
Abstract
Bone marrow-derived mesenchymal stromal stem cells (BMSCs) are a promising cell source for treating articular cartilage defects. The objective of this study was to assess a protocol that involved autologous transplantation of BMSCs into full-thickness cartilage defects in sheep following isolation, expansion, and a short period (4 days) of chondrogenic priming. The impact of oxygen tension during preimplantation culture was investigated. It was hypothesized that chondrogenically primed BMSCs would produce superior cartilaginous repair tissue relative to control defects, and that culture under hypoxia would yield improved repair tissue in comparison to normoxia. Ovine BMSCs were isolated, expanded to passage 2, seeded within a hyaluronic acid (HYAFF) scaffold, and primed ex vivo in chondrogenic medium for 4 days under normoxia (21% oxygen) or hypoxia (3% oxygen). Full-thickness, 7-mm-diameter articular cartilage defects were created in the femoral condyles of five sheep. Twenty defects were treated with normoxia-cultured, autologous BMSC-seeded scaffolds (eight); hypoxia-cultured, autologous BMSC-seeded scaffolds (eight); cell-free scaffolds (two); or no implants (two). Preimplantation priming was evaluated through gene expression analysis using reverse transcription quantitative polymerase chain reaction. After 6 months, histological assessment was performed on repair tissues with a modified O'Driscoll scoring system and tissue dimension analysis. Priming of preimplantation BMSC-seeded scaffolds in chondrogenic medium for 4 days resulted in significantly increased gene expression of hyaline cartilage-related collagen II and aggrecan relative to unprimed BMSCs (p < 0.05). Defects implanted with chondrogenically primed BMSC-seeded scaffolds developed cartilaginous repair tissues that contained safranin O-positive proteoglycans, and had significantly larger repair tissue areas, higher percentages of defect fill, and improved histological scores than cell-free controls (p < 0.05). Although hypoxic culture improved the preimplantation gene expression profile, a consistent difference in histological scores was not found between normoxia- and hypoxia-seeded BMSC-seeded scaffolds after 6 months (p = 0.90). This study demonstrates in a sheep model that (1) chondrogenic priming ex vivo improves the gene expression profile of BMSCs; (2) chondrogenically primed BMSCs are associated with the development of superior cartilaginous tissue to cell-free controls within cartilage defects; and (3) oxygen tension during preimplantation ex vivo culture does not consistently modulate cartilaginous repair tissue formation following BMSC transplantation into cartilage defects.
Introduction
B
Several BMSC transplantation protocols have been proposed to date with no clear consensus on technique established. 1 One protocol that has been assessed in a clinical realm involves implantation of bone marrow aspirate concentrate (BMAC), which contains a heterogeneous bone marrow-derived cell collection that includes BMSCs, on a biomaterial scaffold into a focal cartilage defect.11–13 This protocol allows for aspiration, processing, and implantation to occur during the same operative period and avoids the need for ex vivo cell culture. 12 However, the number of BMSCs present in each concentrate is not known at the time of implantation. Furthermore, there is evidence that the BMSC content of concentrates varies between donors and depends on the processing system used.18,19 It is therefore unclear whether the implanted concentrate contains an adequate number of BMSCs to support the development of hyaline-like cartilaginous repair tissue. Tissue consistent with fibrocartilage has been found in some defects on histological analysis and MRI in preclinical and clinical studies following transplantation of heterogeneous cell collections.11–13,17,20
A second clinical protocol entails isolation and expansion of BMSCs in the laboratory and implantation on a scaffold into a focal cartilage defect.9,10 Although the BMSCs in this protocol are enriched during the isolation process, they are undifferentiated and there is only limited evidence to suggest that they are capable of producing ECM consistent with hyaline cartilage rather than fibrocartilage or bone.10,21–23 Undifferentiated BMSCs are predisposed to entering an osteogenic pathway following sustained expansion. 21
A third protocol that has been investigated in preclinical models involves creation of a tissue-engineered construct developed through ex vivo culture of a BMSC-seeded scaffold for 2–3 weeks to allow for differentiation and ECM production before implantation within a focal cartilage defect.24–27 Although the transplanted construct contains cells that may be inclined to continue down a chondrogenic lineage, chondrogenic predifferentiation prolongs the culture period and may be time and resource intensive relative to other protocols. 25 Furthermore, there is in vitro evidence to suggest that prolonged predifferentiation periods may not be required to induce chondrogenesis. 28
The first objective of this pilot study was to assess a protocol for BMSC transplantation that involved BMSCs that were isolated, expanded, seeded within esterified hyaluronic acid (HYAFF) scaffolds, and chondrogenically primed using a short (4-day) culture period in chondrogenic medium before autologous implantation in an ovine model. The chondrogenic priming period of 4 days was shorter than previously described chondrogenic culture periods of 10–21 days used within in vivo studies that involved transplantation of cells into cartilage defects in animals24–27 and subcutaneous tissues in nude mice.29–31 The rationale of this shortened protocol was to expose BMSCs to chondrogenic factors with the goal of predisposing them to the chondrogenic lineage while avoiding a time- and resource-intensive differentiation period. It was hypothesized that chondrogenically primed, autologous BMSC-seeded HYAFF scaffolds implanted into full-thickness cartilage defects would lead to the development of superior cartilaginous repair tissue relative to control defects.
Tissue engineering strategies have been employed within transplantation protocols with the goal of promoting BMSC chondrogenesis and hyaline-like cartilaginous ECM formation. Incubator oxygen tension is a culture variable that has gained attention given the posited role of oxygen in guiding BMSC differentiation in musculoskeletal development. 32 Our group and others have established extensive in vitro support for the use of hypoxic incubation to drive BMSC chondrogenesis.33–38 Hypoxic culture has been shown to augment chondrogenesis following subcutaneous implantation in mouse models.39,40 However, preimplantation hypoxic culture has not been investigated in BMSCs implanted within joints in animals or humans to date.
The second objective of this study was to assess the impact of oxygen tension during preimplantation culture of BMSCs on neo-cartilage formation in vivo within full-thickness cartilage defects. It was hypothesized that BMSC isolation, expansion, and chondrogenic priming under hypoxia would yield improved cartilaginous repair tissue in comparison to normoxia.
Materials and Methods
Ethical approval
The Animal Care and Use Committee at the University of Alberta provided ethical approval for this study. All methods were performed in accordance with the relevant guidelines and regulations.
Oxygen tension
Oxygen tension was defined relative to standard incubator conditions. Normoxic culture was performed within a humidified incubator containing 21% oxygen and 5% carbon dioxide with a temperature of 37°C. Hypoxic culture was performed in a separate humidified incubator containing 3% oxygen and 5% carbon dioxide with a temperature of 37°C.
BMSC isolation, expansion, and priming
Bone marrow aspirates were obtained from the iliac crest of five female Suffolk sheep with mean age ± standard error of the mean (SEM) of 2.6 ± 0.3 years and mean mass ± SEM of 75.0 ± 5.6 kg (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/tea). The general anesthesia and aspiration procedures are described in Supplementary Materials and Methods S1. Bone marrow aspirate was collected and mixed with 8 mL of heparin (10,000 units/10 mL; Pharmaceutical Partners of Canada, Richmond Hill, Canada). Each aspirate was then filtered with a cell strainer (100-μm pore size; Becton Dickinson Canada, Mississauga, Canada). Staining with crystal violet (Sigma-Aldrich, Oakville, Canada) and hemocytometer counting were used to determine the number of bone marrow mononucleated cells (BMNCs) within each aspirate. Bone marrow aspirates containing 8 × 107 BMNCs were seeded within each 150-cm2 tissue culture plastic flask and submersed in expansion medium containing fibroblast growth factor two (Supplementary Materials and Methods S2). BMSCs were isolated from these aspirates and expanded to passage 2 (P2) under normoxia (21% oxygen) or hypoxia (3% oxygen) using the protocol described in Supplementary Materials and Methods S2. The BMSCs produced by this protocol were previously shown to display multilineage differentiation potential and are capable of producing tissue consistent with cartilage, bone, and fat.7,37
Expanded BMSCs were seeded at 1 × 107 BMSCs/cm3 onto scaffolds composed of nonwoven HYAFF fibers (10–20 μm fiber diameter, varying inter-fiber spaces, 2 cm × 2 cm × 0.2 cm scaffold dimensions; Anika Therapeutics S.r.l., Abano Terme, Italy) and cultured statically for 4 days in chondrogenic medium containing dexamethasone and transforming growth factor-beta three (TGF-β3) within a humidified incubator containing an oxygen tension consistent with the oxygen tension used for isolation and expansion (Supplementary Materials and Methods S3).
Assessment of proliferation and population doubling
Proliferation during expansion was assessed via cell counts. Total cell counts of trypsinized BMSCs at P0, P1, and P2 were calculated using trypan blue (Sigma-Aldrich) staining and hemocytometer counting of small aliquots of BMSCs in expansion medium. The number of BMSCs isolated in each flask containing 8 × 107 BMNCs was extrapolated from a colony-forming unit fibroblastic assay previously described. 37 Population doubling was determined using the method described by Solchaga et al. 41
Assessment of chondrogenic priming
BMSC-seeded scaffolds that were cultured for 4 days in chondrogenic medium to induce chondrogenic priming were assessed with reverse transcription quantitative polymerase chain reaction (RT-qPCR) to quantify gene expressions of aggrecan, collagen II, and SOX9 that are related to hyaline cartilage, gene expression of collagen I that is related to fibrocartilage, and collagen X that is related to hypertrophic cartilage. These gene expressions were compared to unprimed BMSCs that were not exposed to chondrogenic medium and BMSCs that were seeded at 1 × 107 BMSCs/cm3 onto HYAFF scaffolds and cultured in chondrogenic medium for 14 days to induce chondrogenic differentiation.
To perform gene expression analysis, total ribonucleic acid (RNA) was extracted from cell-seeded scaffold constructs using TRIzol Reagent (Life Technologies) after grinding with a pestle (Geno Technology, St. Louis). Total RNA (100 ng) in a 40-μL reaction was reverse transcribed to complementary deoxyribonucleic acid (cDNA) using GoScript Reverse Transcription System (Promega, Madison) primed in the presence of oligo(dT) primers (1 μg). qPCR was performed with a DNA Engine Opticon I Continuous Fluorescence Detection System (Bio-Rad) using HotGoldStar Taq polymerase and SYBR Green detection (Eurogentec North America, San Diego). Custom-designed primers (Supplementary Table S2) using Primer Express software (Applied Biosystems, Foster City) were obtained from Invitrogen (Life Technologies). 37 Gene (messenger RNA [mRNA]) expression levels for each primer set were normalized to the expression level of ovine beta-actin (β-actin) by the 2−ΔΔC(T) method. These gene expressions were compared to unprimed BMSCs that were not exposed to chondrogenic medium and BMSCs that were seeded at 1 × 107 BMSCs/cm3 onto HYAFF scaffolds and cultured in chondrogenic medium for 14 days to induce chondrogenic differentiation. To ensure that the reference gene, β-actin, was not modulated by oxygen tension, the threshold cycles for normoxia- and hypoxia-primed BMSCs were compared and confirmed to be not significant (p = 0.30).
BMSC-seeded scaffolds that were cultured for 4 days in chondrogenic medium to induce chondrogenic priming were also assessed biochemically for proteoglycan production through the quantification of sulfated glycosaminoglycan (GAG) per DNA (GAG/DNA). To perform this biochemical analysis, constructs were rinsed in phosphate-buffered saline and digested in proteinase K (1 mg/mL in 50 mM Tris with 1 mM ethylenediaminetetraacetic acid [EDTA], 1 mM iodoacetamide, and 10 mg/mL pepstatin A; all from Sigma-Aldrich) for 16 h at 56°C. Sulfated GAG content was measured by 1,9-dimethylmethylene blue binding (Sigma-Aldrich) using chondroitin sulfate as the standard (Sigma-Aldrich). De novo sulfated GAG production was determined by subtracting GAG quantities of BMSCs that were not exposed to chondrogenic medium from GAG quantities in constructs that were primed for 4 days or differentiated for 14 days in chondrogenic medium. DNA content was determined using the CyQUANT Cell Proliferation Assay Kit (Life Technologies) with supplied bacteriophage λ DNA as the standard.
Joint surgery and treatment groups
Sheep were positioned in supine position following general anesthesia. The surgical approach is described in detail in Supplementary Materials and Methods S1. A midline skin incision was made and parapatellar windows were used to access the medial and lateral condyle of each stifle joint. A 6.95 mm-diameter, full-thickness articular cartilage defect was created in the weight-bearing zone of each condyle. An autologous BMSC-seeded scaffold or cell-free scaffold (control) was cut with a biopsy punch to create a 7 mm-diameter circular implant (Fig. 1A). Tisseel fibrin sealant (Baxter, Mississauga, Canada) was used to secure each implant. A defect that was treated with Tisseel fibrin sealant alone without implantation of cells or a scaffold served as another control.

BMSC ex vivo culture and transplantation.
The treatment groups in this study included cell-free controls (with and without scaffold), normoxia-cultured BMSC-seeded scaffolds and hypoxia-cultured BMSC-seeded scaffolds (Fig. 1B). Four defects in two stifle joints were treated as cell-free controls by leaving defects empty or implanting cell-free scaffolds. Eight defects in four stifle joints were treated with normoxia-cultured, autologous BMSC-seeded scaffolds. The BMSCs in this group were isolated, expanded, and chondrogenically primed under normoxic conditions. Eight defects in four other stifle joints were treated with hypoxia-cultured, autologous BMSC-seeded scaffolds. The BMSCs in this group were isolated, expanded, and chondrogenically primed under hypoxic conditions. Both defects in each joint received the same cell condition of no cells (control), normoxia-cultured BMSCs, or hypoxia-cultured BMSCs to avoid interaction of one treatment group with another.
In vivo reassessment of implants and explantation
To ensure that the BMSC-seeded scaffolds were capable of remaining in the defect following implantation, sheep Z01 was reassessed in a second-look procedure 1 week after the initial surgery (Supplementary Materials and Methods S1). Six months after the transplantation surgery, sheep were sacrificed using sodium pentobarbital (WDDC) and the stifle joints were extracted for processing.
Histological processing and assessment of repair tissue
Processing for histological analysis involved reducing each condyle to osteochondral blocks centered on the defect with 5–10 mm of native articular cartilage surrounding the defect on all sides. Each specimen was fixed using 10% wt/vol buffered formalin for 7 days (Sigma-Aldrich). Thereafter, decalcification was performed for four months in a solution containing 10% wt/vol EDTA tetrasodium salt dehydrate and 0.1% wt/vol formalin followed by two months in a solution containing 20% wt/vol EDTA tetrasodium salt dehydrate and 0.1% wt/vol formalin (Sigma-Aldrich). Specimens were then dehydrated, processed into paraffin wax and sectioned at a thickness of 8 μm in a longitudinal plane. Sections were stained with 0.1% wt/vol safranin O to reveal proteoglycan matrix depositions and counterstained with 1% wt/vol fast green. Microphotographs of each section were captured using an Eclipse Ti-S microscope (Nikon Canada, Mississauga, Canada) fitted with NIS Elements Basic Research Imaging Software Version 4.20 (Nikon Canada). The microscope automatically stitched magnified photomicrographs into larger images containing the entire defect, repair tissues, and native tissues. Images were subsequently reoriented in Photoshop (Adobe Systems, San Jose). For histological assessment (Supplementary Materials and Methods S4) microphotographs were selected, coded, placed in random order, and assessed by two blinded observers (T.D.B. and N.M.J.) using a modified version of the O'Driscoll histological scoring system (Supplementary Materials and Methods S5 Table 1 and Supplementary Materials and Methods S5).42–44 Averages of total scores and subscores from both observers were used for statistical analysis. Microphotographs were also assessed in a blinded fashion for quantitative measures using ImageJ software (National Institutes of Health, Bethesda).
Macroscopic assessment of repair tissue
Photographs of each defect were coded, placed in random order, and assessed by the same two blinded observers using a modified Goebel macroscopic scoring system (Supplementary Materials and Methods S6 Table 1 and Supplementary Materials and Methods S6). 45 Averages of total scores and subscores from both observers were used for statistical analysis.
Statistical analysis
Statistical analysis was performed using SPSS Statistics 23 (IBM, Armonk) and significance was concluded when p < 0.05. All data were subjected to Levene's test to assess for homogeneity of variance and Shapiro–Wilk test to assess for normality of distribution. For parametric analysis, a t-test was used for comparison of two groups, while a one-way analysis of variance (ANOVA) was used for comparison of three or more groups. Nonparametric analysis was performed when either Levene's test or Sharpiro–Wilk test was significant. A Mann–Whitney U test was used for comparison of two groups and a Kruskal–Wallis ANOVA was used for comparison of three or more groups. A detailed description of statistical analysis is found within Supplementary Materials and Methods S6.
Results
BMSC isolation and expansion
Bone marrow aspirates with a mean volume ± SEM of 32 ± 2 mL were collected from five ovine donors. BMSCs were isolated by plastic adherence and expanded in tissue-culture flasks under normoxic or hypoxic conditions (Fig. 1A). Cell count, cell population doublings per day, and cumulative population doublings were not significantly different between oxygen tension groups over the course of the expansion period (p = 0.11, 0.40, and 0.79; Fig. 2A–C). Cell count increased progressively with each passage (p < 0.001; Fig. 2A). Once 80% confluence was reached at P2, there were 16.1 ± 2.7 × 106 BMSCs derived from each flask seeded and cultured under normoxia and 18.7 ± 4.3 × 106 BMSCs derived from each flask seeded and cultured under hypoxia (p = 0.16). Population doublings per day were fastest at P0 and decreased with passage thereafter (p = 0.03; Fig. 2B). Cumulative population doublings by the end of the expansion period were 13.1 ± 0.5 doublings for normoxia-expanded BMSCs and 13.0 ± 1.1 doublings for hypoxia-expanded BMSCs (p = 0.86; Fig. 2C).
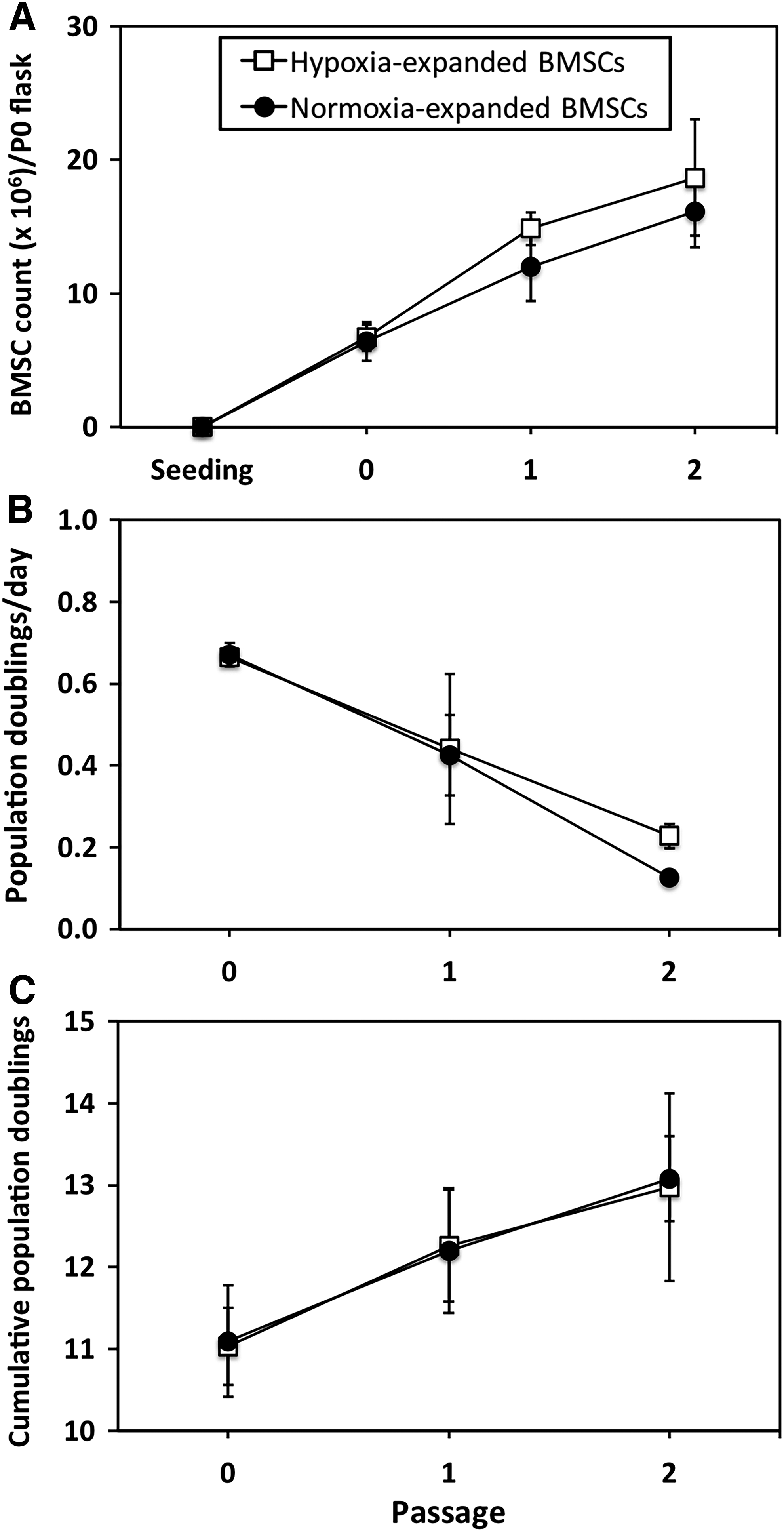
Expansion of BMSCs.
Chondrogenic priming of BMSCs
Expanded BMSCs were seeded onto HYAFF scaffolds, and cultured for 4 days ex vivo in chondrogenic medium containing TGF-β3 and dexamethasone to induce chondrogenic priming. This 4-day chondrogenic priming period resulted in significantly increased hyaline cartilage-related aggrecan and collagen II gene expressions of BMSCs relative to unprimed controls (p < 0.05; Fig. 3A, B). Expressions of these genes after 4 days of priming were not different than after a longer course (14 days) of chondrogenic differentiation (p ≥ 0.67). There was a trend for sex determining region Y-box nine (SOX9) gene expression to increase following priming although a significant difference was not found between time points (p = 0.10 for normoxia- and 0.13 for hypoxia-cultured BMSCs; Fig. 3C). Fibrocartilage-related collagen I and hypertrophic cartilage-related collagen X gene expression were significantly augmented after 4 days of chondrogenic priming under normoxia in comparison to unprimed controls (p < 0.05), while expressions of these genes were not significantly increased after 4 days of chondrogenic priming under hypoxia (p = 0.34 and 0.39; Fig. 3D, E). Although the 4-day culture period was shown to prime BMSCs through augmentation of genes associated with chondrogenesis, proteoglycan production that was seen after 14 days of chondrogenic differentiation was not yet established after 4 days of chondrogenic priming based on GAG/DNA (Fig. 3H).
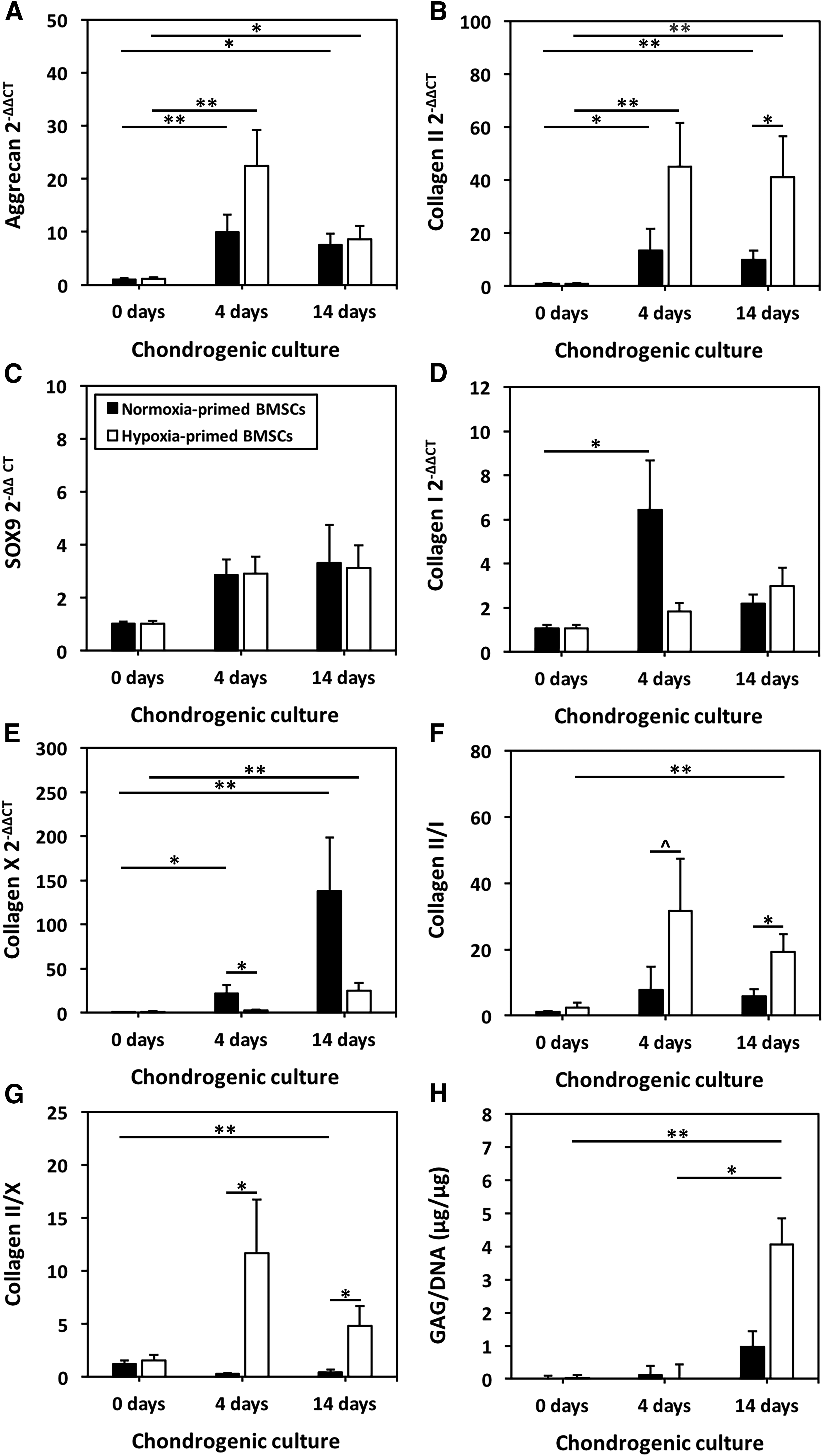
Chondrogenic priming of BMSC-seeded scaffolds. BMSCs were isolated and expanded under either normoxia or hypoxia and not exposed to chondrogenic medium (0 days) or seeded onto HYAFF scaffolds and either primed in chondrogenic medium for 4 days or differentiated in chondrogenic medium for 14 days within the same oxygen condition as isolation and expansion. Thereafter,
Oxygen tension during culture modulated gene expressions of BMSCs seeded within HYAFF scaffolds after 4 days of chondrogenic priming. The increase in collagen II gene expressions between 0 and 4 days of culture in chondrogenic medium was more significant in hypoxia-cultured BMSCs (p = 0.004) than normoxia cultured BMSCs (p = 0.01; Fig. 3B). However, aggrecan and collagen II gene expressions were not significantly different in a direct comparison of priming for 4 days under hypoxia and normoxia (p = 0.22 and 0.12; Fig. 3A, B). Collagen X gene expression was significantly lower when priming was performed under hypoxia versus normoxia (p = 0.03; Fig. 3E). The ratio of collagen II-to-X expression was significantly increased with hypoxic priming relative to normoxic priming after 4 days (p = 0.03; Fig. 3G), while the ratio of collagen II-to-I expression approached significantly higher levels under hypoxia in comparison to normoxia (p = 0.09; Fig. 3F).
Postoperative course following transplantation surgery
Following joint surgery for BMSC transplantation, sheep returned to full activity. One week from the time of surgery, a second-look procedure was performed on sheep Z01 to assess the position of each implant. BMSC-seeded scaffolds were noted to be in place within the femoral condyle cartilage defects. None of the sheep displayed signs of postoperative infection or lameness during the 6-month post-transplantation period.
Histological assessment of repair tissue
Femoral condyle defects were assessed histologically 6 months after BMSC implantation surgery. Full resurfacing of defects with hyaline-like cartilaginous repair tissue that stained strongly with safranin O occurred in some cases following transplantation of chondrogenically primed BMSCs (Fig. 4B, J, K). Other defects treated with BMSC transplantation were partially filled with heterogeneously stained repair tissue that resembled mixed tissue or fibrocartilage (Fig. 4C, L) while other defects contained predominantly fibrous tissue (Fig. 4D–F, M). Defects that received cell-free treatment were empty (Fig. 4D, G) or partially filled with mixed repair tissue (Fig. 4A; Supplementary Fig. S1A, C).
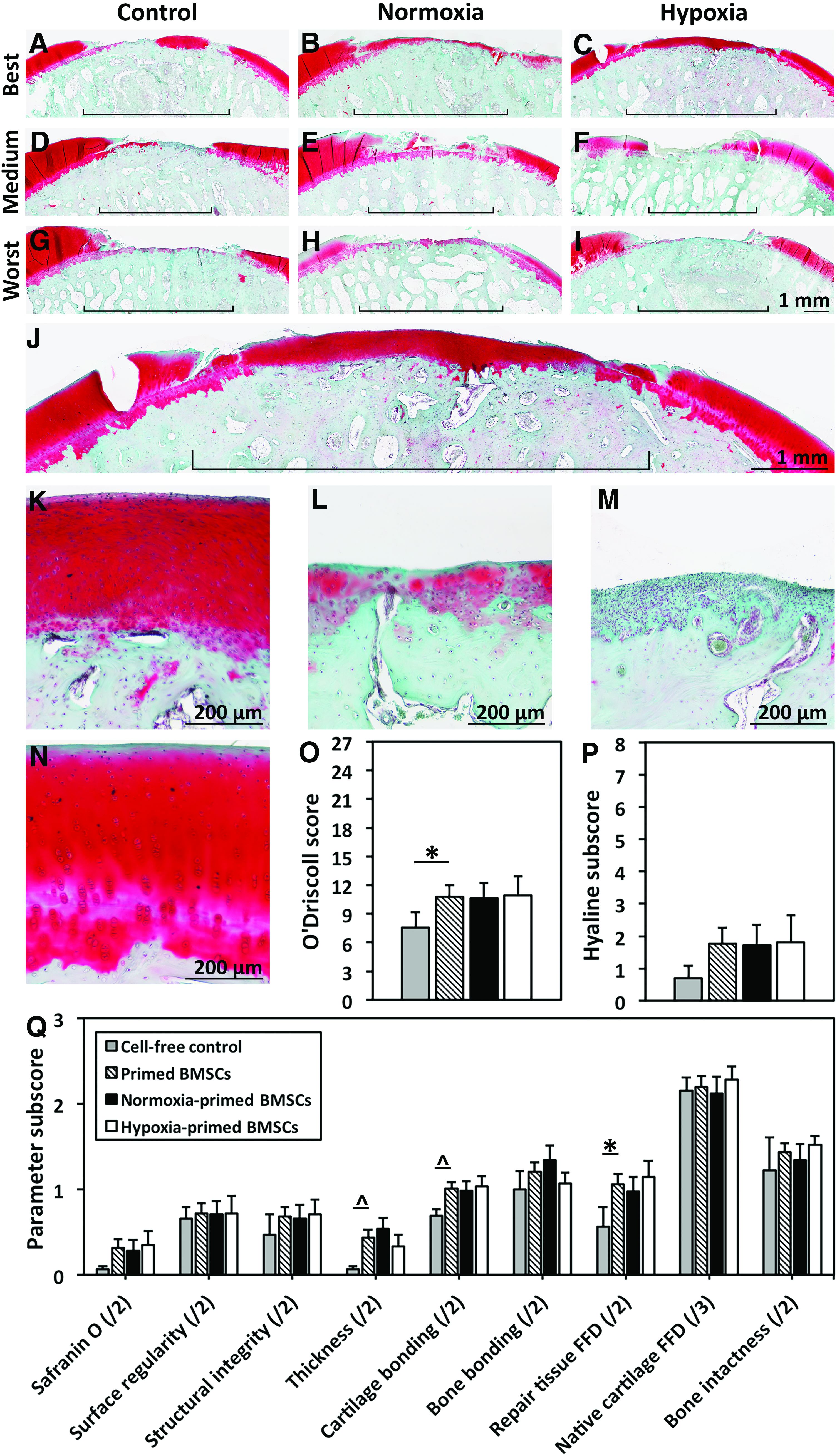
Histological staining and scoring of cartilaginous repair tissue. Six months following transplantation, femoral condyles were explanted, processed, sectioned at 8-μm thickness, and stained with safranin O and fast green.
Repair tissue quality was assessed with a modified O'Driscoll scoring system (Supplementary Materials and Methods S5 Table 1). In a comparison of all BMSC-seeded scaffolds—that included a pooled group of all scaffolds seeded with either hypoxia- or normoxia-cultured BMSCs (Fig. 4O)—and cell-free controls, defects implanted with BMSC-seeded scaffolds had significantly higher modified O'Driscoll scores (mean ± SEM of 10.8 ± 1.1, range of 4.5–23.0) than cell-free controls (7.6 ± 0.9, range of 4.5–12.5, p = 0.04; Fig. 4O). Subscores for freedom from degeneration in repair tissue were significantly higher in defects filled with BMSC-seeded scaffolds than cell-free control defects (p = 0.04), while subscores for repair tissue thickness and bonding of repair tissue to native cartilage approached significantly higher levels in defects filled with cell-seeded scaffolds than control defects (p = 0.08 for both; Fig. 4Q).
Modified O'Driscoll scores were not significantly different in the comparison of cell-free controls (7.6 ± 0.9, range of 4.5–12.5), normoxia-cultured BMSC-seeded scaffolds (10.6 ± 1.4, range of 4.5–21.8), and hypoxia-cultured BMSC-seeded scaffolds (10.9 ± 1.8, range of 5.0–23.0; p = 0.41 between all groups; p = 0.90 between normoxia- and hypoxia-cultured BMSC-seeded scaffolds; Fig. 4O). Differences between these groups in subscores for repair tissue thickness and freedom from degeneration in repair tissue approached significance (p = 0.09 for both). In the direct comparison of normoxia- and hypoxia-cultured BMSC-seeded scaffolds within each of three sheep (Y08, Z01, and Z33) that received normoxia-cultured BMSCs in one joint and hypoxia-cultured BMSCs in the contralateral joint, modified O'Driscoll scores were significantly higher following treatment with hypoxia-cultured BMSCs in two of three animals (p < 0.05; Supplementary Fig. S1K).
Quantitative analysis of tissue dimensions demonstrated that the area of all forms of repair tissue was significantly larger within defects implanted with BMSC-seeded scaffolds than cell-free controls (p = 0.003; Fig. 5A). Hyaline-like cartilaginous tissue area approached significantly larger levels in defects implanted with BMSC-seeded scaffold than cell-free controls (p = 0.07; Fig. 5A). Percentage of defect fill was significantly higher in BMSC-seeded scaffolds (47.4% ± 5.0%) than cell-free controls (35.0% ± 2.1%, p = 0.04; Fig. 5B). Percentages of defect fill with hyaline-like cartilaginous tissue and percentages of all repair tissue with features of hyaline-like cartilage were not significantly different between groups (p = 0.26 and 0.92).

Quantitative analysis of repair tissue and defect dimensions. Six months following transplantation, femoral condyles were explanted, processed, sectioned at 8-μm thickness, and stained with safranin O and fast green. Histological sections were assessed for
In the assessment of oxygen tension on repair tissue dimensions, areas of all forms of repair tissue and hyaline-like cartilaginous tissue were not found to be significantly different in the comparison of cell-free controls, normoxia-cultured BMSC-seeded scaffolds, and hypoxia-cultured BMSC-seeded scaffolds (p = 0.21 and 0.56, respectively; Fig. 5A). For cell-free controls, normoxia-cultured BMSC-seeded scaffolds, and hypoxia-cultured BMSC-seeded scaffolds, percentages of defect fill were 35.0% ± 2.1%, 50.4% ± 6.5%, and 44.4% ± 8.0%, respectively (p = 0.42; Fig. 5B). Percentages of defect fill with hyaline-like cartilaginous tissue and percentages of all repair tissue with features of hyaline-like cartilage were not significantly different between groups (p = 0.74 and 0.87; Fig. 5B, C). Assessment of these parameters within each animal is shown in Supplementary Fig. S1.
Macroscopic assessment of repair tissue
Defects were assessed macroscopically at the time of explantation. Variation was seen in the quality of repair tissue within each treatment group as some defects were resurfaced with tissue while other defects were relatively unfilled (Fig. 6A–C). Analysis using a modified Goebel scoring system (Supplementary Materials and Methods S6 Table 1) demonstrated that scores between cell-free controls (11.4 ± 1.3) and BMSC-seeded scaffolds (13.0 ± 0.9) were not significantly different (p = 0.37; Fig. 6D). Subscores based on color of repair tissue, surface quality of repair tissue, defect fill, and degeneration of adjacent cartilage were similar between groups (p ≥ 0.44), while the subscore for presence of blood vessels approached a significantly higher level (less blood vessels) in BMSC-seeded scaffolds (p = 0.09). Modified Goebel scores were not significantly different between cell-free controls (11.4 ± 1.3) and defects implanted with normoxia-cultured BMSC-seeded scaffolds (12.8 ± 1.3) and hypoxia-cultured BMSC-seeded scaffolds (13.2 ± 1.4; p = 0.73; Fig. 6D).

Macroscopic images and scoring of cartilaginous repair tissue. Six months following transplantation, distal femora were explanted and photographed.
Discussion
The main finding of this preclinical study is that BMSCs that were isolated, expanded, seeded within HYAFF scaffolds, chondrogenically primed ex vivo for 4 days, and implanted into full-thickness articular cartilage defects were associated with the development of hyaline-like cartilaginous repair tissue that was superior to controls in an ovine model. Hypoxia improved the chondrogenic gene profile and reduced the expression of hypertrophic cartilage-related collagen X in BMSCs during preimplantation ex vivo priming. However, incubator oxygen tension during ex vivo culture of BMSCs did not consistently modulate the quality of repair tissue in ovine joints present at 6 months following implantation.
The proposed preimplantation protocol of BMSC isolation, expansion, and chondrogenic priming is a hybrid of BMSC implantation protocols that have been assessed in preclinical and clinical studies.8,10–12,25,29–31 The goal of this protocol was to reduce the duration of the ex vivo chondrogenic culture period required by the predifferentiation protocol of BMSC transplantation while providing chondrogenic priming to undifferentiated BMSCs before implantation. Four days of culture in chondrogenic medium was shown to significantly enhance collagen II and aggrecan gene expressions of ovine BMSCs within HYAFF scaffolds relative to unprimed controls. This finding is consistent with the limited literature available before this study that assessed chondrogenic priming of BMSCs in serum-free medium containing TGF-β3 and dexamethasone for periods of less than or equal to 7 days within in vitro culture systems.46–50
Gene expression after 4 days of chondrogenic priming was compared to that after 14 days of chondrogenic differentiation. Collagen II and aggrecan mRNA expressions after 4 days of priming were enhanced to a level similar to that after 14 days of differentiation. Other groups reported similar findings.46–50 Murdoch et al. followed collagen II gene expression each day over the initial 6 days of chondrogenic culture and noted an augmentation after 4 days relative to earlier time points of 1–3 days. 48 A second rise in expression was noted again between 5 and 6 days. The findings of the present study suggest that a short period of priming is sufficient to direct BMSCs toward a chondrogenic lineage. Future study is required to determine the ideal time of exposure to chondrogenic medium for priming within short culture periods under 2 weeks.
BMSCs that were isolated, expanded, seeded within HYAFF scaffolds, chondrogenically primed, and implanted within full-thickness cartilage defects led to superior cartilaginous repair tissue formation and quality based on modified O'Driscoll scores, repair tissue area, and defect fill measurements than cell-free controls. This finding provides preliminary evidence that this specific protocol is promising and may serve as an option for treatment of full-thickness cartilage defects. Furthermore, this finding indicates that the HYAFF scaffold is suitable for use in this BMSC transplantation protocol as it has been with other protocols involving undifferentiated BMSCs, BMACs, and differentiated chondrocytes.11,12,15,51,52 Future work should involve more rigorous study of the proposed protocol to establish that it is capable of consistently fostering chondrogenesis and to compare BMSCs that are chondrogenically primed to BMAC, undifferentiated BMSCs, and BMSCs that have been predifferentiated for multiple weeks before transplantation.
The effect of incubator oxygen tension during preimplantation ex vivo culture on quality of cartilaginous repair tissue developed in vivo within large-animal joint cartilage defects was assessed for the first time in this study. Hypoxic exposure during isolation, expansion, and differentiation was shown previously to improve chondrogenic gene expression and ECM formation within in vitro pellet, micromass, hydrogel, and scaffold culture systems.33–40 Our group demonstrated that hypoxic culture during distinct periods of isolation/expansion and differentiation augmented in vitro BMSC chondrogenesis within a HYAFF scaffold. 37 In the present study, culture in hypoxic conditions significantly increased the collagen II-to-X ratio, increased the collagen II-to-I ratio to a level that approached significance, and significantly reduced collagen X gene expression. A consistent effect of oxygen tension during preimplantation ex vivo culture on histological scoring, repair tissue dimensions, and macroscopic scoring was not found. In the paired assessment of these parameters within each of three donors that were implanted with normoxia-cultured BMSC-seeded scaffolds in one stifle joint and hypoxia-cultured BMSC-seeded scaffolds in the contralateral stifle joint, variability between donors was apparent. Modified O'Driscoll histological scores were higher following transplantation with hypoxia-cultured BMSCs than normoxia-cultured BMSCs in two animals, while the opposite was found in a third animal.
Lack of a consistent difference between oxygen tension groups could relate to several factors. First, the duration of exposure of differentiating cells to specific oxygen tensions within ex vivo and in vivo environments could modulate chondrogenesis. In vitro experiments involve sustained control of oxygen tension during BMSC culture periods of isolation, expansion, and differentiation.35–37 In vivo experiments that utilize subcutaneous implantation of BMSC-seeded biomaterials into mice allow for control of oxygen tension during ex vivo culture periods that are similar in duration to in vivo periods during which oxygen tension is not controlled.36,40 However, in vivo experiments like those performed in this study involve ex vivo culture periods that are much shorter than in vivo periods. The effects induced during ex vivo culture periods may become less relevant than the in vivo environment to modulation of cellular activity and quality of remodeling repair tissue as the duration of in vivo period is lengthened. Since joints have been shown to be naturally hypoxic, 53 it is possible that sustained exposure of implanted cells to hypoxic joint environments could modulate chondrogenesis and reduce differences between groups. Similar to this proposed effect but within a shorter time frame, it was previously shown in vitro that hypoxic exposure during differentiation offsets the differences established between hypoxia and normoxia during isolation and expansion.35,37 In addition to the effects of oxygen tension, other modulatory factors that are present within joints—such as mechanical loading—could become more prominent in affecting tissue formation as time between implantation and joint assessment is lengthened for in vivo studies. 54
A second factor that could be involved in absence of a consistent difference between oxygen tension groups in this study is the involvement of a donor-specific impact of oxygen tension on tissue quality. Although hypoxic culture has been shown to augment chondrogenesis in BMSCs derived from the majority of donors in the studies published to date, our group has reported evidence that human and ovine BMSCs have varying in vitro chondrogenic responses to oxygen tension with BMSCs from a minority of donors demonstrating more pronounced chondrogenesis under normoxia.35,37 Boyette et al. reported superior in vitro chondrogenesis of BMSCs following a protocol of normoxic preconditioning and hypoxic differentiation, but noted variability between human donors in the positive impact of hypoxia on chondrogenesis. 55 More recently, Anderson et al. demonstrated a wide range of GAG production by BMSCs across human donors in an in vitro pellet culture model. 38 They divided donors into high-GAG and low-GAG categories based on GAG production under normoxia. It was noted that hypoxia enhanced GAG production and collagen II gene expression more significantly in BMSCs from low-GAG donors.
To fully appreciate the general effect and variability of effect of oxygen tension during preimplantation ex vivo culture on in vivo cartilaginous tissue formation, further study is required. Given the interdonor variability noted in the optimizing effect within the in vitro literature,35,38 an in vivo study involving a larger number of animals may be required to determine the interdonor variability in vivo, individual animal variations ex vivo versus in vivo, and whether hypoxic culture improves cartilaginous tissue formation across most animals. We postulate that the positive impact of hypoxic incubation during preimplantation culture on in vivo cartilaginous tissue formation would be apparent in a larger study.
The impact of incubator oxygen tension on BMSC proliferation during ex vivo expansion was also assessed. Larger cell counts were found at the end of the expansion period following expansion under hypoxia, although the difference between groups was not statistically significant. Population doublings per day and cumulative population doublings were similar under normoxia and hypoxia. These results are consistent with previous studies reporting nonsignificant differences during early passages and enhanced expansion under hypoxia with prolonged expansion periods beyond P2.40,56
The specific cartilage injury and transplantation model used in this study allowed for preclinical investigation of BMSC transplantation principles, although a number of limitations of this model existed. Sheep were used given that this species is commonly utilized in the assessment of cell-based treatment modalities for cartilage restoration.25,27,57,58 Simulated defects were created during the same surgical procedure and just before construct implantation. This protocol has been described previously and allows for minimization of surgical insult to each stifle joint given that separate surgical procedures are not required for defect creation and treatment.19,24,57 However, it fails to represent the time period between cartilage injury and surgical intervention that chronic models account for. 25 The defect diameter of 7 mm used is consistent with previous literature and considered to be a critical-sized defect.25,59 Each defect was created to involve the full thickness of cartilage without compromising the subchondral bone plate to prevent the involvement of a subchondral source of cells. The downside of this technique is that the cartilage of the ovine femoral condyle is quite thin, which leads to relatively shallow defects that potentially increase the likelihood of implant delamination in comparison to osteochondral defects. Two defects were situated in each stifle joint with one on the medial condyle and another on the lateral condyle, which allowed for defects to be situated within different microenvironments within each joint. Exposure and visualization of the lateral condyle was more difficult than the medial condyle given the origin of the extensor digitorum longus. 60
Other possible limitations of the sheep model used in this study relate to implant fixation and postoperative activity. Construct implantation was supplemented with Tisseel fibrin sealant. This technique is consistent with multiple clinical studies that have used fibrin sealant or platelet-rich fibrin gel to promote implant fixation.11,12,14,15,17,61 Indeed, in this study, it is possible based on the histological appearance of certain defects implanted with cell-seeded scaffolds that those implants may have delaminated causing a lowering of scoring for cell-based implants. The use of suture to augment fixation has also been reported. 15 It is unclear whether suture augmentation is required for the model used in this study. Damage to adjacent native cartilage by sutures has been described previously in a sheep study that utilized full-thickness cartilage defects. 62 Postoperatively, animals were allowed to perform weight bearing as tolerated. In contrast to clinical studies in which guarded weight bearing is used in the postoperative setting, sheep returned to full weight bearing immediately following surgery. 11
In conclusion, ovine distal femoral cartilage defects that were implanted with ex vivo chondrogenically primed BMSC-seeded scaffolds developed hyaline-like cartilaginous repair tissues with improved quality relative to cell-free controls. These findings provide preliminary evidence that this protocol may serve as an option for treatment of full-thickness cartilage defects. Incubator oxygen tension during ex vivo culture improved the preimplantation gene expression profile, although the quality of repair tissue present 6 months after implantation was not modulated consistently between animals. It is currently unclear whether preimplantation ex vivo culture of BMSCs in hypoxic conditions improves cartilaginous tissue formation following transplantation. Therefore, further study is required to assess the use of oxygen tension modulation in the optimization of BMSC transplantation.
Footnotes
Acknowledgments
Financial support was provided by the University Hospital Foundation at the University of Alberta Hospital, Canadian Institutes of Health Research (CIHR MOP 287058) and the Edmonton Orthopaedic Research Committee. Anika Therapeutics S.r.l. (Abano Terme, Italy) graciously supplied the HYAFF scaffold. Baxter (Mississauga, Canada) graciously provided the Tisseel fibrin sealant. We would like to thank Dr. Thomas Churchill for providing us with histology equipment, Mrs. Aillette Mulet-Sierra for her expertise in histology, and Dr. Craig Wilkinson and Ms. Brenda Tchir for their involvement in animal monitoring and care. Stipend support and research allowance for Dr. Troy Bornes was provided by an Alberta Innovates–Health Solutions Clinician Fellowship and Canadian Institutes for Health Research Frederick Banting and Charles Best Doctoral Research Award.
Authors' Contributions
T.D.B. was involved in study design and performed aspiration surgeries, cell culture and tissue engineering, implantation surgeries, explantation procedures, cell and tissue processing, experimental assays, data acquisition, histological scoring, macroscopic scoring, data analysis, and article writing. A.B.A. was involved in study design, co-supervised the study, guided cell culture and tissue engineering, oversaw experimental assays, and revised the article. N.M.J. was involved in study design, co-supervised the study, performed aspiration and implantation surgeries, conducted histological and macroscopic scoring, and revised the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.