
Abstract
Extracellular matrix (ECM)-ornamented biomaterials have attracted attention due to their high potential to improve the biofunctionality of original materials. It is thought that ECM with a bone mimetic microenvironment generated by the specific induction of osteoblasts would be more beneficial for bone regeneration than a regular ECM. In this study, we developed an osteogenic and mineralized ECM construct (Os/M-ECM-SIS) under the guidance of osteoblasts on a small intestinal submucosa (SIS) scaffold cotreated with icariin and calcium. The generated Os/M-ECM-SIS scaffolds exhibited similar morphology and inorganic components as natural bone and higher mechanical strength than ECM-SIS. Cell adhesion, proliferation, and differentiation of osteoblasts and fibroblasts were also enhanced in the cells cultured on the Os/M-ECM-SIS scaffolds. The Os/M-ECM-SIS scaffolds even promoted transdifferentiation of fibroblasts with an upregulation of osteogenic differentiation markers. In a calvarial defect model, new bone formation was greatly enhanced in defects implanted with the Os/M-ECM-SIS scaffolds compared with ECM-SIS scaffolds. Further study showed that the Os/M-ECM-SIS scaffolds promoted bone regeneration in vitro and in vivo via the Bmp/Smad-signaling pathway. Thus, this work proposes a valuable method for generating a mineralized bone mimetic scaffold with SIS as off-the-shelf bone graft substitute that provides an excellent osteogenic microenvironment, making it suitable for application in bone tissue engineering.
Introduction
A
Scaffolds with bone mimetic microenvironment were essential for bone regeneration. 6 Recently, the extracellular matrix (ECM) with excellent biocompatibility has been proven to provide a suitable microenvironment to influence cell fate and to contribute to tissue regeneration, including bone formation. 7
Small intestine submucosa (SIS), derived from the submucosal layer of the porcine intestine, is an acellular, naturally occurring collagenous ECM material with a wide variety of cytokines. 8 These growth factors control osteogenesis, vascularization, bone tissue regeneration, and ECM formation by recruiting osteoprogenitor cells and stimulating their differentiation to specific lineages.1,9 Other components of SIS include glycosaminoglycans, fibronectins, chondroitin sulfates, heparins, heparin sulfates and hyaluronic acids,10,11 all of which play important roles in bone remodeling or are bone matrix components.
The similarities in matrix composition make SIS a potential scaffold for bone tissue engineering. Studies have used SIS to repair bone defects12–14 ; however, SIS typically does not provide beneficial mechanical support, especially during the early stages of implantation, because of its soft tissue origin. Moreover, SIS from the submucosal layer of the intestine may provide different microenvironments for bone tissue, which limits its application to bone regeneration.
Several studies have attempted to generate bone mimic ECM by integrating one or more ECM components in supported biomaterials.15–18 However, there are some drawbacks associated with the use of such natural molecules. First, they are not easy to integrate into compliant biomaterials. Second, synthetic scaffolds with one or more artificial ECM components cannot replicate the complexity of natural ECM, including abundant inorganic and organic components, the ratio of each component, and the spatial distribution of such molecules.
Recently, the creation of a cell-generated ECM coating on scaffold surfaces that mimics the composition of native bone has attracted much attention.3,15,17 These “off-the-shelf” bone substitute materials enhanced biological functionality and possibly with osteoinductivity. Among these investigations, osteoblasts or osteogenically differentiated stem cells have been typically used to generate the ECM coating, which may be potentially capable of regulating the composition and organization of the ECM and thus be similar to native bone environment.3,19,20 However, the ECM generation procedure has not been well studied in these reports, including the key factors that might significantly affect ECM secretion. Moreover, synthetic materials were selected as basic scaffolds in these studies and could not maximize the potential ability of natural ECM.
ECM-generating cell culture was conducted under static or flow conditions. Studies on the effects of the culture medium on the properties of the secreted ECM and the effects of such conditional ECMs to bone regeneration are very limited. The culture medium for ECM production should be optimized based on the nature of the original scaffold material. Two key points were considered for the ECM coating on SIS: mechanical properties and osteogenic properties. Our previous work21,22 showed that icariin is a potent osteogenic compound that significantly enhances osteoblast differentiation in vitro and bone regeneration in vivo by activating the BMP and Runx2 signaling pathways. Icariin was incorporated into SIS scaffolds (Ic-SIS), which were demonstrated to facilitate bone regeneration in vivo. 13 We also previously showed that calcium-enriched medium was able to induce significant biomineralization in vitro within 4 weeks. 23
The purpose of this study was to generate a bone mimetic ECM construct for bone regeneration: osteogenic and mineralized ECM coating-SIS scaffolds (Os/M-ECM-SIS). Osteoblastic MC3T3-E1 cells were stimulated with a special osteogenic cocktail (icariin with calcium-enriched medium) to reorganize the ECM of the SIS-based construct and to engineer osteogenic and highly mineralized artificial bone (Os/M-ECM-SIS) after decellularization. The mechanical and osteogenic properties of the composite were studied and compared with those of raw SIS and SIS with a regular ECM coating (ECM-SIS). An experimental mouse calvarial bone defect model was also used to investigate the bone-regenerating capability of our bone mimetic construct.
Materials and Methods
Generation of decellularized osteogenic and mineralized ECM-modified SIS scaffold
Lyophilized SIS scaffolds were kindly supplied by Cook Biotech, Inc. (West Lafayette, IN). The SIS scaffolds were cut into round constructs measuring 5 mm in diameter for in vitro experiments and 4 mm in diameter for in vivo experiments by a biopsy punch (Miltex, Loznica, Serbia). Icariin and MC3T3-E1 cells were obtained as previously described. 13
Before cell culture, the scaffolds were rehydrated in complete culture medium at 37°C for at least 24 h. Pretreated SIS scaffolds were spread on 96-well plates. Approximately 5 × 104 MC3T3-E1 cells were dropped on the SIS scaffolds. The next day, SIS scaffolds with cells were transferred to new wells and were cultured with regular medium or induction medium (containing 10 mM CaCl2 and 10 μM icariin) for 4 weeks. The concentration of CaCl2 and icariin was selected as previous studies.21–24 The medium was changed every other day. The decellularization process was performed as previous reported. 3 Briefly, the samples underwent three freeze/thaw cycles in a −80°C freezer and a 37°C water bath (15 min for each step). After each thaw step, the scaffolds were rinsed in sterile PBS to remove lysed cells.
The DNA and calcium contents were quantified to verify the extent of decellularization. Two kinds of ECM-ornamented SIS were generated. (1) Osteoblasts on SIS scaffolds were cultured in regular medium followed by decellularization of osteoblasts. The decellularized scaffolds were defined as ECM-SIS scaffolds and (2) osteoblasts on SIS scaffolds were cultured in calcium and icariin medium followed by decellularization of osteoblasts. The decellularized scaffolds were defined as osteogenic and mineralized ECM-SIS scaffolds (Os/M-ECM-SIS).
Elastic modulus (E-modulus) of the scaffolds
The scaffolds measured in an elastic modulus assay were cut to dimensions of 2 × 5 cm2. Cells were seeded on the scaffolds and cultured in regular medium or induction medium for 4 weeks. The decellularized scaffolds were then immersed in PBS or dried in a FreeZone 2.5 lyophilizer (Labconco, Kansas City, MO). The E-modulus was measured using an Instron universal testing machine (Instron Corporation, Canton, MA) at a speed of 1 mm/min. Triplicates were performed for each group.
Morphology and inorganic components of the scaffolds
The surface and atomic compositions of scaffolds were analyzed via scanning electron microscope (SEM) scanning. Samples were fixed in 4% paraformaldehyde PBS solution for 30 min at room temperature, washed with PBS, and dehydrated in 50%, 70%, 90%, 95%, and 100% ethanol for 15 min each. The fixed samples were vacuum dried, sputter-coated with platinum, and examined under an SEM (SU-70, Hitachi, Japan).
X-ray diffraction (XRD) was used to analyze the deposited inorganic components in the Os/M-ECM-SIS scaffolds. Mouse calvarial bone was used as a positive control. XRD patterns were recorded using a Bruker D8 Advance X-ray diffractometer (Ettlingen, Germany) with graphite monochromatized CuKα irradiation (λ = 0.154 nm) using a scan step size of 2θ = 0.02° over a 2θ range of 10–80°.
Quantification of deposited calcium was performed as described in the Methods section of the Supplementary Data (Supplementary Data are available online at www.liebertpub.com/tea). 21
Protein components of the scaffolds
Scaffolds were lysed in intensive RIPA buffer overnight at 4°C. The total amount of ECM protein was quantified using a BCA protein assay kit (P0012S; Beyotime, Shanghai, China) according to the manufacturer's instructions. Triplicates were performed for each group.
Osteocalcin (Ocn) is one of the most important ECM proteins for bone regeneration. The expression and distribution of Ocn protein on decellularized scaffolds were examined via immunofluorescence (IF) staining. The performance was following the protocol of Cell Signaling Technology (CST). Briefly, the samples were fixed in 4% formaldehyde for 15 min at room temperature and rinsed with PBS. Then, the samples were covered with ice-cold 100% methanol for 10 min at −20°C. After washing with PBS, blocking buffer (5% normal serum/0.3% Triton X-100/PBS) was applied to block unspecific staining. The samples were incubated with Ocn antibody (1:200, ab93876; Abcam, Shanghai, China) in 1% BSA/0.3% Triton X-100/PBS at 4°C overnight. The secondary antibody was conjugated with Alexa Fluor® 555 (CST, Shanghai, China). Nuclei were stained with DAPI, and the samples were mounted with antifade solution. The samples were imaged under a confocal microscope (Olympus FluoView™ FV1000, Tokyo, Japan).
Effect of Os/M-ECM-SIS scaffolds on the attachment of osteoblasts and fibroblasts
Approximately 3000 cells were seeded on SIS, ECM-SIS, and Os/M-ECM-SIS scaffolds. After 1 and 4 h, the cells were washed with PBS, fixed in 4% formaldehyde, and stained with β-actin (1:2000, 3700P; CST) by IF staining following the manufacturer's instructions. The samples were imaged under a confocal microscope. Perimeter and area of attached cells were measured using ImageJ. Triplicate staining was performed for each group, and eight random areas from each staining were selected for measurement.
Effect of Os/M-ECM-SIS scaffolds on the proliferation of osteoblasts and fibroblasts
Approximately 1000 cells were seeded on SIS, ECM-SIS, and Os/M-ECM-SIS scaffolds in a 96-well plate. The next day, scaffolds were transferred to new wells and cultured in 100 μL fresh medium. DNA was quantified at indicated times (days 1, 3, and 6) using a CyQUANT® Cell Proliferation Kit (C7026; Invitrogen, Shanghai, China). Briefly, cells were frozen at −70°C and thawed at room temperature for efficient cell lysis. We added 200 μL of CyQUANT GR dye/cell lysis buffer to each sample well. The sample fluorescence was measured under an excitation wavelength of 480 nm and an emission wavelength of 520 nm. A cell number standard curve was generated to calculate the number of attached cells. Four repeats were performed for each group.
Effect of Os/M-ECM-SIS scaffolds on the osteogenic differentiation of osteoblasts and fibroblasts
Bone-specific differentiation markers [alkaline phosphatase (Alp), bone sialoprotein (Bsp), Ocn, and collagen type I (Col1)] were used to evaluate the osteogenic differentiation of cells cultured on the SIS scaffolds. Total mRNA was extracted using TRIzol® reagent (Invitrogen). RNA concentration and purity were measured by a Thermo Scientific Multiskan® GO (Thermo Scientific). The first strand of cDNA was synthesized using a PrimeScript™ RT reagent kit (RR037A; Takara, Dalian, China). Quantitative real-time polymerase chain reaction (real-time qPCR) was performed to assess mRNA expression levels using SYBR green master mix (RR091A; Takara). The relative expression levels were calculated by the comparative Ct method (2−(ΔΔc(t))). 25 Expression of actin was used as an endogenous control in all experiments and relative fold changes were calculated. All reactions were carried out in triplicate, and data expressed as mean ± standard deviation (SD). Specific primer sequences are shown in Supplementary Table S1(Supplementary Data are available online at www.liebertpub.com/tea).
Implantation in a mouse calvarial defect model
Eight-week-old male C57BL/6 mice weighing 20–25 g were purchased from Charles River (Beijing, China) and used in a mouse calvarial defect model to assess new bone formation. The mice were randomly divided into four groups: control (defect only), SIS, ECM-SIS, and Os/M-ECM-SIS groups. All operations were performed under general anesthesia induced by an intraperitoneal injection of 1.2% avertin (T48402-5G; sigma, Shanghai, China)-PBS solution (25 μL/g) on a super clean bench. Critical-sized (4 mm) defects were created on the skull using a biopsy punch and filled with different scaffolds. All animals were sacrificed at 4 or 8 weeks after surgery. All experimental procedures involving animals in this study were conducted in compliance with the Chinese legislation regarding the use and care of laboratory animals and were approved by the Animal Care and Use Committee of Ningbo University.
Sample harvesting and histological staining
The calvarias containing the defects and scaffolds (n = 5 for each group) were harvested and fixed in 10% formalin overnight. The samples were rinsed with PBS, decalcified in 10% (w/v) sodium citrate/22.5% (v/v) formic acid (Morse's solution) for 2 days, neutralized with 5% sodium sulfate for 6 h, and washed with water for 6 h. The samples were then dehydrated, embedded in paraffin, and sectioned (5 μm). Hematoxylin and eosin (H&E) and Masson's trichrome staining (MTS) were used for histological and histomorphometric analysis. H&E staining was performed under standard procedures. MTS was performed according to the instructions of the Trichrome staining kit (Masson, HT15; Sigma). Briefly, the deparaffinized slides were washed with PBS and treated for 15 min at 56°C by Bouin's solution (Sigma), followed by washing in running tap water. The sections were incubated in Weigert's hematoxylin with equal volume of ferric chloride (Nanjing SenBeiJia Biological Technology Co., Nanjing, China) for 5 min and then rinsed in deionized water. The sections were sequentially incubated in working phosphotungstic/phosphomolybdic acid solution for 5 min, in aniline blue solution for 5 min, and in acetic acid (1%) for 2 min, followed by dehydration through alcohol and clear in xylene.
Immunohistochemistry staining
Sections were deparaffinized under standard procedures. After washing with PBS, the sections were heated in sodium citrate buffer for antigen retrieval. An EXPOSE mouse- and rabbit-specific HRP/DAB detection immunohistochemistry (IHC) kit (ab80436; Abcam) was used for IHC staining to detect the protein level of Bmp-2 and the protein localization of pSmad1/5/8. A Bmp-2 antibody (1:500, ab 14933; Abcam) and a pSmad1/5/8 antibody (1:500, 13820P; CST) were applied.
Statistical analyses
All quantitative data are expressed as mean ± standard deviation. Statistical analyses were performed using SPSS® software (Chicago, IL). Statistical significance was determined using a Student t-test (two-group comparison) or a one-way analysis of variance (ANOVA) followed by a post hoc test (multigroup comparison). A p-value less than 0.05 was considered statistically significant.
Results
Cotreatment of icariin and calcium enhanced osteogenic protein secretion and mineralization of osteoblasts on SIS scaffolds
To mimic the bone microenvironment and to increase the mechanical strength of SIS scaffolds, osteoblasts were used to modify the scaffold with the secreted bony matrix under the stimulation of the cocktail medium with icariin and calcium (Ic/Ca). As shown in Figure 1A, osteogenic differentiation markers (Alp and Ocn) were significantly increased in the cells treated with the Ic/Ca medium compared to the regular culture. According to mRNA expression, Ocn protein levels were higher in the cells treated with Ic/Ca than in the regularly cultured cells, as indicated by IF staining (Fig. 1B). A special pattern of secreted Ocn protein was also detected in the Ic/Ca-treated cells. After Ic/Ca treatment to osteoblasts cultured on SIS, the surface of the SIS scaffolds became white (Fig. 1C). The quantification of the calcium deposition also showed that the Ic/Ca medium greatly enhanced mineralization by ∼15-fold compared with that measured for the general medium (Fig. 1C). The results indicated that cotreatment of osteoblasts with icariin and calcium enhanced osteogenic protein secretion and mineralization on SIS scaffolds.
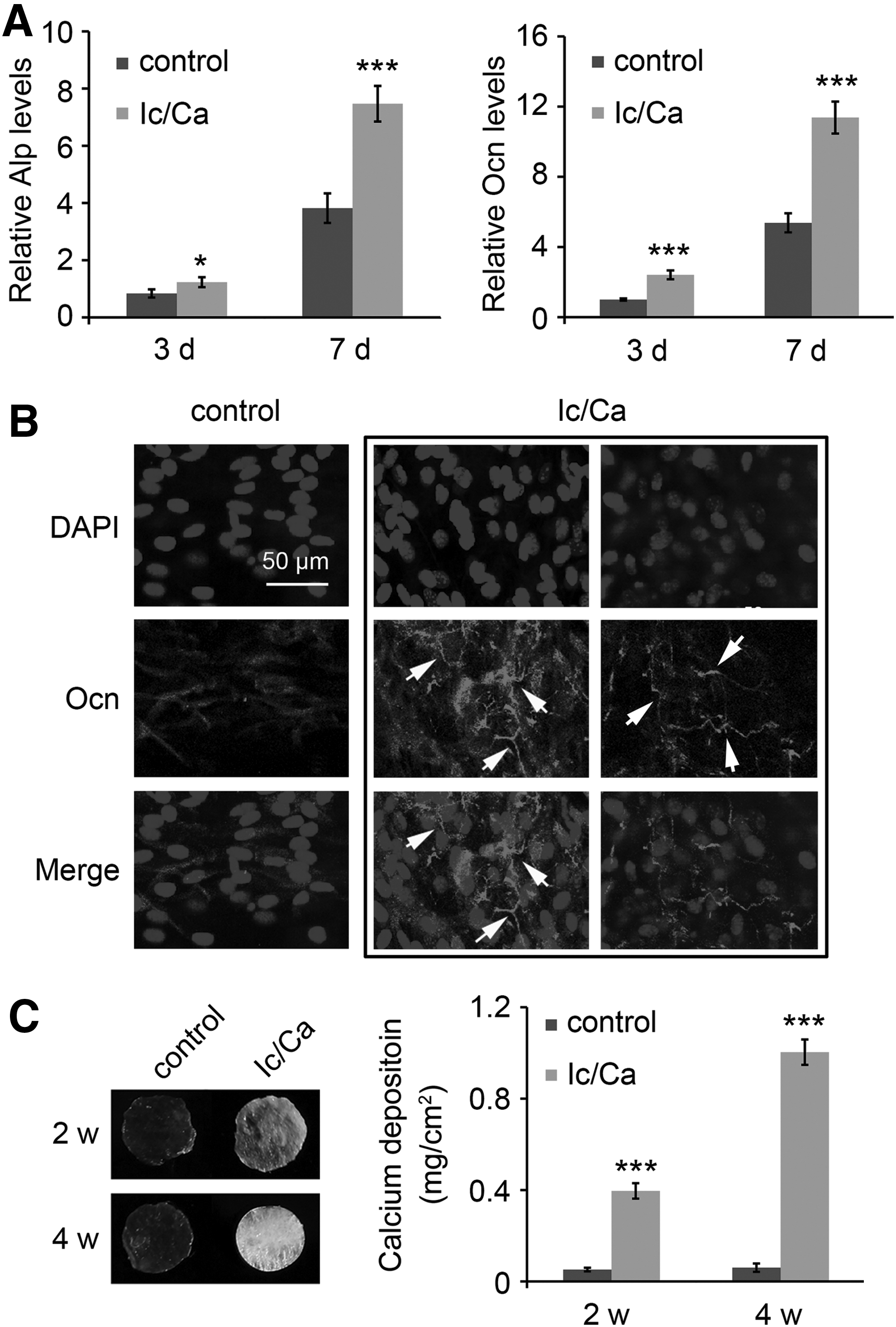
Osteogenic differentiation and mineralization were enhanced in the cells treated with icariin and calcium on SIS scaffolds. MC3T3-E1 cells were seeded on SIS scaffolds and treated with the indicated components of calcium (10 mM) and icariin (10 μM) for the indicated times.
Generation and characterization of osteogenic and mineralized ECM-ornamented SIS scaffolds
To obtain an ECM-ornamented scaffold, osteoblasts were seeded on SIS and cultured with/without the cocktail treatment (Ic/Ca medium). After 4 weeks of culture, cells were removed by washing followed by freeze/thaw cycling. Less than 5% of the original DNA was left in the decellularized scaffolds, whereas no significant reduction in calcium deposition was observed after decellularization (Fig. 2A).
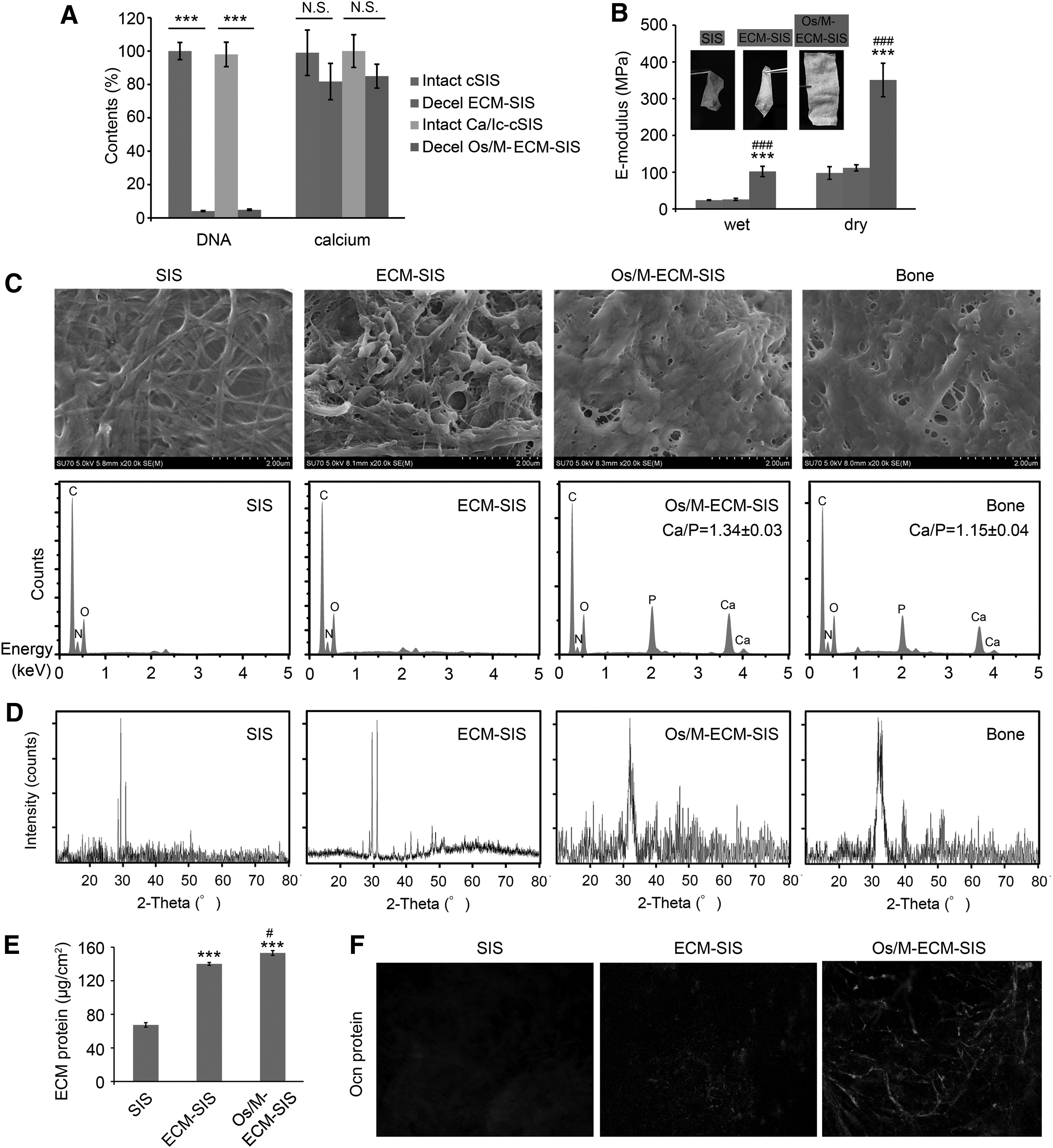
Characterization of the generated Os/M-ECM-SIS scaffolds. MC3T3-E1 cells were cultured on SIS scaffolds with cotreatment of icariin (10 μM) and calcium (10 mM) for 4 weeks and then decellularized by freeze/thaw cycling.
The mechanical properties, morphology, and molecular components of the Os/M-ECM-SIS scaffolds were determined. As shown in Figure 2B, the raw SIS and ECM-SIS scaffolds were soft and translucent, while the Os/M-ECM-SIS scaffolds were obviously rigid and opaque. The elastic modulus of the Os/M-ECM-SIS scaffolds was increased compared with that of the raw SIS scaffolds, from 23 MPa to 134 MPa under wet conditions and from 98 MPa to 442 MPa under dry conditions (Fig. 2B). No significant mechanical enhancement was observed in the ECM-SIS scaffolds compared with the raw SIS scaffolds.
SEM micrographs showed that the surface of the Os/M-ECM-SIS scaffolds was covered by cell-engineered organic and inorganic ECM, similar to natural bone (Fig. 2C). To confirm the atomic composition of the mineral deposits, energy dispersive X-ray spectroscopy (EDS) spectra of the scaffolds were analyzed with SEM. Os/M-ECM-SIS scaffolds revealed high intensities of Ca and P peaks, similar to the EDS spectra of natural bone (Fig. 2C). The Ca/P ratio of Os/M-ECM-SIS is 1.34 ± 0.03, lower than pure hydroxyapatite (1.67) but higher than native calvarial bone. Compared with the XRD pattern of the SIS scaffolds, the pattern of the Os/M-ECM-SIS scaffolds showed new peaks at 2θ values of 32.6° and 39.8° (Fig. 2D), similar to the peaks observed in the XRD pattern of bone. The mineral was later confirmed by the standard powder diffraction file to be hydroxyapatite (HAp), which is the main inorganic component of bone.
Compared with raw SIS scaffolds, ECM-decorated SIS scaffolds had more protein deposition (Fig. 2E). The Os/M-ECM-SIS scaffolds exhibited higher protein content than the ECM-SIS scaffolds. Figure 1B and C shows that the expression of Ocn was increased in the cells cultured on SIS scaffolds after icariin and calcium cotreatment, and secreted Ocn was observed. We further evaluated whether the Ocn protein could be preserved on the Os/M-ECM-SIS scaffolds followed by decellularization. The Ocn protein was markedly detected on the Os/M-ECM-SIS scaffolds, while no Ocn protein was detected on the raw SIS scaffolds and little Ocn protein was found on the ECM-SIS scaffolds (Fig. 2F). Moreover, Ocn protein on the Os/M-ECM-SIS scaffolds exhibited a linear distribution.
Thus, besides HAps, other components, including proteins, were also inside of Os/M-ECM matrix, which contained much more P than Ca, and led to the lower ratio of Ca/P (Fig. 2C). For native calvarial bone, besides the ECM, it also contained cells. Molecules in the cells, especially DNA, contained more P than Ca, which further lowered the ratio of Ca/P.
Effect of Os/M-ECM-SIS scaffolds on the attachment and proliferation of osteoblasts and fibroblasts
Osteoblasts and fibroblasts are major cells that contribute to bone regeneration. The cellular responses of osteoblasts and fibroblasts to the Os/M-ECM interfaced SIS scaffolds were investigated. The initial cell adhesion and spreading behaviors were visualized by confocal microscopy after IF staining of β-actin (Fig. 3). Few cells were found in the raw SIS, and actin staining also corresponded to a limited area, while more cells were found on the ECM-SIS and Os/M-ECM-SIS scaffolds (Fig. 3A). For osteoblasts, the cells exhibited good spreading morphology on the ECM-SIS and Os/M-ECM-SIS scaffolds as early as 1 h, with apparent filopodia extensions. More filopodia were observed on the Os/M-ECM-SIS scaffolds than on the ECM-SIS scaffolds. Fibroblasts spread well on the ECM-SIS and Os/M-ECM-SIS scaffolds for up to 4 h.
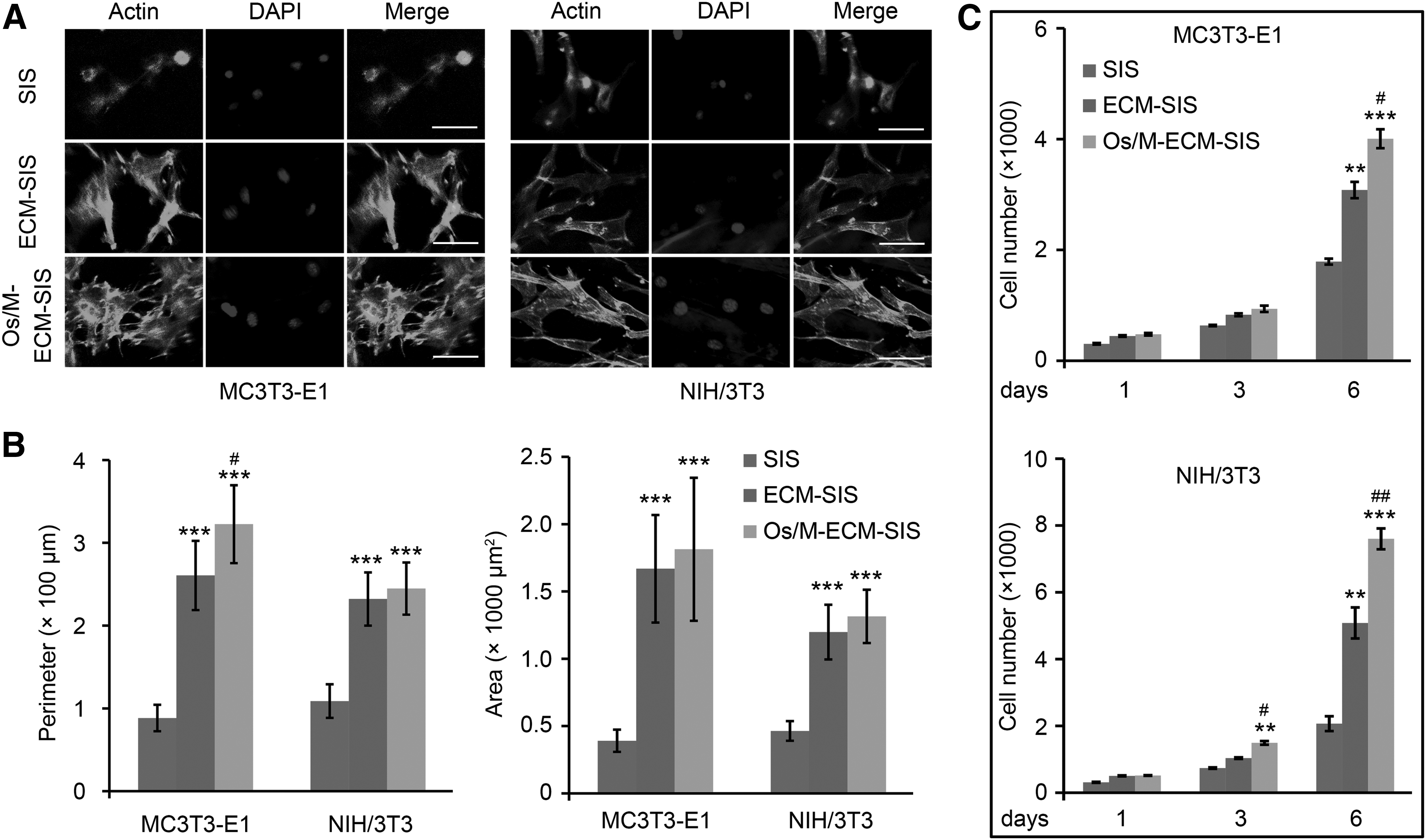
Os/M-ECM-SIS scaffolds promote cell adhesion and proliferation of osteoblasts and fibroblasts. MC3T3-E1 and NIH/3T3 were cultured on SIS, ECM-SIS scaffolds, and Os/M-ECM-SIS scaffolds for 4 h for scaffold cell adhesion analysis and for 6 days for scaffold cell proliferation assay.
The cell spreading area and perimeter were subsequently quantified to assess the cell spreading behavior (Fig. 3B). Compared with raw SIS, the ECM-ornamented SIS (ECM and Os/M-ECM-SIS scaffolds) showed significantly higher values of both indices for fibroblasts and osteoblasts. The perimeter of the osteoblasts on the Os/M-ECM-SIS scaffolds was significantly higher than that on the ECM-SIS scaffolds.
Cell proliferation of osteoblasts and fibroblasts on the Os/M-ECM-SIS scaffolds was later evaluated (Fig. 3C). The cells grew faster on the ECM-SIS scaffolds than on raw SIS. Cell proliferation was further promoted by the Os/M-ECM-SIS scaffolds compared with the ECM-SIS scaffolds. A significant increase in cell proliferation of fibroblasts on the Os/M-ECM-SIS scaffolds was observed as early as 3 days.
Effect of Os/M-ECM-SIS scaffolds on differentiation into osteogenic lineage
Osteoblasts were cultured on SIS, ECM-SIS, and Os/M-ECM-SIS scaffolds for 14 days, and the expression of differentiation markers at different stages (Alp, Bsp, and Ocn) was examined (Fig. 4A). The level of Alp (an early-stage marker) in the cells on the Os/M-ECM-SIS scaffolds was the highest among the three groups on both day 7 and 14. Slight upregulation of Bsp (a middle-stage marker) and Ocn (a late-stage marker) was observed in the cells on the Os/M-ECM-SIS scaffolds on day 7, while their expression was significantly increased in the Os/M-ECM-SIS scaffolds than other scaffolds on day 14.
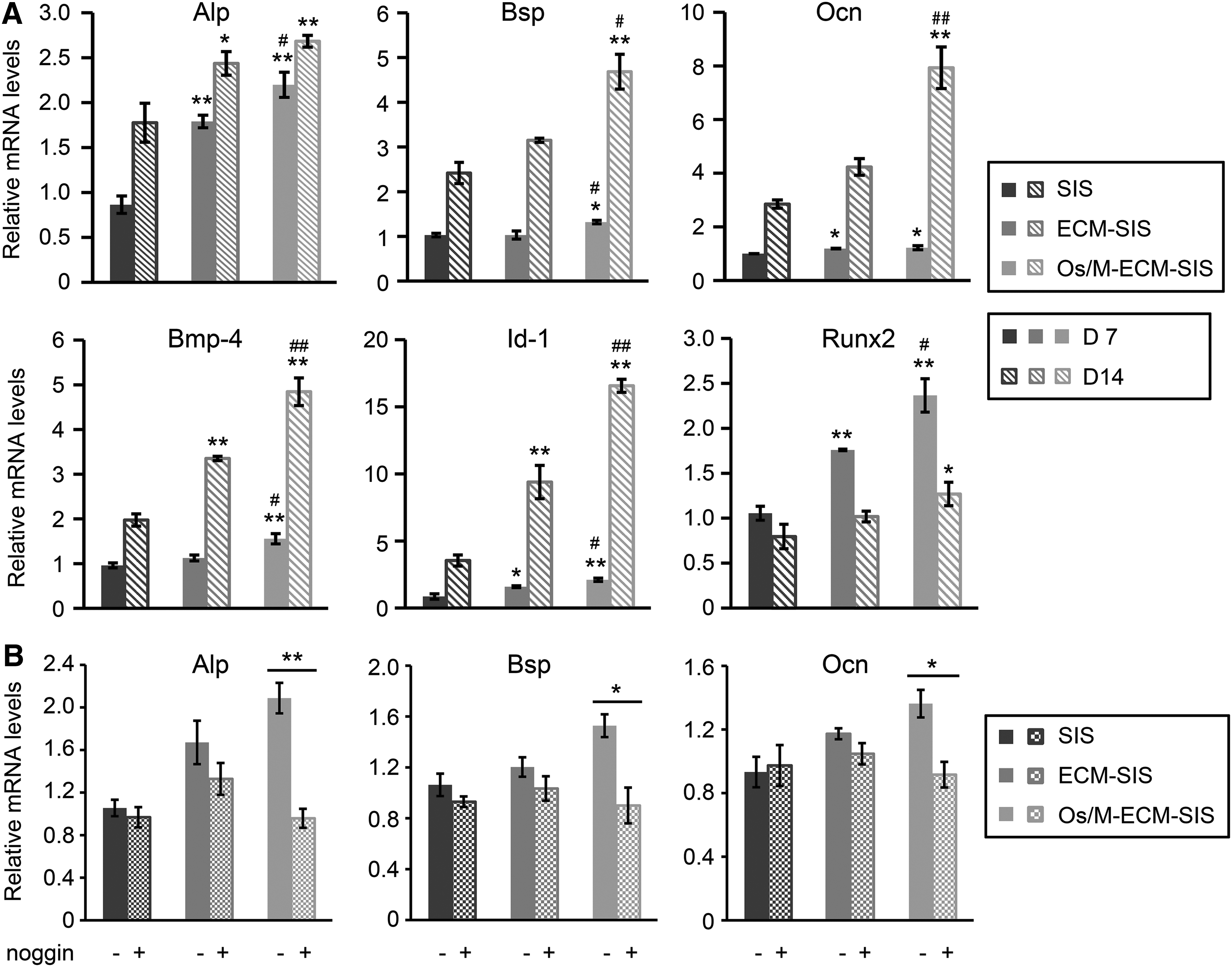
Os/M-ECM-SIS promotes osteogenic differentiation of osteoblasts.
Accordingly, Runx2 was increased by the Os/M-ECM-SIS scaffolds on day 7, and the increase was less significant on day 14. The levels of Runx2 in the three groups on day 14 were lower than those on day 7. Moreover, the expression of Bmp-4 and Id-1 was also upregulated in the cells cultured on the Os/M-ECM-SIS scaffolds compared with those on SIS and ECM-SIS scaffolds on both day 7 and 14. Moreover, noggin, a BMP antagonist, was used to confirm the effect of BMP signaling in the cells on Os/M-ECM-SIS scaffolds. Addition of noggin into the culture medium completely inhibited the upregulation of Alp, Bsp, and Ocn induced by the Os/M-ECM-SIS scaffolds (Fig. 4B). The results indicated that activation of Bmp signaling pathway was a major contributor to osteogenic differentiation of cells on the Os/M-ECM-SIS scaffolds.
Abundant fibroblasts are recruited during the process of bone regeneration in vivo. We seeded fibroblasts on SIS, ECM-SIS, and Os/M-ECM-SIS scaffolds and cultured them for 7 days under Bmp-2 treatment. Interestingly, the Os/M-ECM-SIS scaffolds enhanced transdifferentiation of fibroblasts to an osteogenic lineage induced by Bmp-2, especially to the late stage of osteogenic differentiation (Fig. 5).
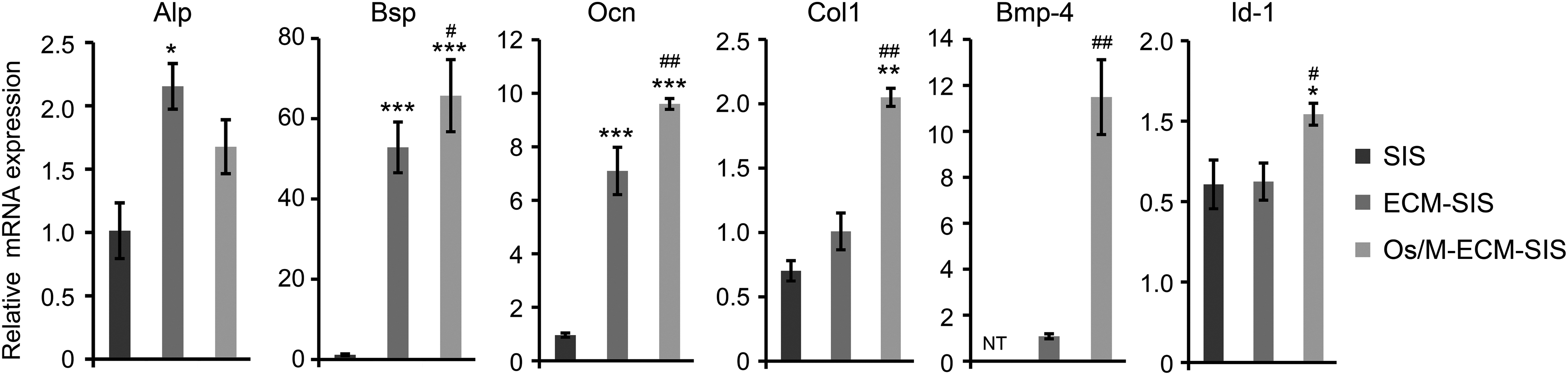
Os/M-ECM-SIS enhanced osteogenic transdifferentiation of fibroblasts induced by Bmp-2. NIH/3T3 cells were seeded on different SIS scaffolds and treated with Bmp-2 (100 ng/mL) for 7 days. The mRNA expression of differentiation marker genes (Alp, Bsp, and Ocn) and key associated genes (Col1, Bmp-4, and Id-1) was assessed by qRT-PCR. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with SIS; #p < 0.05, ##p < 0.01 compared with ECM-SIS.
The Os/M-ECM-SIS scaffolds increased the expression of Alp, Bsp, Ocn, and Col1 in fibroblasts compared with SIS. The levels of Bsp and Ocn were upregulated in the ECM-SIS and Os/M-ECM-SIS scaffolds and were significantly higher in the Os/M-ECM-SIS scaffolds than in the ECM-SIS scaffolds. Furthermore, no Bmp-4 was detected in the fibroblasts on SIS, while it was detectable in the cells on the ECM-SIS scaffolds. Similar to the expression in osteoblasts, Bmp-4 and Id-1 were significantly upregulated in fibroblasts on the Os/M-ECM-SIS scaffolds compared with those on the ECM-SIS scaffolds.
In vivo bone regeneration in a calvarial defect model
A calvarial defect model in C57BL/6 mice was used to investigate the effect of the cell-engineered scaffolds on bone regeneration. At 4 weeks, the defects in the control group were connected by a thin fibrous tissue, while a thicker layer of fibrous tissue was observed in the defects with SIS scaffolds (Fig. 6A). The defects implanted with the ECM-SIS scaffolds exhibited bone-like tissues surrounding the scaffold fibers (Fig. 6B). New bones with the typical structure of mature bone were formed across the defects implanted with the Os/M-ECM-SIS scaffolds at 4 weeks (Fig. 6B). Morphological characterization of the newly formed bone revealed sparse osteocytes embedded in lacunas with osteoblasts lining the outer edge of the bone tissue.
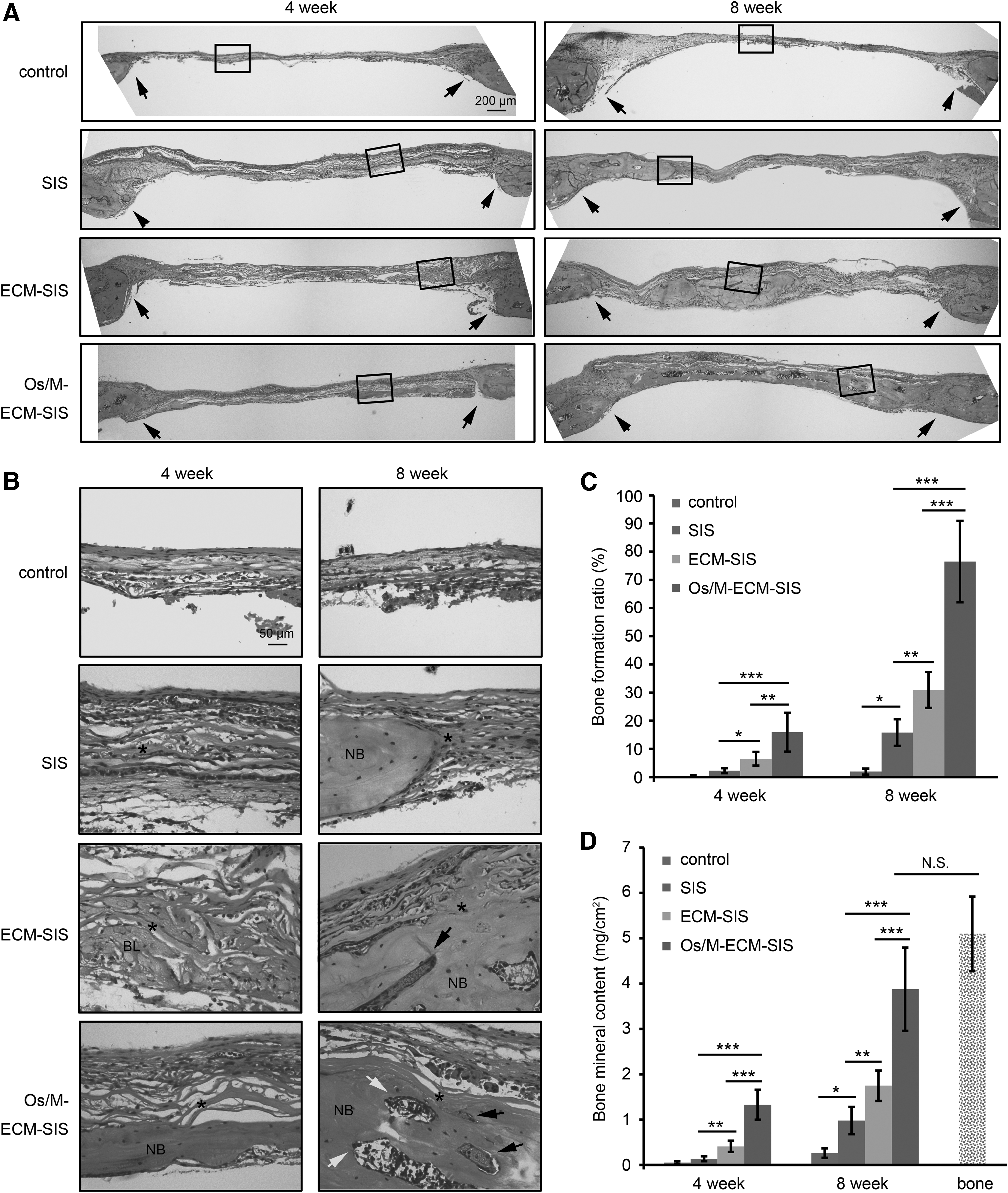
H&E staining in the calvarial defects of the control and in the defects implanted with the scaffolds (SIS, ECM-SIS, and Os/M-ECM-SIS) at 4 and 8 weeks.
By 8 weeks, marrow cavities and blood vessels were observed, and the defects implanted with the Os/M-ECM-SIS scaffolds were almost fully covered with newly formed bone (Fig. 6A, B). High-magnification images demonstrated that the collagenous fibers of the SIS became thinner at 8 weeks, and some fibers were immersed into the newly formed bone (Fig. 6B and Supplementary Fig. S1A). It is possible that the collagen fibers of the SIS scaffolds were reorganized and reused during new bone regeneration. Moreover, the Os/M-ECM-SIS scaffolds with greater mechanical strength were easier to implant and spread, whereas the SIS and ECM-SIS scaffolds bent easily and tended to curve or fold at defective regions (Supplementary Fig. S1B).
New bone formation was quantified by a histological morphology analysis using H&E staining (Fig. 6C), as previously reported.13,14,26,27 The bone formation ratio was defined as area of new formed bone divided by area of defect tissue in HE images, and measured by ImageJ. As expected, a limited amount of new bone was formed in the defect-only groups at 4 and 8 weeks. By 8 weeks, the raw SIS scaffold group exhibited a significantly greater amount of newly formed bone than the control group. Although the bone formation ratio was significantly higher in the ECM-SIS group than in the raw SIS group, the Os/M-ECM-SIS scaffolds exhibited the highest bone formation ratio among all groups at both 4 and 8 weeks.
The calcium deposition was then measured for the harvested calvaria. The calcium content of the defects for all types of implanted SIS scaffolds was significantly higher than that of the control defects (Fig. 6D). The calcium content was the highest in the Os/M-ECM-SIS group at both 4 and 8 weeks.
The sections were also stained with Masson's trichrome (MTS) to further analyze the histological morphology of newly formed bone. As shown in Figure 7A, the control defect was connected by thin fibrous tissue with little blue staining. In all SIS groups, the defect region was almost completely covered with a thick blue collagenous matrix at 4 weeks. Collagen should be secreted by osteogenic cells and fibroblasts that are recruited by the SIS scaffolds. Newly formed mature bone was clearly identified by red staining of the defects implanted with Os/M-ECM-SIS scaffolds at 4 weeks and all types of SIS scaffolds at 8 weeks; this new bone was nearly identical to the original bone at the defect edge.
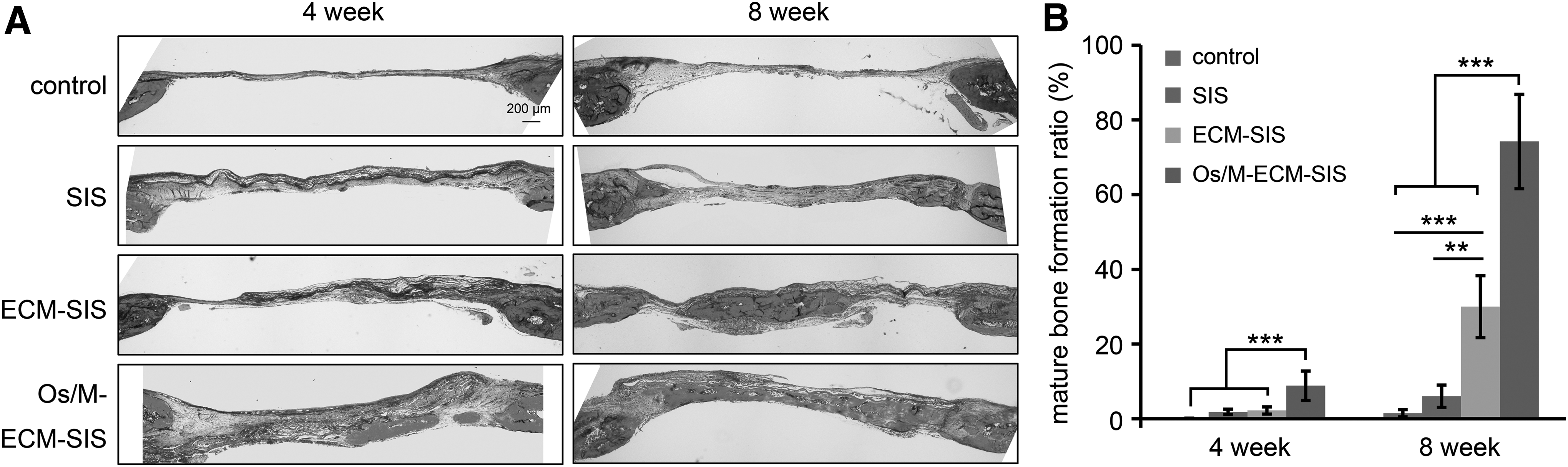
MTS in the defects of the control animals or animals implanted with scaffolds (SIS, ECM-SIS, and Os/M-ECM-SIS) at 4 and 8 weeks.
The new mature bone was quantified based on MTS. Consistent with Figure 6C, more mature bone was formed in the ECM-SIS group and Os/M-ECM-SIS group at 8 weeks, and the Os/M-ECM-SIS group showed the highest ratio of mature bone formation (Fig. 7B).
To further investigate in vivo bone regeneration at the molecular level, the expression of Ocn was assessed by IF staining and was highest in the Os/M-ECM-SIS group at both 4 and 8 weeks (Fig. 8). At week 8, the defect in the Os/M-ECM-SIS group was closely filled with newly formed bone, and the lacuna contained as much Ocn protein as the original bone at the edge (Fig. 8 and Supplementary Fig. S2). Interestingly, most of the Ocn protein was localized on the brain side of the collagenous fibers in all SIS scaffold groups; this pattern was consistent with the localization of newly formed bone as indicated by histological analysis (Fig. 6).

Immunological staining of Ocn, Bmp-2, and pSmad1/5/8 in the defects at 4 and 8 weeks. Ocn was detected by IF staining. Bmp-2 and pSmad1/5/8 were detected by IHC staining.
Our in vitro results revealed that the Os/M-ECM-SIS scaffolds stimulated osteogenic differentiation of osteoblasts and fibroblasts via the Bmp signaling pathway (Fig. 4). In accordance with the in vitro results, Bmp-2 was markedly induced in the defects implanted with the scaffolds, especially in the Os/M-ECM-SIS-implanted defects (Fig. 8). pSmad1/5/8 was detected in nuclei as early as 4 weeks in the defects implanted with the Os/M-ECM-SIS scaffolds. The results confirmed that the Os/M-ECM-SIS scaffolds enhanced bone regeneration in vivo via activation of the Bmp signaling pathway.
Discussion
The design and manufacture of scaffolds with an ideal molecular composition, strong mechanical properties, and superior biological activities are key challenges that limit their successful implementation in bone tissue engineering. 1 Natural biomaterials that are able to mimic the structural, mechanical, and biological properties of natural tissues have attracted significant attention. In the present study, we generated a totally natural construct with the desired bone mimic matrix on SIS scaffolds. The generated construct (Os/M-ECM-SIS) was demonstrated to exhibit similar inorganic and organic components, similar morphology with natural bone, and higher mechanical strength than regular ECM coating scaffolds. Cellular responses, including cell adhesion, spreading, proliferation, and osteogenic differentiation, were greatly improved for use of Os/M-ECM-SIS scaffolds. In addition, the Os/M-ECM-SIS scaffolds induced greater mineralized new bone formation in a calvarial mouse defect model, even with the invasion of vessels and bone marrow.
Disadvantages of synthetic materials such as metals, polymers, and ceramics limit their applications in the clinic. Compared with synthetic materials, naturally derived biomaterials exhibit excellent biocompatibility and biodegradability and low immunogenicity, which have been broadly applied to bone tissue engineering. 28 SIS, a natural collagenous ECM biomaterial, was reported with various bioactive factors. 8 Commercial SIS has been applied in reconstructive therapies, especially for soft tissue repair.29–31 Some studies attempted to transplant SIS sponges into bone defects in vivo, and new bone formation was increased.12,32,33 In those studies, SIS was introduced as the basic scaffold for bone tissue engineering. In our calvarial defect model, SIS scaffold fibers were clearly immersed into newly formed bone, which indicated that collagen fibers of SIS were reused during bone regeneration and further demonstrated the excellent biocompatibility for bone tissue engineering in vivo.
After decellularization, growth factors, such as IGF-1, TGF-α, and bFGF, were still detectable in the SIS scaffold, which indicated that SIS provided effective binding sites for protein combination.8,34 Other studies reported SIS as a useful carrier for sustained protein release.31,35,36 Bovine serum albumin (BSA) was entrapped into SIS gel, and a sustained release of BSA-FITC was observed for up to 30 days in vivo after subcutaneous injections administered to rats. 35 Choi et al. generated a hepatocyte growth factor-containing SIS gel to realize gradient, sustained release. 31 These studies further suggested the effective binding of proteins to SIS scaffolds. Moreover, SIS was not only a useful scaffold for protein binding but also useful for hydrophilic or hydrophobic molecule binding. We previously incorporated icariin into SIS scaffolds, and sustained release of icariin was observed for more than 30 days. 13 These binding sites gave SIS scaffolds the potential ability to preserve the ECM secreted by cells.
In our study, we cultured osteoblasts on SIS scaffolds with special treatments. After decellularization, abundant inorganic and organic components were reserved. Deposited calcium was measured without a significant reduction after decellularization. The protein content was detected in ECM-ornamented SIS scaffolds, and Ocn protein was observed with a special distribution in the Os/M-ECM-SIS scaffolds. Although DNA was detectable in the decellularized scaffolds, the residual DNA was less than 5% and was fragmented. The remaining DNA fragments should not affect transplantation in vivo, even in clinic. For example, Badylak and Gilbert 37 reported that the clinical efficacy of commercial ECM devices had been largely positive despite the presence of DNA remnants. Therefore, the remaining DNA after decellularization in ECM devices leading to any adverse host response is unlikely to be a concern.
Although SIS has been clearly demonstrated as an ideal biomaterial for tissue engineering, original SIS from the small intestine may provide a different microenvironment than that of bone tissue, which limits its application to bone regeneration. Bone tissue is composed of inorganic (mainly hydroxyapatite) and organic proteins (such as osteogenic ECM proteins). Although SIS from the small intestine lacks inorganic calcium and thus mechanical strength, the porous interwoven structure of SIS possesses a large surface area that is beneficial for retaining deposited calcium and other ECM components.
In this study, osteoblasts were used to allow for rapid natural biomineralization and osteogenic ECM secretion on SIS with a special calcium and icariin cocktail treatment. Osteoblasts arise from mesenchymal stem cells and play important roles in bone regeneration. They synthesize crosslinked collagen, special bone matrix proteins, and calcium. In several recent studies,38–40 researchers mixed calcium phosphate-based materials into polymer scaffolds to fabricate bone mimetic constructs that, to some extent, induced bone regeneration in vivo. These artificial complex scaffolds were able to mimic the composition of bone but not the special distribution of calcium in the original tissue. Other studies also cultured cells on synthetic biomaterials. However, the cells were always cultured in regular culture medium, which should only secrete various amounts of partial ECM components, especially key proteins for bone regeneration. We supposed that special cell treatments could alter the composition, distribution, and bioactivity of the secreted ECM, possibly enhancing the osteogenic capability of the construct.
In conditions of committed induction, cells can be induced to function as builders to construct a new mineralized and ECM-ornamented house to better mimic the microenvironment of the host tissue. The new SIS-based house should therefore be designed to integrate these ECM components and to be spatially and functionally organized. This possibility was verified in the study. The elastic modulus of the generated Os/M-ECM-SIS scaffold was significantly increased under both wet and dry conditions. SEM and XRD analyses revealed uniformly mineralized hydroxyapatite and ECM proteins, which were very similar to original bone tissue. The key ECM protein Ocn was distributed in the Os/M-ECM-SIS scaffolds in a special pattern. Most importantly, the Os/M-ECM-SIS scaffolds performed substantially better than the ECM-SIS scaffolds in both the in vitro and in vivo analyses.
Osteoblasts play an essential role during bone formation and remodeling by producing and calcifying bone matrixes.41,42 However, a limited number of osteoblasts were found during bone regeneration, while abundant fibroblasts were recruited and contributed to bone repair in the defect area. Runx2 and Osx, in combination with Oct4 and L-Myc, were transfected into fibroblasts. About 80% fibroblasts were converted into Ocn-producing cells and facilitated bone repair after transplantation into artificial bone defect lesions in immunodeficient mice. 41 Microenvironment has been demonstrated to affect the cell fate of a mesenchymal stem cell, even to convert a particular cell type to another kind of cell. 43 Birk et al. seeded preadipocytes on 3D biomatrices of marine origin, and the cells were modulated and differentiated into bone-forming cells. 43
Bmps, as effective cytokines for bone formation, were commonly used to convert other kinds of cells into osteoblastic cells.44,45 Interestingly, we found that Os/M-ECM-SIS scaffolds greatly enhanced the transformation of fibroblasts to osteoblasts in the presence of Bmp-2, along with the upregulation of osteogenic differentiation markers (Alp, Bsp, Ocn, and Col1). The results indicated that osteoblasts cultured on SIS scaffolds in cocktail medium secreted bone mimetic ECM, and the decellularized Os/M-ECM-SIS scaffolds reserved these ECM components, which contributed to the conversion of fibroblasts into osteoblasts and promoted bone regeneration in vivo.
Bmps have been considered as critical cytokines for regulating osteoblast differentiation and successive bone formation. Bmp-2, the most effective cytokine for bone formation, exhibits strong osteosynthetic activity in vitro and in vivo, even to induce ectopic bone formation.46,47 Bmps modulate osteoblast differentiation by activating Smad1/5/8 phosphorylation and regulating osteoblast-related transcriptional factors, including Runx2.48,49
The mechanism by which the Os/M-ECM-SIS scaffolds facilitate bone regeneration was investigated in vitro and in vivo in the present study. Bmps and Runx2 were significantly increased in the cells cultured on the Os/M-ECM-SIS scaffolds and in the defects implanted with the Os/M-ECM-SIS scaffolds. It is possible that the Os/M-ECM-SIS scaffolds activate Bmp receptors directly via the bioactive factors on the scaffold or indirectly via upregulation of Bmps of cells. Under the activation of Bmp receptors, Smad1/5/8 was phosphorylated, combined with Smad4, and transferred into the nuclei. Increased Runx2 cooperated with pSmad1/5/8/Smad4 and stimulated osteogenic differentiation-associated gene expression, including Alp, Bsp, Ocn, and Col1.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 31300800), the Applied Research Project on Nonprofit Technology of Zhejiang Province (No. 2017C33135), and the K.C. Wong Magna Fund in Ningbo University. The authors thank Cook Biotech Inc. for the supply of lyophilized SIS scaffolds.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.