
Abstract
Extracted forms of collagen are subjected to chemical cross-linking to enhance their stability. However, traditional cross-linking approaches are associated with toxicity and inflammation. This work investigates the stabilization capacity, cytotoxicity and inflammatory response of collagen scaffolds cross-linked with glutaraldehyde (GTA), 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide, 4-arm polyethylene glycol (PEG) succinimidyl glutarate (4SP), genipin (GEN), and oleuropein. Although all cross-linking methods reduced free amine groups, variable data were obtained with respect to denaturation temperature, resistance to collagenase digestion, and mechanical properties. With respect to biological analysis, fibroblast cultures showed no significant difference between the treatments. Although direct cultures with human-derived leukemic monocyte cells (THP-1) clearly demonstrated the cytotoxic effect of GTA, THP-1 cultures supplemented with conditioned medium from the various groups showed no significant difference between the treatments. With respect to cytokine profile, no significant difference in secretion of proinflammatory (e.g., interleukin [IL]-1β, IL-8, tumor necrosis factor-α) and anti-inflammatory (e.g., vascular endothelial growth factor) cytokines was observed between the noncross-linked and the 4SP and GEN cross-linked groups, suggesting the suitability of these agents as collagen cross-linkers.
Introduction
C
Exogenous cross-linking methods, primarily chemical in nature, are customarily used as a means to control the mechanical stability and the degradation rate of collagen-based biomaterials. However, chemical cross-linking of collagen is associated with a predominant proinflammatory macrophage response, inhibition of macrophage polarization, reduced cell infiltration, increased proinflammatory cytokine expression, and delayed wound healing, resulting in cytotoxicity, reduced biocompatibility, and peri-implantation fibrosis.7–10
The mechanism behind the inflammatory and wound healing response to collagen cross-linking is still poorly understood. Macrophages have been described as a “rapid response” cell type, with crucial role in coordinating later inflammatory response and regeneration phases after implantation. 11 Interestingly, recent studies indicate that macrophage response can be modulated by the physicomechanical properties of an engineered scaffold and, in particular, the architectural,12,13 topographical, 14 and chemical 15 properties that can be variably affected as a function of the cross-linking method employed.
Macrophage phenotype switching is controlled by numerous factors, including disease and disease state, drugs, growth factors, cytokines, chemokines, hormones, cell shape, scaffold present, and its properties.16–21 Through direct interactions 12 and through released by-products, 22 cross-linked collagen devices directly modulate macrophage response in vitro. M1 macrophages (proinflammatory or classically activated) are activated by bacterial lipopolysaccharide (LPS), tumor necrosis factor α (TNF-α), and interferon γ (IFN-γ) to stimulate the secretion of large amounts of proinflammatory interleukin (e.g., IL-1α, IL-1β, IL-6, IL-8, IL-12, and IL-23) and TNF-α cytokines, reactive oxygen species, and matrix metalloproteinases to attack and phagocytize pathogens or foreign matter.23–25 M2 macrophages (anti-inflammatory or alternatively activated) are activated by IL-4, IL-10, and IL-13 or a combination of these to produce IL-1ra, IL-4, IL-10, IL-13, vascular endothelial growth factor (VEGF), transforming growth factor β, arginase, and scavenging molecules.23–25 Critically, M2 macrophages display differential functions according to the cellular subphenotype (e.g., anti-inflammatory [M2a], homeostatic [M2b], and prowound healing [M2c]). In instances that macrophages fail to remodel the substrate or to degrade foreign matter (frustrated phagocytosis), they aggregate and form foreign body giant cells, which are generally accepted as cells with higher degradation/resorption capacity than monocytes/macrophages. Furthermore, giant cells are related to peri-implantation fibrosis and foreign body response,26,27 often encountered with heavily cross-linked collagen devices. The mechanism behind this transformation is unclear; however, hydrophobicity, ionic charge, and soluble fusion mediators (e.g., receptor activator of nuclear factor kappa-B ligand, macrophage colony-stimulating factor, TNF-α, IL-1, IL-4, and IL-13) are believed to be important in the formation of foreign body giant cells. 28
Customarily used collagen cross-linking approaches, such as glutaraldehyde (GTA) and 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC), have been associated with cytotoxicity,29,30 calcification,31,32 and foreign body response.8,33 To this end, the use of alternative methods [multibranched polyethylene glycol (PEG) polymers34,35 and plant extracts (e.g., genipin [GEN], 36 myrica rubra, 37 and oleuropein [OLE] 38 ) has been advocated as a means to enhance mechanical and enzymatic stability, while reducing cytotoxicity. However, their immune response is yet to be assessed. Herein, the structural, physical, and biological properties of noncross-linked (NCL) and GTA, EDC, 4-arm PEG succinimidyl glutarate (4SP), GEN, and OLE cross-linked collagen films were assessed.
Materials and Methods
Materials
Bovine Achilles tendons were collected from a local abattoir (steers aged 24 months). 4SP (Mw 10,000), GEN, and OLE were purchased from JenKem Technology, Challenge Bioproducts (Taiwan), and Extrasynthese (France), respectively. All other materials and reagents were purchased from Sigma-Aldrich (Ireland), unless otherwise stated.
Type I collagen isolation
Bovine type I collagen was extracted from Achilles tendons by adapting a previously described protocol. 39 In brief, the tendons were manually separated from the surrounding fascia, cryo-milled (Freezer/Mill 6870; SPEX SamplePrep), and washed with 1× phosphate buffered saline (PBS) solution. Milled tendon tissue was dissolved in 1.0 M acetic acid under orbital agitation for 48 h at 4°C. Subsequently, pepsin (porcine gastric mucosa) was added in the solution at a ratio of 80 U/mg of milled tendon. The solution was incubated at 4°C for 72 h under stirring. Insoluble tendon was separated by filtration and centrifugation (21,000 g at 4°C for 20 min). Collagen solution was purified by repeated salt precipitation (0.9 M NaCl), centrifugation, and resuspension in 1.0 M acetic acid. The final atelocollagen solution was dialyzed (Mw 8000 cutoff) against 1 mM acetic acid and the final solution was kept at 4°C. Collagen concentration was determined using hydroxyproline assay (∼5 mg/mL) 40 and collagen purity was assessed using sodium dodecyl sulfate polyacrylamide gel electrophoresis followed by silver staining 41 (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
Collagen film fabrication
Typical protocols for the fabrication of collagen films were followed 42 with slight modifications. In brief, the pH of the collagen solution was adjusted to ∼7.3 using 1.0 M NaOH and 10× PBS. Cross-linking was conducted as per established protocols (Table 1). The final solutions were placed in silicone molds and incubated for 1 h at 37°C to induce gelation. Subsequently, the water content was evaporated overnight at 25°C.
Treatments: noncross-linked collagen film (NCL), collagen films cross-linked with glutaraldehyde (GTA), 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC), 4-arm PEG succinimidyl glutarate (4SP), genipin (GEN), and oleuropein (OLE).
PBS, phosphate buffered saline; PEG, polyethylene glycol.
Structural characterization
Morphological analysis of the produced films was analyzed using a Stereo Microscope (SZX16; Olympus, United Kingdom) and a Hitachi S-4700 scanning electron microscope (SEM; Hitachi, United Kingdom). Before SEM analysis, collagen films were incubated in 1× PBS overnight, dehydrated in ascending ethanol concentrations (50%, 70%, 90%, 96%, and 100%), and gold coated (Emitech K-550X Sputter Coater; Emitech, United Kingdom).
Quantification of free amines
Free amines were quantified using ninhydrin assay as has been described previously. 43 In brief, ∼3 mg of each film was mixed with 200 μL of deionized water and 1 mL of running buffer, which contained one part of 4% (w/v) ninhydrin in 2-ethoxyethanol and one part of 200 mM citric acid with 0.16% (w/v) tin (II) chloride at pH 5.0. The mixtures were incubated at 95°C for 30 min. The reaction was stopped by cooling down in ice and the addition of 250 μL of 50% isopropanol. After vortexing, the absorbance was read at 570 nm (Varioskan Flash Multimode Reader; Thermo Scientific). Glycine, at different concentrations, was used for the standard curve and the percentage of free amines of each condition was normalized against the NCL collagen films.
Quantification of denaturation temperature
The denaturation temperature was evaluated using a differential scanning calorimeter (DSC-60; Shimadzu, Japan), as has been described previously. 44 In brief, collagen films were incubated in 1× PBS at room temperature overnight and then they were quickly blotted using filter paper to remove surface/unbound water. The samples were then hermetically sealed in aluminum pans and were subjected to a single constant heating ramp at 5°C/min in the range of 25°C to 90°C. An empty pan was used as reference. Denaturation temperature was determined as the maximum heat absorption of the endothermic peak.
Quantification of enzymatic degradation
Resistance to enzymatic degradation was quantified as has been described previously. 45 In brief, collagen films were weighed and hydrated for 2 h in 0.1 M Tris-HCl and 5 mM CaCl2 at pH 7.4. Subsequently, the films were incubated in 10 U/mL bacterial collagenase type IV (Clostridium histolyticum), reconstituted in the same buffer. After 24 h of incubation at 37°C, centrifugation was carried out (10,000 g for 5 min), the supernatant was removed, and the remaining films were freeze dried and weighed. Enzymatic degradation was quantified as weight loss.
Quantification of mechanical properties
Uniaxial tensile test of hydrated (overnight incubation at 37°C in PBS and brief bloating in tissue paper before testing to remove surface liquid) films was performed using an electromechanical testing machine (Z2.5; Zwick, Germany). Uniform strips were prepared and, using a micrometer screw gauge, the width and thickness of the samples were measured. The grips of the testing machine were covered with a rubber film to avoid breakage at contact points. Samples that broke at contact points were excluded. The grips were set at 20 mm distance. The samples were deformed to complete failure (deformation rate of 10 mm/min, 10 N static load cell). The following parameters were assessed: force at break, stress at break, strain at break, and elastic modulus.
Human skin fibroblast response
Basic cellular functions were assessed using human skin fibroblasts (WS1, American Type Culture Collection [ATCC]). WS1 fibroblasts were grown in Eagle's minimum essential medium supplemented with 10% fetal bovine serum (FBS), 1% penicillin, and streptomycin. Films were sterilized in 70% ethanol for 30 min, followed by three washes in sterilized Hank's balanced salt solution (HBSS). Cells were seeded onto the samples at 16 × 103 cells/cm2 and incubated at 37°C, 5% CO2, and 95% humidified air for 1, 3, and 7 days. Phase contrast microscopy images were obtained using an inverted microscope (Leica microsystem, Germany) and images were analyzed with the LAS EZ 2.0.0 software. Cell proliferation was assessed using Quant-iT™ PicoGreen® dsDNA kit (Invitrogen), as per manufacturer's guidelines. Cell metabolic activity was assessed after 2 h incubation at 37°C with 10% alamarBlue® (Invitrogen), as per manufacturer's protocol. Cell metabolic activity was expressed in terms of percentage reduction of alamarBlue and normalized considering metabolic activity of cells in tissue culture plastic (TCP) at each time point as 100%. Cell viability was evaluated using Live/Dead® assay. In brief, samples were incubated in HBSS with 4 μM calcein and 2 μM ethidium homodimer for 30 min. Stained samples were observed using an inverted fluorescence microscope (IX 51; Olympus, United Kingdom). Five images were captured per film. Viable (green) and dead (red) cells were counted using ImageJ 1.48v software (National Institutes of Health).
Human macrophage response and cytokine release
Human-derived leukemic monocyte cells (THP-1; ATCC) were grown in RPMI-1640 supplemented with 10% FBS, 1% penicillin, and streptomycin. Cells were seeded onto the various samples at 26 × 103 cells/cm2 and mature macrophage-like state was induced through treatment with phorbol 12-myristate 13-acetate (PMA) at 100 ng/mL for 6 h, as has been described previously.46–48 Subsequently, plastic-adherent cells were washed with HBSS and incubated with supplemented medium at 37°C, 5% CO2, and 95% humidified air for 1 and 2 days. Activated positive control phenotype was induced with 100 ng/mL of LPS in supplemented medium for 24 h. Cell proliferation and metabolic activity were determined as described in “Human skin fibroblast response” section. Cellular viability was quantified using CytoTox 96® nonradioactive cytotoxicity assay (Promega) to measure released lactate dehydrogenase in the supernatant from dead cells. Phase contrast microscopy images were obtained using an inverted microscope (Leica microsystem) and images were analyzed with the LAS EZ 2.0.0 software.
Inflammatory cytokines (IL-1β, IL-4, IL-6, IL-8, IL-10, IL-12p70, IL-13, TNF-α, and VEGF) were measured using Meso Scale Discovery (MSD) electrochemoluminescense assay, as per manufacturer's guidelines. In brief, cell-free supernatants of each sample and standards were incubated on MSD plates for 2h followed by a wash. The plates were then incubated with detection antibody solution for 2 h. Subsequently, the plates were washed and read using a Meso™ QuickPlex SQ120 instrument (MSD).
The potential effect of released/degradation subproducts of cross-linked collagen films was investigated using the preconditioned medium. Cross-linked films were incubated in supplemented RPMI-1640 medium at 37°C and 5% CO2 for 6 days before being exposed to cells. THP-1 cells were seeded at 26 × 103 cells/cm2 with 100 ng/mL of PMA for 6 h. Subsequently, plastic-adherent cells were washed with HBSS and incubated with preconditioned medium at 37°C, 5% CO2, and 95% humidified air for 48 h. An activated control was induced with 100 ng/mL of LPS for 24 h. Characterization of macrophage cells was performed as already described.
Statistical analysis
All experiments were carried out in triplicate, except swelling, denaturation temperature, and tensile test assays that were carried out in quintuplicate. Numerical data are expressed as mean ± standard deviation. Statistical analysis was performed using MINITAB® (version 16.2; Minitab, Inc.). One way analysis of variance followed by Fisher's post hoc test was employed after confirming normal distribution from each sample population (Anderson–Darling normality test) and the equality of variances (Bartlett's and Levene's tests for homogeneity of variance). Nonparametric statistics were used when either or both of the mentioned assumptions were violated and, consequently, Kruskal–Wallis test for multiple comparison analysis was carried out. Statistical significance was accepted at p < 0.05.
Results
Structural characterization
NCL collagen films and cross-linked collagen films with EDC and 4SP were colorless and totally transparent, GTA and OLE cross-linking made the films yellow/brown and semitransparent, and GEN cross-linking resulted in dark blue and totally opaque films (Supplementary Fig. S2). SEM analysis revealed that NCL and 4SP cross-linked films maintained the fibrillar structure of collagen, which was diminished in EDC, GTA, GEN, and OLE cross-linked films (Supplementary Fig. S2).
Quantification of free amines
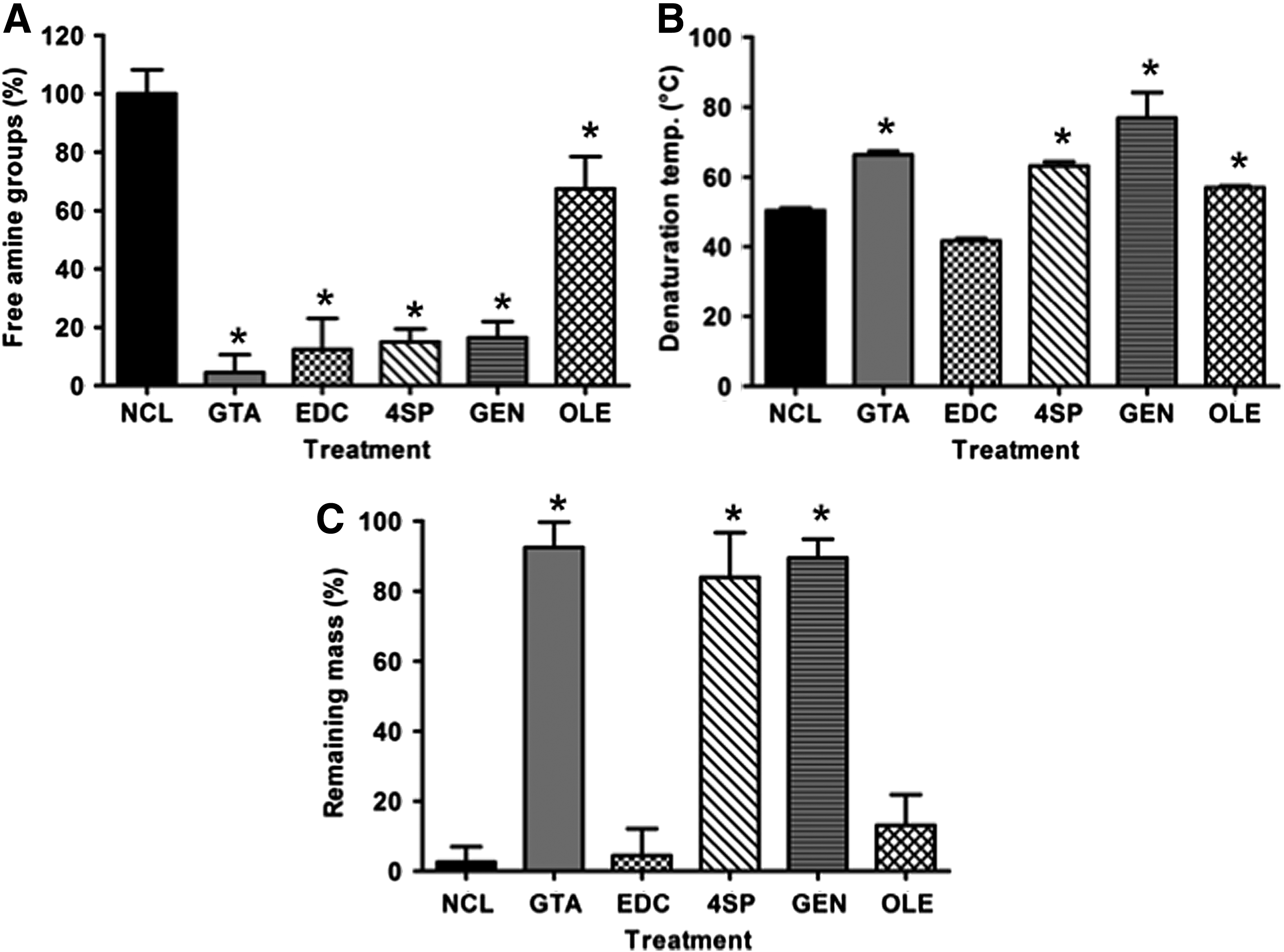
Collagen cross-linking involves the formation of covalent bonds between the free amine or carboxyl groups of collagen with the cross-linking agent; reduction in free amines can be used as a cross-linking efficiency indicator. A significant (p < 0.001) decrease in free amine groups was observed for all cross-linked collagen films (Fig. 1A). Among the cross-linked groups, GTA, EDC, 4SP, and GEN brought about an ∼80% reduction in free amines, whereas OLE induced an ∼40% reduction in the amount of free amines (Fig. 1A).
Quantification of denaturation temperature
DSC analysis revealed that all cross-linking methods, but EDC (p > 0.05), significantly increased (p < 0.001) the denaturation temperature of the produced collagen films, as compared with that of the NCL films (Fig. 1B).
Quantification of enzymatic degradation
In vitro enzymatic degradation analysis (Fig. 1C) revealed that NCL, EDC, and OLE films were almost completely degraded within 24 h, whereas GTA, 4SP, and GEN induced a very high resistance to enzymatic degradation (less than 20% was degraded).
Quantification of mechanical properties
Stress–strain curves consisted of a small toe region, a region of steeply rising stress, and a long region of constant gradient until fracture (Supplementary Fig. S3). GEN-stabilized films exhibited the highest (p < 0.001) force at break, stress at break, and elastic modulus values, whereas EDC cross-linked films exhibited the highest (p < 0.001) strain at break values (Table 2). EDC-stabilized films exhibited the lowest (p < 0.001) force at break, stress at break, and elastic modulus values, whereas GTA cross-linked films exhibited the lowest (p < 0.001) strain at break values (Table 2).
The highest (bp < 0.001) force at break, stress at break, strain at break, and elastic modulus values were obtained from the GEN, GEN, EDC, and GEN cross-linked films, respectively. The lowest (ap < 0.001) force at break, stress at break, strain at break and elastic modulus values were obtained from the EDC, EDC, GTA and EDC cross-linked films, respectively. Treatments: NCL collagen film, collagen films cross-linked with GTA, EDC, 4SP, GEN, and OLE.
Human skin fibroblast response
Phase contrast microscopy analysis revealed that human skin fibroblasts maintained their spindle-shaped morphology, independently of the cross-linking method and culture time (Supplementary Fig. S4). By day 7, DNA quantification (Fig. 2A; only OLE was significantly higher [p < 0.001] than the NCL control), cell metabolic activity (Fig. 2B), and cell viability (Fig. 2C) assays revealed no apparent differences between the NCL and cross-linked groups (p > 0.05).
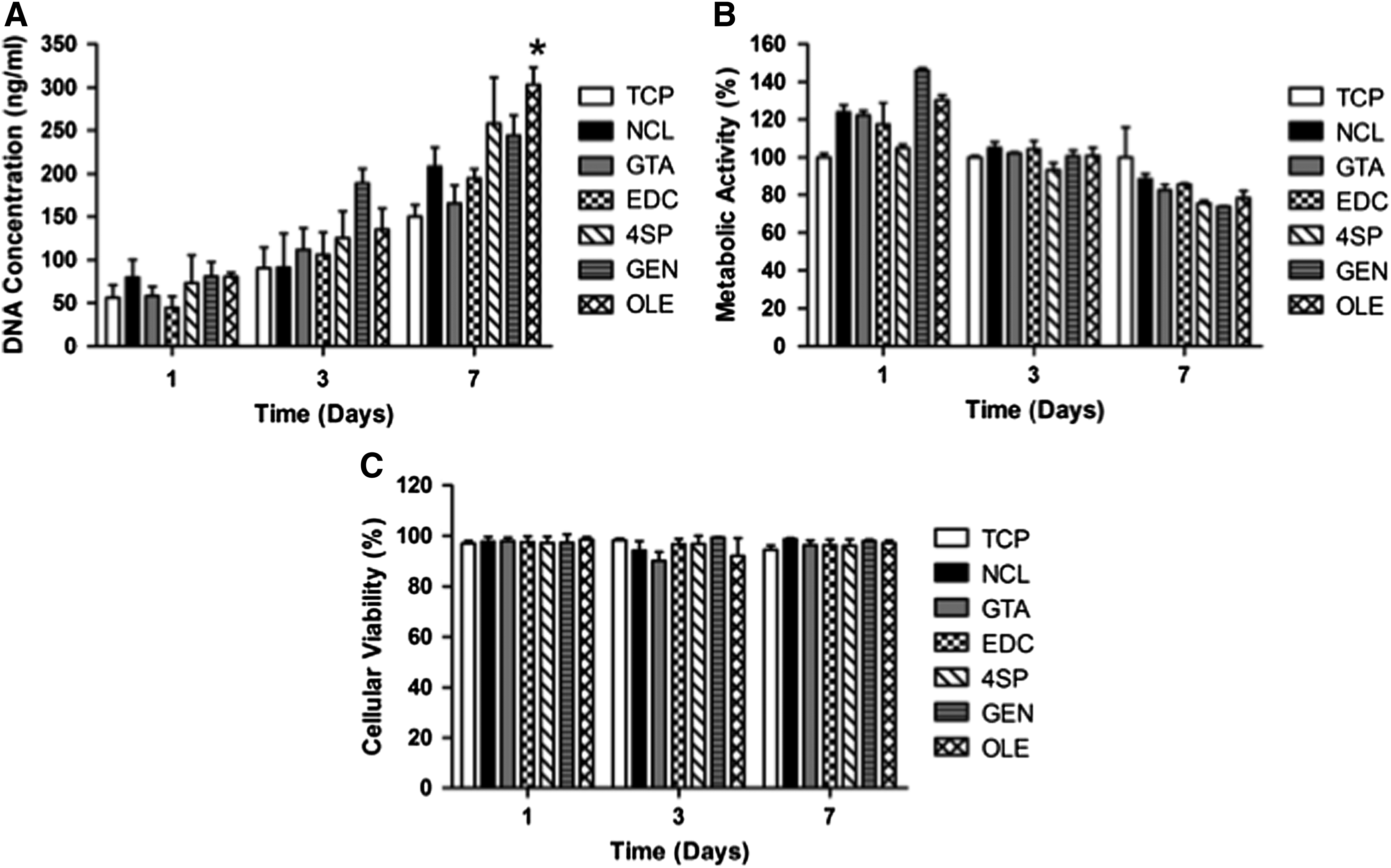
Human skin fibroblasts onto cross-linked collagen films after 1, 3, and 7 days of culture. Cellular proliferation was assessed through DNA concentration quantification
Human macrophage response and cytokine release
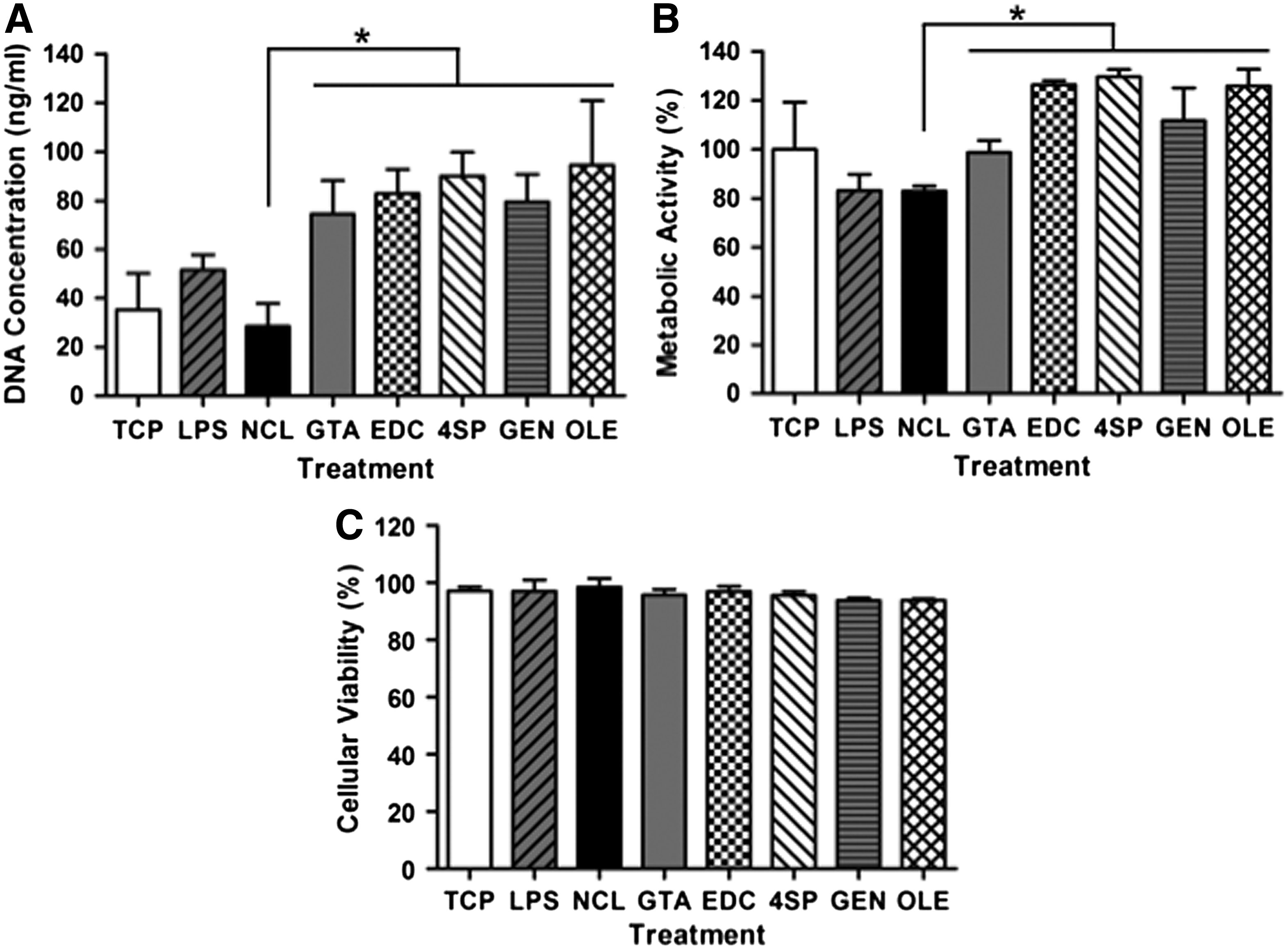
Phase contrast microscopy analysis revealed that by day 2, (1) most macrophages, independently of the treatment and time in culture, adopted a round morphology; (2) some elongated cells were observed on TCP, GEN, LPS, 4SP, and OLE; and (3) all treatments, but GTA, formed aggregates (Supplementary Fig. S5). Cells grown on TCP, LPS, and NCL exhibited significantly higher (p < 0.001) DNA concentration than cells grown on GTA, EDC, 4SP, and GEN at day 1, whereas at day 2, cells grown on TCP, LPS, and NCL exhibited significantly higher (p < 0.001) DNA concentration than cells grown on GTA, EDC, 4SP, GEN, and OLE (Fig. 3A). GTA cross-linked films induced the lowest (p < 0.001) cell metabolic activity at both time points; TCP, LPS, and OLE induced the highest (p < 0.001) cell metabolic activity at day 1; and TCP, LPS, GEN, and OLE induced the highest (p < 0.001) cell metabolic activity at day 2 (Fig. 3B). GTA cross-linked films induced the lowest cell viability (p < 0.001) at both time points; no significant difference was observed between EDC, 4SP, GEN, and OLE at both time points (Fig. 3C).
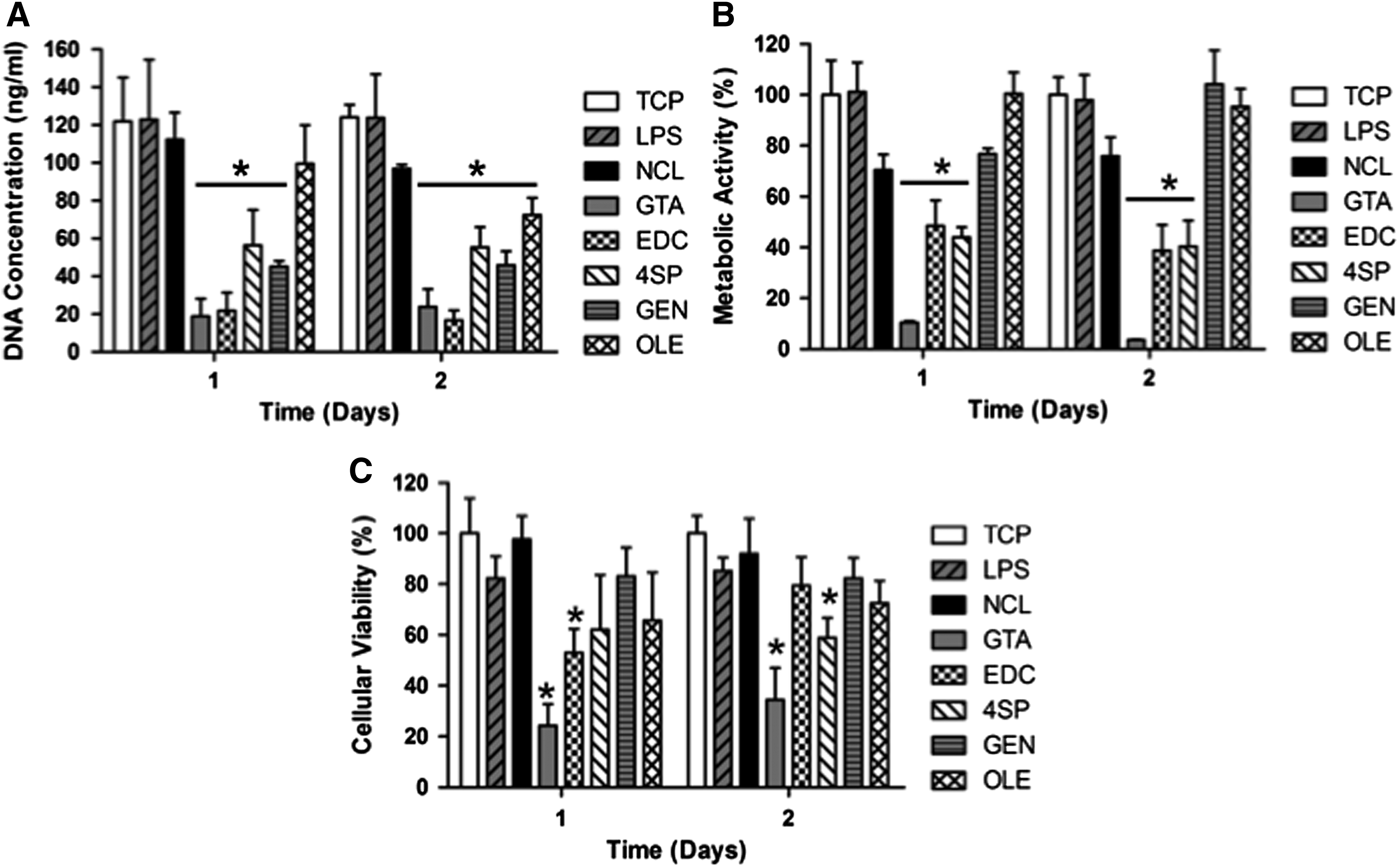
THP-1 cells onto cross-linked collagen films after 1 and 2 days of culture. Cellular proliferation was assessed through DNA concentration quantification
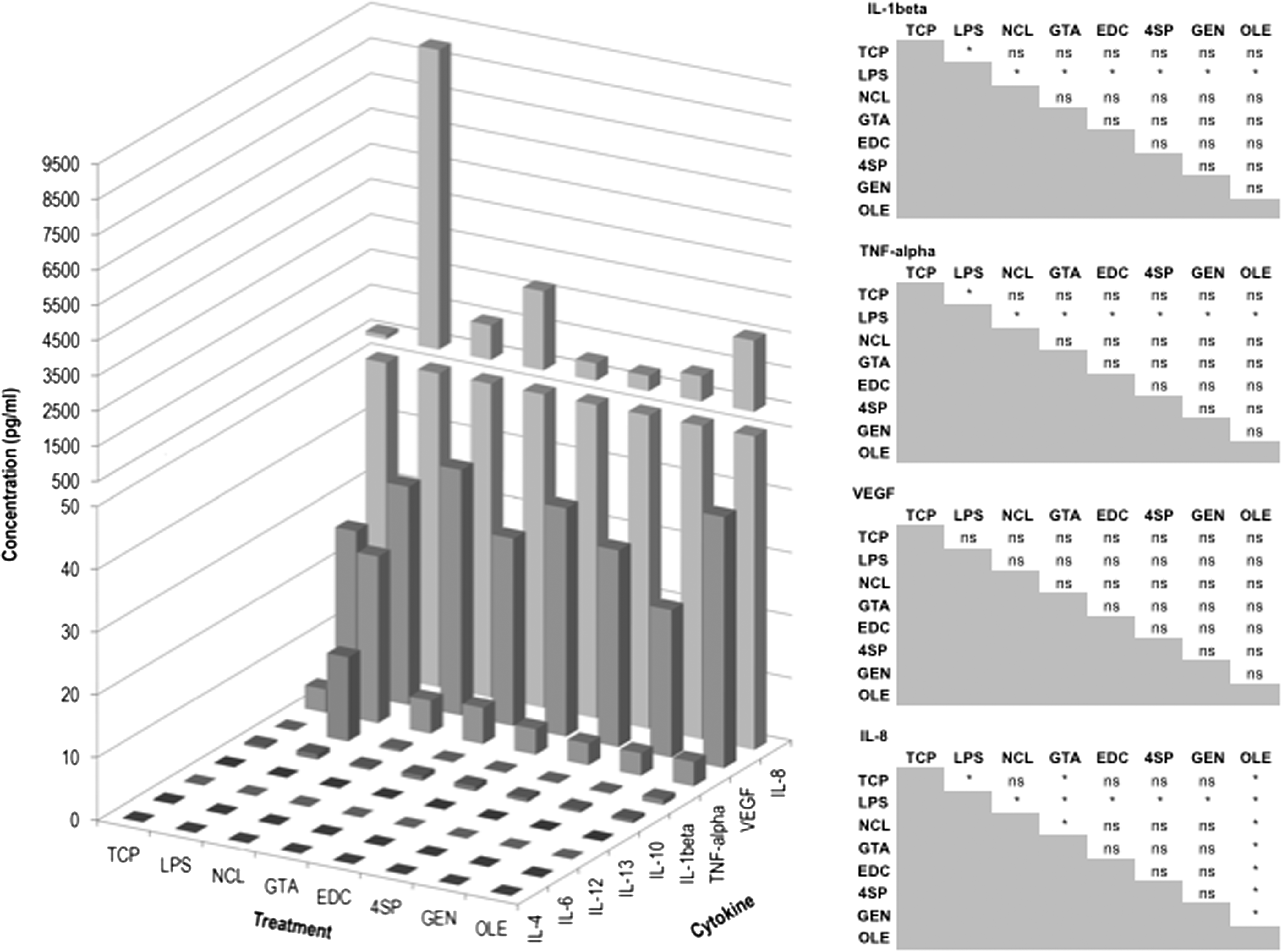
When THP-1 cells were seeded on the various substrates, no significant difference (p > 0.05) in secretion of IL-1β, IL-8, TNF-α, and VEGF was detected between NCL, 4SP, and GEN groups at both time points (Fig. 4). Cytokine release of IL-4, IL-6, IL-12, IL-13, and IL-10 was below the detection limit for all treatments for both time points (Fig. 4). Cytokine release from cells seeded on GTA films was very low (Fig. 4) because of the low number of cells (Fig. 3C).
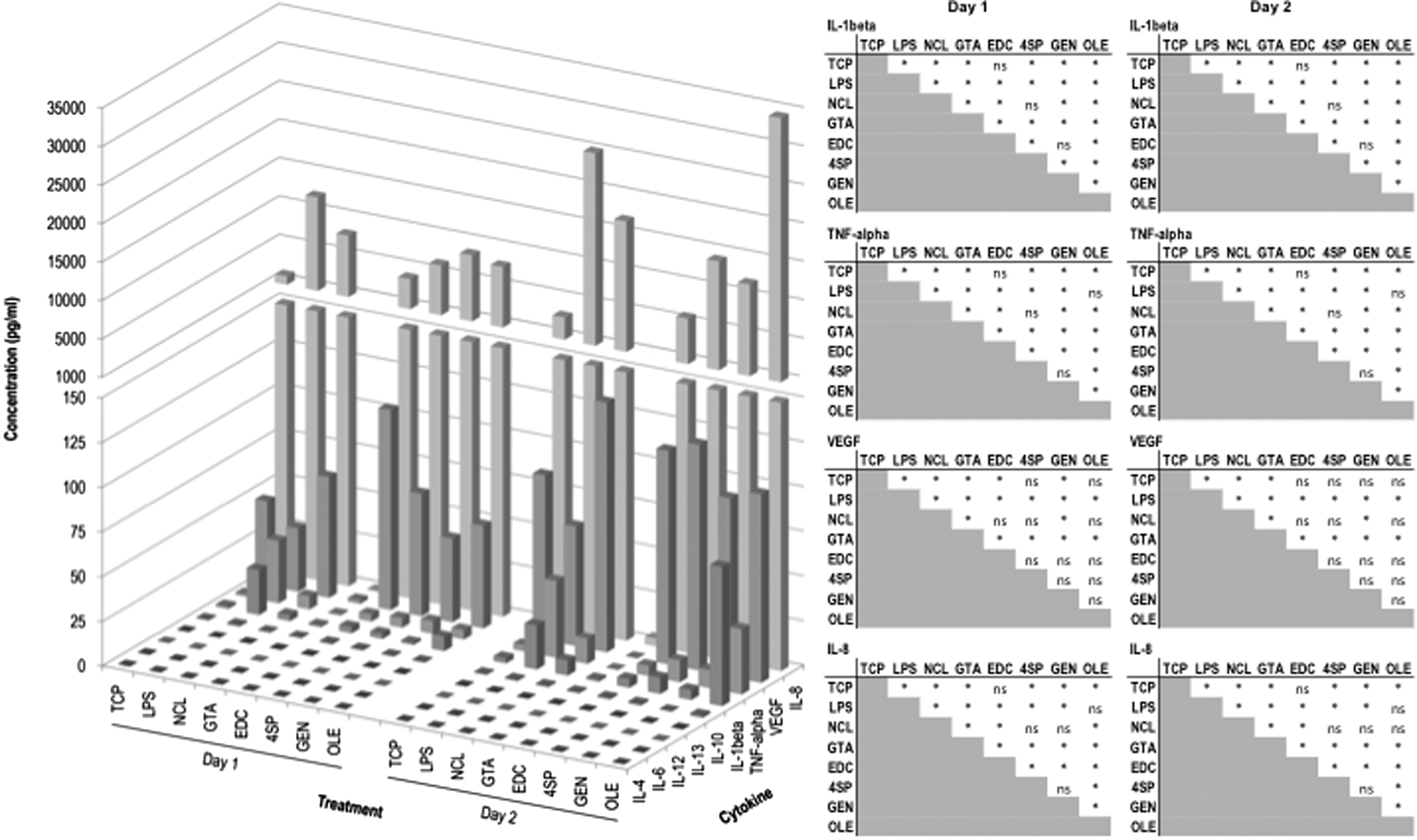
Multiplex ELISA on proinflammatory and anti-inflammatory cytokines of THP-1 cells cultured on distinct films for 1 and 2 days. Left: the bar represents the average. Right: multiple comparison tests for every pair of conditions (*p < 0.05, ns: not significant). Treatments: NCL collagen film, collagen films cross-linked with GTA, EDC, 4SP, GEN, and OLE. TCP and LPS-stimulated TCP (LPS) were used as controls.
Phase contrast microscopy analysis revealed that by day 2, (1) most macrophages, independently of the treatment, adopted a round morphology, (2) some elongated cells were observed on all samples, but TCP and OLE, and (3) all treatments, but OLE, formed aggregates (Supplementary Fig. S6). All cross-linking treatments exhibited significantly higher (p < 0.05) DNA concentration (Fig. 5A) and metabolic activity (Fig. 5B) than the NCL counterparts, whereas no significant difference (p > 0.05) was observed in cell viability (Fig. 5C). With respect to inflammatory cytokine release, only preconditioned medium with GTA and OLE cross-linked films significantly increased (p < 0.05) IL-8 release (Fig. 6), in comparison with the NCL samples. No significant difference (p > 0.05) was detected for VEGF, TNF-α, and IL-1β between the treatments (Fig. 6). Cytokine release of IL-4, IL-6, IL-12, IL-13, and IL-10 was below the detection limit for all treatments (Fig. 6).
THP-1 cells cultured with preconditioned medium from the cross-linked collagen films after 2 days of culture. Cellular proliferation was assessed through DNA concentration quantification
Multiplex ELISA on proinflammatory and anti- inflammatory cytokines of THP-1 cells cultured with preconditioned medium from the cross-linked collagen films after 2 days of culture. Left: the bar represents the average. Right: multiple comparison tests for every pair of conditions (*p < 0.05, ns: not significant). Treatments: NCL collagen film, collagen films cross-linked with GTA, EDC, 4SP, GEN, and OLE. TCP and LPS-stimulated TCP (LPS) were used as controls.
Discussion
Collagen-based devices represent one of the largest subsets of biomaterials currently in clinical practice. 49 Chemical cross-linking methods50–52 are customarily employed to induce stability and to offer control over enzymatic degradation, yet collagen biomaterials are commonly associated with persistent and chronic activity of M1 macrophages, 33 resulting in inflammation, suboptimal in vivo performance, and, in several cases, implant failure.7,8 Herein, we ventured to assess the influence of various cross-linking methods (e.g., GTA, EDC, 4SP, GEN, and OLE) on the structural, biochemical, biophysical, and biological properties of collagen-based films (Table 3 summarizes our findings).
= denotes nonsignificant (p > 0.05) alterations with respect to the NCL group; ↑ denotes a significant (p < 0.05) increase with respect to the NCL group; and↓ denotes a significant (p < 0.05) reduction with respect to the NCL group. Treatments: NCL collagen film, collagen films cross-linked with GTA, EDC, 4SP, GEN, and OLE.
IL, interleukin.
Although collagen can be cross-linked through its free amine or carboxyl groups, each cross-linking method has a unique mode of action, which consequently would be responsible for the properties of the resultant scaffold. GTA, for example, is considered a potent, but cytotoxic, collagen cross-linker that, in physiological conditions, reacts with the amine groups (primarily lysine, hydroxylysine, histidine, arginine, and tyrosine) to form Schiff bases. Carboxylic acid groups of aspartic and glutamic acid residues in collagen react with EDC and then with N-hydroxysuccinimide (NHS); the NHS-activated carboxylic acid groups then react with the amine groups of lysine and hydroxyl lysine residues and release the NHS. EDC has limited cross-linking ability because of its short length structure and inability to polymerize, but is not as cytotoxic as GTA. The succinimidyl groups of 4SP react with the amine groups present on the collagen molecule. 4SP is not as potent as GTA, but it is significantly more cytocompatible. GEN cross-links free amine groups (e.g., lysine, hydroxylysine, and arginine) and forms intramolecular and intermolecular cross-links with collagen. OLE is an iridoid glycoside that contains a glucose molecule, which is cleaved by β-glucosidase to produce an aglycone, which then reacts with the lysine residues of proteins. Both GEN and OLE are almost as efficient cross-linkers as GTA and less cytotoxic. A direct comparison between these prominent collagen cross-linkers is yet to be reported.
NCL collagen films and cross-linked collagen films with EDC and 4SP were clear and transparent, cross-linked collagen films with GTA and OLE were brown, and cross-linked films with GEN were dark blue. The observed discoloration is attributed to the reaction between the cross-linking agents and the amino acids of collagen, as has been reported previously.36,38,53–55 OLE presented a few brown dots that we attributed to clusters of the β-glucosidase enzyme used to activate OLE aglycones, which bind to collagen amines. 38 The surface morphology was found to be cross-linking dependent, as it has been reported previously,55–58 but in general a typical fibrous surface 59 was observed. GTA and OLE exhibited the least fibrous surface/smoothest surface, which is in agreement with previous observations, in which cross-linking reduced/preferentially altered surface roughness.42,60,61
Overall, cross-linking reduced the percentage free amines and increased the denaturation temperature and the resistance to enzymatic degradation of the collagen films. 4SP and GEN exhibited values similar to GTA, further advocating previous studies on their use as collagen cross-linking agents.34,35,53–55,62,63). Although EDC is used extensively as a GTA alternative,64–68 herein EDC samples despite exhibiting lower percentage of free amines than the NCL samples, neither the denaturation temperature nor the resistance to enzymatic degradation was improved. Furthermore, although OLE has been claimed to be a potent collagen cross-linker,38,69 herein it reduced the percentage of free amines and increased the denaturation temperature, but did not improve resistance to enzymatic degradation. Overall, these data illustrate that an array of assays should be conducted to assess the potential of a collagen cross-linking agent.
The mechanical properties of the cross-linked scaffolds also appeared to be method dependent, with EDC producing the most extendable scaffolds and GEN resulting in scaffolds with the highest force at break, stress at break, and elastic modulus values. Although both GTA70–73 and EDC74–76 have been reported repeatedly to produce scaffolds with mechanical properties superior to NCL counterparts and/or other cross-linkers, our data are in agreement with previous observations, in which GEN significantly improved mechanical resilience77–80 of collagen-based scaffolds and EDC failed to yield scaffolds with strength similar to more potent cross-linkers.55,81,82 With respect to deformation mechanism, typical stress–strain curves of collagen materials that yield and undergo plastic flow were observed. The low modulus of the toe region, which is succeeded by the nonlinear stress–strain curve, has been attributed to reorientation and uncramping of the collagen fibrils and the initiation of stretching of the triple helix, the nonhelical ends, and the cross-links.36,83–85
With respect to human skin fibroblast compatibility assessment, none of the cross-linking methods affected cell morphology (in all cases, human skin fibroblasts maintained their spindle-shaped morphology) and they were not cytotoxic (no significant reductions were detected in DNA concentration, cell viability, and cell proliferation at a given time point between NCL and cross-linked scaffolds). Previous studies have reported both cytotoxic 86 and noncytotoxic 87 effects of EDC and GTA cross-linked collagen scaffolds. These contradictive data suggest that the cross-linking concentration/postcross-linking treatments (e.g., number, duration, and composition of washing bath) should be optimized for a specific device to identify the optimal cross-linker concentration with the desired cell response.
Macrophage polarization has been closely linked to its shape. For example, macrophage elongation has been associated with transition from a proinflammatory M1 phenotype into an anti-inflammatory M2 phenotype, 18 whereas cell aggregates are indicative of fusogenic phenotype and foreign body response. 26 Qualitative cell shape analysis revealed that most THP-1 macrophages exhibited a round morphology, whereas some elongated THP-1 cells were also identified. Furthermore, THP-1 aggregates were also formed. Such heterogeneous macrophage populations have been reported previously, when mature human monocyte-derived macrophages were exposed to polyethylene particulate 88 and have been associated with macrophage phenotype variations.18,89–91
By day 2, all cross-linked scaffolds significantly reduced THP-1 DNA concentration; GTA, EDC, and 4SP significantly reduced THP-1 metabolic activity; and GTA and 4SP significantly reduced THP-1 viability in comparison with the NCL samples. When macrophages were treated with preconditioned medium, NCL medium significantly reduced THP-1 DNA concentration and metabolic activity, whereas cell viability was not affected. Reduction in cell proliferation, viability, and metabolic activity as a function of cross-linking method employed indicates cytotoxicity, as has been observed previously for macrophages exposed to GTA cross-linked bovine pericardium-derived scaffold, 12 but was not verified herein with dermal fibroblasts and was not verified with the preconditioned medium. Although previous data have demonstrated the influence of substrate rigidity on macrophage response,92,93 no clear correlation was observed herein between scaffold rigidity, as a function of the different cross-linking method utilized, and macrophage response. Previous studies have demonstrated the influence of surface topography and chemistry on macrophage response.94–97 We believe that this may be the reason for the variable cell response to the various scaffolds, but more detailed analysis (e.g., scaffolds with very specific topography, cross-linkers with similar chemistry) is required to verify this assumption.
THP-1 cells seeded on collagen scaffolds secreted significant amounts of 1 L-1β (proinflammatory), TNF-α (proinflammatory), IL-8 (proinflammatory), and VEGF (proangiogenesis), with OLE samples exhibiting the highest 1 L-1β, TNF-α, and IL-8 values and EDC and 4SP samples exhibiting the highest VEGF values. Previous studies have demonstrated high IL-1β, IL-6, and TNF-α secretion when GTA was used98,99; this was not verified here because of the toxicity of the cross-linker (although GTA because of its self-polymerization capacity can cross-link amines that are relatively far apart,100,101 degradation products and unreacted GTA result in high cytotoxicity29,102 and calcification103,104). With respect to EDC, similarly to our work, previous studies have demonstrated comparable to the control bovine pericardium tissue proinflammatory cytokine profile. 12 In vitro data for GEN are not available; however, previous in vivo studies have shown inflammatory response to GEN cross-linked bovine pericardium as a function of cross-linking concentration. 105 Although in our hands OLE films exhibited TNF-α production similar to LPS, previous studies have shown OLE to attenuate IL-1β, IL-6, and TNF-α secretion in LPS-activated macrophages. 106 With respect to 4SP inflammatory response, in vitro data are not available as yet. To date though, all published articles have shown promise in various clinical indications, including tendon, 55 nucleus pulposus, 35 cartilage,34,107 and cornea. 108
Conclusions
During the extraction and purification of collagen, native cross-linking is degraded. To reinstate integrity to reconstituted forms of collagen, exogenous cross-links are incorporated into the supramolecular structure, primary through chemical approaches. However, traditional cross-linking methods (e.g., GTA and EDC) are often linked to poor cytocompatibility, high inflammation, and poor tissue integration. Herein we assessed the stability and biological response of collagen-based devices stabilized with GTA, EDC, 4SP, GEN, and OLE. All cross-linking methods reduced free amine groups. Denaturation temperature, resistance to collagenase digestion, and mechanical properties were cross-linking method dependent. No significant differences were observed between the treatments in fibroblast cultures. Only the GTA-cross-linked scaffolds were cytotoxic to human-derived leukemic monocyte cells, whereas cultures supplemented with conditioned medium from the various groups showed no significant difference between the treatments. No significant difference in proinflammatory and anti-inflammatory cytokines was observed between the NCL and the 4SP and GEN cross-linked groups, suggesting the suitability of these agents as collagen cross-linkers.
Footnotes
Acknowledgments
This work was supported by the European Union Seventh Framework Programme (FP7/2007-2013), under grant agreement number 263289 (Green Nano Mesh); European Union Seventh Framework Programme (FP7/2007-2013), under grant agreement number 251385 (Tendon Regeneration); Health Research Board, Health Research Awards Programme, under grant agreement number HRA_POR/2011/84; and Enterprise Partnership Scheme, Irish Research Council, under grant agreement number EPSPG/2011/64. This publication has also been supported by Science Foundation Ireland and the European Regional Development Fund, under grant agreement number 13/RC/2073.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.