differentiation into chondrogenic lineage.
Effect of compression on ESCs seeded in PDMS scaffolds
Since mechanical stresses are known to play a role in cell fate, including processes of self-renewal and differentiation, we wondered if PDMS, due to its high elasticity, would be able to transduce compressive signals to ESCs cultured in the 3-D scaffolds. In our preliminary experiments, we evaluated the effect of various levels of compressive stress ranging from 0 to 0.1 MPa on ESCs grown in PDMS scaffolds. An optimal effect on chondrogenic differentiation was observed when ESCs were grown in 3-D PDMS scaffolds for 7 days, followed by the application of a 0.05 MPa compressive stress for 24 h.
To decouple the effect of 3-D culture in PDMS scaffolds on ESC differentiation from induction due to compressive stress, gene and protein analysis was compared in ESCs cultured in PDMS scaffolds in the presence and absence of compression. Analysis of the ESCs grown under these conditions showed reduced expression of master pluripotent marker, Oct4 (Fig. 5A), suggesting that cells were undergoing differentiation. When various lineage-specific markers were probed, Myog and Opn markers were downregulated, but Nestin was slightly upregulated (1.3-fold) as a result of 0.05 MPa compressive stress applied for 24 h to ESCs grown in PDMS for 7 days. However, more striking results were the 13-fold versus 4-fold increase in the expression of chondrogenic marker, Col2, in 3-D PDMS scaffolds with and without compression, respectively. Immunocytochemical analysis depicted in Figure 5B and quantified in Figure 5C also showed that ESCs grown in PDMS scaffolds had significantly higher expression of COL2 protein as a result of compression. Further transcriptional analysis showed that compression induced expression of early chondrogenic markers, Sox9 and Acan, but not hypertrophic markers, including Runx2, collagen type X (Col10), and matrix metallopeptidase 13 (Mmp13) (Fig. 5D). Overall, culture in PDMS scaffolds alone had only a slight chondroinductive effect on ESC differentiation, which was upregulated by application of compressive stress resulting in early chondrogenic differentiation of ESCs.
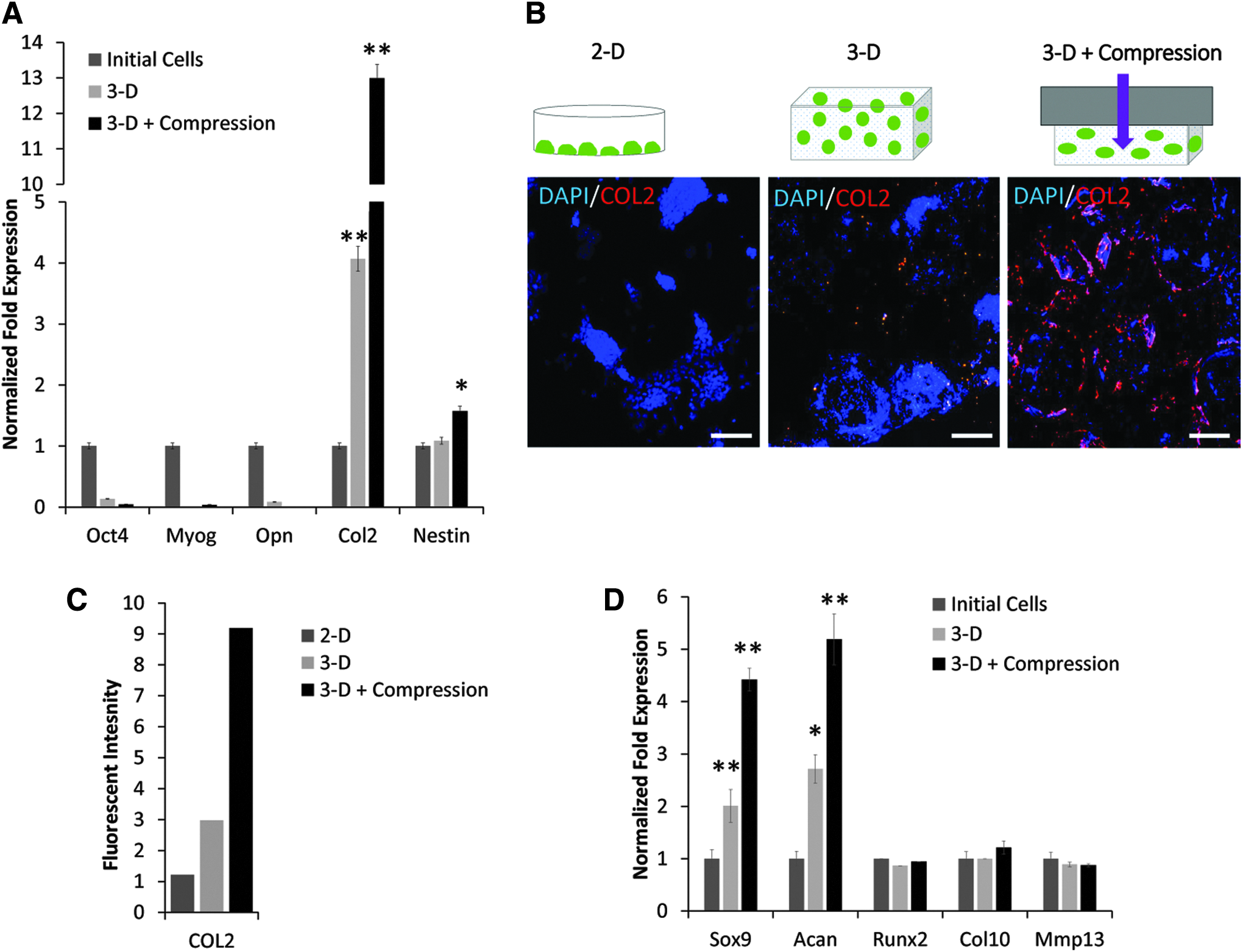
Effect of compressive stress on differentiation of ESCs cultured in 3-D PDMS scaffolds.
Mechanotransduction of compressive signals
Mechanotransduction of biophysical signals from the microenvironment can regulate cell shape and fate. 26 A role in mechanical signaling has recently been reported in numerous pathways, including RhoA signaling, responsible for actinomyosin contractility, 27 and Yes-associated protein (YAP)/PDZ-binding motif (TAZ) signaling. 28 We also investigated the mechanism of selective chondrogenic differentiation by compression. The results reported in Figure 6A indicated that compressive stress, while upregulating the chondrogenic markers, Sox9 and Col2, resulted in the suppression of β-Catenin, but had no effect on Tcf, a downstream effector of the Wnt pathway or Rac1 expression involved in RhoA signaling. Interestingly, downstream effectors of RhoA pathway, including Rhoa, Yap, and Taz, were upregulated (2.7-fold, 3.1-fold, and 1.5-fold, respectively) in cells subjected to compressive stress.
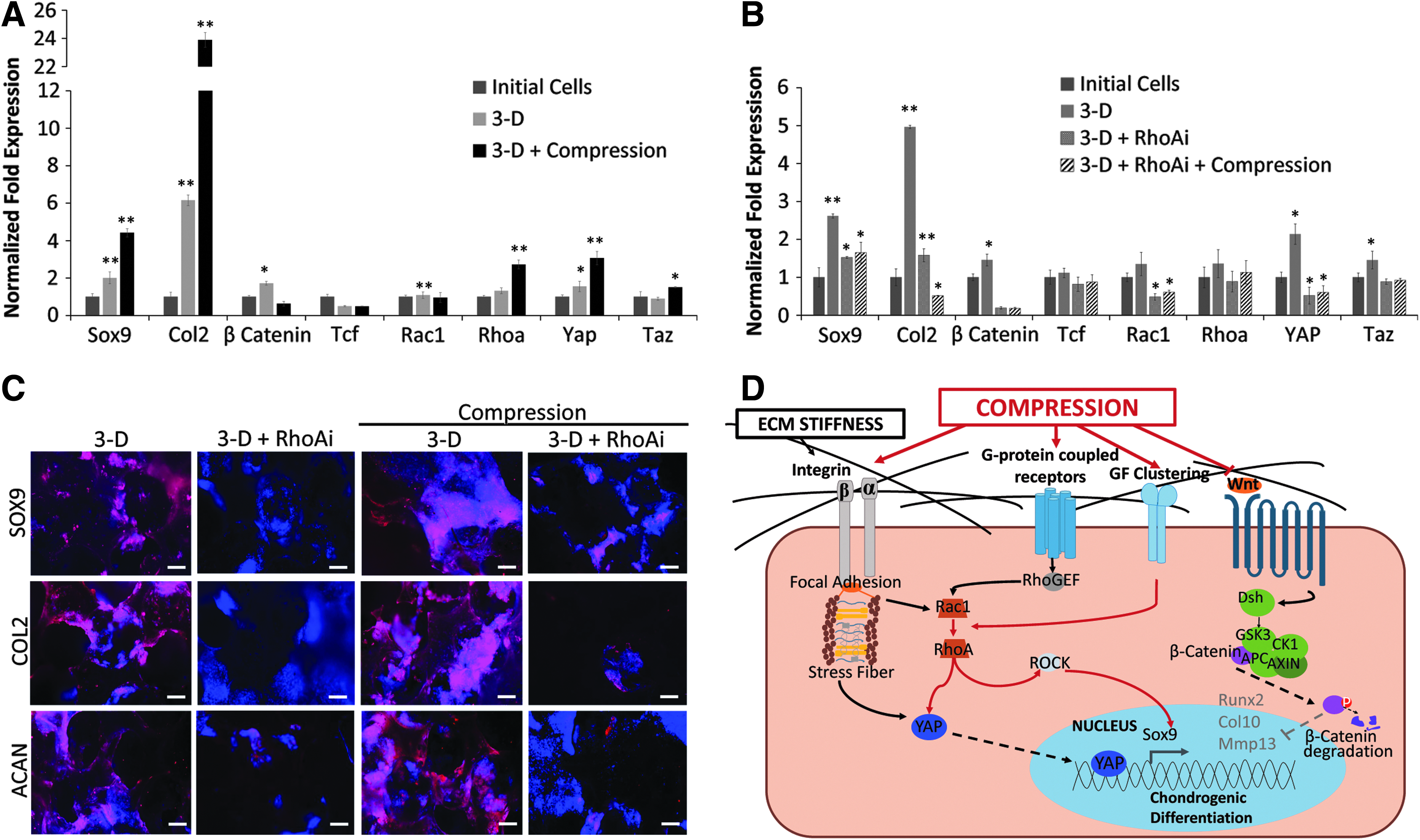
Compression induced chondrogenic differentiation of ESCs through RhoA and YAP signaling pathway.
ESCs were grown in PDMS scaffolds for 7 days, then treated with RhoAi, and subjected to compressive stress for 24 h to elucidate the role of RhoA signaling in chondrogenic differentiation. These results depicted in Figure 6B, C show that RhoAi abrogated the increase in Rhoa, Yap, and Taz expression. Not surprisingly, RhoAi treatment also resulted in the reduction in transcription of Sox9 and Col2, as well as the translation of SOX9, COL2, and ACAN, suggesting the inhibition of chondrogenic differentiation of ESCs in PDMS scaffolds even under compressive stress.
Based on these results, a mechanism of compression induced chondrogenic differentiation of ESCs in 3-D PDMS was proposed (Fig. 6D). Briefly, compressive stress downregulated Wnt and modulated RhoA signaling, which led to upregulation of downstream mechanosensitive transcription factors, YAP/TAZ, and induction of chondrogenic lineage differentiation.
Discussion
ESCs are a promising source to produce chondrogenic derivatives for the regeneration and repair of damaged cartilage. 13 Studies have shown that maintenance and differentiation of stem cells in vitro can be improved by mimicking the native microenvironment.15,29 This can be achieved by altering factors that influence cell fate determination. In fact, matrix elasticity and stiffness of substrates have been reported to play a role in directing differentiation of stem cells into specific lineages.20,30,31 These reports showed that softer, stiffer, and rigid matrices directed stem cells into neurogenic, myogenic, and osteogenic cell types. Despite the important role of mechanical signals in the cell fate determination, they have been poorly investigated.
In this study, we describe the effects of internal cell responses to matrix stiffness and external compressive stress on the growth and differentiation of ESCs. Although PDMS has commonly been used in microfluidic devices and long-term medical implants, different substrate formulations have been reported to hinder cell viability and adhesion,21,32 limiting the use of PDMS for macroculture of cells. Other reports have suggested that PDMS scaffolds promoted cell adhesion following incubation in FBS or concentrated proteins.21,33–35 Similar to these reports, our study also showed that PDMS scaffolds incubated in growth medium containing 10% FBS allowed successful ESC attachment, with an increase in colony size in a time dependent manner in 3-D culture. ESCs cultured in 3-D PDMS scaffolds exhibited compact colony morphology similar to early 2-D growth for several days; however, cell morphology changed to fibroblast-like type upon further incubation, suggesting that ESCs had undergone differentiation. PDMS scaffolds fabricated from Sylgard 527 and Sylgard 184 are more rigid than the scaffolds composed of Ecoflex 30 used in this study and have been shown to support differentiation of PC12 and C2C12 cell lines into neurons and myotubes. 36 Flexible biomaterials with a pore size ranging from 200 to 600 μm have been used for tissue engineering applications for myocardium, intestine, liver, and cartilage,37–40 whereas scaffolds with large pore sizes have been shown to promote chondrogenic differentiation and matrix deposition. 41 Since the 3-D PDMS scaffolds described in this study, composed of Ecoflex 30 with 400 to 500 μm pores, are highly flexible, we reasoned that they could be used to investigate the effects of matrix stiffness and compressive stress on cell fate determination.
ESCs grown in 2-D cultures have been shown to spontaneously differentiate into various cell lineages. 15 Long-term culturing of ESCs in 3-D PDMS scaffolds showed that cell growth was not limited by 3-D culture confinement conditions as the cell proliferation was progressively increased in a time dependent manner. However, 3-D culture of ESCs in PDMS scaffolds selectively induced differentiation into early chondrogenic lineage, expressing Sox9, Col2, and Acan, but not hypertrophic lineage markers. Compressive stress appeared to have a synergistic effect on the differentiation of ESCs. These results suggested that the low stiffness and high elasticity of 3-D PDMS scaffolds with the addition of compressive stress may have provided a more conductive microenvironment for chondrogenic differentiation. PDMS scaffolds and compression did not cause hypertrophic differentiation, as evident from the lack of expression of Runx2, Col10, and Mmp13. This is in line with previous studies that have shown that chondrocytes and chondroprogenitors are capable of transducing and responding to compressive stress.13,19,20 In a previous study, a compressive stress of 0.1 MPa applied for 10 min stimulated proteoglycan deposition of chondrocytes, but compression for 20 h suppressed ECM production. 42 In this study, ESCs grown in PDMS scaffolds subjected to a compressive stress of 0.05 MPa for 24 h resulted in induction of chondrogenic differentiation. Taken together, these findings suggest that both substrate elasticity and mechanical compressive stresses play a role in induction of chondrogenic differentiation in ESCs grown in 3-D PDMS scaffolds.
Elucidation of the mechanosensing response of cells to their microenvironment due to matrix substrate elasticity and compressive stresses is important to improve in vitro differentiation of ESCs into a more homogenous population of derivatives. Although mechanotransduction through changes in the actin cytoskeleton is the most studied of the pathways, YAP/TAZ signaling is also important. YAP and TAZ are ideally positioned at the center of biomechanical, Hippo, TGFβ/BMP, and Wnt signaling pathways, 43 suggesting that YAP may play a role in the induction of chondrogenic differentiation through matrix elasticity and is further upregulated upon compression.
Mechanotransduction using the Rho signaling pathway has been thoroughly studied in adult stem cell differentiation; RhoA is activated by growth on rigid substrates, leading to induced cell spreading, cell contractility, BMP-dependent signaling, and osteogenic differentiation.26,44 In ESCs, Rho signaling plays an important role in the maintenance of pluripotency, viability, and fate determination. 26 In articular chondrocytes, dynamic compression and TGFβ activated RhoA/ROCK signaling resulted in a ROCK-dependent expression in Sox9. 45 Overall, RhoA/ROCK signaling has been shown to be context dependent, altered by cell type and mechanical stimulation.
In our study, compression significantly increased RHOA expression, signifying that cells underwent morphological changes in cell shape. Rac1 often antagonizes RhoA and has been shown to aid in chondrogenic differentiation in adult stem cells by inhibiting cell spreading and stress fibers.20,44 Conversely, in our study, we found that Rac1 expression remained unchanged following compression. In fact, compression activated RhoA, resulting in increased YAP signaling. In addition, inhibition of Wnt signaling and degradation of β-Catenin in the cytoplasm43,46 adversely impact ESC self-renewal and expression of hypertrophic chondrogenic genes, Runx2, Col10, and Mmp13, which are implicated in the disease process of osteoarthritis. 47 When RhoA signaling was inhibited with RhoAi, CCG-1423, chondrogenic gene and protein expression decreased even in the presence of compressive stress, suggesting that RhoA plays an important role in the chondrogenic differentiation of ESCs in 3-D PDMS scaffolds. However, understanding the precise role of RhoA in mechanosignaling warrants further investigation.
In conclusion, our findings demonstrated a robust and simple approach for induction of ESC differentiation into early chondrogenic lineage. This research may lead to large scale production of a homogenous population of chondrogenic derivatives of ESCs and ultimately increase their use for cell therapy, tissue engineering, and regenerative medicine.
Footnotes
Acknowledgments
This research was supported by Oakland University, Michigan Head and Spine Institute (MHSI), and the Oakland University William Beaumont Institute for Stem Cell and Regenerative Medicine (OU-WB ISCRM). C. McKee was supported by the Oakland University Provost Graduate Student Research Award.
Disclosure Statement
No competing financial interests exist.