
Abstract
Macrophages are pivotal cells during the foreign body reaction (FBR), as they orchestrate the proinflammatory microenvironment inside and around biomaterials by secretion of inflammatory mediators. Furthermore, they are responsible for the degradation of biomaterials and are thought to instruct the fibroblasts that generate a fibrous capsule around implanted biomaterials. In this study, we investigated the events during the FBR when macrophages are not present. Hexamethylenediisocyanate crosslinked collagen scaffolds were implanted in “Macrophage Fas-Induced Apoptosis” mice, which allow “on demand” macrophage depletion. We observed that macrophage depletion completely inhibited inflammatory ingrowth into the scaffolds and resulted in an increased capsule size. Quantitative polymerase chain reaction analysis revealed decreased expression levels of proinflammatory mediators such as TNFα and IL1β, and increased expression levels of collagens and fibroblast-stimulating growth factors such as EGF, FGF1, FGF2, and TGFα. Our results indicate that macrophages are indeed crucial for the generation of a proinflammatory microenvironment inside implanted biomaterials, leading to inflammatory ingrowth. In contrast, macrophages do not appear to be important for the generation of a fibrous capsule around implanted biomaterials. In fact, our data suggest that the macrophages present in the capsule might instruct the surrounding fibroblasts to produce less fibroblast-stimulating factors and less collagens.
Introduction
U
In this study, we interrogated the dogma that macrophages are the most important cell type during the FBR by investigating the FBR in a mouse model that enables conditional macrophage ablation. While many models for experimental macrophage depletion have been described, for example, administration of clodronate liposomes 21 or administration of diphtheria toxin to CD11b-DTR mice, 22 depletion efficiency varies dramatically in these models (typically between 50% and 95%). In this study, we employed macrophage fas-induced apoptosis (MaFIA) mice, which enabled highly efficient (>95%) macrophage depletion. In these mice, a transgene is expressed under control of the c-fms promotor, resulting in expression that is restricted to the monocytic lineage, including monocytes, macrophages, dendritic cells (DC), Kuppfer cells, Langerhans cells, osteoclasts, and microglial cells.23,24 The transgene encodes an inducible suicide protein, which drives Fas-mediated apoptosis 25 after stimulation with exogeneous AP20187.26,27 After induction of depletion, we implanted hexamethylenediisocyanate-crosslinked dermal sheep collagen (HDSC) discs in the mice. The ingrowth of inflammatory cells and capsule formation were studied using histology and immunohistochemistry, and mRNA expression was analyzed using quantitative polymerase chain reaction (qPCR).
Materials and Methods
Animals and the depletion of macrophages
All animal experiments and breeding protocols were performed according to governmental and international guidelines on animal experiments, and were approved by the local Animal Care and Use Committee of the University of Groningen. MaFIA mice were purchased [C57BL/6-Tg(Csf1r-EGFP-NGFR/FKBP1A/TNFRSF6)2Bck/J, JAX Laboratories] and subsequently housed and bred in the central animal facility of the University of Groningen. During experiments, animals were weighed daily and blood was drawn at regular intervals to quantify the number of white blood cells, red blood cells, and platelets using a pocH-100i machine. Similarly, hemoglobin and hematocrit levels were assessed. Conditional depletion of macrophages was achieved by daily intraperitoneal injection of 10 mg/kg AP20187 (Ariad Pharmaceuticals) for 12 days. AP20187 was dissolved in diluent consisting of 4% ethanol, 10% PEG-400, and 1.72% Tween-20 in saline. Control animals were injected with diluent without AP20187 according to the same administration regime. After 4 days, the depletion of macrophages was considered to be complete and biomaterials were implanted.
Biomaterials
The impact of macrophage depletion on the FBR was studied in our standard model of the FBR.28–30 Sheep-derived dermal collagen (Zuid Nederlandse Zeemleder fabriek, Oosterhout, The Netherlands) was crosslinked with hexamethylenediisocyanate to form noncytotoxic HDSC, 31 a porous and degradable biomaterial devoid of immunogenic groups. Potential endotoxin contamination was assessed using the LAL-method (LAL kinetic-QCL; Cambrex), and no contamination was found (i.e., <0.05 EU/mL). Cylinder-shaped discs with a height of 1 mm and a diameter of 6 mm were made out of a sheet of HDSC using a biopsy punch.
Surgical procedures
Macrophage-depleted and control mice (N = 3) were anesthetized with 4% isoflurane. Two percent isoflurane was used to maintain anesthesia. After shaving and disinfection, two subcutaneous pockets were created from a single midline incision on the back of the mice, toward both flanks of the animals, at about 1 cm from the midline of the animal. HDSC discs were implanted in both pockets. Perioperatively, buprenorphine (0.01 mg/kg) was administered as pain medication. After 7 days, HDSC discs plus surrounding tissue were dissected from the pockets. After dissection, one of the discs was used for RNA isolation, and the other disc was cut in half. One half was used for histological analysis, and the other half was used for immunohistochemistry. Spleens, livers, and kidneys were explanted, weighed, and subsequently used for histological analysis. After explantation, the animals were killed under anesthesia using cervical dislocation.
RNA isolation, cDNA synthesis, and qPCR
RNA isolation was performed using a FavorPrep Total RNA Isolation Mini Kit (FATRK001; Favorgen). HDSC discs were pulverized in FARB lysis buffer using plungers provided in the kit. Subsequent RNA isolation was performed according to the manufacturer's instructions. Isolated RNA was transcribed into cDNA using a First-Strand cDNA Synthesis Kit (#K1622; Fermentas). qPCR analysis was performed with 5 ng cDNA using SYBR Green Supermix (Bio-Rad) to quantify the expression of selected genes. Primers were developed using Roche Assay Design Center (Table 1). The reaction mixtures were incubated at 95°C for 15 s, 60°C for 30 s, 72°C for 30 s for 40 cycles in a ViiA 7 qPCR machine (Invitrogen). ACTB was used as a reference gene and relative expression was calculated as 2−ΔCT.
Histological analysis and immunohistochemistry
After explantation, one half of the HDSC discs was fixed in 2% (v/v) glutaraldehyde in 0.1 mM phosphate buffer (pH 7.4). Glutaraldehyde-fixed materials were embedded in Technovit 7100 (Heraeus Kulzer) according to the manufacturer's protocol, after dehydration in graded alcohol dilution series. Two micrometer sections were cut and stained with Toluidine Blue (Fluka Chemie) and subsequently analyzed by light microscopy. The other half of the discs was fixed in zinc fixative, consisting of 100 mM Tris, 3.2 mM calcium acetate, 27 mM zinc acetate, and 37 mM zinc chloride. Zinc-fixed materials were embedded in paraffin after dehydration in graded alcohol dilution series followed by xylol. Sections of 5 μm were cut and used for immunohistochemistry. Sections were deparaffinized in xylol followed by graded series of ethanol. Proteinase K treatment was used as antigen retrieval method. Sections were preincubated with 10% rabbit serum and subsequently incubated in 10 μg/mL F4/80 antibody (MCA497; Serotec) to detect macrophages. Endogeneous peroxidase activity was blocked by treatment with H2O2. Primary antibodies were detected with 13 μg/mL peroxidase-conjugated rabbit–anti-rat IgG (P0450; Dako). 3-Amino-9-ethylcarbazole (A6926; Sigma) was used for color development, and Mayer's Hematoxylin (51260; Fluka) was used as counterstain. Spleens, livers, and kidneys were fixed in 4% paraformaldehyde, subsequently dehydrated in graded alcohol dilution series, and finally embedded in paraffin. Sections of 5 μm were cut and stained according to the Periodic Acid–Schiff method.
Quantifications of histological and immunohistochemical analyses
Six representative photographs were taken from histological samples using Leica Qwin software on a Leica microscope. The area occupied by a fibrous capsule, inflammatory ingrowth, and the number of fibroblast-like cells (i.e., spindle-shaped elongated cells with flattened nuclei) were quantified in these photographs using ImageJ. Similarly, the area positive for F4/80 immunostaining was quantified in six micrographs using a NuanceFX Multispectral Imaging System (PerkinElmer).
Statistical analysis
Data are expressed as mean ± standard error of the mean. Statistical significances were evaluated by a Student's t-test. A difference was considered statistically significant when p < 0.05.
Results
Validation of macrophage depletion model
MaFIA mice were treated with AP20187 for 4 days (from d4 until d0) to induce complete depletion of macrophages. 26 Subsequently, at d0, HDSC discs were implanted into dorsolateral subcutaneous pockets to induce the FBR. Treatment with AP20187 was continued to maintain macrophage depletion. After 7 days (day 7), the HDSC discs and surrounding tissue were explanted and processed for further analysis. Treatment with AP20187 caused a drop in white blood cell numbers in peripheral blood as observed at d0, which persisted as long as treatment was continued, until d7 when the experiment was terminated (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
To verify the successful depletion of macrophages, immunohistochemistry for F4/80 was performed on sections from the HDSC scaffolds and surrounding tissue (Fig. 1A). In nondepleted mice, that is, diluent-treated controls, high macrophage numbers (as indicated by positivity for F4/80) were present in the capsule and in the ingrowth inside the implanted discs. A few F4/80-positive multinucleated giant cells were also observed in the ingrowth. In sharp contrast, hardly any macrophages were present in the same compartments in depleted mice, and no giant cells were observed. Quantification of the F4/80 immunostaining confirms this observation, as the F4/80 signal is virtually absent in depleted mice, in both the capsule and the ingrowth (Fig. 1B). These data indicate that AP20187-induced depletion of macrophages in MaFIA mice during the FBR was indeed successful and effective (>95%).
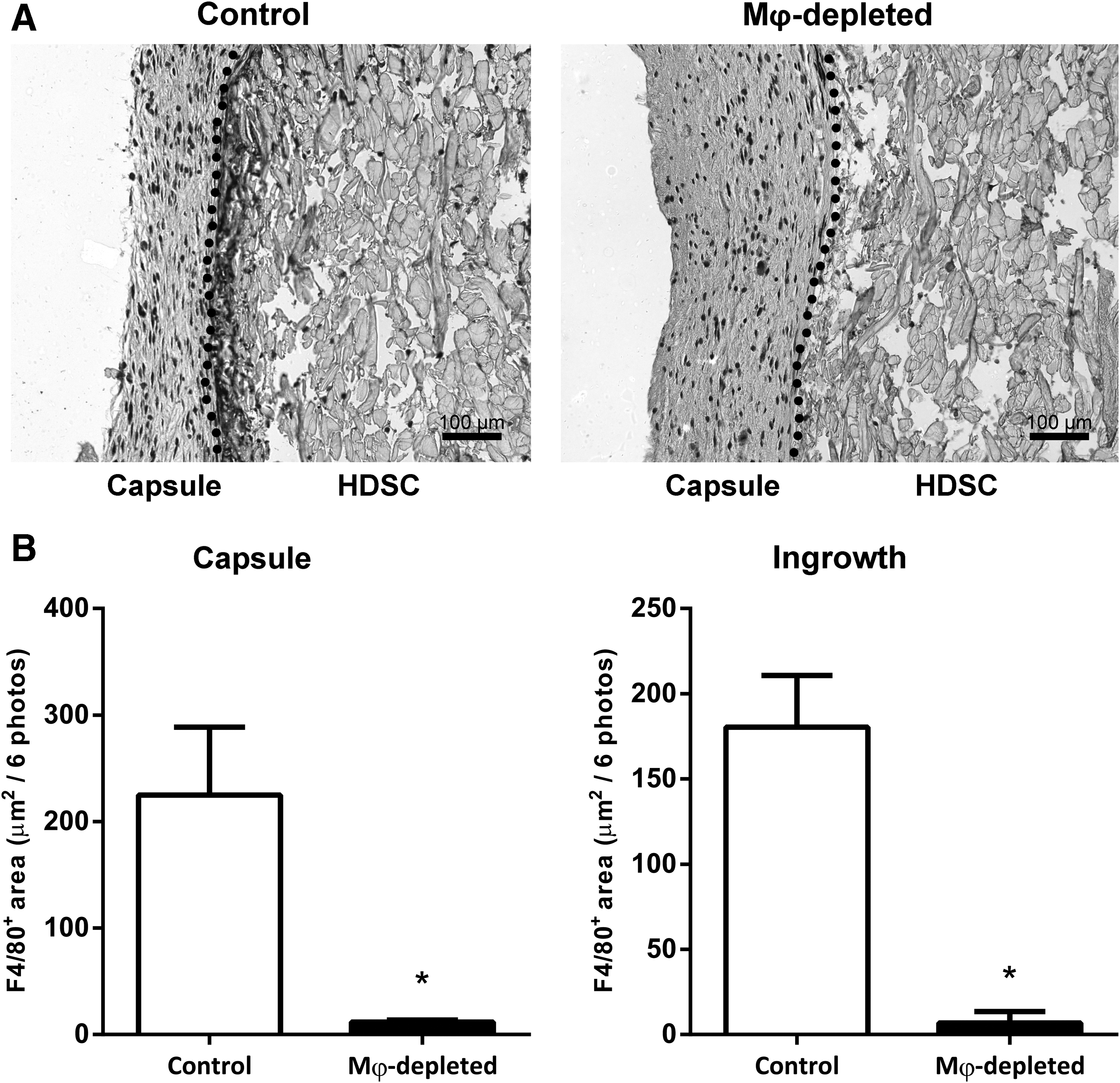
Immunohistochemical detection of F4/80 in HDSC scaffolds in nondepleted and Mφ-depleted mice
In sham-operated animals, no indication of inflammation or fibrosis was observed at the sites of mock implantation. In a qualitative comparison between the FBR to HDSC in nondepleted MaFIA mice (diluent-treated controls), and the FBR to HDSC in C57BL/6 mice (MaFIA background strain), no differences were found.
Side effects of depletion
During our experiment, the animals were weighed daily and blood was drawn before implantation and during explantation to assess the influence of macrophage depletion on body weight and several blood parameters (Fig. 2). Based on previous research26,27 (Sandra H. Burnett, pers. comm.), weight loss of around 20%, anemia, and peritoneal adhesions were to be expected in this model.
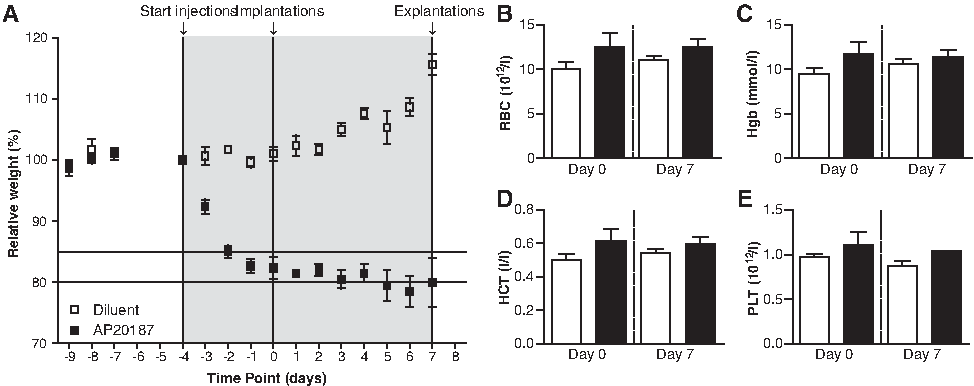
Effect of diluent (white squares and bars) and AP20187 (black squares and bars) treatment on body weight
Indeed, the body weight of AP20187-treated mice dropped rapidly after initiation of AP20187 treatment (Fig. 2A). After 4 days, the body weight of these mice stabilized somewhat at around 80% of their original weight. In contrast, the nondepleted mice, that is, diluent-treated controls, steadily gained weight over the course of the experiment. In addition to the weight loss, anemia was expected to occur after depletion of macrophages. In recent years, evidence is accumulating that macrophages play a pivotal role in iron recycling and erythroid proliferation and differentiation, thereby contributing to the maintenance of red blood cell levels.32–34 However, no detrimental effect of the depletion of macrophages on red blood cell counts, platelet counts, hemoglobin, and hematocrit levels were observed (Fig. 2B–E).
Peritoneal adhesions were not observed during the terminal surgery, where discs and organs were excised for further analysis. Macrophage depletion did induce splenomegaly, disrupted the normal cellular organization of the spleen, and induced the presence of megakaryocytes (Fig. 3A), as has been described before. 26 A trend toward enlarged livers was observed (p = 0.05) and the glycogen granules that are normally present in livers are lost upon macrophage depletion (Fig. 3B). Furthermore, interstitial, perivascular, and periductal inflammatory ingrowth was observed.

Effect of diluent and AP20187 treatment on spleen
In set of control experiments, AP20187 was administered to nontransgenic C57BL/6 mice. Since these mice lack the receptor for AP20187, weight loss, peritoneal adhesions, splenomegaly, and macrophage depletion were not observed (data not shown).
Effects of macrophage depletion on the FBR
The effects of macrophage depletion on the FBR were initially studied histologically. In macrophage-depleted mice, larger capsules were developed in comparison to nondepleted mice, that is, diluent-treated controls (Fig. 4A, quantified in Fig. 4B left panel). Furthermore, the capsules in macrophage-depleted mice contained less fibroblast-like cells (Fig. 4B middle panel), suggesting that fibroblasts in capsules of macrophage-depleted mice exhibit higher extracellular matrix production. In nondepleted mice a large number of inflammatory cells were present within the HDSC discs, forming a solid mass of ingrowth. In sharp contrast, HDSC discs in macrophage-depleted mice contained only a few mononuclear leukocytes (Fig. 4B right panel) indicating that macrophages are indispensable for the generation of the inflammatory ingrowth during the FBR. Neutrophils were not readily observed in histological sections of either macrophage-depleted or diluent-treated control mice.

Histological analysis of the foreign body reaction to HDSC scaffolds at day 7
qPCR analysis
The expression of proinflammatory cytokines, chemokines, fibroblast-stimulating growth factors, collagens, MMPs, and tissue inhibitors of MMPs was analyzed in macrophage-depleted and nondepleted mice (diluent-treated controls) using qPCR (Fig. 5). The expression of the proinflammatory cytokines, IL1β and TNFα, was decreased in macrophage-depleted mice (Fig. 5A), whereas the expression of the proinflammatory chemokine MCP1 was increased (Fig. 5B). The expression of the proinflammatory chemokine MIP1α was similar in macrophage-depleted and nondepleted mice (Fig. 5B). The expression of fibroblast-stimulating growth factors, such as CTGF, EGF, FGF1, FGF2, and TGFα, was consistently upregulated or showed a trend toward upregulation in macrophage-depleted mice (Fig 5C). The expression of collagens, such as collagen I, III, IV, V, and VI, was consistently upregulated in macrophage-depleted mice (Fig. 5D). The expression of TIMP1 and TIMP2 was increased or showed a trend toward an increase (Fig. 5E). The expression of MMP2 was increased in macrophage-depleted mice, whereas MMP13 was decreased.

Gene expression analysis of proinflammatory cytokines
Discussion
Macrophages are, arguably, the most important cells during the FBR.5–8 They generate and maintain a proinflammatory environment in and around the introduced biomaterial. Furthermore, they contribute to the removal of the foreign material by secreting factors that degrade the biomaterial extracellularly and/or by phagocytosis of small (predegraded) biomaterial particles. In addition, extensive crosstalk between macrophages and fibroblasts is likely to occur, especially in the capsule around the biomaterial. During this crosstalk, macrophages may instruct fibroblasts through secretion of soluble factors, which may be crucial for the generation of a well-organized collagen-rich fibrous capsule.
In this study, we challenged the dogma that macrophages are the most important cells during the FBR by studying the FBR after macrophage depletion in MaFIA mice. Depletion was very efficient (>95%), which is an improvement over previous studies on the FBR after macrophage depletion, using CD11b-DTR mice (50% depletion), 35 or clodronate liposomes (36–98% depletion). 36 Interestingly, it is also slightly more efficient than a previous study that used the MaFIA mouse model to study the FBR to solid alginate spheres in the peritoneum (85% depletion). 37 It is noteworthy that DC are also expected to be depleted in the presented mouse model, and a large degree of (potential) functional overlap exists between DC and macrophages. Although incompletely understood, DC are expected to play an important role in the FBR.38,39 Obviously, a more specific and refined depletion model is required to discriminate between DC and macrophage functions in the FBR.
We found that in macrophage-depleted mice, no inflammatory cells migrate toward (and into) the biomaterial and less proinflammatory cytokines were expressed. Others have found similar results when studying the FBR to alginate microspheres in the peritoneum after macrophage depletion. 37 Furthermore, we found that capsules were thicker, which coincided with increased expression of fibroblast-stimulating growth factors, collagens, and tissue inhibitors of metalloproteinases (TIMPs).
Since fibroblasts and neutrophils are both capable of secretion of proinflammatory mediators and phagocytosis,40–42 one could hypothesize that these cells may take over functions of macrophages inside the biomaterial. However, neither neutrophils nor fibroblasts were observed inside biomaterials in macrophage-depleted mice. This indicates that there is no redundancy of macrophage functionality inside biomaterials during the FBR, which emphasizes their importance in this context.
In contrast, macrophages appear to be dispensable for the generation of a well-organized fibrous capsule. In fact, capsules were bigger in macrophage-depleted mice, which coincided with increased expression of collagens, TIMPs, and fibroblast-stimulating growth factors. This suggests that macrophages actually dampen the fibrotic response of the fibroblasts during the FBR. Previously, in vitro experiments on (indirect) cocultures of macrophages and fibroblasts suggested that proinflammatory macrophages are able to instruct fibroblasts to adopt a proinflammatory and nonfibrotic phenotype through paracrine signaling. 17 These proinflammatory fibroblasts secrete decreased amounts of collagen type I, increased amounts of MMPs, and display an increase in MMP activity. 16 This may explain why the size of capsules is reduced in the presence of macrophages. The increased expression of fibroblast-stimulating growth factors suggests that the self-organizing capacity of fibroblasts in macrophage-depleted mice may be mediated through paracrine signaling.
The striking difference in expression pattern between MMP2 and MMP13 may be explained by the substrate specificity of these enzymes. MMP2 is a gelatinase, which is likely to be more efficient in degrading the relatively immature collagenous structures found in the developing capsule. It has been suggested that MMP2 is more suitable for tightly regulated cell-directed focal proteolysis required in tissue homeostasis. 43 In this case, MMP2 may be more suitable for the subtle remodeling of the fibrous capsule in a microenvironment where structural integrity is crucial. MMP13 is a collagenase, which is able to degrade relatively mature collagenous structures, such as the densely crosslinked implanted dermal sheep collagen. In this case, MMP13 may be more suitable for the blunt destruction of large foreign collagenous structures such as HDSC. We, therefore, speculate that the increased expression of MMP2 in macrophage-depleted mice is secreted by fibroblasts in an attempt to remodel the fibrous capsule, whereas the increased expression of MMP13 in nondepleted mice is secreted by macrophages in an attempt to degrade the implanted biomaterial. Indeed, in rodent models of liver fibrosis, scar-associated macrophages were demonstrated to be the main source of MMP13, which was shown to be important for the spontaneous resolution of experimental liver fibrosis.44,45
Conclusions
In conclusion, we have successfully studied the FBR in a mouse model for conditional macrophage ablation. Our data indicate that macrophages are, as expected, crucial for the generation and orchestration of proinflammatory ingrowth into biomaterials. Somewhat surprisingly, macrophages appeared to be dispensible for the generation of a fibrous capsule around biomaterials. Paracrine signaling between fibroblasts may play a pivotal role during this process, and the presence of macrophages may—in the context of the FBR—actually dampen the fibrogenic response.
Footnotes
Acknowledgments
This work was supported by The Netherlands Institute for Regenerative Medicine (NIRM, Grant No. FES0908), and the Dutch Kidney Foundation.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.