
Abstract
Extracellular matrix (ECM) maintains a reservoir of bioactive growth factors and matricellular proteins that provide bioinductive effects on local cells that influence phenotype and behaviors. Bioactive acellular ECM scaffolds can be used therapeutically to stimulate adaptive tissue repair. Fibroblast growth factor-2 (FGF-2) attenuates transforming growth factor-β1 (TGF-β1)-mediated cardiac fibrosis. Heparin glycosaminoglycan can influence FGF-2 bioactivity and could be leveraged to enhance tissue engineering strategies. We explored the effects of heparin on FGF-2 enhancement of bioactive ECM scaffold biomaterials for its antifibrotic effect on attenuating human cardiac myofibroblast activation. Increasing heparin concentration at a fixed concentration of FGF-2 markedly increased the amount of FGF-2 retained and eluted by ECM scaffolds. To explore synergistic bioinductive effects of heparin and FGF-2, collagen gel contraction assay using human cardiac myofibroblasts was performed in vitro. Myofibroblast activation was induced by profibrotic cytokine, TGF-β1. FGF-2 and heparin in combination reduced human cardiac myofibroblast-mediated collagen gel contraction to a greater extent than FGF-2 alone. These observations were confirmed for both human atrial and human ventricular cardiac fibroblasts. Cell death was not different between groups. In summary, heparin is an effective adjuvant to enhance FGF-2 loading and elution of acellular ECM scaffold biomaterials. Heparin increases the bioactive effects of FGF-2 in attenuating human cardiac myofibroblast activation in response to profibrotic TGF-β1. These data may inform tissue engineering strategies for myocardial repair to prevent fibrosis.
Introduction
T
Heparin is a negatively charged glycosaminoglycan with affinity for ECM protein components such as FGF-2. Heparin, when bound to FGF-2, has been shown to prevent proteolytic degradation and thermal or pH-dependent denaturation of FGF-2, as well as facilitate FGF-2 binding to cell membrane receptors.7–11 Furthermore, heparin administration alongside FGF-2 in a coacervate vehicle has been shown to reduce cardiac fibrosis and myocardial infarction (MI)-induced inflammation beyond FGF-2 alone. 12 We therefore sought to determine whether heparin could facilitate FGF-2 enhancement of an acellular ECM scaffold biomaterial (SIS-ECM). We also explored the bioactive effects of heparin augmentation on FGF-2-mediated human cardiac myofibroblast inhibition.
Materials and Methods
SIS-ECM biomaterial enhancement
Porcine SIS-ECM (CorMatrix®-ECM®; CorMatrix Cardiovascular, Inc., GA) was obtained. One-cm diameter pieces were prepared by soaking them in 0.9% NaCl with or without varying concentrations of FGF-2 (Invitrogen, Waltham, MA) and 10,000-units/mL heparin (PPC, Richmond Hill, ON) for 2 h at room temperature.
FGF-2 elution from SIS-ECM
FGF-2-enhanced SIS-ECM biomaterial was incubated in a serum-free cell culture medium (Iscove's Modified Dulbecco's Medium [IMDM], 1% penicillin-streptomycin) for 1, 3, or 24 h. Eluate was transferred to a 1.5 mL Eppendorf tube and centrifuged at 10,000 rpm for 10 min at 4°C. Supernatants were diluted to appropriate ranges and FGF-2 was measured by enzyme-linked immunosorbent assay (ELISA).
SIS-ECM biomaterial homogenization
In preparation for FGF-2 quantification, SIS-ECM biomaterial samples were minced and incubated in 300 μL of lysis buffer solution prepared from 10 × radioimmunoprecipitation assay buffer (Millipore, Temecula, LA) and one tablet of protease inhibitors (Roche, Indianapolis, IN) for 10 min at 4°C. Samples were then homogenized using a tissue homogenizer. Homogenates were centrifuged at 10,000 rpm for 10 min at 4°C. Thirty microliters of the supernatant was collected and added to 10 μL of 4 × NuPage LDS sample buffer (Invitrogen, Waltham, MA).
Western blot
FGF-2 bound to SIS-ECM biomaterial was quantified by Western blot. SIS-ECM biomaterial homogenate samples were heated at 95°C for 10 min and 20 μL of each sample was run on a 12% SDS-PAGE gel at 200 V for 1 h and then transferred onto a nitrocellulose membrane (Millipore, Temecula, LA) at 100 V for 1 h. Nitrocellulose membranes were blocked in 5% skim milk in Tris-buffered saline +0.05% Tween (TBST) for 1 h at room temperature and incubated with primary FGF-2 rabbit polyclonal IgG antibody (1:1000 dilution; Santa Cruz Biotechnology, Dallas, TX) overnight at 4°C. Membranes were then washed (3 × ) with TBST and incubated with secondary IgG rabbit horseradish peroxidase-linked antibodies (1:2500 dilution; Sigma Aldrich, St. Louis, MO) for 1 h in 10 mL of blocking solution. Membranes were washed with TBST thrice and incubated for 1 min in luminol and peroxide (SuperSignal West Femto Maximum Sensitivity Substrate; Thermo Scientific, Waltham, MA). The chemiluminescent signal was measured using a gel imaging system (ImageQuant LAS 4000–General Electric, Toronto, ON). The optical density of each band was quantified relative to a known concentration of FGF-2 using Li-Cor Western Blot Analysis Software (Li-Cor, Lincoln, NE) and ImageJ.
Enzyme-linked immunosorbent assay
FGF-2 bound to SIS-ECM biomaterial was measured by ELISA. SIS-ECM biomaterial homogenate was tested using a human FGF-2 ELISA kit (Abcam, Cambridge, MA) in accordance with the manufacturer's specifications. Ninety-six-well microplate colorimetric absorbance values were measured using Gen5 Microplate Reader software, using Synergy Mx Scanner at 450 nm wavelength.
Cardiac fibroblast cell isolation
Following consent, right atrial appendage or left ventricular tissue samples were obtained from patients undergoing cardiac surgery at the Foothills Medical Centre (Calgary, Alberta). Experiments involving human tissue usage were approved by the Conjoint Health Research Ethics Board at the University of Calgary and conform to the Declaration of Helsinki. Biopsy samples were rinsed in phosphate-buffered saline (Lonza, Walkersville, MD) to remove blood and visible fat was removed as much as possible. Tissues were minced and cultured as explants on 100 mm Petri dishes (Falcon, Corning, NY) with IMDM supplemented with 10% fetal bovine serum (Gibco Life Technologies, Burlington, ON) and 5% penicillin–streptomycin (Life Technologies, Burlington, ON). Cells were incubated at 37°C at 5% CO2. The cell culture medium was replaced every 2 days and passaged with 4 mL 0.25% Trypsin-EDTA when cells reached 85% confluency. Only cells arising from the first explant were used for experiments.
3D collagen gel contraction assay
As previously described, three-dimensional (3D) collagen gel contraction assay13,14 was performed. Five hundred microliters of IMDM alone (serum-free medium control) or containing 10 ng/mL human recombinant transforming growth factor-β1 (TGF-β1; Gibco Life Technologies, Burlington, ON) was added into each well, with or without FGF-2. To determine whether heparin affected the myofibroblast activity in the presence of FGF-2, heparin was added in addition to FGF-2 in a fourth group. Images of the collagen matrix surface area were obtained 48 h from the time of release.
Annexin V/propidium iodide cell apoptosis assay
As previously described, 14 cells isolated from collagen matrices were resuspended in 300 μL of cold binding buffer and stained with annexin V and propidium iodide (BD Pharmingen, Becton Dickinson, San Jose, CA), according to the manufacturer's instructions. Percentages of annexin V and propidium iodide-positive cells were measured using flow cytometry.
Statistics
Multiple comparisons were performed using Bonferroni's multiple comparisons test, p < 0.05, for Western blot and ELISA experiments. Unpaired t-tests were performed for 3D collagen matrix gel contraction experiments.
Results
Heparin augments FGF-2 enhancement of SIS-ECM biomaterial
SIS-ECM biomaterial was treated with increasing concentrations of FGF-2 to determine the optimal dose for FGF-2 enhancement. Increasing the concentration of FGF-2 within the enhancement solution resulted in increased FGF-2 loading into the SIS-ECM biomaterial (Fig. 1A, B). We next determined if an adjuvant could be used to augment FGF-2 loading into the SIS-ECM biomaterial. The lowest concentration of FGF-2 (2 μg/mL) shown to enhance FGF-2 levels in the biomaterial was combined with increasing concentrations of heparin. Increasing heparin concentration at a fixed FGF-2 dose resulted in increased FGF-2 loading into the SIS-ECM biomaterial (relative density values: 1.98 at 0 mg/mL heparin; 2.35 at 1 mg/mL heparin; 3.86 at 10 mg/mL heparin; and 4.37 at 50 mg/mL heparin; N = 1; Fig. 1A–C). We confirmed these observations in a larger sample using ELISA, which also demonstrated that heparin augmented FGF-2 enhancement of SIS-ECM in a dose-dependent manner (FGF-2 concentration values: 11,413 ± 493.3 ng/mL at 10 mg/mL heparin; and 17,880 ± 1113.6 ng/mL at 50 mg/mL heparin; N = 3, p < 0.05, Bonferroni's multiple comparison test; Fig. 1D–E).

Heparin does not limit FGF-2 elution from enhanced SIS-ECM biomaterial
Given that heparin increased FGF-2 loading of SIS-ECM biomaterial, we next explored if FGF-2 loaded into the SIS-ECM biomaterial was able to elute from the SIS-ECM biomaterial and provide bioactive effects in conditions that would mimic an in vivo setting. FGF-2-enhanced SIS-ECM biomaterial was incubated in a cell culture medium for 1, 3, and 24 h after which the concentration of FGF-2 was measured in the eluate (Fig. 2). FGF-2 was detected in the eluate of both FGF-2-enhanced SIS-ECM biomaterial and SIS-ECM biomaterial alone (Eluate FGF-2 concentration at 1-h time point: 72.96 ± 5.44 ng/mL for SIS-ECM biomaterial alone and 6854.67 ± 918.35 ng/mL for FGF-2-enhanced SIS-ECM; N = 3, p < 0.05, Bonferroni's multiple comparison test; Fig. 2). Increasing amounts of heparin in the SIS-ECM biomaterial resulted in higher concentrations of FGF-2 in the eluate (Fig. 2). These data show that heparin enhanced loading of FGF-2 into SIS-ECM biomaterial without limiting the subsequent elution of FGF-2 from the biomaterial.

FGF-2 concentration calculated from eluted media at 1, 3, and 24 h after enhancing SIS-ECM with FGF-2 ± heparin (ng/mL) after 2 h. Values shown are mean ± standard deviation; N = 3; *p < 0.05—statistical significance compared to ♦ as determined by unpaired t-test.
Heparin augments FGF-2-mediated attenuation of human cardiac myofibroblast activation
The bioactive effects of heparin on FGF-2 mediated attenuation of human cardiac myofibroblast activation were determined using an in vitro 3D collagen gel contraction model. Human atrial and ventricular cardiac myofibroblasts embedded in collagen matrices produced a baseline level of collagen matrix gel contraction and the addition of TGF-β1, a profibrotic cytokine, was used to stimulate an increased myofibroblast activity (Figs. 3 and 4). In the presence of TGF-β1, FGF-2 attenuated collagen matrix gel contraction by both atrial (normalized contraction ratio compared to baseline: 1.42 ± 0.18 for TGF-β1 vs. 1.17 ± 0.16 for TGF-β1+FGF-2; N = 3, p = ns; unpaired t-test; Fig. 3) and ventricular (normalized contraction ratio compared to baseline: 1.16 ± 0.06 for TGF-β1 vs. 1.10 ± 0.03 TGF-β1+FGF-2; N = 3, p = ns; unpaired t-test; Fig. 4) human cardiac myofibroblasts. Addition of heparin to the FGF-2 solution significantly reduced human atrial (normalized contraction ratio compared to baseline: 1.42 ± 0.18 for TGF-β1 vs. 0.902 ± 0.10 for TGF-β1+FGF-2+heparin; N = 3, p < 0.05; unpaired t-test; Fig. 3) and ventricular (normalized contraction ratio compared to baseline: 1.16 ± 0.06 for TGF-β1 vs. 0.809 ± 0.08 for TGF-β1+FGF-2+heparin; N = 3, p < 0.05; unpaired t-test; Fig. 4) cardiac myofibroblast-mediated collagen matrix gel contraction beyond FGF-2 alone. Heparin was effective in augmenting FGF-2 attenuation of human atrial and ventricular cardiac myofibroblast-mediated collagen matrix gel contraction.
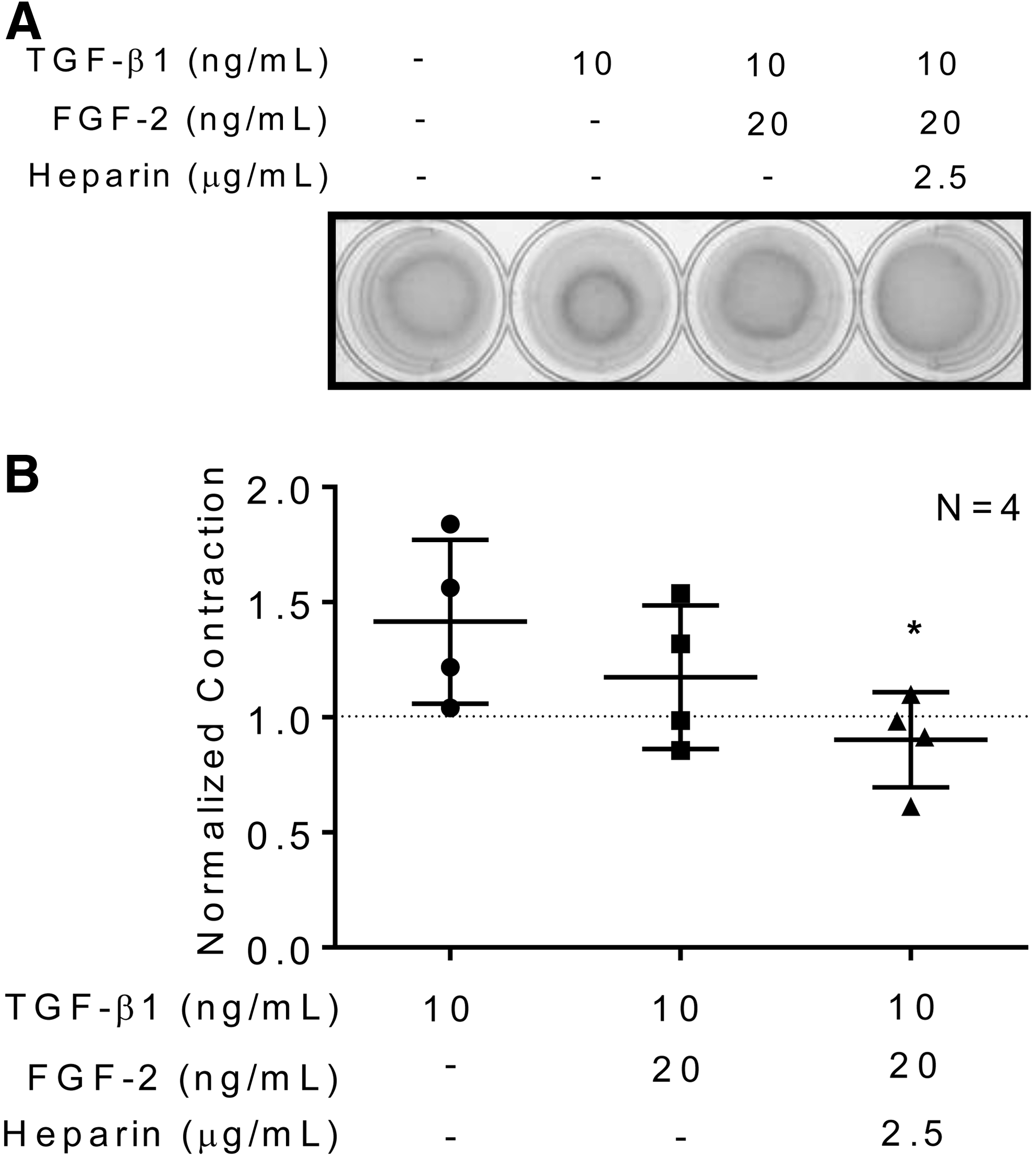
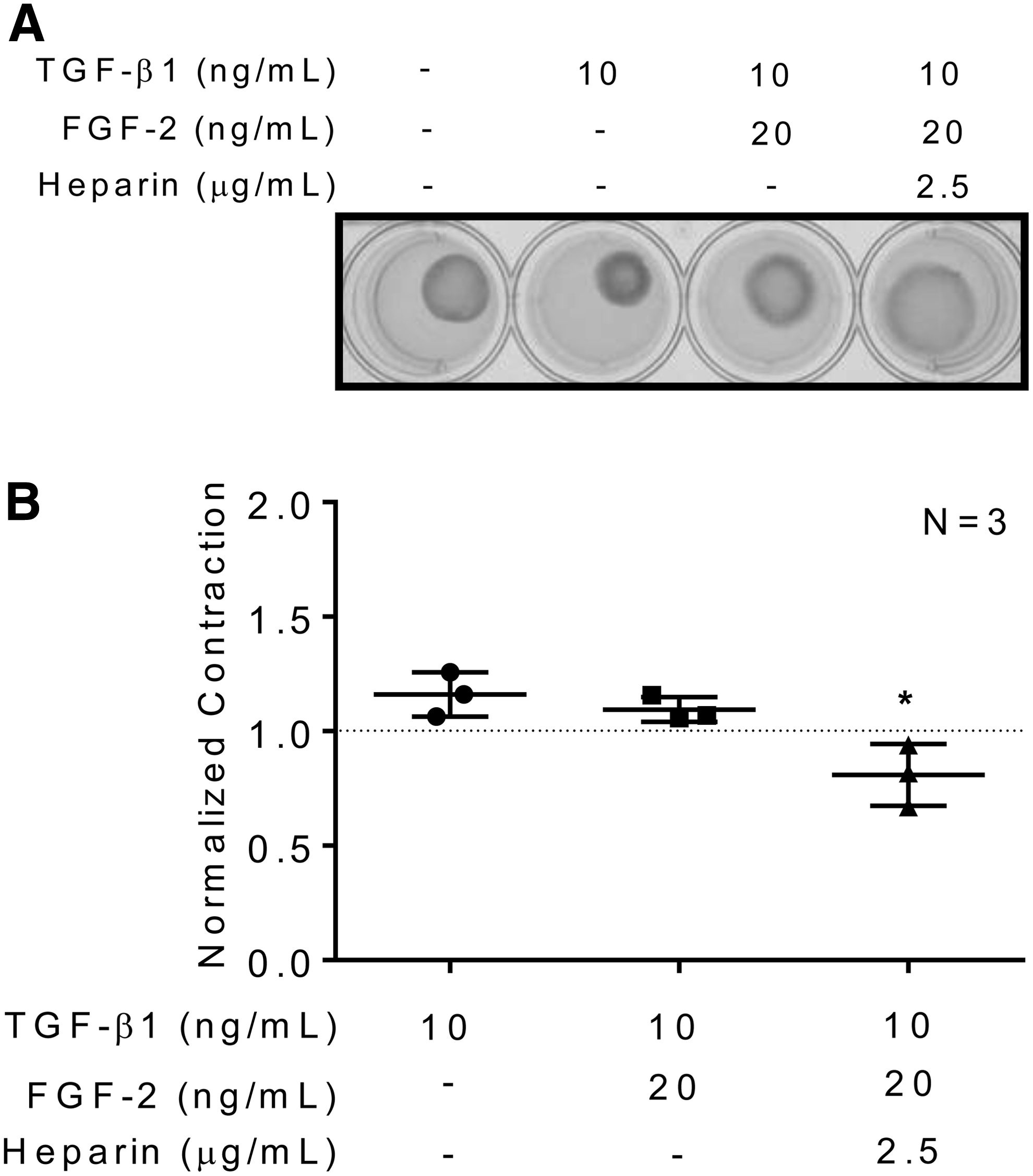
Heparin does not induce human cardiac myofibroblast cell death
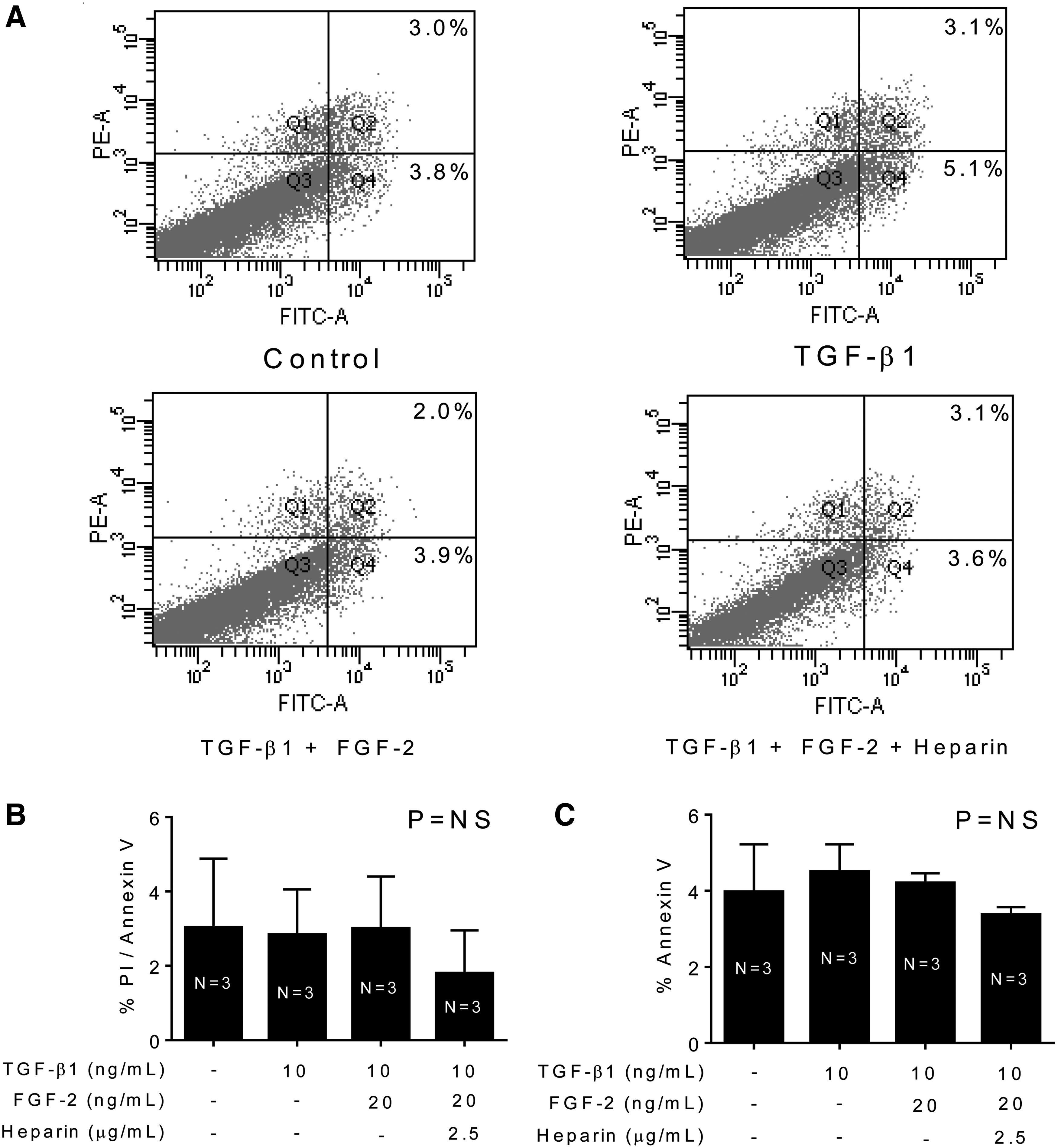
To show that attenuation of gel contraction was not due to cell toxicity from the treatment dosages, annexin V and propidium iodide dye-based apoptosis and necrosis measurements were performed using fluorescence. There were no significant differences in the percentage of cells that were detected as dead or in the early stages of apoptosis in comparison to the controls (Fig. 5).
Discussion
The potential of acellular ECM scaffolds for tissue repair has been well established. Enhancement of biomaterials with targeted growth factors to optimize their bioactive and repair potential is an emerging area for discovery. The ideal approach to enhance biomaterials with growth factors is unclear. Covalent attachment or physical encapsulation of growth factors to delivery vehicles has been explored, but these techniques often rely upon matrix degradation or external triggers for release. 15
We and others have employed a surgical tissue engineering strategy in rat and porcine models that leverages a bioactive decellularized porcine-derived ECM scaffold (SIS-ECM). SIS-ECM supports tissue regeneration, is well tolerated without immune reaction, and has been tested in other cardiac interventions.16–21 Biologic ECM acts as a natural reservoir for bioactive growth factors and may be ideally suited for growth factor enhancements. SIS-ECM contains growth factors, such as FGF-2 and TGF-β1, and ECM components, such as collagen I, III, and IV, fibronectin, and heparin glycosaminoglycan.5,17,22–24 We enhanced SIS-ECM with FGF-2 for the prevention of cardiac remodeling following MI.3,16 FGF-2 enhanced SIS-ECM as a biomaterial for myocardial repair, preserved ECM architecture, prevented remodeling, and improved cardiac performance. 3
We believe SIS-ECM is a suitable delivery vehicle for growth factors in treating cardiac fibrosis. In this study, we uniquely explored a noncovalently incorporated growth factor enhancement technique of SIS-ECM biomaterial with FGF-2 using heparin glycosaminoglycan. The increase in FGF-2 isolated from enhanced SIS-ECM due to an increase in the FGF-2 enhancement concentration may involve more than the ability of the biomaterial tissue to soak in a solution and could be explained by the potential to saturate the heparin molecules found natively in SIS-ECM with FGF-2. Heparin molecules have a high negative charge density, while FGF-2 and its receptor, fibroblast growth factor receptor 1 (FGFR1), have positively charged and polar amino acid residues found in heparin binding domains, such as lysine or arginine and asparagine or glutamine. These have been found to be critical residues in forming a bonded complex through electrostatic molecular interactions.12,25
Even at a fixed FGF-2 concentration, increasing heparin dosage in the enhancement solution led to an increase in the amount of FGF-2 retained and then eluted by SIS-ECM. We believe that the addition of heparin leads to greater incorporation of the glycosaminoglycan into the SIS-ECM, creating a greater potential to be enhanced with FGF-2. This method may be advantageous in comparison to a covalently modified enhancement technique because it preserves the biological structure of the heparin molecule and is more representative of native biochemistry. While heparin can increase the FGF-2 enhancement of SIS-ECM, it does not limit the elution of FGF-2. Release from the biomaterial may be critical for its bioactive effects in host tissues to influence adaptive tissue repair. However, this method may not be ideal for a more controlled release of FGF-2 over a prolonged period of time as may be seen with physical encapsulation or covalent attachment.
Heparin enhances FGF-2 effect in attenuating human cardiac myofibroblast activation
Heparin not only contains binding sites for FGF-2 increasing the potential for its enhancement of biomaterials but also aids in the binding of FGF-2 and its receptor, FGFR1, and potentiates the biological activity of FGF-2.25,26 Without heparin, FGF-2 binds to FGFR1 in a low-affinity complex, 27 but is stabilized to form a dimerized FGF-2-FGFR1 complex with heparin.25,28 In addition, the ability of heparin to increase the half-life of FGF-2 and protect it from proteolytic degradation or denaturation7–11 may explain the augmented antifibrotic effects of FGF-2 on human cardiac myofibroblasts seen with heparin. FGF-2 and heparin did not induce a toxic effect on human cardiac myofibroblasts, proving that the attenuation response was not due to cell death. We have previously shown that reduced collagen matrix gel contraction is correlated with a decrease in the expression of alpha-smooth muscle actin (α-SMA) by cardiac myofibroblasts.14,29 This is correlated with reduced collagen synthesis and deposition, and reduced collagen fiber alignment, representative of local ECM remodeling.14,30 Further study will be required to elucidate the precise mechanisms underlying the observed effects of heparin augmentation on cardiac fibroblasts.
Using heparin to enhance FGF-2 uptake into ECM scaffolds may provide several advantages for in vivo effects when used as a therapy: (1) biocompatibility of SIS-ECM biomaterial with host tissue and documented integration of host cells into implanted material; (2) enhancement with angiogenic and antifibrotic FGF-2 using a noncovalently modified technique preserving biological activity and structure; (3) high loading efficiency of the growth factor in the biomaterial, reducing the translational barriers with respect to cost associated with growth factor usage; and (4) protection of the therapeutic protein from proteolytic degradation or denaturation, as well as augmented and potentiated binding between FGF-2 and FGFR1 and the biological activity of FGF-2.
Limitations
We recognize several limitations of this study. We did not determine the mechanisms by which heparin increases FGF-2 enhancement of SIS-ECM. Second, the FGF-2/heparin tested in the 3D collagen gel matrix assay was not directly eluted from the enhanced SIS-ECM. As a result, the study did not investigate whether FGF-2 eluted from FGF-2-/heparin-enhanced SIS-ECM biomaterial was eluted as free FGF-2 or in interaction with heparin glycosaminoglycan. Last, although reduced collagen gel contraction has been previously correlated to decreased α-SMA expression, a direct analysis of this myofibroblast marker was not measured.
Further study is needed to address several questions regarding the following: (1) the dosage of heparin used to enhance SIS-ECM with FGF-2 and its potential effects on hemostasis at the surgical site; (2) in vivo assessment of FGF-2-/heparin-enhanced SIS-ECM on myocardial fibrosis and heart function; (3) imaging of single cell morphological and ECM alterations; and (4) mechanisms involved in FGF-2-/heparin-mediated reduction of myofibroblast activation.
Conclusion
In conclusion, heparin glycosaminoglycan can augment growth factor enhancement of acellular ECM biomaterial. Heparin augmentation increased the FGF-2 bioactivity in inhibiting human cardiac myofibroblast activation. Glycosaminoglycans should be investigated as a technique for growth factor enhancement of ECM scaffolds for use in biosurgical strategies.
Footnotes
Disclosure Statement
No competing financial interests exist.