
Abstract
Chondrogenesis and mechanical stimulation of the cartilage template are essential for bone formation through the endochondral ossification process in vivo. Recent studies have demonstrated that in vitro regeneration strategies that mimic these aspects separately, either chondrogenesis or mechanical stimulation, can promote mineralization to a certain extent both in vitro and in vivo. However, to date no study has sought to incorporate both the formation of the cartilage template and the application of mechanical stimulation simultaneously to induce osteogenesis. In this study, we test the hypothesis that mimicking both the biochemical and mechanical extracellular environment arising during endochondral ossification can enhance the in vitro mineralization potential of human mesenchymal stem cells (hMSCs). hMSC aggregates were cultured for 21 days under the following culture conditions; (1) Growth Medium − hydrostatic pressure (HP), (2) Chondrogenic Priming–HP, (3) Growth Medium + HP, and (4) Chondrogenic Priming +HP. Each group was then further cultured for another 21 days in the presence of osteogenic growth factors without HP. Biochemical (DNA, sulfate glycosaminoglycan, hydroxyproline, alkaline phosphatase activity, and calcium), histological (Alcian Blue and Alizarin Red), and immunohistological (Col I, II, and X, and BSP-2) analyses were conducted to investigate chondrogenic and osteogenic differentiation at various time points (14, 21, 35, and 42 days). Our results showed the application of HP-induced chondrogenesis similar to that of chondrogenic priming, but interestingly, there was a reduction in hypertrophy markers (collagen type X) by applying HP alone versus chondrogenic priming alone. Moreover, the results showed that both chondrogenic priming and HP in tandem during the priming period, followed by culture in osteogenic medium, accelerated the osteogenic potential of hMSCs.
Introduction
T
Endochondral ossification is the process by which all long bones are formed during early fetal development, as well as in fracture healing. It relies on the production of a cartilage template, which is followed by vessel invasion that allows osteoblast precursors to colonize the tissue.27–30 Chondrocyte differentiation is an integral part of the endochondral ossification process. At each stage of differentiation of the cartilage template, the cells express growth factors that have a significant effect on the endochondral ossification process. For example, proteins produced by hypertrophic chondrocytes (i.e., vessel endothelial growth factor and matrix metallopeptidase) stimulate the breakdown of the hypertrophic cartilage template and induce the invasion of endothelial and mesenchymal stem cells (MSCs), which lead to vessel formation and thereafter mineralization. 31 Recent studies have suggested that in vitro approaches, which mimic certain aspects of the endochondral ossification process, can promote mineralization and vascularization to a certain extent both in vitro and in vivo.32–36 Specifically, studies have shown that chondrogenic priming can significantly increase the osteogenic potential of human MSCs (hMSCs) in vitro compared to culture in osteogenic growth factors.32,34
Mechanical stimulation is also extremely important during endochondral ossification. In utero, mechanical loading of the cartilage template is achieved by external loads imparted by movements and muscular activity of the mother.37,38 Moreover, previous studies have also shown that fetal muscle contractions are crucial for the development of bone tissue in utero. 39 Mathematical models of the endochondral ossification process have been developed and applied to predict how specific mechanical stimuli (e.g., hydrostatic pressure [HP], shear stress, and compressive strain) can both promote and inhibit cells to undergo endochondral ossification.38,39,40–45 Previous in vivo computational models have found that the application of cyclic HP can influence the rate of endochondral ossification in vivo.46–48 Previous in vitro studies have also shown that the application of HP can aid and enhance the chondrogenic differentiation of stem cells and thus promote the production of the cartilage template.49–57
In vivo cartilage within a joint can be exposed to a range of stresses between 3–18 MPa58,59 and strains of −7.2 ± 4.2%, 60 with a typical human's cadence of ∼1 Hz. 61 In vitro studies have applied pressures to stem cells (bone marrow or adipose derived) within the range of in vivo values using HP vessel-based bioreactors that enhanced chondrogenesis.49–57,62 It has been shown that application of HP (0.1 MPa–10 MPa) to human bone marrow stem cells aggregates49,50,53 or those seeded on a collagen52,56 or agarose scaffold 51 can significantly enhance expression of chondrogenic markers (aggrecan and SOX-9).49–53,56 The highest amount of matrix deposition was found when a pressure of 10 MPa was applied for a minimum of 5 days.49,50,53,57 However, in vivo, the formation of the cartilage template and the mechanical stimulation of the template are essential for bone formation. To date, no approach has sought to recreate both the biochemical and mechanical extracellular environment of the endochondral ossification process to enhance the mineralization potential of hMSCs.
This study tests the hypothesis that an in vitro bone regeneration strategy that mimics both the biochemical and mechanical extracellular environment of the endochondral ossification process will provide an effective strategy for in vitro mineralization of MSCs. The main objective of this study was to evaluate the mineralization potential of hMSCs by using cellular aggregates cultured with and/or without external chondrogenic growth factors and simultaneous application of HP. The chondrogenic and osteogenic potentials of hMSC aggregates were assessed using biochemical assays (DNA, alkaline phosphatase activity (ALP), sulfate glycosaminoglycan (sGAG), hydroxyproline, and calcium), along with histological (Alcian Blue and Alizarin Red) and immunohistological (collagen type I, II, and X and Bone Sialoprotein 2 [BSP2]) staining.
Materials and Methods
Human donor MSCs: isolation and characterization
Bone marrow was collected from the iliac crest as described previously, 63 by standard puncture and aspiration into heparinized syringes, from two healthy human male donors (aged 25 and 27 years) after receiving informed consent according to the Declaration of Helsinki and approval by the Ethics Committee of Ulm University. hMSCs were isolated ex vivo by conventional plastic adherence and expanded in vitro in triple-layered cell stack flasks in a standard growth medium consisting of α-minimum essential medium (MEM) supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin, and 10% fetal bovine serum (FBS).
Flow cytometry was performed to determine the phenotypic characterization of hMSCs as described previously. 64 Briefly, cells were washed with phosphate-buffered saline (PBS) and harvested using trypsin, followed by neutralization in 10% FBS and further washing with PBS. Cells were suspended in PBS (1 × 105 cells/mL) with 0.5% bovine serum albumin (BSA) to prevent nonspecific binding and then stained using the following antibodies: CD3-FITC, CD34-FITC, CD45-PE, CD73-PE, CD90-FITC, and CD105-PE. All antibodies were sourced from BioLegend through Ozyme (Paris, France), BD Biosciences (Le Pont de Claix, France), and Beckman Coulter (Paris). Following washing steps in PBS, cells were analyzed by using a Cytomics FC 500 (Beckman Coulter).
Trilineage potentiality of the hMSCs was verified by osteogenic/adipogenic/chondrogenic induction for 21 days. Cells were induced toward osteogenic lineage in monolayer culture in α-MEM media containing 50 μM ascorbic acid, 10 mM β-glycerolphosphate, and 100 nM dexamethasone, and osteogenic differentiation was detected by Alizarin red staining. hMSCs were induced toward the adipogenic lineage in monolayer using a StemPro Adipogenesis Differentiation Kit (Gibco, Life Technologies, France) and adipogenesis was confirmed using Oil Red O solution in 2-propanol, diluted to 60% using deionized water. hMSCs were induced toward the chondrogenic lineage in aggregate culture by using a Chondrogenesis Differentiation Kit (Gibco) and chondrogenic differentiation was identified by Alcian Blue staining. hMSCs were further cultured to passage 3 in normoxic (20% Oxygen) conditions.
Cellular aggregate formation
Once the cells reached a confluency of ∼90%, the cells were trypsinized, counted, and centrifuged at 650 g, 22°C for 5 min. The cells were then resuspended in the standard growth medium at 1 × 106 cells/mL. This cell suspension was divided into 1.5 mL tubes so that there were 250,000 cells in each tube, and then centrifuged for 5 min at 400 g to create cellular aggregates as previously described.30–32,35
Application of biochemical induction and hydrostatic pressure
After 24 h, hMSC aggregates were transferred from tubes to an Evolve (EVO) Cell Culture bag (Quest Biomedical, West Midlands, United Kingdom), containing 25 mL of either a chondrogenic medium (high-glucose DMEM GlutaMAXTM [Gibco, Life Sciences, Dublin, Ireland], 10 ng/mL TGF-β3 [ProSpec-Tany TechnoGene Ltd., Ness-Ziona, Israel], 50 μg/mL ascorbic Acid [Sigma Aldrich], 4.7 μg/mL linoleic Acid [Sigma Aldrich], 100 nM dexamethasone [Sigma Aldrich]), and 1 × insulin-transferrin-selenium (BD Biosciences, Bedford, MA), or a standard growth medium, as described above. Nine aggregates were placed within each EVO cell culture bag and air was removed using a DMSO Resistant Needle Free Valve (Quest Biomedical). hMSC aggregates were cultured within the EVO cell culture bags for 21 days under biochemical or HP stimulation under the following experimental conditions:
1. Cellular aggregates cultured in the standard growth medium for 21 days without HP (Growth Medium − HP). 2. Cellular aggregates cultured in the chondrogenic medium for 21 days without HP (Chondrogenic Priming − HP). 3. Cellular aggregates cultured in the standard growth medium for 21 days with HP (Growth Medium + HP). 4. Cellular aggregates cultured in the chondrogenic medium for 21 days with HP (Chondrogenic Priming+HP).
HP of 10 MPa was applied to the two groups every day at a frequency of 1 Hz for 1 h/day using a custom-built bioreactor, see Figure 1 (A). After 21 days of mechanical stimulation and/or chondrogenic priming in normoxic conditions, cellular aggregates were cultured for up to 42 days in the osteogenic medium consisting of α-MEM media containing 50 μM ascorbic acid, 10 mM β-glycerophosphate, and 100 nM dexamethasone, see Figure 1 (B). At specific time points (14, 21, 35, and 42 days), the culture medium from the cellular aggregates was collected, snap frozen, and stored at −80°C until biochemical assays could be performed. The cellular aggregates were washed with PBS and then treated in one of the following two ways: (1) snap frozen and stored at −80°C for biochemical analysis or (2) fixed overnight in 4% paraformaldehyde before being placed in PBS and refrigerated for histochemical analysis. For each condition, two independent experiments were carried out with at least two repeats in each experiment (n = 2 for histological analysis and n = 3 for biochemical analysis).

Schematic of the experimental design: a custom-built hydrostatic pressure (HP) system
Quantitative biochemical analysis
DNA content
DNA content was analyzed in hMSC aggregates. Five hundred microliters of papain digest (100 mM sodium phosphate buffer containing 10 mM L-cysteine [Sigma Aldrich], 125 μg/mL papain [Sigma Aldrich], and 5 mM Na2EDTA [Sigma Aldrich] in ddH2O at pH 6.5) was added to the cellular aggregates and they were placed in an oven at +60°C overnight as previously described. 65 Once the cellular aggregates were digested, the biochemical assays were performed straight away or samples were stored at −80°C until the assays could be performed. DNA content was performed using the Hoechst 33258 DNA assay with calf thymus DNA (Sigma Aldrich) as a standard, following a previously published protocol.65,66 Briefly, in minimal light, 20 μL of papain digest of the sample/standard is added to a 96-well plate in triplicate. To this, 200 μL of working solution (assay buffer and 1 mg/mL Hoechst Dye solution [Sigma Aldrich]) was added. The plate was incubated away from light for 10 min and then read on a microplate reader (Wallac Victor3 1420 Multilabel Counter) at an excitation of 350 nm and emission of 450 nm as previously described.32,34,65,67
sGAG content
The chondrogenic differentiation of hMSC aggregates was determined by measuring the sGAG content on the papain-digested samples (prepared for the Hoechst assay) and dimethylmethylene blue (DMB) dye (26.25 μg/mL 1,9 DMB, 0.625% ethanol, and 2.5 μg/mL sodium formate, all Sigma Aldrich, in ddH2O at pH 3) with 10 mg/mL bovine-derived chondroitin-4-sulfate (Sigma-Aldrich) as a standard, according to a previously published protocol. 68 Briefly, 40 μL of each sample and standard in triplicate was added to a 96-well plate and 125 μL of the DMB dye was added. The plate was read on the microplate reader at an optical density of 595 nm as previously described. 32
Hydroxyproline content
Hydroxyproline content was determined using the papain-digested samples according to a previously published protocol. 68 Briefly, 50 μL of each sample was added to prelabeled tubes, 50 μL of 6 M HCl was added, and the tubes were placed overnight in an oven at +110°C. Samples were then placed in a laminar flow hood to dry for 3–7 days. The dried samples were reconstituted in 200 μL of acetate-citrate buffer (12 g/L of sodium acetate trihydrate, 5 g/L of citric acid monohydrate, 1.2 mL/L of acetic acid, and 3.4 g/L of sodium hydroxide, all Sigma Aldrich, in ddH2O at a pH of 6.5). Fifty microliters of each sample was added in triplicate to a 96-well plate. Fifty microliters of Chloramine-T reagent (141 mg of chloramine-T in 2.07 mL of dH2O, 2.6 mL of n-propanol, and 5.33 mL of acetate buffer) was added to the 96-well plate and placed on an orbital shaker and allowed to oxidate for 15 min at room temperature. Fifty microliters of the p–DMBA reagent (1.4 g p-dimethyl-aminobenzaldehyde, 7 mL n-propanol, and 3 mL of perchloric acid) was added, mixed gently, and incubated at +37°C for 30 min. The plate was read on the microplate reader at an optical density of 550 nm.
ALP activity
Osteogenic differentiation of hMSC aggregates was studied by measuring the ALP activity in the supernatant of hMSC aggregates using a colorimetric assay of enzyme activity (SIGMAFAST p-NPP Kit; Sigma Aldrich), which uses p-nitrophenyl phosphate (pNPP) as a phosphatase substrate with ALP enzyme (Sigma Aldrich) as a standard. Forty microliters of the medium was added to a 96-well plate in triplicate with a 50 μL of 5 mM pNPP solution, which contains both pNPP and assay buffer. The samples were shielded from direct light at room temperature for 1 h. After this, 20 μL of stop solution (3 N NaOH) was added to the wells and the plate was read at 405 nm in a microplate reader as previously described.34,35,67
Calcium content
Calcium deposition within the aggregates was measured in the hMSC aggregates using the Calcium Liquicolor kit (Stanbio Laboratories) according to the manufacturer's protocol. Briefly, aggregates were digested by adding 1 mL of 0.5 M HCl and rotating the solution overnight in a cold room, and then dissolved with a homogenizer. Ten microliters of each of the digested samples and assay standard was added to a 96-well plate and 200 μL of the working solution was added. The plate was read on a microplate reader at an absorbance of 550 nm as previously described.34,69
Histology
Aggregates from hMSCs were examined by histological methods on days 14, 21, 35, and 42. After each sample had been fixed overnight in 4% paraformaldehyde, samples were dehydrated through a series of increasing grades of ethanol, cleared with toluene, and embedded in paraffin wax using an automatic tissue processor (Leica ASP300; Leica). All samples were sectioned with a thickness of 8 μm using a rotary microtome (Leica microtome; Leica) and mounted on glass slides. Sections were stained with 1% Alcian Blue 8GX solution at pH 1 for sGAG and with Nuclear Fast Red Solution for Nuclei, as previously described.32,34,35,65 For mineralization, 2% Alizarin Red solution was added to sections (all Sigma Aldrich), as previously described.32,34,35
Immunohistochemical analysis
Immunohistochemical analysis was used to detect collagen type I, collagen type II, collagen type X, and BSP2. Sections were deparaffinized overnight in an oven before undergoing a series of rehydration steps through varying ethanol grades (100–50%). The samples were then treated with 40 μg/mL of proteinase K for 20 min at +37°C (Sigma Aldrich), rinsed with 0.5% (v/v) PBS-Tween, and blocked with PBS with 1% w/v BSA (Sigma Aldrich) for 60 min. Sections were then incubated overnight at +4°C with either goat polyclonal anti-Collagen Type I (1:400; sc-8783, Santa Cruz) or rabbit polyclonal anti-Collagen type II (1:100; ab734712 Abcam) in PBS with 1% of BSA (w/v). After three washing steps with PBS containing 1% w/v BSA, the sections were incubated with Alexa Fluor® 488 donkey anti-goat secondary antibody (1:200; Jackson Immunoresearch 705-545-003) and Alexa Fluor® goat anti-rabbit secondary antibody (1:200; Jackson Immunoresearch, 115-485-209), respectively, for 1 hour at room temperature in the dark. The samples were washed thrice in PBS with 1% w/v BSA, and the slides were then incubated overnight at +4°C with mouse monoclonal Collagen Type X antibody (1:1000; ab49945 Abcam) or mouse monoclonal anti-BSP2 (1:200; sc-73497, Santa Cruz). After three washing steps with PBS and 1% w/v BSA, the sections were incubated with DyLight 549 goat anti-mouse secondary antibody (1/200; Jackson Immunoresearch, 115-505-062) for 1 h at room temperature in the dark. Finally, samples were washed thrice with PBS and 1% w/v BSA, and the sections were mounted using 4′,6-diamidino-2-phenylindole mounting media (Sigma Aldrich). Fluorescence emission was detected using confocal laser scanning microscopy (Olympus Fluoview 1000).
Statistical analysis
Results are expressed as mean ± standard deviation. For all the biochemical analysis, two-way analysis of variance was conducted, with time and media type as the independent factors followed by pairwise multiple comparison procedure (Tukey's HSD test). All analyses were performed with GraphPad. For all comparisons, the level of significance was p ≤ 0.05.
Results
hMSC tripotentiality assay
As seen in Figure 2 (A), hMSCs displayed the classical MSC CD90+/CD73+/CD105+/CD34−/CD3−/CD45− phenotype. The differentiation assays confirmed the multipotency of isolated hMSCs through positive markers of (1) osteogenic differentiation; as seen by the Alizarin Red staining of hMSCs cultured in osteogenic differentiation media, but a lack of staining in the negative control group (standard growth media) [Figure 2 (B), left], (2) adipogenic differentiation; as seen by Oil Red O-stained lipid nodules within MSCs cultured in adipogenic induction medium, but a lack of lipid staining in the negative control group (growth media) [Figure 2 (B), centre], and (3) chondrogenic differentiation, as seen by the positive sGAG staining compared to a lack of sGAG staining in the negative control group (growth media) [Figure 2 (B), right].

Characterization of hMSCs.
Viability of hMSCs
As shown in Figure 3, there was no significant difference in the viability of hMSCs aggregates as determined by DNA content between any of the groups at any time point.

DNA content of the hMSCs on day 14, 21, 35, and 42. DNA content per aggregate was measured with a Hoechst assay. n = 6 samples per group per time point. Error bars denote standard deviation.
Production of the cartilage template
To effectively demonstrate the influence of HP on sGAG and hydroxyproline content by hMSCs, the data were analyzed in two ways. (1) The absolute amount of sGAG/hydroxyproline and (2) the normalized amount of sGAG/hydroxyproline secreted over the entire study.
sGAG content
The Growth Medium + HP group had a significant increase (p < 0.0001) in absolute sGAG content from day 21 to 35, see Figure 4 (A). On day 35, there was significantly (p < 0.0001) more absolute sGAG content in the Growth Medium + HP group compared to all other groups. However, this increase in absolute sGAG content is no longer present by day 42.
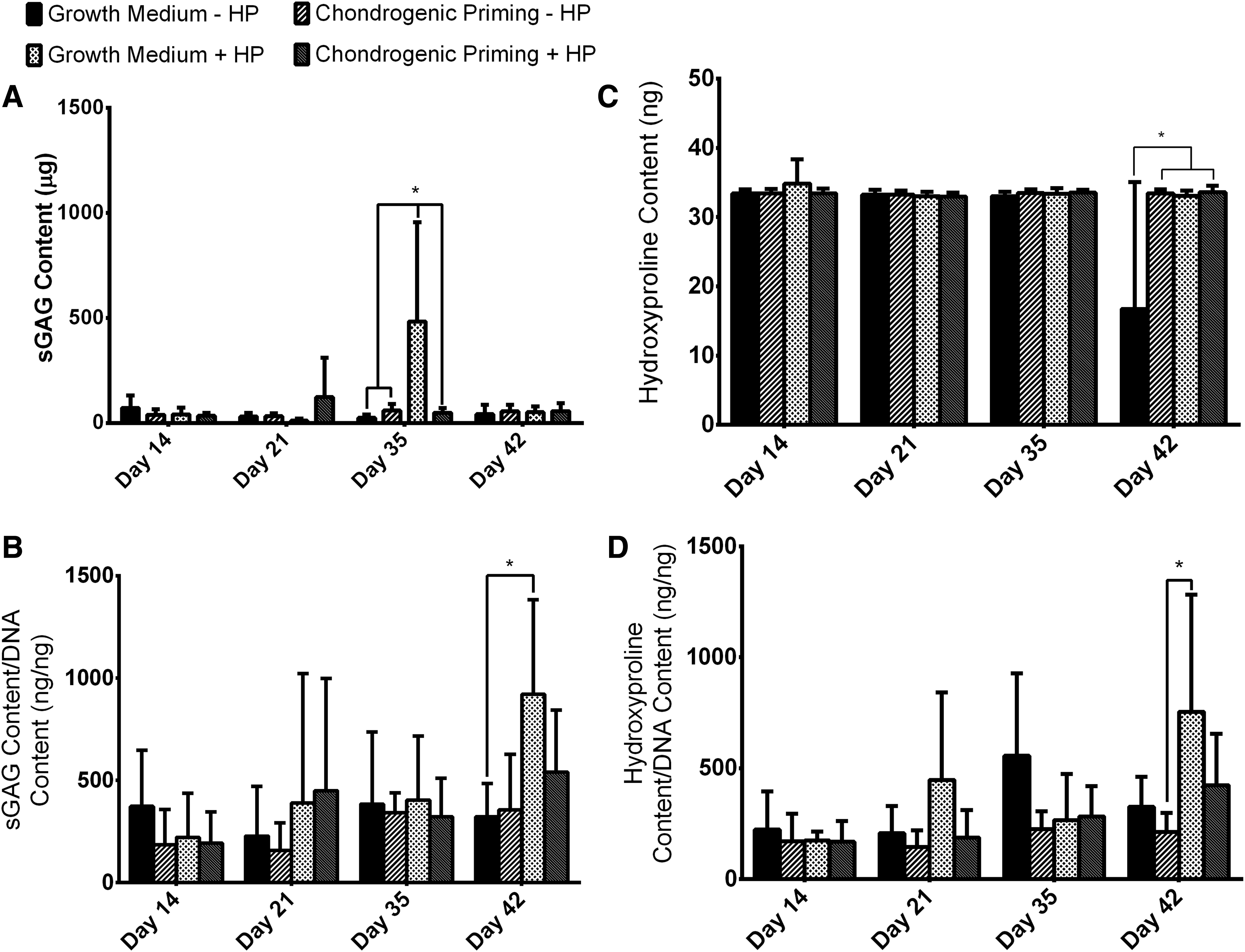
Chondrogenic matrix production by hMSCs in aggregates.
The Growth Medium + HP group had a significant increase (p < 0.001) in normalized sGAG content over the course of the study, see Figure 4 (B). By day 42, there was significantly higher (p < 0.001) sGAG content in the Growth Medium + HP group compared to the Growth Medium − HP group.
Hydroxyproline content
The Growth Medium − HP had a significant decrease (p < 0.0001) in the absolute values of hydroxyproline from day 35 to 49, see Figure 4 (C). By day 49, the Growth Medium − HP group had significantly less (p < 0.0001) hydroxyproline content compared to all other experimental groups.
The Growth Medium + HP group had a significant increase (p < 0.001) in normalized Hydroxyproline content over the course of the study, see Figure 4 (D). By day 42, there was significantly higher (p < 0.05) hydroxyproline content in the Growth Medium + HP group compared to the Chondrogenic Priming − HP group.
Alcian Blue staining
The sGAG results are supported by the Alcian Blue-stained sections of each of the groups on day 21 and 42, see Figure 5. On day 21, the least amount of sGAG deposition was seen in the two groups without HP. The intensity for sGAG staining throughout the aggregate was highest in the Growth Medium + HP group on day 21. The Alcian Blue staining increased over the course of the experiment in the groups without HP and by day 42, there was no difference in sGAG staining between any of the groups.
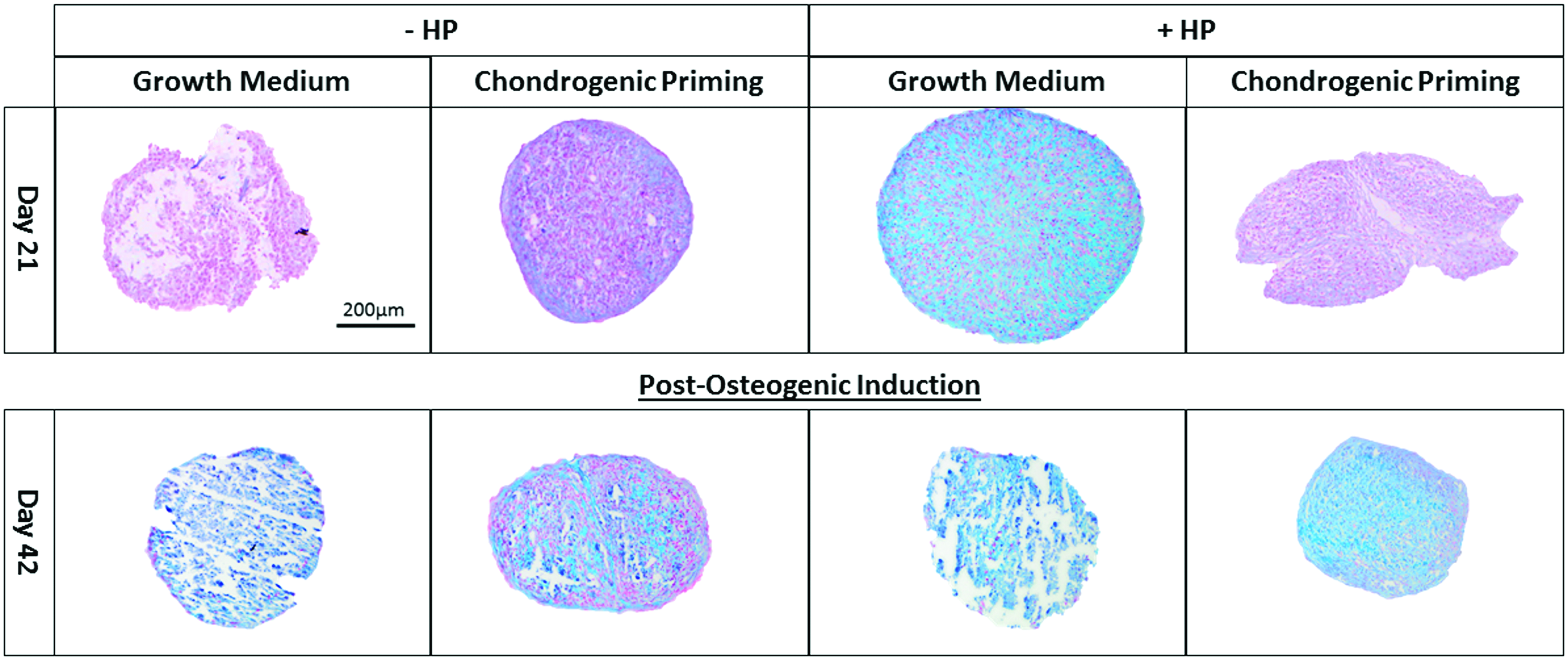
sGAG synthesis in hMSC aggregates. Alcian Blue staining of the groups on day 21 (after application of HP) and day 42 (postosteogenic induction) was performed on cellular aggregates to highlight sGAG in blue and nuclei in red with Nuclear Fast Red. Images were taken at a magnification of 10X. Color images available online at www.liebertpub.com/tea
Collagen type II/collagen type X dual staining
The two groups cultured in the presence of chondrogenic growth factors showed the least amount of positive collagen type II staining, see Figure 6. On day 21, positive collagen type X staining was only present in the Chondrogenic Priming ± HP groups irrespective of the application of HP. However, by day 42, the intensity of collagen type X staining in the Chondrogenic Priming ± HP groups had reduced compared to day 21. On the other hand, the intensity of collagen type X staining increased in the Growth Medium ± HP groups from day 21 to 42.
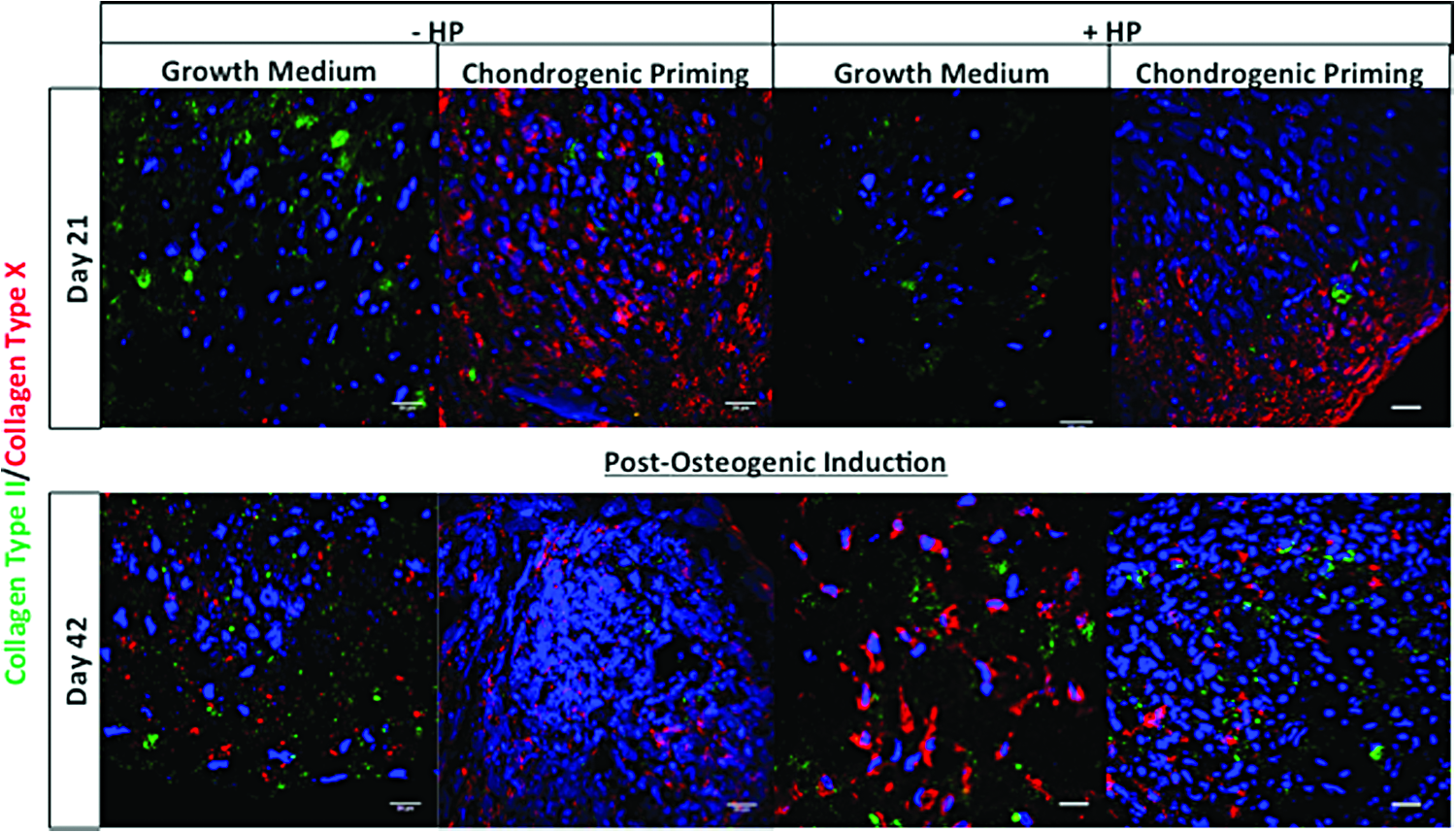
Influence of treatments on chondrogenic differentiation of hMSC aggregate. Collagen type II (green) and collagen type X (red) dual staining were performed on sections of cellular aggregates of the groups on day 21 (after application of HP) and day 42 (postosteogenic induction). Nuclei were labeled with DAPI (blue). Images were taken at a magnification of 60×. DAPI, 4′,6-diamidino-2-phenylindole. Color images available online at www.liebertpub.com/tea
Mineralization of the cartilage template
To effectively demonstrate the influence of HP on ALP and calcium content by hMSCs, the data were analyzed in two ways. (1) The absolute amount of ALP/calcium and (2) the normalized amount of ALP/calcium secreted over the entire study.
ALP activity
On day 14 and 21, there was a significant increase (p < 0.0001) in the absolute ALP activity in the Growth Medium ± HP groups compared to the Chondrogenic Priming ± HP groups, see Figure 7 (A). On day 35, there was a significant increase (p < 0.0001) in the ALP activity in the Chondrogenic Priming ± HP groups. There was no significantly difference in the ALP activity between the groups on day 35 and 42.
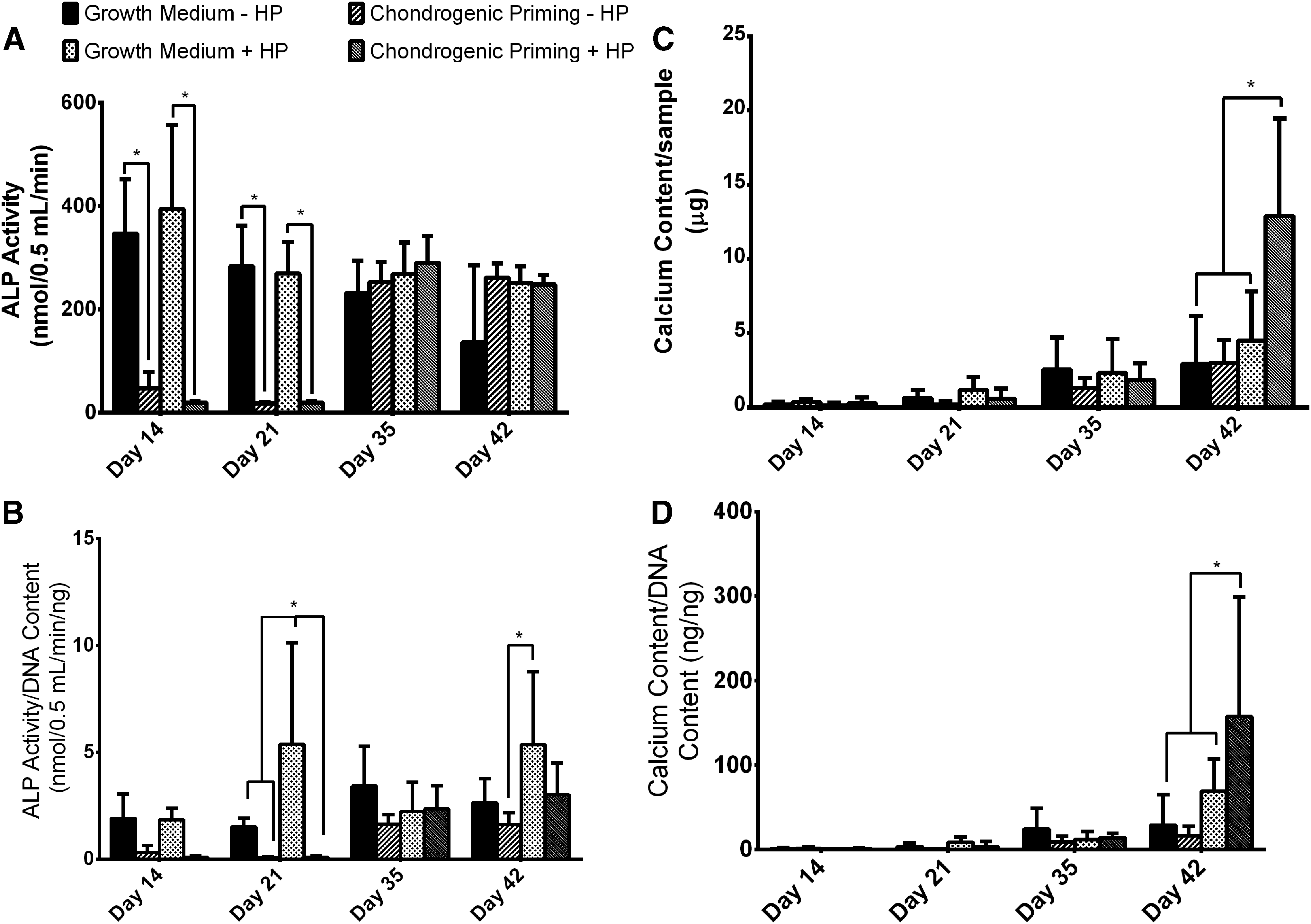
Effect of treatments on early- and late-stage osteogenesis markers in hMSC aggregates.
On day 14, there was no significant difference in the normalized ALP Activity between any of the groups, see Figure 7 (B). On day 21, there was significantly higher (p < 0.001) ALP activity in the Growth Medium + HP group compared to all other groups, however. By day 35, there was no significant difference between any of the groups. On day 42, there was significantly higher (p < 0.05) ALP activity in the Growth Medium group + HP group compared to the Chondrogenic Priming − HP group.
Calcium content
There was no significant difference in absolute calcium content between any of the groups on day 14, 21, and 35, see Figure 7 (C). However, by day 42, there was significantly higher (p < 0.0001) calcium content in the Chondrogenic Priming + HP group compared to all the other groups. On day 42, there was also a significantly higher (p < 0.05) normalized calcium content in the Chondrogenic Priming + HP group compared to all the other groups.
Calcium staining
The calcium results were supported by the Alizarin Red-stained sections of the groups on day 21 and 42, see Figure 8. On day 21, there was no positive staining for mineralization in any of the groups. However, by day 42, there was positive staining for mineralization in all of the groups. Similar to the calcium results, the least amount of positive red staining was seen in the Chondrogenic Priming − HP group and the highest amount of positive red staining was seen in the Chondrogenic Priming + HP group.

Mineralization in hMSC aggregate. Alizarin Red staining of the groups on day 21 (after application of HP) and day 42 (postosteogenic induction) was done on aggregate sections. Images were taken at a magnification of 10×. Color images available online at www.liebertpub.com/tea
Collagen type I/bone sialoprotein 2 dual staining
On day 21, the highest amount of positive staining for both collagen type I and BSP-2 was seen in the Chondrogenic Priming + HP group, see Figure 9. There was minimal positive staining in the Growth Medium ± HP groups and no BSP-2 staining in the Chondrogenic Medium − HP group on day 21. However, by day 42, there was an increase in positive staining for collagen type 1 and BSP-2 in the Growth Medium ± HP groups and an increase in BSP-2-positive staining in the Chondrogenic Priming − HP group. On the other hand, there was a decrease in positive staining in the collagen type 1 and BSP-2 staining in the Chondrogenic Priming + HP group from day 21 to 42.
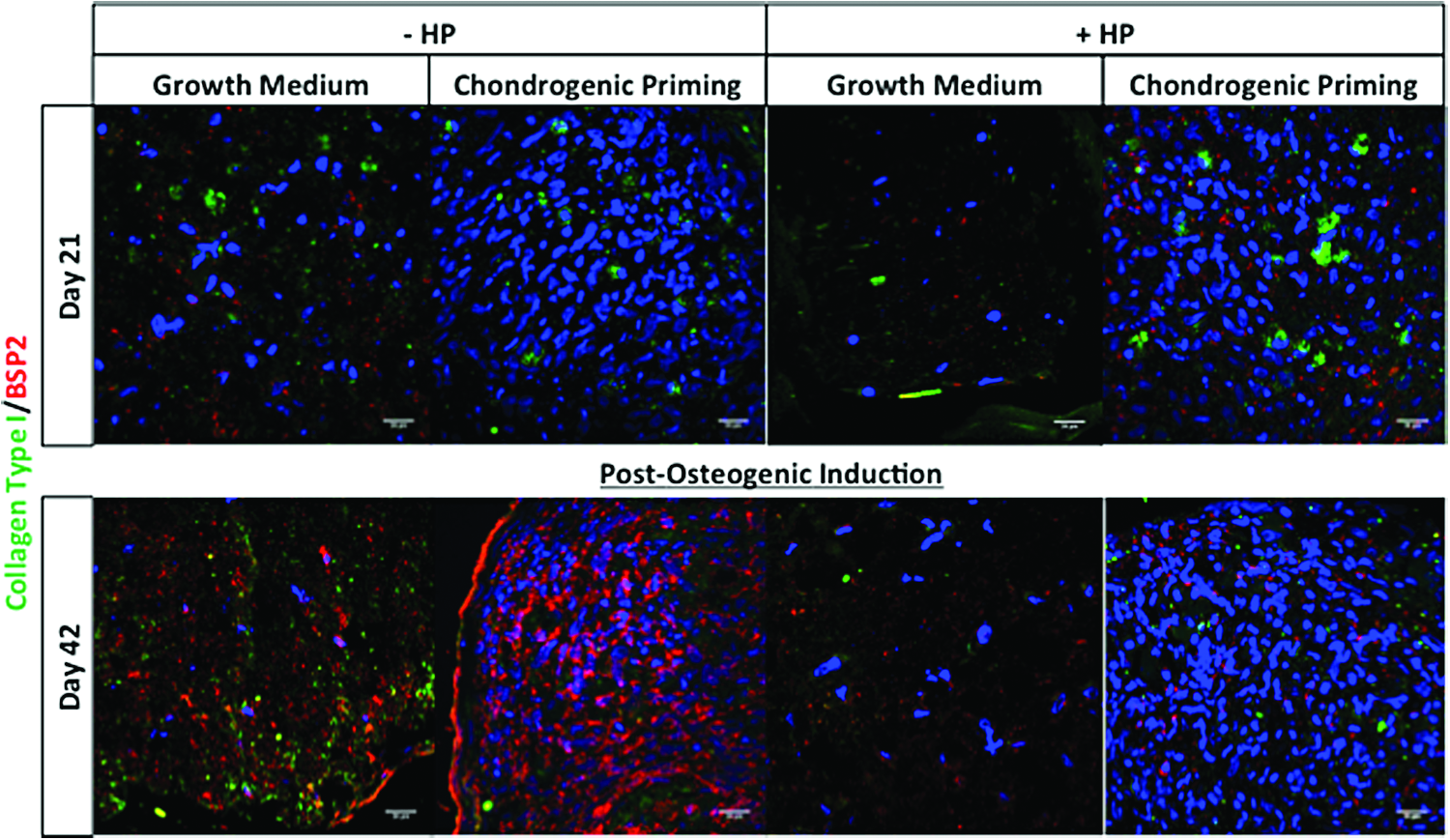
Bone matrix proteins in hMSC aggregate. Collagen type I (green) and BSP-2 (red) dual staining was performed on aggregate sections of the groups on day 21 (after application of HP) and day 42 (postosteogenic induction). Nuclei were labeled with DAPI (blue). Images were taken at a magnification of 60×. Color images available online at www.liebertpub.com/tea
Discussion
This study investigated whether an in vitro bone regeneration strategy that mimics both the biochemical and mechanical extracellular environment of the endochondral ossification process would provide an effective strategy for in vitro mineralization of hMSCs. The results from this study found that the application of HP had a significant effect on both early and late osteogenic markers compared to the same groups cultured without HP. Specifically, the results from this study show that the osteogenic potential (calcium content) of hMSCs was favorably enhanced by applying both chondrogenic priming and HP in tandem during the priming period. Interestingly, this study also demonstrated that postosteogenic induction without HP for 21 days induced the conversion of the chondrogenic aggregates into mineralized pellets through a process resembling endochondral ossification. This conversion was enhanced by applying both chondrogenic priming and HP for 21 days before postosteogenic induction.
One limitation of this study was that osteogenic and chondrogenic factors (ascorbic acid dexamethasone, β-glycerol, and TGF-β3) were introduced into the culture media of hMSCs to encourage MSC differentiation down specific pathways. It was unlikely that these factors are present in the combinations used in this study during endochondral ossification in vivo, and some concern may exist regarding cell viability in long-term in vitro studies. However, previous research studies have exposed MSCs to both chondrogenic and osteogenic differentiating agents for long durations (14 days to 5 weeks) and have demonstrated long-term viability and matrix production by MSCs.25,26,53,65,68,70–75 Furthermore, we had control groups without any chondrogenic growth factors and there was no difference in cell viability between those cultured with/without growth factors. Another potential limitation of this study is that MSCs from two male donors were pooled and we did not directly explore whether the hMSCs displayed a donor-dependent response to mineral formation. Previous studies have reported donor variability in the expression of osteogenic markers in vitro. 76 However, the control groups also contained pooled cells, so the differences observed between the groups cannot be explained by donor variability. Another limitation of this study is the use of osteogenic growth factors to induce mineralization after application of mechanical stimuli, which allowed us to investigate the in vitro osteogenic potential of our approach. However, we have previously shown that chondrogenic and vascular priming in vitro, in the absence of osteogenic growth factors, ultimately led to enhanced vessel formation and increased mineralization within the cellular aggregate when they were implanted in vivo. 33 Therefore, we expect that these cellular aggregates would also show enhanced mineralization once implanted in vivo, as the body would provide the natural biochemical cues. However, future studies should investigate the in vivo potential of mimicking the biochemical and mechanical environment of endochondral ossification. Moreover, future studies should investigate in vivo mineralization potential of combining all three vital stages of the endochondral ossification process: (1) biochemical and (2) mechanical environment, as well as (3) the cellular niche. The final limitation of this study was the drop in cell number in the groups over time course of the study. This reduction in cell number may be due to long in vitro culture, however, as stated in the above MSCs, that have been exposed to both chondrogenic and osteogenic differentiating agents for long durations25,26,53,65,69,70–75 and have demonstrated long-term viability and matrix production by MSCs. Moreover, there was no significant difference between our experimental groups and our control groups.
The first stage of endochondral ossification is the aggregation of MSCs to form cellular aggregates, after which the cells undergo chondrogenic differentiation to form the cartilage template.27,29,77 It is during this early stage of limb development that mechanical stimuli have been found to have a large effect on the rate of endochondral ossification.38,39,40–45 Specifically, computational models of embryonic chick hind limb rudiments predicted that HP was at its highest toward the center of the rudiment, which is where the periosteal collar begins to form 1–1.5 days later. 39 Our previous studies showed that mimicking the cellular niche and biochemical environment of the endochondral ossification process can promote mineralization and vascularization to a certain extent both in vitro and in vivo.32–35 This study aimed to mimic not only the biochemical environment but also the mechanical extracellular environment of the endochondral ossification process. This was achieved by allowing the cells to form a cellular aggregate and undergo chondrogenic differentiation, during which time HP was applied. Similar to previous computational studies,46–48 the results of this study found that the application of HP can have a significant effect on the rate of mineralization potential of hMSCs. However, unlike previous studies, the results also show that by mimicking both the formation of the cartilage template and the application of HP in tandem, there was a significant increase in late osteogenic markers (calcium), and a significant decrease in early bone (ALP and BSP-2) and cartilage markers (sGAG and collagen type II). Moreover, the values of calcium content seen in the two groups exposed to HP were up to seven times larger than those seen in previous studies, which investigated the mineralization potential of chondrogenic priming alone 32 and chondrogenic and vascular priming.34,35 This increase in calcium content might be explained by the fact that by mimicking both vital stages of the endochondral ossification process, the maturation of the cartilage template can be increased, specifically, the process in which the cartilage template undergoes hypertrophy, which occurred by day 21, as seen with collagen type X staining. As cartilage hypertrophy is a known precursor of mineralization, this led to rapid ossification of the cellular aggregates, as seen with our Alizarin Red and calcium content. However, when only HP was applied, it slowed down the maturation of the cartilage template, with hypertrophy of the cartilage template and production of early osteogenic markers (ALP) only occurring after 42 days of in vitro culture. Ultimately, if left for longer durations, this group, we expect that this may have led to a significant increase in mineralization. Future studies should investigate longer time points and which genes and genetic pathways are being affected through the application of the mechanical stimuli.
HP has been well established as a mechanical stimulus to induce chondrogenesis in hMSCs.49–57 Specifically, previous studies have found that the application of HP within ranges similar to that seen in vivo to hMSCs as a cellular aggregate49,50,53 or seeded on a collagen52,56 or agarose scaffold 51 can significantly enhance the chondrogenic differentiation process, specifically the expression of aggrecan and SOX-9.49–53,56 Interestingly, the results we present in this study further verify that the application of HP to hMSCs can not only enhance chondrogenic differentiation in the presence of chondrogenic growth supplements but also, the application of HP without any external growth factors resulted in reduced hypertrophy, while allowing for chondrogenesis; however, once the HP was removed and the cells were exposed to osteogenic growth factors, the cells began to produce hypertrophic markers, but at a much slower rate than those exposed to chondrogenic growth factors alone. One of the major limitations to the use of MSCs for cartilage-based tissue engineering strategies is that the cells have limited success postimplantation due the constructs undergoing hypertrophy and thus inducing bone formation.78–84 We propose that the application of HP alone on hMSCs can obviate the need for chondrogenic supplements to induce chondrogenesis by hMSCs, while also reducing hypertrophy within a 3D cartilage-based tissue-engineered constructs.
Conclusions
The results from this study showed that the application of HP has a significant effect on both the chondrogenic and osteogenic differentiation of hMSC aggregates. Specifically, the results from this study also show there was a reduction in hypertrophy markers (collagen type X), a leading limitation to current cartilage tissue engineering approaches by applying HP alone. Moreover, the results from this study showed an acceleration of hMSC osteogenesis (calcium content), due to the combination of chondrogenic priming and the application of HP in tandem. We propose that this approach mimicked endochondral ossification in vivo. Therefore, we conclude that to induce stable chondrogenesis, it is more beneficial to apply HP alone, without any external growth factors. However, to enhance the osteogenic potential of hMSCs, it is more beneficial to apply both chondrogenic priming and HP in tandem. We propose that there may be a trade-off between mineralization and chondrogenesis of hMSCs for in vitro HP strategies.
Footnotes
Acknowledgments
This project was supported by the European Research Council Grant 258992 (BONEMECHBIO), European Regional Development Fund through the Science Foundation Ireland (SFI) Investigators Programme Grant 258992, and the seventh Framework programme through the project REBORNE Grant No. HEALTH-2009-1.4.2-241879. The authors would like to thank Markus Rojewski and Hubert Schrezenmeier of the Institute for Clinical Transfusion Medicine and Immunogenetics Ulm, German Red Cross Blood Donor Service, University of Ulm, for supplying the bone marrow stromal cells.
Author Disclosure Statement
There are no financial and personal relationships between the authors and others who might bias their work. All authors state that they have no conflicts of interest.