
Abstract
The application of bone morphogenetic protein 2 (BMP-2) has been extensively investigated to improve diabetes-impaired bone healing; however, the delivery of BMP-2 by gene therapy for bone regeneration has rarely been investigated in diabetic animals. In this study, we aimed to evaluate which cells induce more new bone formation in diabetic animals when cell-based BMP2 gene therapy is applied. For this purpose, we harvested bone marrow stromal cells (BMSCs) twice in the same animal before (non-diabetic BMSCs; nBMSCs) and after diabetes induction (diabetic BMSCs; dBMSCs) using modified bone marrow ablation methods. And then, cells were transduced by adenoviral vectors carrying the BMP2 gene (AdBMP2). In in vitro, AdBMP2-transfected dBMSCs (B2/dBMSCs) produced higher BMP-2 mRNA levels over 48 h, whereas AdBMP2-transfected nBMSCs (B2/nBMSCs) exhibited a transient increase in BMP-2 mRNA followed by a decrease to the baseline level within 48 h. Both B2/dBMSCs and B2/nBMSCs induced secretion of BMP-2 for 3 weeks. However, B2/dBMSC BMP-2 secretion peaked from day 3 to 10, whereas B2/nBMSC BMP-2 secretion peaked from day 1 to 7. The analysis of osteogenic activity revealed that mineralization nodule formation and the expression levels of osteogenic genes were significantly higher in B2/dBMSCs than B2/nBMSCs and were accompanied by upregulation of canonical Wnt/β-catenin and Smad signaling. AdBMP2-transfected autologous cells were implanted into critical-sized calvarial defects in diabetic animals and induced significantly more bone regeneration than non-AdBMP2-transfected cells. In addition, B2/dBMSCs led to significantly more new bone formation than B2/nBMSCs. Thus, BMP2 gene therapy using diabetic cells effectively supported diabetic bone healing and it was related to the enhanced responses to AdBMP2 of dBMSCs.
Introduction
D
To improve diabetic bone healing, bioactive molecules, such as growth factors, have been applied to these types of defects; bone morphogenetic protein 2 (BMP-2) is a candidate as the most potent molecule for inducing bone formation.4–6 For osteogenesis, supraphysiologic doses of BMP-2 are needed and are dependent on the condition of animals, species of animals, and defect type in recombinant human BMP-2 (rhBMP-2) application.7–9 Zara et al. demonstrated that less than 30 μg/mL of rhBMP-2 was not sufficient to induce bone formation and more than 150 μg/mL of rhBMP-2 induced adverse effects such as activation of osteoclasts and increase of adiposity of bone marrow in rat fracture healing models. 10 On the other hand, using vectors carrying the BMP2 gene, it is possible to obtain a therapeutic effect at a lower concentration. BMP2 gene was directly or indirectly delivered to the defect site by the transplantation of transfected cells, subsequently inducing BMP-2 production in vivo.11–14 In our previous studies, mesenchymal cells (MSCs) were transfected by adenoviral vectors carrying the BMP2 gene (AdBMP2), and less than 0.01 μg/mL BMP-2 was produced over a period of 2 weeks and effective for de novo bone formation in an 8-mm critical-sized calvarial defect in healthy animals. 15 However, bone regeneration using vectors carrying BMP2 gene was not attempted in diabetic bone healing.
When the cell-based gene therapies like our previous study are designed for diabetic animals, 15 there are some questions about the results when diabetic cells are applied for BMP2 gene delivery because osteogenic differentiation of MSCs is closely related to glucose level and is regulated through runt-related transcription factor 2(RUNX2)-glucose transporter 1 (GLUT1). 16 Generally, osteogenic differentiation in high glucose is inhibited and cell apoptosis is increased. 17 Diabetic MSCs have also shown that the number of colonies is decreased and osteogenic gene expression such as Cbfa1/runx2 and osteocalcin, osteoblastic differentiation, and mineralization processes are reduced comparing to healthy MSCs.18–21 However, MSCs harvested from rat spinal ligaments increased osteogenic gene expression in high glucose concentration and BMP-2-induced osteogenic gene expression was also enhanced in high glucose. This phenomenon is suggested as one of the mechanisms of pathological ossification of the posterior longitudinal ligament. 22 Similarly, diabetes activates vascular BMP-2 expression and BMP-2/Smad signaling pathway and it is associated with an increased osteogenesis and calcium accumulation in aorta. 23 Therefore, osteogenic differentiation and BMP-2 responsiveness of MSCs are dependent on the environment, and response about AdBMP2 of diabetic MSCs is unknown. If so, it is also a question of whether bone regeneration would be more beneficial for diabetic bone healing if healthy cells that were harvested before diabetes induction are applied for BMP2 gene delivery.
In this study, we aimed to evaluate diabetic cell-based BMP2 gene therapy on bone regeneration in diabetic animals. For this purpose, non-diabetic bone marrow stromal cells (nBMSCs) were harvested from healthy animals; these animals were then injected with streptozotocin to induce type 1 DM. After 8 weeks of diabetic induction, diabetic BMSCs (dBMSCs) were harvested from the diabetic animals. Cell-based BMP2 gene delivery was then conducted with these two types of BMSCs, and we examined the resulting differences both in vitro and in vivo.
Materials and Methods
Animals
In this study, 36 male Sprague-Dawley rats (8 weeks of age) were used and housed in specific, pathogen-free facilities. The animals were cared for and treated in accordance with guidelines established by the Seoul National University Institutional Animal Care and Use Committee. This study conformed to the Animal Research: Reporting In Vivo Experiments (ARRIVE) guidelines for preclinical studies. The animal research protocols were approved by the Institute of Laboratory Animal Resources, Seoul National University (SNU-120309-1).
Experimental design
The experimental design is summarized in Figure 1. In brief, while the animals were under general anesthesia with ketamine (90 mg/kg; Yuhan Co., Seoul, Korea) or xylazine (10 mg/kg; Bayer), nBMSCs were harvested from the tibiae (mean body weight, 338.8 ± 3.5 g and fasting blood glucose level, 104.8 ± 11.7 mg/dL). 24 The wounds were allowed to heal for 2 weeks; subsequently, the animals were fasted for 4 h, and diabetes was induced with a single intraperitoneal injection of streptozotocin (Sigma-Aldrich, St. Louis, MO; 60 mg/kg, freshly dissolved in 5 mmol/L citrate buffer, pH 4.5).25,26 Two weeks after diabetes induction, blood glucose levels were monitored using a glucometer (Accu-Check; Boehringer Mannheim Corporation, Indianapolis, IN). Rats with blood glucose levels greater than 300 mg/dL were considered diabetic (mean body weight, 396.3 ± 29.7 g). The diabetic rats showed marked polyphagia, polydipsia, and polyuria. After DM was confirmed, dBMSCs were harvested in the same manner after 8 weeks of DM. After 2 weeks of healing, the diabetic animals underwent regenerative therapy using either autologous nBMSCs or dBMSCs, according to the experimental group.
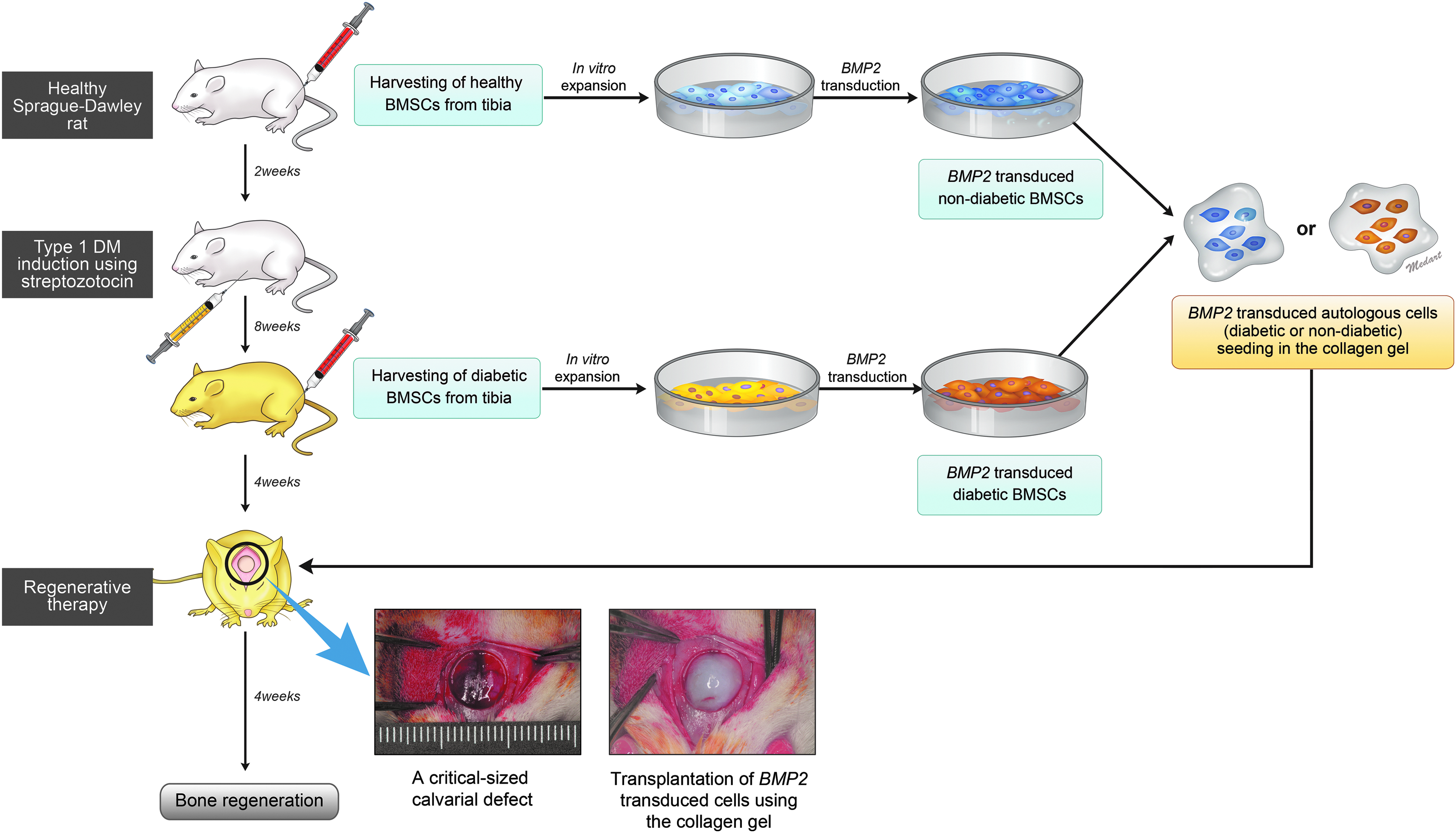
Diagram of the experimental design. nBMSCs were harvested from both tibiae of each healthy animal under general anesthesia before diabetes induction. After 2 weeks of wound healing, type I DM was induced in each animal with a single intraperitoneal injection of streptozotocin. Blood glucose levels were measured 2 weeks later to confirm hyperglycemia (≥300 mg/dL). Eight weeks after DM induction, dBMSCs were harvested from both tibiae of each animal under general anesthesia. The harvested cells were expanded in culture media and stored in liquid nitrogen until transplantation. Before in vivo cell transplantation according to their designated in vivo experimental group, the autologous non-diabetic or diabetic cells were thawed and expanded to passage two or three. The cells were seeded at a density of 1 × 106 cells/dish and transduced with AdBMP2. The cells were transplanted into the critical-sized calvarial defects in diabetic rats using collagen hydrogel, and bone regeneration was evaluated after 4 weeks of healing. DM, diabetes mellitus; nBMSCs, non-diabetic bone marrow stromal cells; dBMSCs, diabetic bone marrow stromal cells.
Isolation and culture of BMSCs
To isolate BMSCs in vivo, the modified bone marrow ablation technique was used to aspirate the bone marrow from the tibia. 18 Under general anesthesia, an oblique incision was made over the body of the tibia. A 1.0-mm hole was punctured into the marrow cavity at the lower 2/3 area of the tibial body using a round bur (RA2-010, Komet, Kometgroup, Besigheim, Germany) at low speed under sterile saline irrigation. A sterile 10-mL syringe was used to aspirate the cells and blood from the bone marrow cavity, and the marrow was then back-flushed by injecting 5 mL of culture media into the tibiae. Layered sutures were performed using 4-0 vicryl for muscle and 4-0 silk for skin. All the animals received a single intramuscular injection of cefazolin (30 mg/kg) for 2 days after surgery.
After centrifugation, the cell pellet was resuspended in expansion media consisting of alpha-minimum essential medium (α-MEM) (Gibco) supplemented with 10% fetal bovine serum (FBS; Gibco) and a 1% penicillin–streptomycin solution (Gibco). The cells were incubated at 37°C in a humidified atmosphere of 95% air and 5% CO2. Media were changed twice per week. Upon reaching confluence, the cells were released with 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (EDTA; Gibco). Cells from passages two to three were used for transfection.
Construction of BMP-2-producing BMSCs
The construction of an adenoviral vector encoding human BMP-2 (AdBMP2) has been previously described. 27 The BMSCs were plated at a density of 1 × 104 cells/well on a 24-well plate for the alkaline phosphatase (ALP) activity assay, BMP-2 expression quantification, and the mineralization assay, and at 5 × 103 cells/well on a 96-well plate for the cell proliferation assay. The BMSCs were transduced with AdBMP2 at a multiplicity of infection of 100 pfu for 4 h in medium consisting of α-MEM supplemented with 10% FBS and a 1% antibiotic–antimycotic solution, and then an equivalent volume of medium was added. BMP-2 expression was confirmed by enzyme-linked immunosorbent assay (ELISA; R&D Systems).
Cell proliferation assay
Cell proliferation after 3 and 7 days was assessed using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma, St. Louis, MO). MTT solution was added to each cultured sample at the indicated days, and the cells were further incubated for 4 h to allow MTT formazan formation. The purple formazan of each sample was extracted with dimethyl sulfoxide (Sigma), and the optical density (OD) of each extracted solution was measured using an automatic microplate reader (ThermoMax; Molecular Devices, Sunnyvale, CA) at a wavelength of 540 nm. Each sample was measured in triplicate.
Measurement of BMP-2 expression level
The expression of BMP-2 was quantified using a commercial ELISA kit. The culture medium in each well of the 24-well plates was sampled at 3, 7, 10, 14, 17, and 21 days and frozen at −80°C for later analysis. The amount of BMP-2 in the collected medium sample was measured using a commercial enzyme immunoassay kit. All experiments were performed in triplicate.
Measurement of osteogenic activity
To assess the cellular osteogenic activity, the ALP activity and mineralization assays were performed. The ALP activity was measured at 7 and 14 days according to the protocol of the ALP activity assay kit (AnaSpec Co.). The OD of the p-nitrophenol (pNP) produced by the enzymatic hydrolysis of the p-nitrophenylphosphate substrate at 405 nm was measured using an absorbance microplate reader. The protein concentration in the samples was measured using a protein assay kit (iNtRON Biotechnology, Seoul, Korea). The ALP activity is expressed as the concentration of pNP per microgram of protein.
For the mineralization assay, the cells were cultured in osteogenic media (α-MEM with 15% FBS supplemented with 0.2 mM ascorbic acid [Gibco] and 10 mM β-glycerol phosphate [Gibco]) for 21 days. Then, the cells were fixed in 95% cold ethanol. Next, the cells were stained with a 1% alizarin red S solution (Wako Chemicals, Osaka, Japan) for 5 min. Mineralization was examined and imaged. Finally, measurements were obtained using a quantitative eluting procedure with cetylpyridinium chloride in 10 mM sodium phosphate, and the OD of each sample at 540 nm was measured using an absorbance microplate reader.
Gene expression analysis using quantitative real-time polymerase chain reaction
The cells were plated in a dish at a density of 1 × 106 cells and transfected with AdBMP2 in α-MEM supplemented with 10% FBS and an antibiotic–antimycotic solution. Total RNA was isolated at 0, 6, 12, 24, and 48 h using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. To remove any residual genomic DNA in the RNA solutions, an RNase-free DNase treatment (Qiagen) was performed, and the first strand of single-stranded cDNA that was synthesized from 5 μg of RNA using a Superscript III first-strand synthesis system (Invitrogen) was used as a polymerase chain reaction (PCR) template. Quantitative real-time PCR (qRT-PCR) was performed using a Real-Time PCR system (Applied Biosystems), cDNA, primers (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/tea), and a reaction mixture containing SYBR Premix Ex Taq II (Takara Bio Co.). The thermal cycling conditions were as follows: one cycle of 95°C for 15 s followed by 40 cycles of 95°C for 15 s, 60°C for 15 s, and 72°C for 33 s. Post-PCR melting curves confirmed the specificity of the single-target amplification, and the fold change of the gene of interest relative to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was determined in triplicate.
Western blot analysis
The cells were plated in a dish at a density of 1 × 106 cells and transfected with AdBMP2 in α-MEM supplemented with 10% FBS and an antibiotic–antimycotic solution. These cells were washed with ice-cold phosphate-buffered saline and harvested with Cell Extraction Solution (iNtRON Biotechnology, Korea) and a protease inhibitor cocktail solution (Sigma-Aldrich) according to the manufacturer's protocol. Total cell lysates (30 μg) were separated with sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. The membranes were blocked with 5% bovine serum albumin (BSA) at room temperature for 1 h. β-Catenin (Cell Signaling Technology, Danvers, MA), active β-catenin (Cell Signaling Technology), phospho-p44/42 mitogen-activated protein kinase (MAPK; Cell Signaling Technology), p44/42 MAPK (Cell Signaling Technology), β-actin (Sigma), Smad-5 (Cell Signaling Technology), and phopho-Smad-1/5/9 (Cell Signaling Technology) primary antibodies were added to a tris-buffered saline and Tween 20 (TBS-T) solution containing 5% BSA, and incubated with the membranes overnight at 4°C. After being washed with TBS-T thrice, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (Cell Signaling Technology) or anti-mouse IgG (Santa Cruz Biotechnology, Dallas, TX) in TBS-T containing 5% BSA for 1 h. Subsequently, the membranes were washed thrice with TBS-T buffer and analyzed with enhanced chemiluminescence detection reagents (Dogen, Korea).
Transplantation of BMP-2-producing BMSCs into calvarial defects in diabetic animals
Before in vivo cell transplantation, the autologous cells were thawed and expanded to passage two or three. Before 1 day of surgery, the cells were seeded at a density of 1 × 106 cells/dish and transduced with AdBMP2 according to their designated experimental group for the in vivo experiment. BMP-2 expression was confirmed by ELISA before the surgery. The cells were applied with 200 μL of a 1% collagen hydrogel, which was prepared from purified porcine skin-derived type I atelocollagen, sterile, lyophilized collagen (Matrixen-PSP; Bioland, Seoul, Korea), 0.001 N HCl, 26 mM NaHCO3, 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, and 0.025 N NaOH at pH 7.4.
The surgical procedure was performed while the animals were under general anesthesia. A midline incision was made over the calvarium and a full-thickness flap was elevated. An 8-mm, critical-sized calvarial defect was created using a trephine bur (3i Implant Innovation, West Farm Beach, Florida) under sterile saline irrigation. The animals were divided into the following six experimental groups: (1) negative control group—empty; (2) vehicle group—collagen hydrogel alone; (3) nBMSC group—collagen hydrogel containing autologous nBMSCs; (4) dBMSC group—collagen hydrogel mixed with dBMSCs; (5) B2/nBMSC group—collagen hydrogel containing autologous AdBMP2-transfected nBMSCs; and (6) B2/dBMSC group—collagen hydrogel containing autologous AdBMP2-transfected dBMSCs. All animals were treated with their own dBMSCs or nBMSCs. The incisions were sutured in layers with 5-0 chromic gut and 4-0 silk. All the animals received a single intramuscular injection of cefazolin (30 mg/kg) for 2 days after surgery. Six rats from each group were sacrificed at 4 weeks after surgery.
Micro-computed tomography analysis of the regenerated bone tissue
Tissue samples, including the surgical sites, were harvested and fixed in a 10% neutralized buffered formalin solution, and micro-computed tomography (CT) scans were obtained using a SkyScan 1172 (SkyScan, Kontich, Belgium).
The regenerated bone volume and other indexes were measured morphometrically using a computer program (CT-analyzer; SkyScan, Kontich, Belgium) with a lower gray threshold level of 65.28,29
Histological and histomorphometric analyses
The specimens were decalcified with a 10% EDTA solution for 2 weeks, dehydrated through a series of ethanol solutions of increasing concentration, and embedded in paraffin. Five-micrometer-thick coronal sections through the center of the circular defects were obtained and stained with hematoxylin and eosin. The prepared specimens were examined using light microscopy.
After microscopic examination, an image of each slide was captured using a digital camera, and the resulting images were saved to a computer for histomorphometric analysis. The area of newly formed bone (mm2) within the defect was outlined in the magnified histological images and calculated using an automated image analysis system (Tomoro Scope Eye 3.5 Image Analyzer; Techsan Digital Imaging, Seoul, Korea). The defect closure (percentage) was measured as the ratio of the area of newly formed bone divided by the area of the whole defect.
Statistical analysis
Triplicate results for each in vitro experiment were analyzed using paired t-tests to compare nBMSCs and dBMSCs with or without BMP2 gene delivery. ANOVA was performed with Tukey's post hoc test to evaluate the differences among the in vivo experimental groups; p-values less than 0.05 were considered statistically significant.
Results
Effects of diabetes on osteogenic activity and gene expression
Cell proliferation was not significantly influenced by diabetes or by BMP2 gene delivery (Fig. 2A). However, the ALP activity and Alizarin red S staining were significantly increased by diabetes, as shown by both B2/BMSCs and non-B2/BMSCs (Fig. 2B, C). Although the peak concentrations of BMP-2 secretion in both B2/dBMSCs and B2/nBMSCs were similar, BMP-2 production in B2/dBMSCs peaked from day 3 to 10, whereas BMP-2 production in B2/nBMSCs peaked from day 1 to 7 (Fig. 2D).
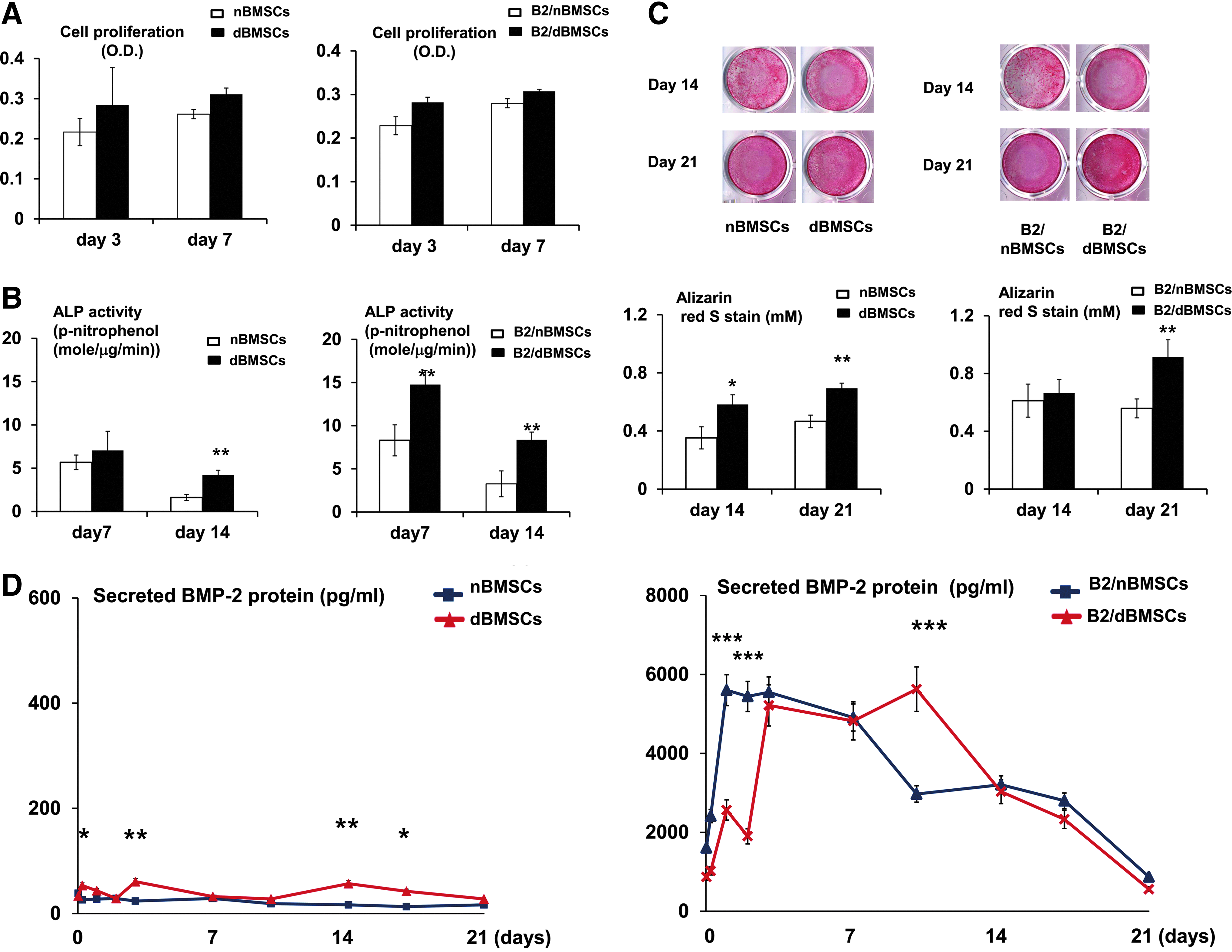
Effects of diabetes on the proliferation and osteogenic differentiation of B2/nBMSCs and B2/dBMSCs.
Diabetes significantly influenced the cellular BMP-2 mRNA expression patterns (Fig. 3A). BMP-2 mRNA expression in B2/nBMSCs showed a transient pattern, as the level of BMP-2 mRNA peaked at 24 h and decreased to the baseline level over 48 h. In contrast, B2/dBMSC BMP-2 mRNA expression gradually increased until 24 h and remained increased until 48 h. Although the level of BMP receptor type II (BMPRII) was not significantly different between cells, BMPRIA expression levels were coordinated with the BMP-2 mRNA expression (Fig. 3B, C). β-Catenin expression remained at low levels throughout the 48 h (Fig. 3D). Gremlin and noggin expression levels were increased in B2/dBMSCs and B2/nBMSCs in conjunction with BMP-2 expression (Fig. 3E, F).
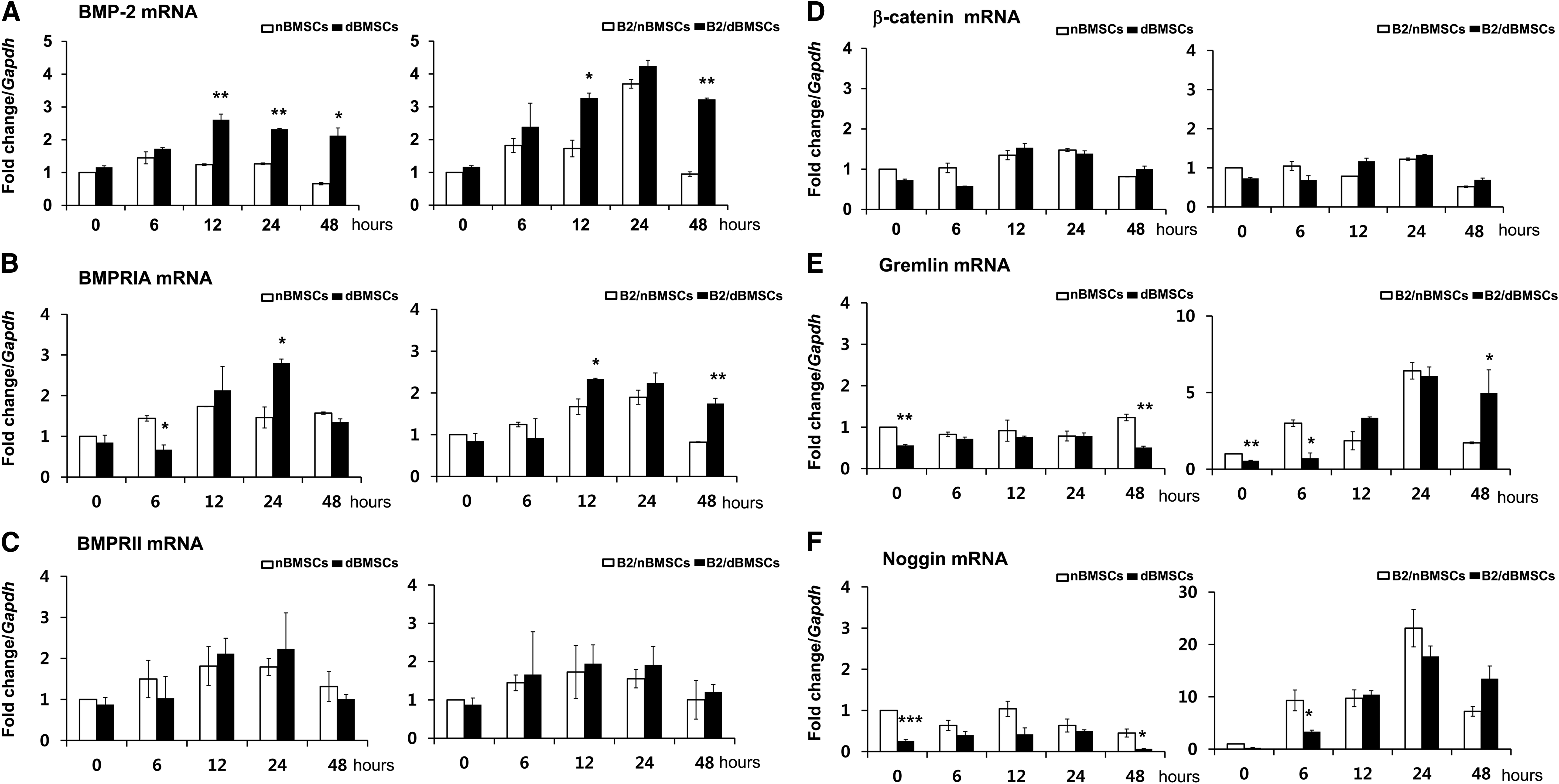
Effects of diabetes on gene expression related to BMP-2 signaling–qRT-PCR result up to 48 h after BMP2 gene delivery.
The expression pattern of osteogenic genes was also coordinated with BMP-2 mRNA expression. They were highly expressed until 48 h in B2/dBMSCs, whereas they transiently increased at 24 h and then decreased to the baseline level in B2/nBMSCs (Fig. 4A–D). Although the adipogenic gene expression of peroxisome proliferator-activated receptor (PPAR)γ was more suppressed in dBMSCs than in nBMSCs, it was significantly higher at 48 h in B2/dBMSCs than in B2/nBMSCs (Fig. 4E).
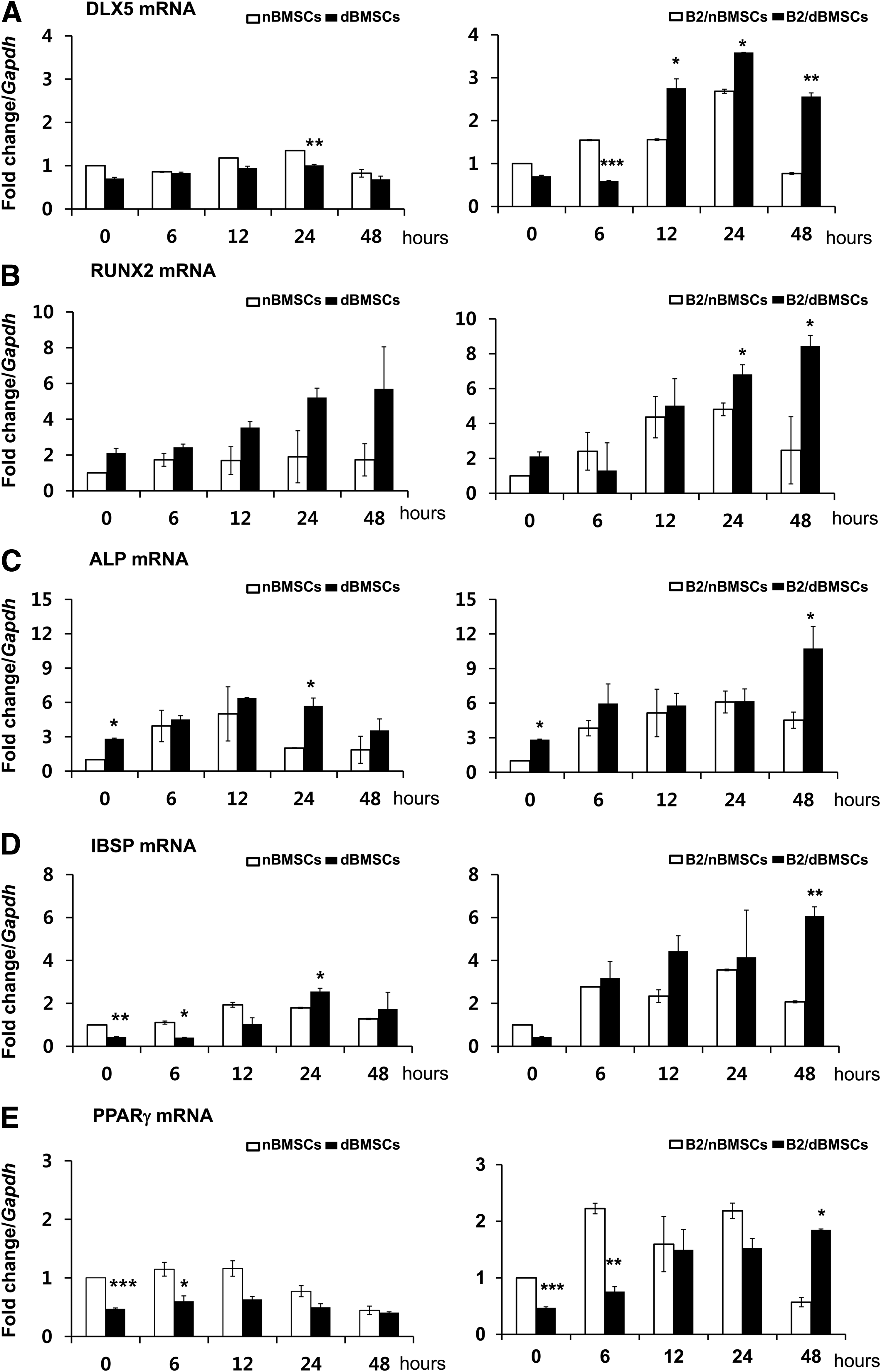
Effects of diabetes on gene expression related to osteogenic and adipogenic differentiation—qRT-PCR results up to 48 h after BMP2 gene delivery.
Signaling molecules related to gene expression changes in diabetes
The Western blot results showed that elements of the Wnt signaling pathway, including β-catenin and active β-catenin, were upregulated in diabetic cells compared with non-diabetic cells (Fig. 5). The p44/42 MAPK (ERK 1/2) pathway was also influenced by diabetes. While the BMP-2 signaling pathway, including Smad-1/5/9, was upregulated in both B2/dBMSCs and B2/nBMSCs, the upregulation was more intense in diabetic cells than in non-diabetic cells at 24 h.

Effects of diabetes on signaling molecules related to osteogenic differentiation.
Cell transplantation into calvarial defects of diabetic animals
Histologically, the B2/dBMSC group produced more new bone than did the B2/nBMSC group (Fig. 6A). However, the newly formed bone in both the B2/nBMSC and B2/dBMSC groups exhibited an immature, woven structure in the magnified images. The new bone formation in the nBMSC and dBMSC groups occurred slightly, near the margin of the defects. The negative control and vehicle groups showed limited bone regeneration and only in the margin of the defects (Supplementary Fig. S1).
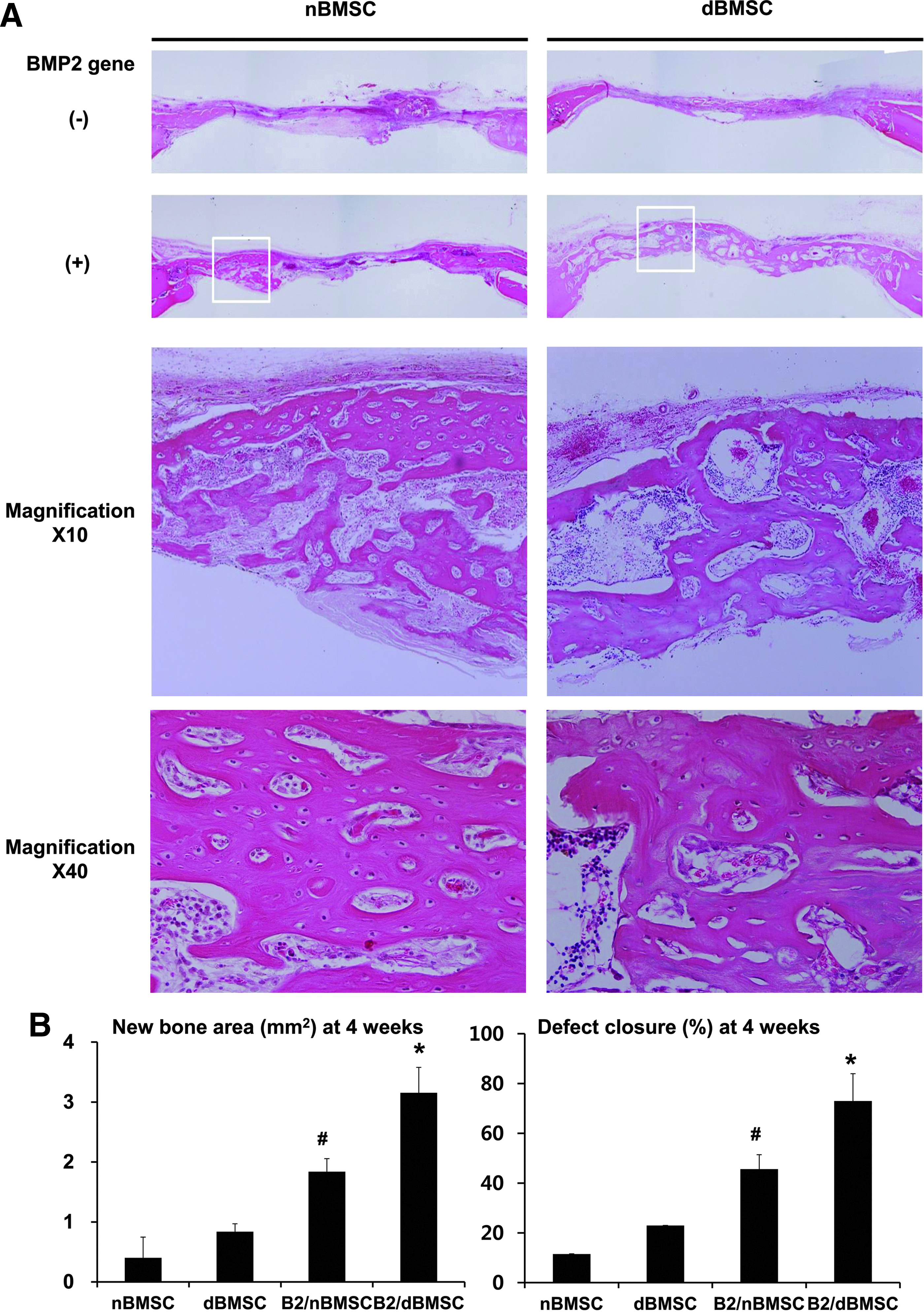
Transplantation of BMSCs into calvarial defects in diabetic animals—histological and histomorphometric analyses.
The histomorphometric analysis showed that the B2/dBMSC group produced a significantly greater area of newly formed bone than the other groups, including the B2/nBMSC group (Fig. 6B; p < 0.001). The average defect closure was 73% in the B2/dBMSC group and 46% in B2/nBMSC group, and this difference was significant (p < 0.001).
In the three-dimensional observation of the defects using micro-CT images, the newly formed bone was clearly demarcated due to its lower radiopacity compared with the normal bone. Similar to the histological examination, the B2/dBMSC group showed the most extensive formation of new bone, followed by the B2/nBMSC, dBMSC, and nBMSC groups (Fig. 7).
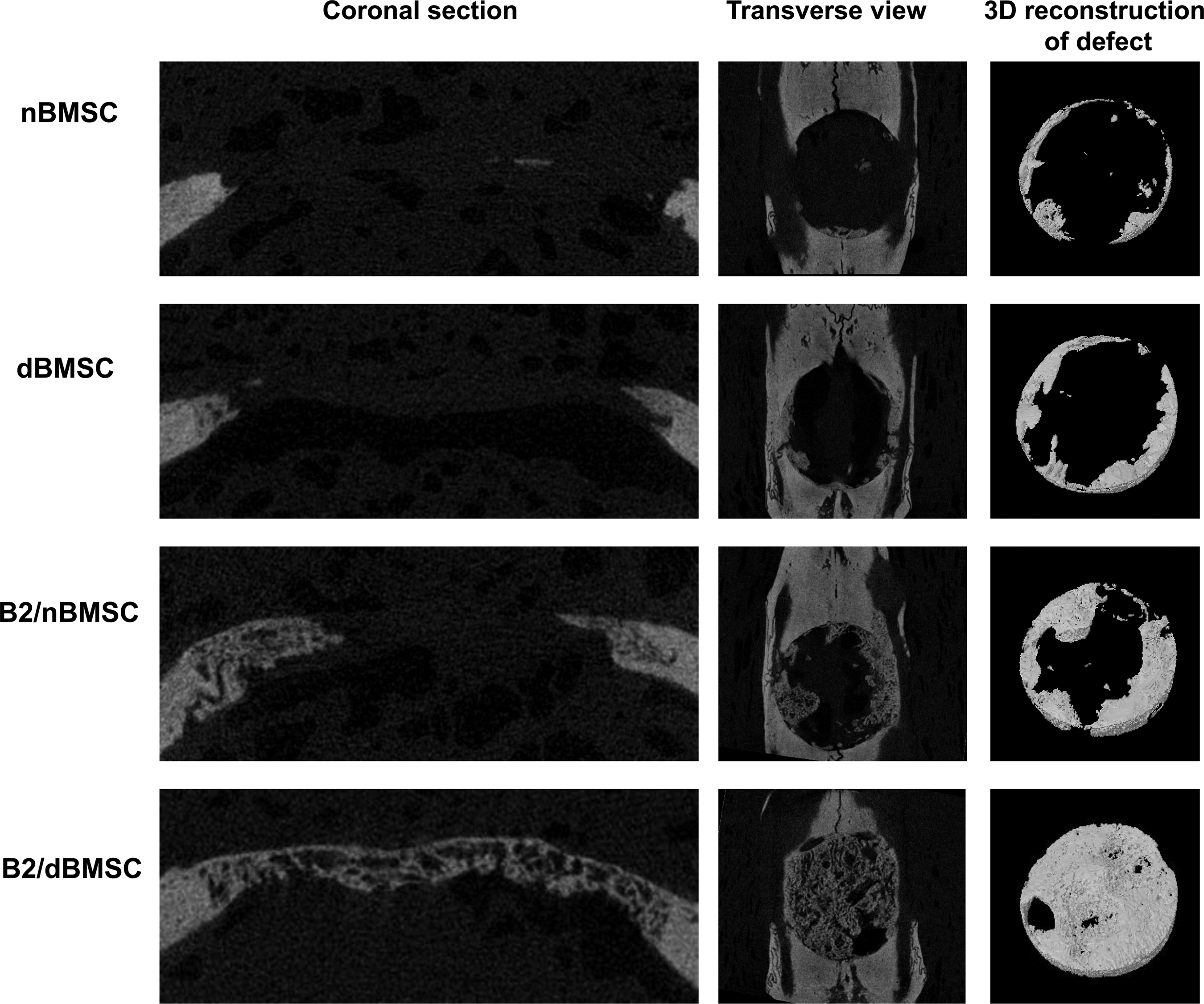
Micro-CT images from each group. The B2/dBMSC groups showed the most extensive bone formation compared with the other groups, and complete calvarial defect closure was observed in some animals. In all groups, the newly formed bone was clearly demarcated from the original bone. The B2/nBMSC group also showed enhanced bone regeneration compared with the nBMSC group, but complete defect closure was not observed. The nBMSC and dBMSC groups exhibited slight new bone formation at the defect border. Micro-CT images: left, coronal section view; middle, transverse view; right, 3D reconstruction of newly formed bone in the defect site. CT, computed tomography.
In the micro-CT analysis of the defects, the bone volume fraction was also significantly greater in the B2/dBMSC group than in the other groups (Fig. 8A; p = 0.002). According to measurements referring to connectivity, the newly formed bone in the B2/dBMSC group had a more well-connected trabecular structure than that in the other groups, whereas new bone in the nBMSC group exhibited less-connected, spongy lattices with a more rod-shaped trabecular structure (Fig. 8B–F). The mineral density of the newly formed bone ranged from 0.695 to 0.775 g HA/cm3, which was not statistically significant among the groups (Fig. 8H).
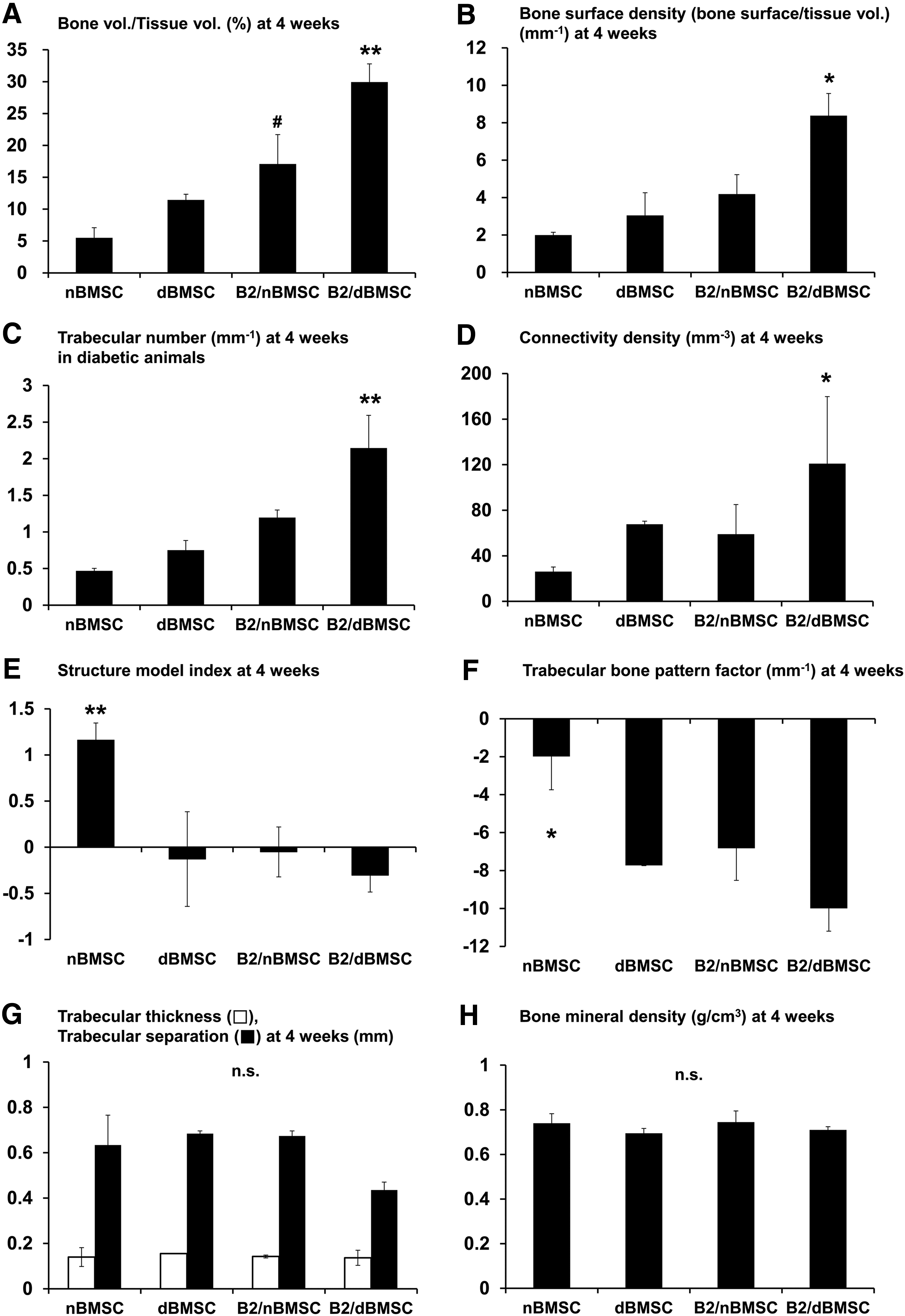
Micro-CT analysis of each group.
Discussion
This study showed that cell-based BMP2 gene delivery is effective for bone regeneration in diabetic animals. However, diabetic experiences of cells significantly influence the regenerative outcome. Cells harvested after the initiation of diabetes express more BMP-2 mRNA than cells harvested before diabetes initiation. As a result, B2/dBMSCs produce a significantly greater amount of newly formed bone than B2/nBMSCs.
Some previous studies have revealed that the cells from diabetic animals show reduced cell proliferation and osteogenic differentiation compared to age-matched healthy rats.19–21 Although a direct comparison to previous studies is impossible due to the differences in the protocols used, our observed increase in the osteogenic activity of dBMSCs remains a conflicting result. However, it is well known that osteoporosis and vascular calcification coexist in diabetes, although the mechanisms are unknown. Recently, Wnt/β-catenin is mentioned with diabetic vascular calcification. In general, functional Wnt/β-catenin transiently acts to maintain the stemness of stem cells in normoglycemic conditions. 30 In contrast, the hyperglycemic conditions of diabetes induce the activation of constitutive Wnt/β-catenin, which controls the osteogenic commitment and BMP-2 responsiveness of MSCs.31–35 Due to the continuous activation of the Wnt/β-catenin signaling pathway, MSCs in hyperglycemic conditions have a tendency to differentiate into osteoblasts, which has been suggested to be responsible for the development of pathological vascular calcification in diabetic patients. 34 Fowlkes et al. suggested that diabetic bone impairment is related to downregulation of the DLX5-RUNX2 pathway, not the BMP-2 pathway. 34 Similarly, the dBMSCs in this study also showed an increase in the Wnt/β-catenin pathway accompanied by increases in ALP, IBSP, and BMPRIA gene expression and a decrease in DLX5 expression at 24 h. In addition, B2/dBMSCs showed upregulation of the Wnt/β-catenin pathway and the BMP-2/Smad signaling pathway. Therefore, osteogenic stimulation by BMP-2 on dBMSCs was not hindered in the diabetic condition, but rather strengthened by the Wnt/β-catenin pathway.
Using adenoviral vectors to deliver the BMP2 gene, B2/dBMSCs produced more BMP-2 mRNA than B2/nBMSCs in vitro. Although B2/nBMSCs transiently expressed BMP-2 mRNA over 48 h, the B2/dBMSCs continuously produced BMP-2 mRNA for more than 48 h. In general, genes introduced into cells through adenoviral delivery tend to become transcriptionally silent after short periods of expression, although the extrinsic gene remains within the transduced cells. Although the exact mechanism underlying this effect is unknown, covalent linkage to bacterial DNA by methylation has been proposed as a potential explanation. 35 Brooks et al. found that genes delivered into skeletal muscle by adenoviruses became extensively methylated as early as 24 h after delivery; de novo methylation is a cellular defense mechanism against the transcription of foreign genes.35,36 However, alterations in the DNA methylation process are frequently observed in diabetes because the DNA methylation process changes in response to glucose concentration.37,38 In other words, healthy cells have their own defense mechanism against the invasion of a foreign gene, whereas diabetic cells are vulnerable to foreign invasion. As a result, BMP2 gene delivery to diabetic cells amplified the transcription of BMP-2 mRNA, leading to enhanced bone regeneration in vivo.
In vivo, the regenerative potential of B2/nBMSCs was similar in both diabetic and healthy rats. The amount of newly formed bone area was similar to that found in our previous study performed in healthy animals. 15 However, the newly formed bone in the diabetic and healthy animals exhibited different levels of maturity. The remodeling process from woven bone to lamellar bone was incomplete in diabetic animals. Different from lamellar bone, woven bone is the most disorganized bone tissue and is known to be more brittle and less stiff than other bone tissues. 39 Therefore, delayed bone turnover could be a possible mechanism of impaired bone healing in diabetes. 40 Some studies have suggested that insufficient amounts of insulin and such growth factors as insulin-like growth factor 1 and platelet-derived growth factor (PDGF) might be related to impaired diabetic healing, which was improved by the application of these molecules in animal studies.41–44 Our previous study also showed the potential of PDGF-BB for improving the quality of BMP-2-induced bone. 15 Accordingly, combination therapy using BMP-2 and other growth factors is recommended to improve diabetic bone healing.
There are some limitations related to this study. The cells in this study were not age-matched cells and the differences between dBMSCs and nBMSCs can be related to the age of animals, although ages 8–18 weeks of the rats were puberty and were before adulthood, and they both correspond to the young age. In addition, we used cell harvesting methods modified from bone marrow ablation procedures to keep the bone structure and function intact. The bone marrow ablation model has been used to investigate mechanisms through which factors influence intramembranous bone formation.45,46 The effect of bone marrow ablation is reported to be transient within 10 days and diminished after 56 days in previous study.47,48 In this study, the time interval between cell harvesting was 10 weeks and the effects seems to be theoretically minimal, but its effects could not be ignored.
In conclusion, BMP-2 delivery using BMP2 gene therapy effectively induces de novo bone formation in a critical-sized calvarial defect during diabetic bone healing. Analysis of the regenerative outcome revealed that BMP2 gene delivery using dBMSCs is advantageous for bone regeneration. B2/dBMSCs showed an increase in BMP-2 expression, and the enhanced osteogenic activity detected was mediated by upregulation of Wnt/β-catenin and BMP-2/Smad signaling. However, BMP2 gene delivery alone did not overcome the impairment in diabetic bone healing, and further strategies to improve the quality of bone in diabetes are needed.
Footnotes
Acknowledgments
The authors are grateful to Ms. Eun-Hye Gwak (Department of Periodontology, School of Dentistry, Seoul National University, Seoul, Korea) for providing technical help with Western blotting assays in this study. This research was supported by the Bio & Medical Technology Development Program of the NRF, which is funded by the Korean government, MSIP (NRF-2014M3A9E3064466) and also supported by Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education (NRF-2014R1A1A2058346).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.