
Abstract
Engineered hypertrophic cartilage (HC) represents an attractive bone substitute material, capable to induce bone formation by endochondral ossification. Since bone formation by HC depends on factors released from the extracellular matrix, in this study, we hypothesized that HC seeding with monocytes committed to osteoclastogenesis could enhance its remodeling, improve chemotaxis of skeletal and vascular cells, and consequently enhance bone formation. This would be particularly relevant for devitalized HC, which currently exhibits only limited osteoinductivity. Living or devitalized HC engineered from human bone marrow-derived mesenchymal stromal cells (MSCs) was seeded or not with human monocytes in the presence of macrophage colony-stimulating factor and RANK-ligand, cultured for up to 15 days, or implanted ectopically in nude mice. Monocytes seeded on devitalized, but not living, HC induced its in vitro resorption, resulting in 30-fold higher release and 2.7-fold lower content of glycosaminoglycans compared with unseeded samples. In vitro, supernatants from monocyte-seeded devitalized HC attracted more monocytes compared with unseeded samples, but did not enhance chemotaxis of MSCs or human umbilical vein endothelial cells. In vivo, however, neither remodeling nor invasion by osteoclasts, endothelial cells, and mouse MSCs were significantly affected by the seeding with monocytes. Finally, in vitro priming of living or devitalized HC by monocytes did not enhance their bone-forming capacity. Further investigations should test the proposed approach on HC engineered to prevent rapid degradation and support osteoclastogenesis, or identify alternative strategies to enhance engineered HC remodeling and bone-forming capacity.
Introduction
B
To create a bone substitute inspired by natural healing, it is possible to in vitro engineer HC from adult bone marrow-derived mesenchymal stromal cells (MSCs), which has been reported to undergo EO in vivo. 3 During HC remodeling, digestion of the HC extracellular matrix, predominantly by osteoclast-derived matrix metalloproteinase (MMP)-9 and chondrocyte-derived MMP-13,4,5 releases factors playing a pivotal role in EO, including VEGF (vascular endothelial growth factor) and BMPs (bone morphogenetic proteins).6–8 This release prompts chemotaxis of endothelial cells, osteoclasts, and MSCs and ultimately guides bone formation. The described mechanism can also be recapitulated by using engineered and devitalized HC matrices, thereby offering the possibility to avoid the logistic/standardization challenges of prolonged autologous cell culture or the immunogenicity issue of living allogeneic HC. 9 However, the bone formation efficiency is reduced compared to living HC, an issue which has previously led us to investigate a possible “reactivation” through addition of stromal vascular fraction (SVF) cells derived from human adipose tissue. 10 Although SVF contains endothelial cells, MSCs, and monocytes, which are precursors to osteoclasts, the specific role of additional monocytes/osteoclasts on HC remodeling remained elusive. In particular, it was not clear whether monocytes attracted as part of the initial immune reaction 11 were sufficient or whether additional monocytes on the surface of HC, possibly primed to become osteoclasts, could enhance the release of cytokines and thus promote osteoinductivity.
In the present study, we addressed whether addition of osteoclastogenically primed monocytes would increase (i) the remodeling of engineered HC, (ii) its chemotaxis of endogenous skeletal cells, and ultimately (iii) its induction of bone formation. To test this hypothesis, we sorted monocytes from peripheral blood12,13 and primed them to osteoclastogenesis through stimulation with RANKL and macrophage colony-stimulating factor (M-CSF).14,15 Then we analyzed both the isolated effect of the primed monocytes on engineered living or devitalized HC in vitro and the efficiency of bone formation in vivo.
Materials and Methods
Generation of HC
HC was generated as described previously.
3
Bone marrow MSCs were extracted from five healthy human donors (male, 33.8 ± 6.8 years) after informed consent (EKBB reference 78/07, Switzerland) and expanded using complete medium (α-MEM, 10% v/v fetal bovine serum [FBS], 1 mM sodium pyruvate, 10 mM HEPES, and 1% v/v penicillin–streptomycin–glutamine; all from Gibco, Basel, Switzerland) with 5 ng/mL fibroblast growth factor-2 (R&D Systems, Wiesbaden, Germany) up to four passages. Cells were detached using 0.05% Trypsin–ethylenediaminetetraacetic acid (EDTA; Gibco), collected, and resuspended in chondrogenic medium consisting of serum-free medium (Dulbecco's modified Eagle's medium [DMEM], 1.25 mg/mL human serum albumin, 1 mM sodium pyruvate, 10 mM HEPES, 1% v/v penicillin–streptomycin–glutamine, and 1% v/v ITS-A; all from Gibco) together with 10 ng/mL tranforming growth factor-β1 (R&D Systems, Minneapolis, MN), 10−7 M dexamethasone, and 0.1 mM ascorbate-2-phosphate (Sigma-Aldrich, Buchs, Switzerland). In the first experiment, a transwell system (0.4 μm membrane pores 24-well-plate; Corning, Root, Switzerland) was used. All subsequent experiments were performed with 1.5 mL screwcap tubes (Sarstedt, Sevelen, Switzerland). 5 × 105 MSCs were centrifuged at 400 g for 5 min to form cell aggregates and the resulting pellets incubated at 37°C and 5% CO2. Medium was changed twice per week. After 3 weeks with chondrogenic medium, hypertrophic medium (serum-free medium with 0.05 μM
Devitalization of HC
Samples were subjected to freeze/thaw devitalization as described previously. 9 In brief, samples were transferred to 15 mL plastic tubes, frozen in liquid nitrogen for 10 min, and then thawed in a water bath at 37°C for 10 min. This cycle was repeated three times, and the samples were finally rinsed with bidistilled water.
Isolation of CD14 positive monocytes
Peripheral blood buffy coats from five healthy donors were used, and mononuclear cells were separated by gradient centrifugation (Ficoll, Histopaque 1077; Sigma-Aldrich) as previously described. 16 CD14+ monocytes were sorted using anti-CD14-coated magnetic beads (Miltenyi Biotec, Auburn, CA) according to the manufacturer's instructions.
Cocultures
CD14+ monocytes were resuspended in coculture medium (α-MEM, 1 mM sodium pyruvate, 10 mM HEPES, 1% penicillin–streptomycin–glutamine; all from Gibco) with 25 ng/mL M-CSF and 50 ng/mL RANK-ligand (R&D Systems, Minneapolis, MN). Living or devitalized hypertrophic matrix was incubated with 1–5 million monocytes on an orbital shaker for 24 h at 37°C. Seeding efficiency was analyzed by incubation with 0.05 mg/mL tetrazolium salt (MTT; Sigma-Aldrich) in phenol-free medium (DMEM; Gibco) for 4 h at 37°C. MTT was then dissolved with DMSO (Sigma-Aldrich) and absorption measured at 575 nm. After seeding, cocultures were gently transferred to conical 96-well plates (Corning). Unseeded samples were kept in the same medium as cocultures. Medium changes were performed twice per week and the supernatants from each medium change were frozen at −20°C for later analysis.
Glycosaminoglycan and DNA content
For the measurement of glycosaminoglycans (GAGs) and DNA, coculture samples were digested overnight with Proteinase K solution (1 mg/mL Proteinase K in 50 mM Tris pH 7.6, 1 mM EDTA, 1 mM Iodoacetamide, and 10 μg/mL Pepstatin; all from Sigma-Aldrich). Supernatants were used without digestion. GAG measurements were performed as described previously. 17 In brief, GAG was precipitated with dimethylmethylene blue (DMMB), centrifuged, and redissolved. Absorption was measured at 656 nm, and GAG concentrations were calculated using a standard curve. DNA measurements were performed with the CyQuant kit (Thermo Fisher Scientific, Zug, Switzerland) according to the manufacturer's protocol.
Chemotactic experiments
Chemotaxis of human MSCs, human umbilical vein endothelial cells (HUVEC), or human CD14+ monocytes toward supernatants from cocultures was analyzed. MSCs of three donors and monocytes from two buffy coats were prepared as described above. HUVEC expanded for four passages were used as provided (Prof. Anna Marsano). Ninety-six-transwell plates (HTS transwell, 8.0 μm pore size, polyester membrane; Corning) were precoated with 1% w/v gelatin and 5 × 104 of MSCs, monocytes, or HUVEC and were placed in fresh coculture medium in the top chambers. Supernatants were diluted 1:2 in fresh coculture medium and distributed in the bottom chambers. Fresh coculture medium was used as a negative control, and coculture medium with 10% v/v FBS was used as a positive control. Plates were incubated for 24 h at 37°C, then centrifuged at 400 g for 2 min, and bottom chambers were incubated with 1 μM Calcein AM for 15 min at 37°C. Fluorescence of the bottom chambers was read at 480 nm excitation and 520 nm emission.
Implantations
Seeded or unseeded samples after 24 h of incubation in coculture medium were used for implantations. At least two replicates were implanted, and each replicate was implanted without the use of gels or scaffolds. All animal procedures were reviewed and approved by the Swiss Federal Veterinary Office (Permit No. BS 1797). Female 6- to 8-week-old nude mice were anaesthetized using 2% isoflurane in 1 L/min O2. The skin was disinfected and cut 8 mm along the dorsal midline. Subcutaneous pockets were prepared by blunt dissection on both sides, and one sample per pocket was implanted. The wounds were closed with surgical staples, and the animals were allowed to recover under red light. Animals were monitored daily, and staples were removed after 7–10 days. Euthanasia was performed in a CO2 chamber after 3 or 8 weeks, and samples were fixed overnight in 1.5% v/v paraformaldehyde before further analysis.
Microtomography
Microtomography was performed using a tungsten X-ray source at 70 kV and 260 μA with a 0.5 mm aluminium filter (Nanotom M; GE). Transmission images were acquired for 360° with an incremental step size of 0.25°. Volumes were reconstructed using a modified Feldkamp algorithm (software supplied by manufacturer) at a voxel size of 2.5–3 μm. Thresholding, segmentation, and 3D measurements were performed using the ImageJ 18 software with the BoneJ 19 and 3D Shape 20 additions. The results are shown as ratios of mineralized volume to tissue volume, since the procedure does not allow to reliably distinguishing mineralized regions from frank bone tissue.
Histology
Paraffin-embedded samples were sectioned at 5 μm thickness. Safranin-O, Masson's tri-chrome, and tartrate-resistant acid phosphatase (TRAP) stainings were performed as described previously. 3 Primary antibodies for collagen type I (ab6308), collagen type II (ab34712), collagen type X (ab49945), bone sialoprotein (ab52128), MMP-9 (ab38898), MMP-13 (ab39012), CD31 (ab28364), and Sca-1 (ab51317; all from Abcam, United Kingdom) were used. After washing and incubation with an appropriate biotinylated secondary antibody (Dako, Denmark), the stainings were developed with the Vectastain ABC kit (Vector Laboratories) according to the manufacturer's protocols.
Quantifications
Samples were completely sectioned, and four to six sections per sample spaced at regular intervals were quantified. Only cells inside or attached to the surface of the implanted HC matrix were counted, and the total sample area was considered for counting. For osteoclasts, TRAP-positive multinucleated cells were counted. For endothelial cells, CD31-positive cells were counted.
Statistical analysis
Data were analyzed by t-test and analysis of variance using the GraphPad Prism v2.2 software. Results were considered significant if p < 0.05. Average values and standard deviations are reported.
Results
Generation of HC
After 5 weeks of in vitro culture, both disks and pellets of hypertrophic cartilaginous matrix showed a typical cellular morphology and positive staining for molecular markers of HC (collagen I/II/X, BSP, MMP-13) as previously published 3 (Supplementary Fig. S1A, B; Supplementary Data are available online at www.liebertpub.com/tea). Given that transwell cultures randomly formed disks or pellets, standardized pellet cultures were used for all subsequent experiments.
In vitro remodeling of HC
Cocultures were initiated by seeding 1 million monocytes (determined as sufficient by titration, see Supplementary Fig. S2A) on living or devitalized HC. GAG was detected in supernatants of all groups throughout the observation time (15 days), with the highest concentrations observed after 3 days (Fig. 1A). Addition of monocytes to devitalized HC led to a 30-fold higher release of GAG, a strongly reduced volume with loss of Safranin-O positivity (Fig. 1B) and a 2.7-fold lower amount of GAG contained in the matrix, compared with unseeded devitalized samples (Fig. 1C). No TRAP-positive multinucleated cells were present on the seeded devitalized matrix, though few TRAP-positive monocytes were found inside the matrix after 7 days (Supplementary Fig. S2B). Addition of monocytes to living samples did not significantly change their release or content of GAG or their histological appearance. However, TRAP-positive multinucleated cells were found within the living matrix after 15 days (Fig. 1D). Monocytes thus appeared to rapidly digest devitalized samples, whereas they transformed into invading osteoclastic cells when seeded on living samples.
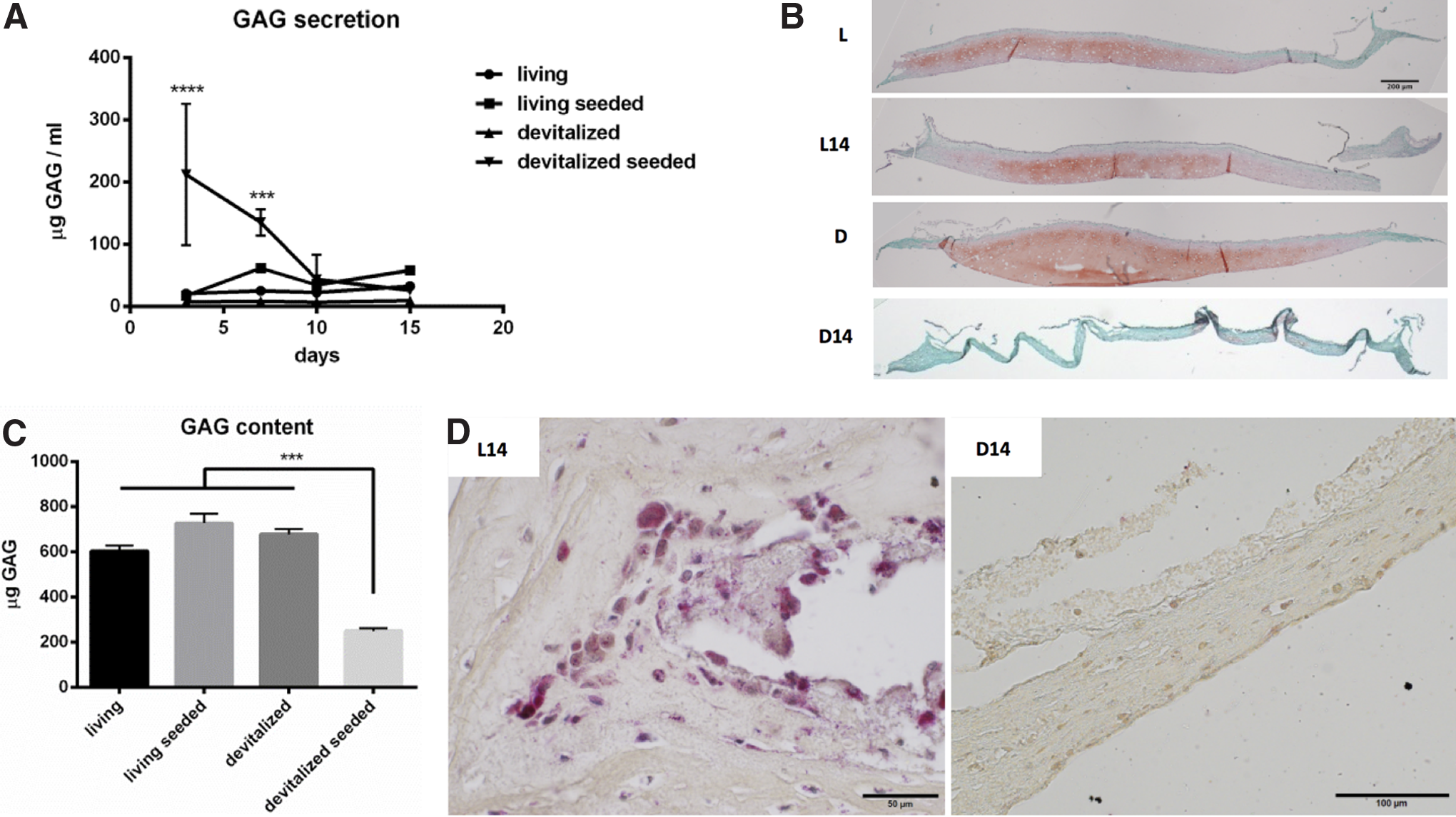
In vitro remodeling of hypertrophic cartilage.
In vitro chemotaxis of osteogenic progenitors
To investigate the effect of the factors released during monocyte/osteoclast digestion, chemotactic assays for human MSCs, HUVEC, and human CD14+ monocytes were performed with supernatants from the first 3 days of culture. No chemotaxis of MSCs or HUVEC toward any supernatant of the experimental groups was found (Fig. 2). Instead, supernatants of seeded devitalized samples induced a significantly increased chemotaxis of monocytes compared with unseeded ones (1.6-fold), reaching the level of corresponding living HC. These results indicated that cocultures primarily attracted monocytes and that devitalized samples could only attract monocytes when seeded with monocytes.

Chemotaxis using supernatants from cocultures after 3 days. Values are fold of negative controls using fresh coculture medium. Human MSCs and HUVEC do not show any chemotaxis. Human CD14 monocytes show significantly decreased chemotaxis toward devitalized samples (*p < 0.05;**p < 0.01). HUVEC, human umbilical vein endothelial cells; MSCs, mesenchymal stromal cells.
Early in vivo remodeling and presence of endogenous osteogenic progenitors
Next, we investigated the effect of monocyte addition on in vivo HC matrix remodeling and chemotaxis of endogenous progenitors as early as 3 weeks after subcutaneous implantation. At this time, devitalized implants regardless of seeding modality appeared devoid of GAG and with a sparse fibrotic matrix surrounded by denser tissue (Fig. 3A). Living implants both with and without monocytes displayed a very faint Safranin-O staining, suggesting loss of GAG. To assess in vivo chemotaxis, the presence of endogenous osteoclasts, endothelial, and MSC-like cells was analyzed histologically. The number of osteoclasts was not significantly influenced by addition of monocytes. Devitalized implants contained 7.6-fold less osteoclasts than living implants (Fig. 3B). Endothelial cells showed a high variability without a significant trend (Fig. 3C) and did not organize into vessel-like structures. Sca-1-positive mouse MSCs were so rare that meaningful quantitative analysis of these cells was not feasible. It thus appeared that neither matrix remodeling nor recruitment of endogenous osteoclasts, vascular, and MSCs was affected by addition of monocytes.
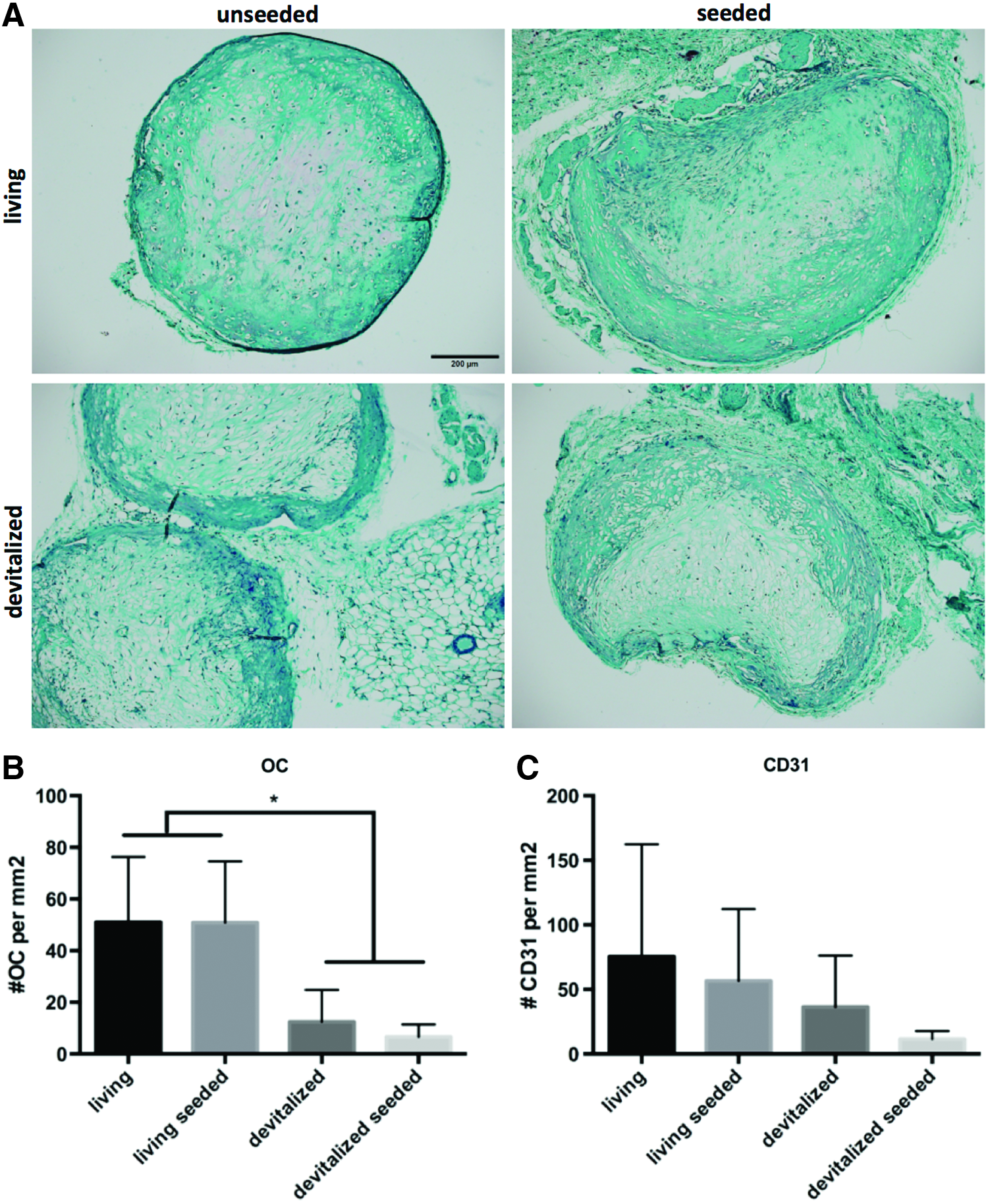
Remodeling and chemotaxis after 3 weeks of subcutaneous implantation in nude mice.
In vivo bone and mineralized tissue formation
To investigate the efficiency of bone formation, samples were analyzed 8 weeks after implantation. The mineralized fraction of total tissue volume was quantified by using microtomography and by defining a threshold for bone and mineralized tissue (Fig. 4A). Addition of monocytes did not significantly change mineralized volumes of either living or devitalized implants. Devitalized implants displayed up to threefold less mineralized volume than living implants.

Bone formation after 8 weeks of subcutaneous implantation.
Histological analysis revealed that devitalized implants regardless of monocyte seeding contained either cell-free fibrotic matrix or cell-rich dense matrix, the latter most likely corresponding to the mineralized tissue found by microtomography. Conversely, both seeded and unseeded living implants displayed frank bone tissue with bone marrow and a peripheral bone lamina. Interestingly, seeded implants more frequently showed red-stained elastin inside the osteoid, suggesting a more mature bone (Fig. 4B).
Thus, it appeared that addition of monocytes did not improve bone formation efficiency in devitalized samples and only minimally affected bone maturation of living implants.
Discussion
In this present study, we demonstrated that addition of osteoclastogenically primed monocytes increased the remodeling of engineered devitalized but not of living HC and led to the release of factors that further attracted monocytes in vitro. However, this did not result in enhanced chemotaxis of endogenous skeletal cells or in more efficient bone formation in vivo by either devitalized or living HC.
We previously showed that addition of adipose-derived SVF cells, a highly heterogenous cell mixture containing endothelial, stromal, and osteoclastic progrenitors led to improved bone formation in devitalized HC. 10 Although previous work explored the effect of additional endothelial cells and MSCs on HC, 21 the possible role of additional monocyte/osteoclast addition has not yet been investigated. Monocytes/osteoclasts could mediate the release of factors from the engineered HC matrix, in line with their physiological role in EO, and thus enhance its bone formation efficiency. To test this hypothesis, we used a pure population of monocytes by processing peripheral human blood, which would also be compatible with an intraoperative setting. In addition, we used freeze/thaw cycles as a simple and effective method to devitalize engineered HC, even though this may have reduced the content of cytokines and growth factors in the matrix when compared to a more targeted apoptosis of hypertrophic chondrocytes. 9
The seeded monocytes invaded both living and devitalized samples in vitro but resulted in markedly different outcomes. Monocytes formed multinucleated osteoclasts on living samples in vitro, without being associated with extensive matrix digestion or increase in the total density of osteoclasts following in vivo implantation. Instead, monocytes seeded on devitalized samples induced in vitro a dramatic loss of GAG and volume upon coculture. TRAP-positive monocytes 22 invaded devitalized samples at early time points without forming multinucleated osteoclasts and simultaneously attracted further monocytes, reminiscent of an immunologic response to a foreign body.23,24 Together with the fibrous appearance and lack of osteoclasts in devitalized implants, these findings suggest that monocytes did not find the appropriate signals/substrate to form osteoclasts on devitalized HC and rather initiated an inflammatory reaction. Indeed the higher release of GAG from devitalized samples may have introduced a higher amount of hyaluronic acid into the surrounding tissue, which is known for its proinflammatory role. 25 An exploration using array analysis for chemotactic factors could contribute to identify processes and molecular mechanisms mediating the observed immune response.
Unexpectedly, neither MSCs nor HUVEC were attracted by any of the culture supernatants, and they were also not affected by the addition of monocytes. Although endothelial cells were present after 3 weeks in vivo, they were overall scarce and their frequency was highly variable between replicates, possibly because vascularization was not yet organized at this early time during remodeling. Also, MSCs appeared to be mostly absent from both living and devitalized HC at this time, and their recruitment was not enhanced by the presence of monocytes. The absence of increased recruitment of MSCs and macrophages could be related to the features of the model, which lacks the relevant signals of an orthotopic environment and of an immunocompetent recipient. However, it should be highlighted that previous studies using the same model have shown the active involvement of resident mouse cells for the remodeling into bone tissue of living3,26 or even devitalized 9 HC. An alternative explanation could be based on the fact that the degradation of the devitalized matrix may happen too rapidly, before the different target cells would be available at the implant site. Thus, a burst release of factors may be unable to meet the need of bone regeneration, where a series of molecular and cellular events needs to be carefully orchestrated.
In conclusion, addition of monocytes did not enhance bone formation of living or devitalized HC. However, our experimental model allowed us to analyze monocyte-mediated digestion of HC. We found a more rapid degradation and lack of osteoclasts in devitalized HC, which likely resulted in a lack of bone formation. To address the challenge of rapid degradation observed in the current study when using devitalized HC, the HC matrix could be crosslinked or otherwise modified after devitalization. As an alternative, the matrix could be tuned through the use of customized cell lines, 9 for example, by enriching it with costimulatory factors for osteoclasts 27 or factors to prevent the early invasion of monocytes28,29 in immunocompetent animals.
Footnotes
Acknowledgments
The AO Foundation (AO S-11-13P) and the Swiss National Science Foundation (Grant No. 310030_133110) supported this study. We thank Prof. Anna Marsano, Dr. Sarah Dimeloe, and Dr. Marco Fischer for their technical support.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.