
Abstract
This study aimed to determine the effect of fibronectin (FN) on cartilage regeneration through the activation of chondrogenic progenitor cells (CPCs). Cells were isolated from the knee cartilage of mice and cultured in the presence of various concentrations of FN. Proliferation, migration, and chondrogenic differentiation assays were performed in vitro. In some experiments, CPCs were preincubated with anti-integrin α5β1 antibody for 60 min before FN treatment to block the integrin α5β1 receptor. Soluble FN was mixed with Pluronic F-127 and injected into the joint cavity in an early-stage osteoarthritis model. Cartilage repair was evaluated histologically, biochemically, and biomechanically. In vitro, we observed that the isolated CPCs, which exhibited stem cell-relevant markers, proliferated most at a concentration of 20 μg/mL FN (p < 0.05). In addition, FN enhanced the proliferation, migration, and chondrogenic differentiation capacity of CPCs, and the enhancement was significantly decreased by blockade of the integrin α5β1 receptor (p < 0.05). In vivo, FN also significantly promoted cartilage repair along with increased CPC activation and integrin α5β1 expression (p < 0.05). These findings suggest that FN enhances CPC proliferation, migration, and chondrogenic differentiation through the integrin α5β1-dependent signaling pathway. Based on these results, a novel and promising therapy focused on targeted activation of CPCs by FN could be developed for the treatment of cartilage injuries in a clinical setting.
Introduction
C
The discovery of chondrogenic progenitor cells (CPCs) has cast new light on cartilage repair. Our previous work indicated that CPCs isolated from the superficial zone of cartilage display a phenotype that is distinct from the phenotype of CCs. 5 The current literature reports that CPCs, which cycle slowly and have a quiescent nature, may become activated at the onset of cartilage injury. 6 CPCs are homogeneously distributed along the surface of cartilage, making it difficult for them to migrate to the defect site, and they lack a strong ability to differentiate directly into cartilage. The use of gene transfer or gene silencing techniques to induce CPC activation has been reported, but these techniques are of low specificity and safety and cannot be used in clinical practice.7,8 Therefore, a clinically promising method for CPC activation is urgently needed.
Fibronectin (FN) is a high-molecular-weight (450 kDa) glycoprotein. 8 FN promotes cell–cell and cell–substrate adhesion and cellular locomotion, all of which are necessary for the maintenance of cellular structure and function. 9 A number of studies have shown that FN dose dependently increases the migration of CCs and the rate of cell metabolism, thereby increasing the rate of protein, RNA. and DNA synthesis.10,11 Because of this and because it has a favorable safety profile, FN can be used as an alternative material for cartilage repair. The fact that CPCs have a greater capacity to adherence to FN than CCs suggests that FN might have beneficial effects on CPC activation and chondrogenesis.
FN is known to be the classical ligand of integrin α5β1. Adhesion molecules of the integrin family, which comprise the main extracellular matrix (ECM) receptors, consist of two transmembrane subunits (α and β) that form a functional dimer. 12 Integrins regulate the proliferation, migration, and differentiation of several types of cells. We and other researchers have found that CPCs exhibit integrin α5β1 expression levels 10-fold higher than those found in CCs.5,13 The relationship between the positive effects of FN on chondrocyte function and the integrin α5β1-dependent signaling pathway has been confirmed, 14 but its effect on CPCs remains unexplored. In addition, further studies are needed to determine whether there is a strong association between CPC activation and the activity of integrin α5β1.
We hypothesized that FN could enhance cartilage repair by activating the integrin α5β1-dependent pathway in CPCs. To test this hypothesis, several in vitro cell culture experiments and the topical application of a FN/Pluronic F-127 hydrogel were performed in an early-stage osteoarthritis (OA) in vivo model.
Materials and Methods
Media and reagents
C57BL/6 mice (6 weeks old) were provided by the Animal Center of Nanjing Medical University. All animal protocols were approved by the animal ethics committee of Nanjing Medical University. Trypsin, fetal bovine serum (FBS), phosphate-buffered saline (PBS), and DMEM/F-12 were purchased from Keygen (China). Anti-SOX-9, RINX-2, cyclin D1, PCNA, collagen-I (Col-I), collagen-II (Col-II), collagen-X (Col-X), aggrecan, MMP-13, CD34, CD45, CD105, CD166, β-actin monoclonal antibodies, Hoechst 33342, FN, and anti-mouse and anti-rabbit secondary antibodies were purchased from Abcam. The anti-Notch-1 monoclonal antibody was purchased from Novus. Anti-integrin α5β1 antibody was purchased from Millipore. The Click-iT EdU Assay Kit, cDNA Synthesis Kit, and SYBR Green PCR Core Kit were purchased from Thermo Fisher. Triton and transwell culture chambers were purchased from Sigma. The Cell Counting Kit-8 (CCK-8) was purchased from Dojindo (Japan).
Animals and study design
Seventy-nine mice were utilized. All surgery was performed under sodium pentobarbital anesthesia. The experimental groups included the following: (1) a sham operation group (sham group, n = 15); (2) a sham operation group that received injectable FN/Pluronic F-127 hydrogel treatment (sham with FN/Pluronic group, n = 15); (3) a group that underwent anterior cruciate ligament transection (ACLT)-induced OA without treatment (OA group, n = 15); (4) an ACLT-induced OA group that received injectable FN/Pluronic F-127 hydrogel treatment (OA with FN/Pluronic group, n = 15); and (5) an ACLT-induced OA group that received injectable Pluronic F-127 hydrogel treatment (OA with Pluronic group, n = 15). The soluble FN (20 μg/mL) was incubated at 4°C with 30% Pluronic F-127 (1:1, v/v) dissolved in double-distilled water. Pluronic F-127 was used as a carrier for drug delivery. Normal cartilage in the sham group served as a control. Cartilage was also harvested from normal mice that were not subjected to ACLT for CC and CPC isolation (n = 4). The right knee of each mouse was the experimental joint. All surgical procedures were performed using a surgical loupe, as described previously. 15 In the sham with FN/Pluronic group and the OA with FN/Pluronic group, an injectable FN/Pluronic F-127 hydrogel was injected during the 2nd week after ACLT, which is classified histologically as the early stage of OA. In addition, in the OA with Pluronic group, an injectable Pluronic F-127 hydrogel was injected. The hydrogels were administered by intra-articular injection once per week at a dosage of 10 μL. All animals were humanely sacrificed 6 weeks after operation. The experimental scheme is shown in Figure 6A.
Cell isolation and culture
CCs and CPCs were obtained from four mice (6-week-old males) immediately after euthanasia. Petri dishes (35 mm) were coated with 10 g mL−1 FN in 0.1 M PBS containing 1 mM MgCl2 and 1 mM CaCl2 overnight at 4°C. Trypsin digestion and mechanical dissociation were used to isolate cells from the surface zone of the knee under sterile conditions. At the same time, CCs and CPCs were separated on the basis of the differential adhesion of CPCs to FN as described previously. 5 CPCs and CCs were cultured in DMEM/F12 containing 10% FBS and 1% penicillin/streptomycin.2,16 For subsequent experiments, cells (3rd passage) were incubated in serum-free DMEM/F12 (without any growth factors). To block the integrin α5β1 receptor, anti-integrin α5β1 antibody (8 μg/mL) was added to CPCs 60 min before FN treatment as previously described. 17 The cells were divided into three groups: the control group was cultured in a serum-free medium alone; the activation group was cultured in a serum-free medium containing FN; and the inhibition group was preincubated with anti-integrin α5β1 antibody before FN treatment.
Cell proliferation assay
Cell proliferation was determined using the EdU and CCK-8 assays. For the EdU incorporation assay, CPCs were incubated with EdU (5-ethynyl-2′-deoxyuridine) for various amounts of time in a serum-free medium before fixation and permeabilization. EdU detection was performed according to the manufacturer's instructions. Quantification of EdU-positive CPCs was performed using ImageJ software. For the CCK-8 assay, CPCs were seeded in 96-well plates at a density of 8 × 103 cells/well in the appropriate medium. CCK-8 buffer (10 μL) was added to each well, and the cells were incubated at 37°C. The absorbance was then measured at a wavelength of 450 nm (650 nm reference) using an ELX800 absorbance microplate reader (BioTek), and the results were evaluated using GraphPad Prism software version 5.0c (San Diego, CA).
Cell migration assay
Cell migration was determined using both a transwell chamber assay and a wound healing assay. CPC suspensions (30,000 cells in serum-free medium) were added to the upper chambers of a transwell plate, and the lower wells were filled with a serum-free medium or a medium lacking FN. The plates were incubated for 12 or 24 h before processing. The migrated cells were counted in five visual fields using a microscope. For the wound healing assay, cells were grown to nearly 100% confluence in six-well plates in a serum-free medium. The cell monolayers were wounded by scraping them with a micropipette tip. Photomicrographs were obtained under a phase-contrast microscope (Olympus, Japan) immediately following or 24 h after wounding. These experiments were repeated thrice.
Chondrogenic differentiation assay in vitro
Cells were stimulated for 28 days in a serum-free chondrogenic medium without (control) or with FN. The basic serum-free chondrogenic medium consisted of DMEM-high glucose supplemented with 1% ITS+, 40 μg/mL L-proline, 1% sodium pyruvate, 1% non-essential amino acids, 50 μg/mL ascorbic acid 2-phosphate, 10−7 M dexamethasone, and 100 U/100 μg penicillin/streptomycin. 18 For the monolayer culture assay, chondrogenesis was induced for 28 days and evaluated by RT-PCR and immunofluorescent staining. For the cell pellet assay, 2 × 105 CPCs were pelleted by centrifugation and cultivated in the medium for 28 days, after which their differentiation was evaluated by immunohistochemical staining.
Flow cytometry
One million CPCs (passage 3) were washed in PBS and incubated for 1 h at 4°C with fluorophore-conjugated antibodies against CD34, CD45, CD105, and CD166. The cells were centrifuged at 200 g, the supernatants were removed, and the cells were washed thrice in PBS. The labeled cells were resuspended in 1 mL of PBS and subjected to single-channel fluorescence-activated cell sorting (FACS) analysis. The results were analyzed using FlowJo software (version 7.6.2; Tree Star).
Western blot analysis
Cells were sonicated in a standard lysis buffer containing protease and phosphatase inhibitors. Western blots were performed using standard protocols, and proteins were visualized using Pierce West Dura detection reagent and a Chemi Doc-It imaging system attached to a BioChemi HR camera (Upland, CA). β-actin was used as a loading control.
Quantitative RT-PCR
Total RNA (12 pellets per experimental condition) was isolated using Trizol reagent (Invitrogen) and 2 μg RNA was reverse transcribed using a cDNA synthesis kit according to the manufacturer's recommendations. The relative expression level of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase was used to normalize the samples. RT-PCR was performed in triplicate using 1 μL of each cDNA sample in a 7500 Real-Time PCR System (Applied Biosystems) with a SYBR Green PCR Core Kit. The following primer pairs were used: collagen-I, 5′-TGGTACATCAGCCCGAAC-3′ (forward) and 5′-GTCAGCTGGATAGC GACA-3′ (reverse); collagen-II, 5′-GCGGTGAGCCATGATCCGCC-3′ (forward) and 5′-GCGACTTACGGGCATCCT-3′ (reverse); and aggrecan, 5′-GAAATGACAACCCCAAGCAC-3′ (forward) and 5′-TCTCCGCTGATTTCAGTCCT-3′ (reverse). Relative quantification of the marker genes was performed using the ΔΔCt method. The results are reported as fold change compared to the control.
Immunohistochemistry and Osteoarthritis Research Society International score evaluation
To immunostain cells or paraffin sections of tissues, the samples were fixed in 4% paraformaldehyde in PBS at room temperature. Immunohistochemical staining was performed according to the instructions provided by the manufacturer. Primary antibodies against Notch-1, integrin α5β1, SOX-9, RUNX-2, Col-I, Col-II, aggrecan, and CD105 were diluted 1/100 in PBS/0.1% BSA and incubated with the tissues overnight at 4°C. The antibody complexes were visualized using diaminobenzidine or fluorescein isothiocyanate. Additional sections of the corresponding tissues were stained with hematoxylin and eosin (HE) and Safranin O-Fast Green to reveal tissue morphology. OA severity was evaluated based on six sections of articular cartilage from each mouse using the Osteoarthritis Research Society International (OARSI) scoring system. 19
Biomechanical compression testing
Under view in an operating microscope, uniform 5-mm-diameter discs of cartilage (n = 75) were prepared from the five groups of animals. The specimens were equilibrated in Hamm's F-12 medium for 1 h and then subjected to closed-chamber compression testing as described previously. 20 The specimens were loaded into a micromechanical testing system (Nanjing, China) and subjected to repeated compression testing under identical conditions (0, 25, 50, 75, 100, 125, 150, 175, and 200 newtons) for 1 h. The mean loss of thickness (Δ h) in each group was recorded.
Statistical analysis
All data are presented as the mean ± standard deviation obtained from the results of five repeated experiments (n = 5). One-way analysis of variance, post hoc tests, and paired t-tests were performed to determine statistical significance, with p < 0.05 being considered statistically significant.
Results
Isolation and characterization of CPCs
The morphology of the cells was recorded using a phase-contrast microscope and is shown in Figure 1A. First-passage cells had a spindle-shaped, fibroblastoid morphology. They grew in monolayers and maintained a stable morphology with no sign of granulation. Most of the cells expressed specific markers of CPCs, including Notch-1, integrin α5β1, the cartilage-specific transcription factor SOX-9, and RUNX-2, an important transcription factor that acts during bone development (Fig. 1B). Moreover, the CPCs expressed cell surface antigens that are typically found on stem cells. CPCs at passage 3 were homogeneously positive for CD105 (94.30%) and CD166 (95.11%), but were negative for the hematopoietic surface antigen CD34 (3.91%) and the leukocyte common antigen CD45 (4.61%) (Fig. 1C).

Isolation and characterization of CPCs.
Optimization of FN concentration for stimulation of the proliferation of CPCs
To investigate the optimal FN concentration for enhancing the growth of CPCs, the cells were incubated with varying concentrations of FN (0, 10, 20, 30, or 40 μg/mL) for 12 h, and cell viability was measured using the EdU, CCK-8, and Western blot assays.
In the EdU incorporation assay (Fig. 2A), treatment with 20, 30, or 40 μg/mL FN significantly induced CPC proliferation in comparison with the untreated cells (p < 0.05). Specifically, the number of EdU-positive cells peaked and reached a plateau when the cells were incubated in a medium containing 20 μg/mL or more soluble FN (p < 0.05) (Fig. 2B). Therefore, 20 μg/mL FN was determined as the optimum concentration and was used in subsequent experiments. Similar conclusions were reached based on the results of the CCK-8 assay (Fig. 2C). The absorbance values, which reflected the cell numbers, indicated that an FN concentration of 20 μg/mL showed the best promoting effects over 12 h (p < 0.05). The absorbance value reached its maximum level when the cells were incubated in the medium containing 20 μg/mL soluble FN (p < 0.05). Thereafter, the absorbance value no longer increased as the concentration of FN increased. In the Western blot assay, the expression level of cyclin D1 protein plateaued at 20 μg/mL FN, consistent with the above findings (Fig. 2D).
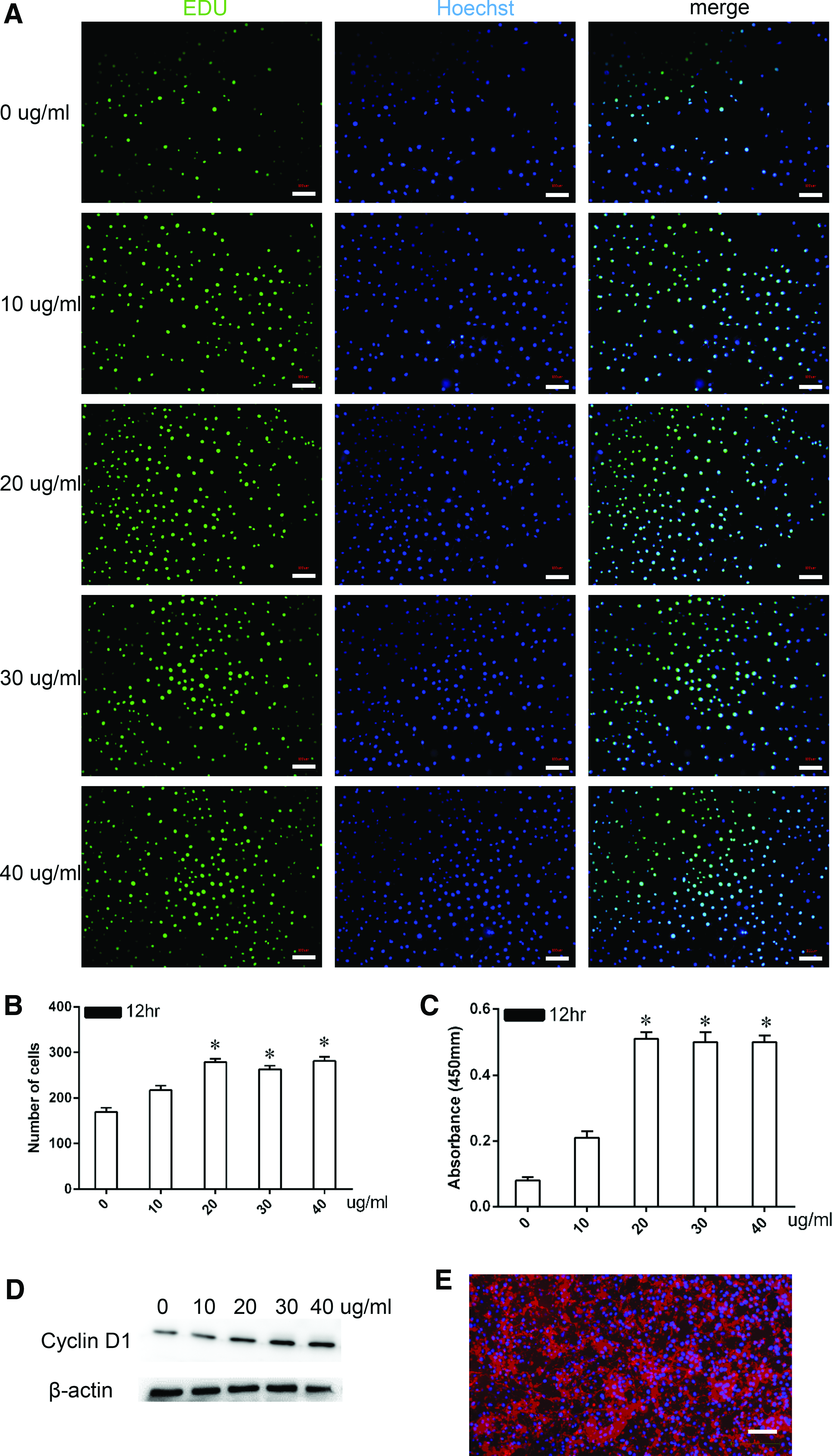
Optimization of FN concentration on the proliferation of CPCs.
The FN in the culture medium remained as freely available or matrix-bound material during in vitro incubation and was detectable by fluorescence imaging (Fig. 2E). The matrix-bound FN was deposited around cells, whereas the freely available fraction displayed a granular appearance.
Effect of FN on the proliferation of CPCs in vitro
A time-dependent increase in CPC proliferation stimulated by FN was observed in the medium containing 20 μg/mL soluble FN in both EdU and CCK-8 assays, as shown by the fact that the cell numbers and the absorbance values increased significantly during 12 h of culture (p < 0.05). No such trend was observed in CC cultures; in those cultures, there was no difference in the cell numbers or absorbance values at the assayed time points (Fig. 3A–C). In the Western blot assay, the expression levels of cyclin D1 protein correlated well with the above findings (Fig. 3D).
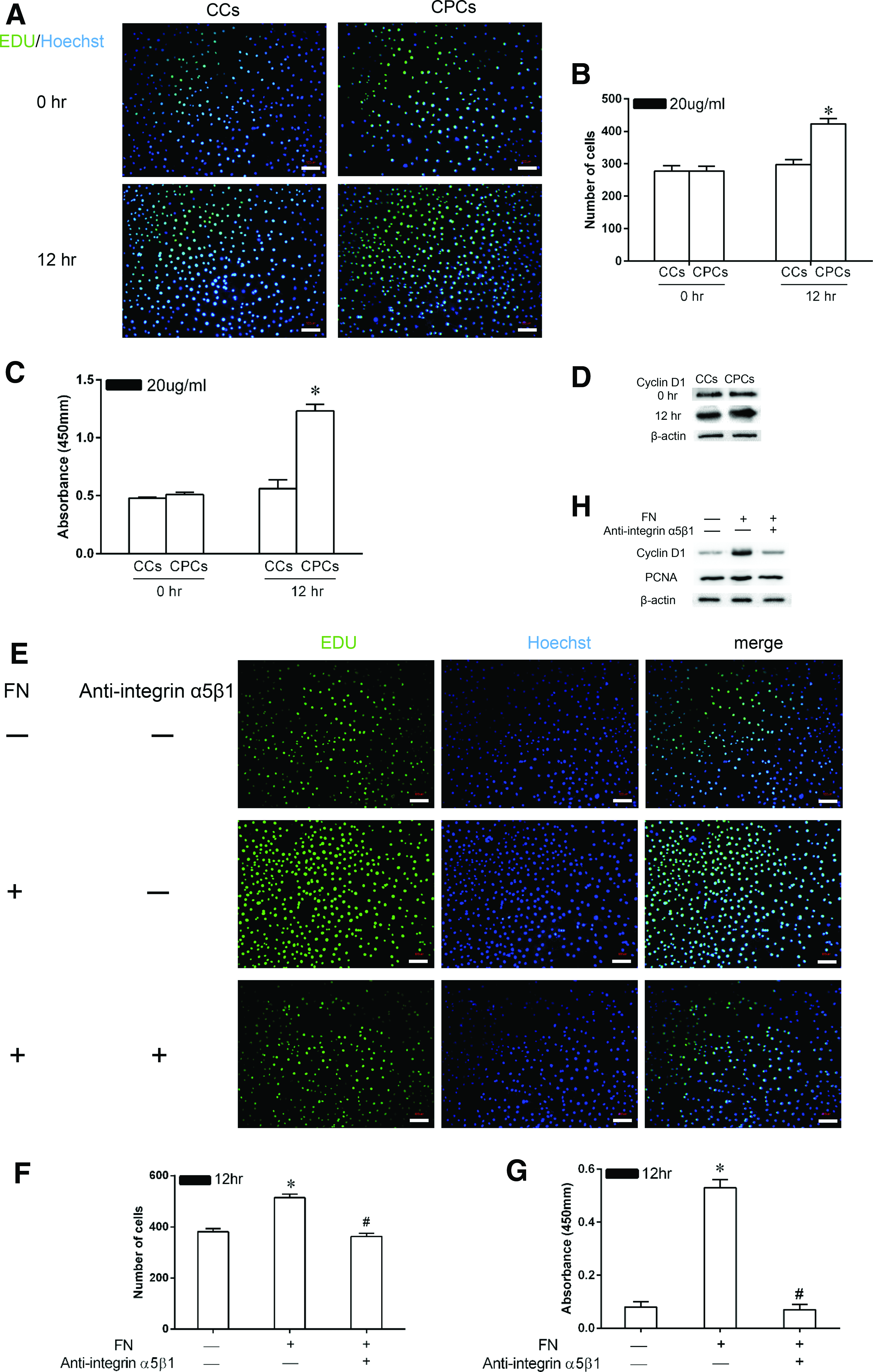
Effect of FN on the proliferation of CPCs in vitro.
To test whether the enhancement of CPC proliferation by FN is mediated through the integrin α5β1 receptor, some of the cells (the “inhibition group”) were preincubated with an antibody against integrin α5β1. FN treatment (the activation group) significantly induced the proliferation of CPCs compared to the control group without FN treatment based on the results of the EdU incorporation and CCK-8 assays (514 ± 14.2 vs. 381 ± 12.5 and 0.53 ± 0.03 vs. 0.08 ± 0.02, respectively; p < 0.05) (Fig. 3E–G). The enhancement of cell proliferation was abolished when the activity of the integrin α5β1 receptor was inhibited, as shown by the fact that the cell numbers and the absorbance values were significantly reduced in the inhibition group compared to those in the activation group (362 ± 13.1 vs. 514 ± 14.2 and 0.07 ± 0.02 vs. 0.53 ± 0.03, respectively; p < 0.05). In the Western blot assay, similar results were obtained; more expression of cyclin D1 and PCNA was observed in the activation group than in the inhibition group (Fig. 3H).
Effect of FN on the migration of CPCs in vitro
To examine the effect of FN on the migration of CPCs and the possible involvement of the integrin α5β1 receptor in this process, transwell migration and wound healing assays were performed (Fig. 4).
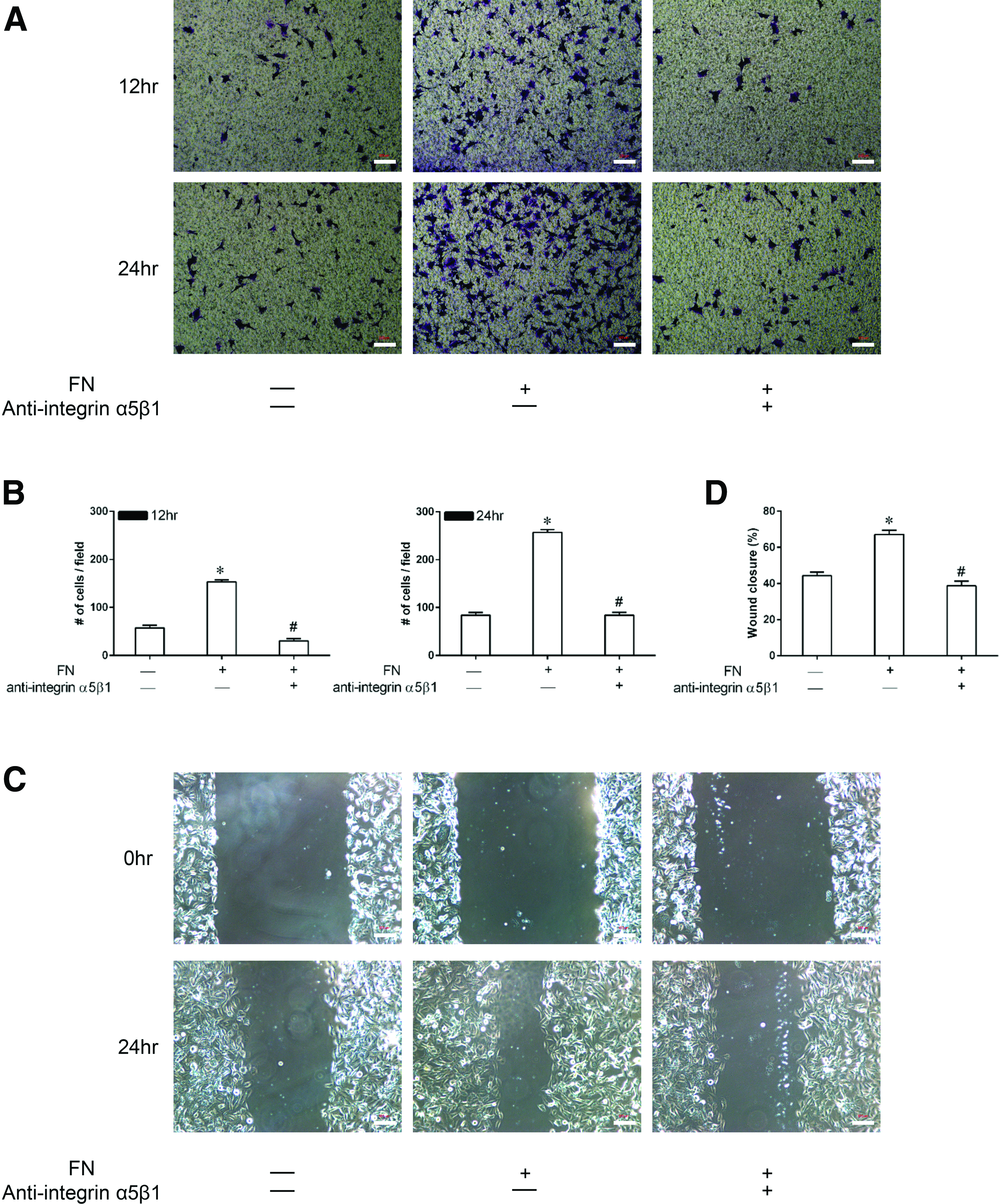
Effect of FN on the migration of CPCs in vitro.
In the transwell assay (Fig. 4A, B), the average number of cells/field was increased by 153 ± 4.2 and 256 ± 6.2, respectively, at 12 and 24 h of culture in the activation group in comparison with the control group (p < 0.05), suggesting that FN increases the migration capacity of CPCs. Blockade of the integrin α5β1 receptor appeared to decrease this effect as the number of migrated cells was significantly lower in the inhibition group than in the activation group (p < 0.05). In the wound healing assay (Fig. 4C, D), consistent results were obtained. After 24 h of incubation, there was a significantly narrower gap area in the activation group compared with that in the control group (67.2 ± 2.3 vs. 44.3 ± 2.1, p < 0.05), but the gap area was enlarged when the integrin α5β1 receptor was inhibited.
Effect of FN on the chondrogenic differentiation of CPCs in vitro
Monolayer and cell pellet culture assays were used to examine the effect of FN on the chondrogenic differentiation of CPCs (Fig. 5). After 28 days of culture, the intensity of immunostaining for collagen-II and aggrecan in both monolayer and cell pellet cultures was significantly increased in the activation group with FN compared to that in the control group without FN, suggesting that FN promotes the chondrogenic differentiation of CPCs (Fig. 5A, B). FN had no effect on the osteogenic differentiation of CPCs, as shown by the fact that the intensity of collagen-I immunostaining remained unchanged even in the activation group to which FN was added. The fact that the staining intensity for collagen-II and aggrecan was significantly decreased in the inhibition group in comparison with the activation group suggests that the integrin α5β1 receptor may be involved in the process of chondrogenic differentiation. Consistent results were obtained by RT-PCR analysis of the above-mentioned cartilage matrices, suggesting that the action of FN on CPC chondrogenesis occurs at the gene level (Fig. 5C).
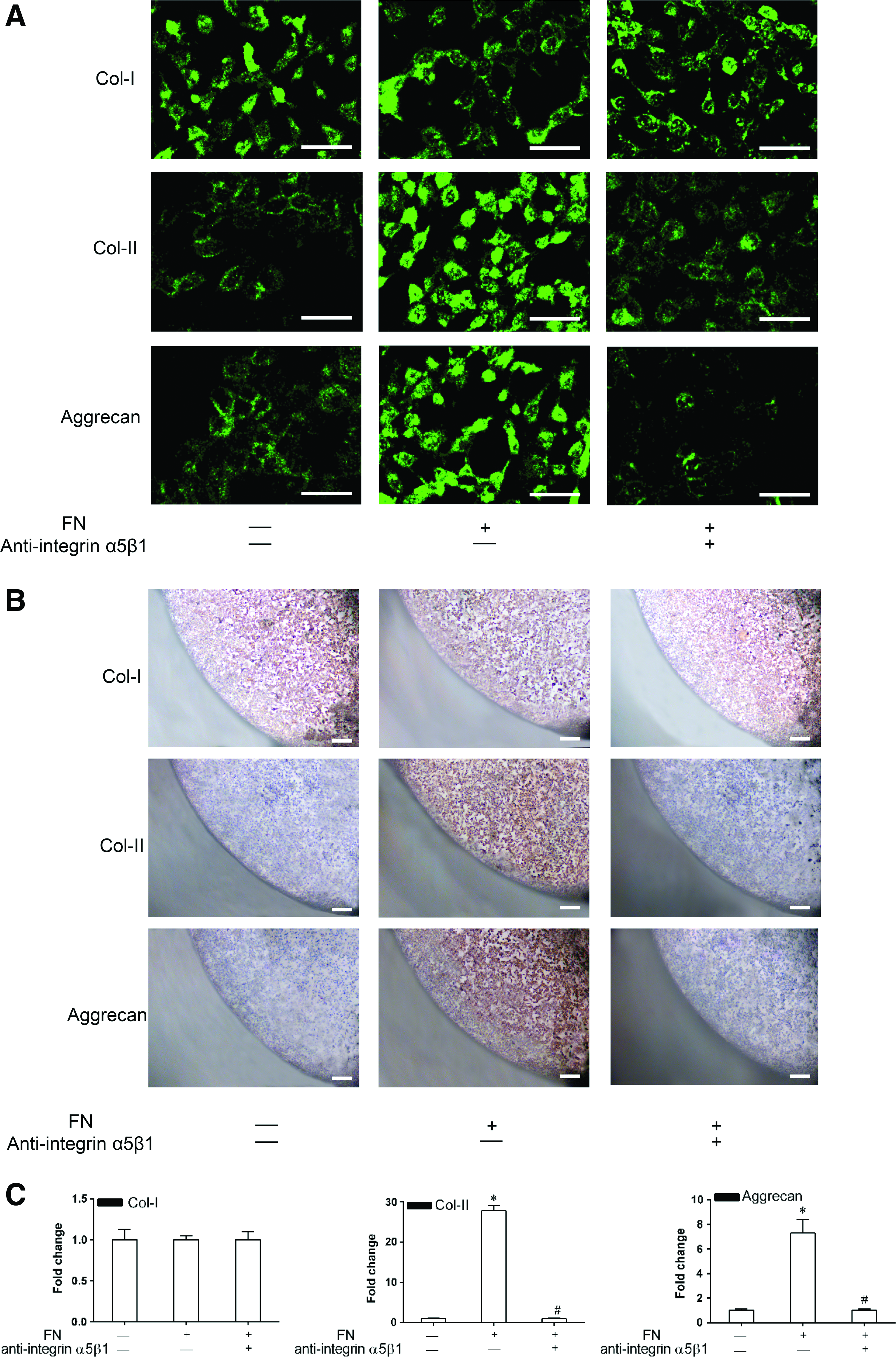
Effect of FN on the chondrogenic differentiation of CPCs in vitro.
Effect of FN on cartilage repair in the early-stage OA model
To investigate the role of FN in cartilage repair in vivo, an early-stage OA model was established. As shown by HE staining, fibrillation and roughening of the articular cartilage prevailed in the OA group relative to the sham and sham with FN/Pluronic treatment groups. After repeated intra-articular injection of FN/Pluronic hydrogel, the articular cartilage surface became smooth and it displayed more cellularity and more staining of the cartilage matrix (aggrecan and collagen), as revealed in the OA with FN/Pluronic group. FN appeared to be the major contributor to the alleviation of OA because the difference in histology between the OA and OA with Pluronic groups was limited. In contrast, FN had no effect on the integrity of normal articular cartilage, as shown by the fact that no difference in histology was found between the sham and sham with FN/Pluronic groups (Fig. 6B).

Effect of FN on cartilage repair in the early-stage OA model.
The OARSI grading system was used for quantitative analysis of the results (Fig. 6C, left). Although FN/Pluronic hydrogel treatment yielded better scores than were found in the OA group (2 ± 0.62 vs. 10 ± 0.58, p < 0.05), the scores were still worse than those obtained for the sham and sham with FN/Pluronic treatment groups (p < 0.05). Pluronic F-127 seemed to have no harmful influence on the beneficial effect of FN; the scores for the OA with Pluronic group were significantly lower than those for the OA group (6 ± 0.73 vs. 10 ± 0.58, p < 0.05), but still higher than those for the OA with FN/Pluronic group (6 ± 0.73 vs. 2 ± 0.62, p < 0.05). A similar beneficial effect of FN was revealed by quantification of collagen-II staining in the different treatment groups (Fig. 6C, right).
In the Western blot assay, the expression levels of collagen-X and MMP-13 were significantly increased in the OA groups compared to the baseline level in the sham groups (Fig. 6D). FN displayed a protective effect on cartilage catabolism; the expression levels of collagen-X and MMP-13 decreased in the FN/Pluronic treatment group compared to the other two OA groups, but Pluronic F-127 had no such protective effect, as shown by the fact that the expression levels of collagen-X and MMP-13 were higher in the OA with Pluronic group than in the OA with FN/Pluronic group.
Biomechanically, the strength of the cartilage in the OA with FN/Pluronic group was superior to that of the cartilage in the other two OA groups, but still weaker than the cartilage in the sham groups (Fig. 6E).
Effect of FN on the activation of CPCs in the early-stage OA model
The influence of FN on the activation of CPCs was also studied. At week 2 in the early-stage OA model, CD105-positive cells were uniformly distributed in the upper two to three cell layers of the cartilage surface (Fig. 7A, top). However, at week 6, more CD105-positive cells appeared in the upper four to five cell layers of the cartilage surface in the OA with FN/Pluronic group than in the OA group (solid arrows vs. open arrows), indicating a higher level of cellular proliferation and migration as a result of FN treatment (Fig. 7A, lower panel). Quantification of CD105-positive cells indicated that the number of CPCs in the mice that received the FN/Pluronic F-127 hydrogel treatment increased from 41.8% at week 2 to 87.5% at week 6 (Fig. 7B). FACS of the CD105- and CD166-positive cells yielded results consistent with the above analysis (Fig. 7C).

Effect of FN on the activation of CPCs in the early-stage OA model.
The potential involvement of the integrin α5β1 receptor in the activation of CPCs was studied by the Western blot assay. In the OA with FN/Pluronic group, the expression level of integrin α5β1 was upregulated at week 6 compared to week 2, whereas in the OA group, the expression level was attenuated at week 6 (Fig. 7D), which suggests that the observed promotion of the activation of CPCs by FN might be mediated by the integrin α5β1 receptor.
Although much of the hydrogel may have been removed during tissue processing, the presence of exogenous FN was confirmed by immunohistological examination of cartilage cross-sections (solid arrows). The exogenous FN appeared as a dense region of the cartilage surface lacking cells, but showing strong immunostaining for FN in the early-stage OA model (Fig. 7E).
Discussion
The development of novel cartilage repair strategies based on stimulation of the homing of endogenous cells is an important current topic in tissue engineering. The use of bone marrow-derived MSCs as well as adipose-derived MSCs and CCs in articular cartilage repair has been extensively studied, but low cell yields and phenotypic alterations during prolonged in vitro culture remain problematic. 21 CPCs may offer a more efficient source of cells for cartilage repair. CPCs display a strong chondrogenic potential and even actively proliferate and migrate toward injury sites. 22 In vitro, we found that FN promotes CPC proliferation, migration, and chondrogenic differentiation. In vivo, topical application of FN promoted cartilage repair in an early-stage OA model. Therefore, CPC-based tissue engineering repair method may offer a potential minimally invasive single-step procedure for cartilage repair.
To isolate and characterize CPCs, we utilized their differential adhesion to FN and found that they are positive for several biomarkers, including Notch-1, integrin α5β1, SOX-9, RUNX-2, CD105, and CD166. These cells express SOX-9 and RUNX-2, demonstrating their potential for chondrogenic and osteogenic differentiation, respectively. Moreover, CPCs are positive for markers of bone marrow stromal cells such as CD105 and CD166 and negative for markers of hematopoietic stem cells and leukocytes such as CD34 and CD45. These characteristics are consistent with the stem cell standards defined by the International Society for Cellular Therapy (ISCT) in 2016. 23
FN has been widely used to treat corneal damage (department of ophthalmology), ulcers (department of dermatology), burn wounds (department of plastic surgery), and other conditions. We investigated the effect of FN on the proliferation, migration, and chondrogenic differentiation of CPCs. Although the effects of growth factors such as TGF-β, PDGF, and IGF on CPC activation may be greater compared with FN, the side effects of these growth factors in vivo are unpredictable due to their multitarget effects.6,9 To the best of our knowledge, the optimal FN concentration for CPC proliferation has never been reported. In this study, we demonstrated that the best promotion of CPC proliferation was obtained using a concentration of soluble FN of 20 μg/mL. Subsequently, we investigated the influence of FN on CPC migration and differentiation at this concentration, and positive effects were also observed. A positive effect of FN has been found in numerous other types of stem cells, including neural crest stem cells, hematopoietic stem cells, and MSCs.24–26
A number of previous studies have demonstrated that integrin α5β1 is strongly expressed on the surface layer of cartilage, and CPCs were mainly located at the same level.6,9,27 It is recognized that integrin α5β1 is the classical FN receptor. 12 In our study, FN promotes the proliferation, migration, and chondrogenic differentiation of CPCs in vitro, but the promotion effect decreased when the integrin α5β1 receptor was blocked. In the early-stage OA model, FN activated CPCs and led to the regeneration of cartilage. At the same time, the expression of integrin α5β1 was also improved. In 1999, Aplin et al. showed that, in addition to regulating cell adhesion to ECM, integrins relay molecular cues about the cellular environment that influences cell shape, proliferation, and migration. 28 Based on our studies and those of others, it can be concluded that FN regulates CPC functions through the integrin α5β1-dependent pathway.
FN is a high-molecular-weight glycoprotein that provides an ideal substrate for CPC attachment. The differential adhesion of CPCs and CCs to FN provides the basis for a classic method for the isolation of CPCs that have been extensively used in the literature.5,29 Using this method, CPCs can be separated from CCs by their different adhesion to FN. CPCs can be harvested either by in vitro cloning based on their stem cell characteristics or by collecting migratory cells from cartilage block cultures in a technique that is based on the higher migration capacity of CPCs compared to that of CCs. However, both of these methods are time-consuming and inefficient for obtaining high numbers of cells in a short time.4,6,9 Therefore, we chose to enrich for CPCs using their differential adhesion to FN. The harvested cells were further characterized by flow cytometry and were shown to possess stem cell characteristics.
The mechanism by which FN improves cartilage regeneration in vivo has not been reported. Our in vivo OA model study revealed that FN treatment leads to abundant ECM formation and high expression of type II collagen, a functional marker for hyaline cartilage. In addition, type X collagen and MMP-13 expression were maintained at surprisingly low levels in FN-treated cultures. Type X collagen is expressed in hypertrophic cartilage before endochondral ossification, and MMP-13 is a major enzyme that degrades the ECM of cartilage. Therefore, FN may modulate the metabolism of CPCs by upgrading anabolism and degrading catabolism. As a result, more newly formed cartilage appeared, and the mechanical strength of the cartilage was improved, as shown by the results of biomechanical tests. Because the mechanical properties of cartilage reflect its functional characteristics, 30 it is not surprising that alleviation of OA was found in our OA with FN/Pluronic group.
During the process of cartilage repair, we observed that CPC proliferation increased significantly in the cartilage surface zone. Similarly, CPC proliferation has been found at sites of cartilage degeneration or injury. Some scientists have even used green fluorescent protein labeling technology to track CPCs and have found that injuries stimulate the emergence and homing of CPCs. 22 Repopulation of the lesion by these cells could promote the repair of chondral damage that might otherwise contribute to progressive cartilage loss.
Carrier material is one of the key factors in cartilage tissue engineering. A suitable carrier material is required to deliver and maintain FN at the injury site. Pluronic F-127 is a synthetic hydrogel that is nontoxic, biocompatible, and bioabsorbable, and has been approved by the FDA for use in humans. 31 It is a liquid at low temperatures and becomes gelatinous at body temperature (37°C). Due to this characteristic, we propose combining FN with Pluronic F-127 to form a drug delivery system (an injectable FN/Pluronic F-127 hydrogel) that could be suitable for clinical intra-articular injection therapy. As revealed by our in vivo study, the novel injectable FN/Pluronic F-127 hydrogel achieved better results, both histologically and biomechanically.
In conclusion, our study presents preliminary evidence that FN activates CPCs and promotes their proliferation, migration, and chondrogenic differentiation both in vitro and in vivo in a manner that is mediated by the integrin α5β1 receptor. Several limitations of our study should be noted. First, because the CPCs were isolated from normal cartilage tissue, further studies are required to elucidate the biological behaviors of CPCs derived from articular cartilage lesions. Second, although Pluronic F-127 is a widely used nontoxic scaffold for the encapsulation of cells, the possible effects of its use in vitro should be further explored. Third, we have not studied the effects of free, nonadherent proteoglycans on CPC behavior. It has been reported that proteoglycans have negative effects on adhesion and cartilage regeneration.32,33 Therefore, the effects of FN might be decreased if they were present.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81601954, No. 81672210 and No. 81673995), Narural Science Foundation for Youths of Jiangsu Province, China (No. BK20151007) and Science and Technology Plan Project Fund of Suzhou (SYSD2014047).
Disclosure Statement
No competing financial interests exist.