
Abstract
Reconstruction of vertebral laminae without epidural scar formation has been challenging. The success of bone formation depends on the biological and mechanical conditions of the surrounding tissues. In this study, we aimed to investigate the roles of biological and mechanical factors in the osteogenesis of artificial laminae. Mesenchymal stem cells derived from rabbit umbilical cord Wharton's jelly were induced for osteogenic differentiation for 3 weeks before seeding on the hydroxyapatite-collagen I scaffolds to construct the tissue-engineered laminae (TEL). TEL were then implanted into the fifth rabbit lumbar vertebrae in both orthotopic lamina (n = 30) and ectopic lamina (n = 30) groups. De novo laminae were examined through histological and radiographic analysis in the 2nd, 4th, 8th, 12th, and 16th weeks postimplantation. Our results showed that de novo laminae formed effectively in both groups, and the osteogenic gene expression levels and cancellous microstructure parameters of de novo laminae in the orthotopic lamina group were significantly higher than those in the ectopic lamina group. In conclusion, the biological stimulation of bone defect initiated the early onset osteogenesis, and the mechanical stimulation of cerebrospinal fluid pulsation stress promoted the osteogenesis of de novo laminae.
Introduction
As the aging of populations and high prevalence of spinal degenerative diseases, total laminectomy and semilaminectomy have been commonly used to treat spinal disorders.1,2 Total laminectomy and semilaminectomy can induce various degrees of scar tissue and adhesion formation in the epidural space and lead to failed back surgery syndrome. 3 Various attempts have been investigated to avoid the occurrence of epidural scar tissue, such as autogenous bone grafting, allograft transplantation, ceramic artificial bone implantation, titanium mesh with autogenous bone grafting, and so on.4–6 However, challenges exist during their clinical applications, including limited supply of autograft bone, donor site pain, immune reaction, potential virus transmission after allogeneic bone grafting, creeping substitution, and unstable titanium mesh fixation.4–6 Tissue engineering techniques, as an ideal alternative, have been applied to reconstruct epidural fat or vertebral laminae by inhibiting the occurrence of epidural scar tissue.7–9
Tissue engineering involves four essential elements such as seed cells, growth factors, scaffolds, and biomechanical environment. Our previous studies have successfully reconstructed artificial vertebral laminae using bone marrow mesenchymal stem cells (BMSCs) combined with collagen scaffolds or β-tricalcium phosphate bioceramics.7,9 But there was still little evidence about the regulating roles of biological and mechanical factors in the bone formation of de novo laminae.
Bone formation starts with an initial anabolic phase, where several biological factors, including IL-1β, IL-6, and TGF-β, are released from the bone defect and initiate the early onset osteogenesis.10–12 In the late stage, mechanical stimulations such as fluid shear stress and pulsation pressure begin to take the preponderant role by regulating the activities of MSCs.13–16 After laminectomy, tissue-engineered laminae (TEL) were influenced by both the biological stimulation of cytokines released from the bone defect and the mechanical stimulation of continuous cerebrospinal fluid pulsation (CSFP) stress.
In this study, we hypothesized that these biological and mechanical stimulations could promote the osteogenesis of de novo laminae in rabbit. We constructed TEL by rabbit umbilical cord Wharton's jelly derived MSCs (WJ-MSCs) and hydroxyapatite-collagen I scaffold and then implanted TEL into the fifth rabbit lumbar vertebrae in both orthotopic and ectopic lamina groups. The orthotopic laminae animal model was made by the removal of native laminae at the size of 10 × 8 × 2 mm, leaving two flesh cancellous bone ends measuring 10 × 2 mm as illustrated in the Figure 1A and C. The ectopic lamina animal model was made by the removal of outer surface cortex of laminae at the size of 10 × 4 mm while preserving the dura surface cortex of laminae, leaving one bone defect at the size of 10 × 4 mm as illustrated in the Figure 1B and D. Osteogenesis in both groups was examined through histological and radiographic analysis for up to 16 weeks postimplantation.

The construction of animal models.
Materials and Methods
Isolation of WJ-MSCs
A pregnant female New Zealand rabbit (27-day pregnancy) was anesthetized using 10% ketamine (40–90 mg/kg; Alfasan, Woerden, Holland) and 2% xylazine (5 mg/kg; Alfasan). The umbilical cords were dissected and flushed thrice using phosphate-buffered saline (PBS; Hyclone, Beijing), supplemented with 100 U/mL of penicillin-streptomycin (Gibco, New York). The epithelium was removed, and the blood vessels were peeled off to expose the WJ. WJ was cut into infinitesimal pieces by ophthalmic scissors and then placed into a 25 cm2 Petri dish. Collagenase II at 1 mg/mL (Sigma-Aldrich) dissolved with PBS was mixed with the WJ. After 4–6 h, the digestion was terminated and centrifuged; the cells in the sediment were resuspended in complete medium containing low-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (Biological Industries, HaEmek, Israel) and 100 U/mL penicillin-streptomycin and then plated in Petri dishes at a concentration of 3 × 104/cm2 in the incubator at 37°C under a 5% CO2 atmosphere. Culture medium was refreshed every 3 days, and the cells were passaged when reaching 80–90% confluence. Cells at passage 4 were used for the following experiments. The protocol was approved by the Committee on the Ethics of Animal Experiments of Fudan University (No. 20160862A003).
Multilineage differentiation
For osteogenic differentiation, WJ-MSCs at passage 4 were cultured in osteogenic induction medium (10 nM dexamethasone [DEX], 10 mM β-glycerophosphate sodium, 50 mg/mL ascorbic acid, and 10 nM 1,25-dihydroxyvitamin D3; Sigma-Aldrich). The cultures were changed every 3 days. On day 21, the mineralization was assessed by Alizarin Red S staining (Sigma-Aldrich).
For adipogenic differentiation, WJ-MSCs at passage 4 were cultured as a monolayer in the presence of adipogenic supplements (1 M DEX, 1 g/mL insulin, and 0.5 mM 3-isobutyl-1-methylxanthine; Sigma-Aldrich). On day 21, cultures were stained with Oil Red O stain (Sigma-Aldrich).
For chondrogenic differentiation, a cell suspension of 1.6 × 107 cells/mL was prepared and seeded at 5-μL droplets of cell solution to generate the micromass cultures in the center of 24-well plate Petri dish. After cultivating micromass cultures for 2 h under high humidity conditions, chondrogenesis media (Gibco) was added to culture vessels and incubated in 37°C incubator with 5% CO2. The cultures were refreshed every 3 days. On day 21, chondrogenic pellets were stained with Alcian Blue staining (Sigma-Aldrich).
TEL construction
Under sterile condition, the hydroxyapatite-collagen I scaffold (Beijing Allgens Medical Science & Technology Co., Ltd., Beijing, China) (Fig. 3A, B) was cut to the size of 10 × 8 × 2 mm. After 3 weeks of culture in osteogenic medium as described in the Multilineage Differentiation section, osteo-differentiated WJ-MSCs were trypsinized and resuspended in media at a concentration of 1 × 106/mL. Then, 100 μL cell suspensions were pipetted on one side of each scaffold, and after 30 min, 100 μL cell suspensions were pipetted on the other side. All constructs were placed in osteogenic medium for another week before implantation.
To observe the cell activity on the scaffold, the osteo-differentiated WJ-MSCs were labeled with fluorescent DiI (Sigma-Aldrich) and then seeded on the hydroxyapatite-collagen I scaffold. After 2 days, the distribution of cells within scaffold was observed under inverted fluorescent contrast phase microscope (Leica).
Construction of orthotopic lamina and ectopic lamina rabbit models
Sixty 2-month-old male rabbits weighing 2.25 ± 0.25 kg were randomly divided into the orthotopic lamina group (n = 30) and the ectopic lamina group (n = 30). The animals were anesthetized with pentobarbital sodium (1 mL/kg intraperitoneally).
For rabbits in the orthotopic lamina group, we located the spinous process of fifth lumbar vertebrate by the anatomical landmark and made a 3-cm longitudinal skin incision. The superficial fascia and paraspinal muscle were retracted to clearly expose the spinous process, which were then removed to expose the native laminae. At last, a bone defect measuring 10 × 8 × 2 mm was created in the laminae by rongeur to expose the dura. The removal of native laminae left two flesh cancellous bone ends measuring 10 × 2 mm at the left and right direction as illustrated in the Figure 1A and C. The TEL were placed in the bone defect and fixed in place at the four corners using a 4-0 polypropylene suture (PROLENE; Ethicon, Tokyo, Japan).
For rabbits in the ectopic lamina group, we removed the paraspinal muscle with a detacher to clearly expose the spinous process and laminae and then removed the spinous processes by rongeur. By removing the spinous processes, a cancellous bone end measuring 10 × 2 mm was created. The cancellous end was then enlarged to the size of 10 × 4 mm by abrasive drilling, similar to that of orthotopic lamina group (10 × 2 × 2 mm), while preserving the dura surface cortex of laminae as illustrated in the Figure 1B and D. The TEL were fixed onto the native laminae.
Then the fenestrated fascia was closed with a 4-0 polyglactin suture (VICRYL; Ethicon). After the operation, sodium penicillin (400,000 U) was administered. All rabbits were housed in separate cages with free access to food and water without immobilization.
Micro-computed tomography examination
The tissue specimens were harvested in the 2nd, 4th, 8th, 12th, and 16th weeks after implantation and were immediately fixed in freshly prepared 4% (w/v) paraformaldehyde. The osteogenesis of specimens was examined using micro-computed tomography (CT) at the Shanghai Public Health Clinical Center. For the orthotopic lamina group, the de novo bone forming in the bone defect was analyzed, and for the ectopic lamina group, the de novo bone forming on the native laminae was analyzed, strictly precluding the native laminae in the two-dimensional images manually using the GEHC MicroView2.0+ABA software. Quantitative analysis was performed using the three-dimensional (3D) model reconstructed by GEHC MicroView2.0+ABA software. Indexes, including bone volume fraction (BVF), trabeculae number (Tb.N), and trabeculae thickness (Tb.Th), were evaluated.
Histology examination
Tissue specimens were decalcified with 10% ethylenediaminetetraacetic acid for 4 weeks. Tissue sections with 6-μm thickness were cut on a microtome and mounted onto glass slides. The sections were processed for routine histological analysis by hematoxylin and eosin (H&E) staining and immunostaining with bone morphogenetic protein-2 (BMP-2) antibody as previously described.7,9
Quantitative real-time PCR
Tissue specimens were harvested in the 2nd, 4th, 8th, 12th, and 16th weeks after implantation and immediately immersed in the TRIzol® Reagent (Life Technologies, USA). The tissue specimens were then grounded until there was no obvious tissue residual. Total RNA was extracted from the grounded tissue using TRIzol Reagent. All RNA samples were then treated with RNase-Free DNase I (Qiagen, Valencia, CA) to digest the genomic DNA. Purified total RNA (500 ng) was reverse transcribed into complementary DNA using the PrimeScript™ RT Master Mix (TaKaRa, Japan). Quantitative real-time PCR (qRT-PCR) was performed using a 7900 Real-Time PCR System (Applied Biosystems, Warrington, UK) with Power SYBR Green PCR Master Mix (Applied Biosystems). The relative gene expression was calculated using the following equation: ΔCt = Ct (test genes)−Ct (ACTB); ΔΔCt = ΔCt (newborn laminae) −ΔCt (normal laminae); fold change = 2−ΔΔCt. The gene-specific primers used for TNF-β, IL-6, OPN, BMP2, RUNX2, and ACTB are listed in Table 1.
Sequences of Oligonucleotide Primers Used for Quantitative Real-Time PCR
Statistical analyses
Statistical analyses were performed using SPSS version 19.0 for Windows. One-way analysis of variance (ANOVA) was used to compare the variables between the groups. Multiple comparisons were performed by one-way ANOVA test followed by post hoc contrasts performed by least significant difference test (if the ANOVA results were significant). Data were presented as mean ± SD (*represents p < 0.05 and **represents p < 0.01).
Results
Characterization of WJ-MSCs
After four passages of culture, the morphology of WJ-MSCs became evenly elongated and spindle shaped and arranged like a vortex (Fig. 2A). For multilineage differentiation, after 3 weeks of osteogenic induction, extensive formation of mineralization nodes was formed as shown by Alizarin Red S staining (Fig. 2B). After 3 weeks of adipogenic induction, the lipid droplets were formed extensively and circled the nucleus in WJ-MSCs, and some WJ-MSCs also formed a big fat vacuole pressing the nucleus (Fig. 2C). Cartilaginous node also formed after 3 weeks of chondrogenic induction as shown by Alcian Blue staining (Fig. 2D).
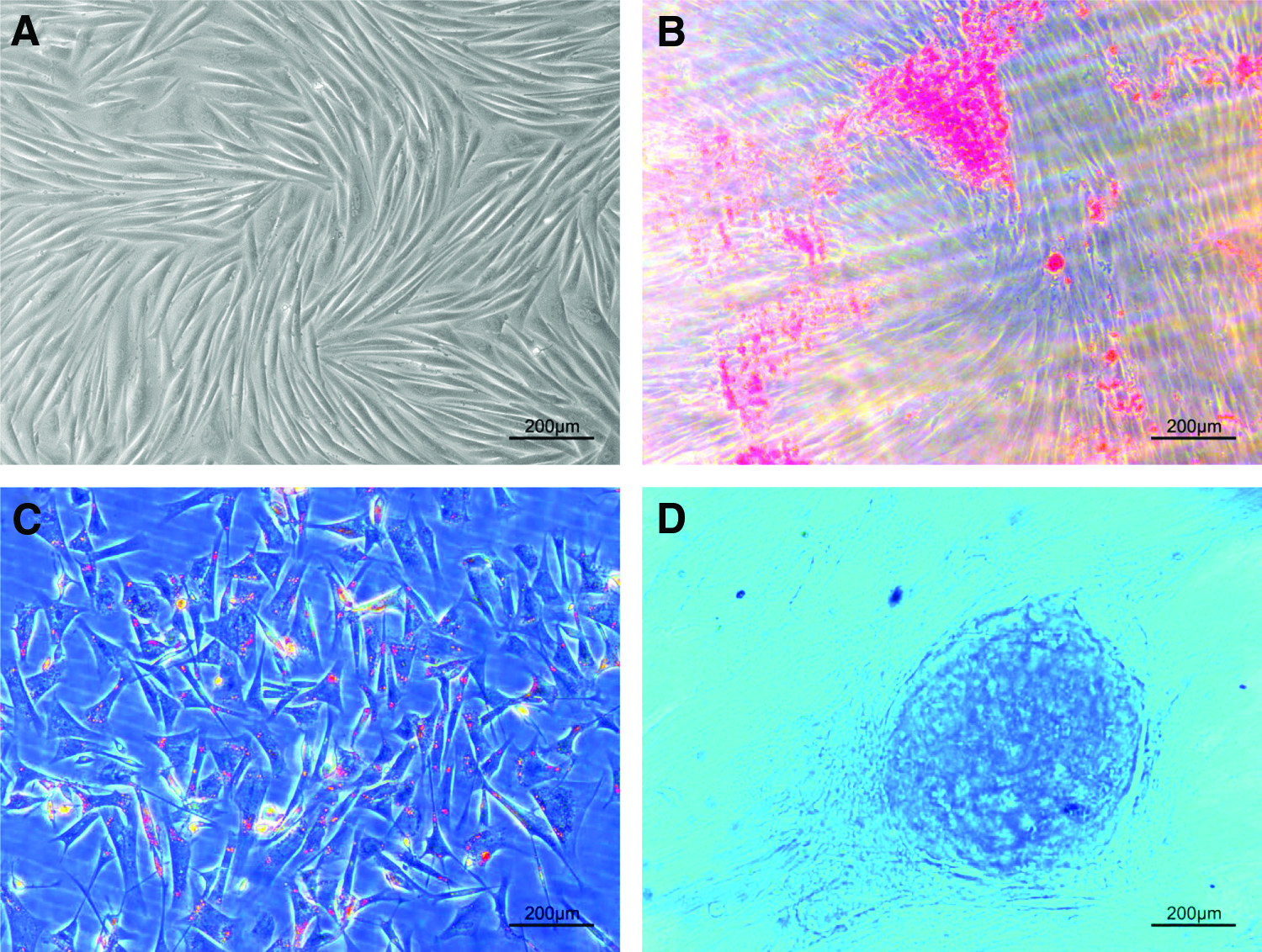
Representative images of WJ-MSCs at passage 4.
Distribution of DiI-labeled osteogenic WJ-MSCs within hydroxyapatite-collagen I scaffold
Osteogenic WJ-MSCs were observed to be evenly adhered to the surface of hydroxyapatite-collagen I scaffold and gradually grew into the scaffolds 2 days after transplantation. In addition, the osteogenic WJ-MSCs within the hydroxyapatite-collagen I scaffold were arranged in multilayers shown by the inverted fluorescence contrast phase microscope (Fig. 3C, D).

Micro-CT examination
Micro-CT scanning showed that bone density of de novo laminae in both groups remained low at the 2nd week and increased in the following weeks. In the orthotopic lamina group, the bone density increased gradually from the bone defect ends toward the middle, and complete de novo laminae were formed in the 12th week, with a disordered arrangement of trabeculae. The trabeculae became more orderly in the 16th week, and the inner surface of de novo laminae showed a curvature and smoothness similar to those of the native laminae. In the ectopic lamina group, the bone density gradually increased from the outer surface to the inner surface, and the ectopic laminae gradually merged with the native laminae in the 16th week (Fig. 4A).
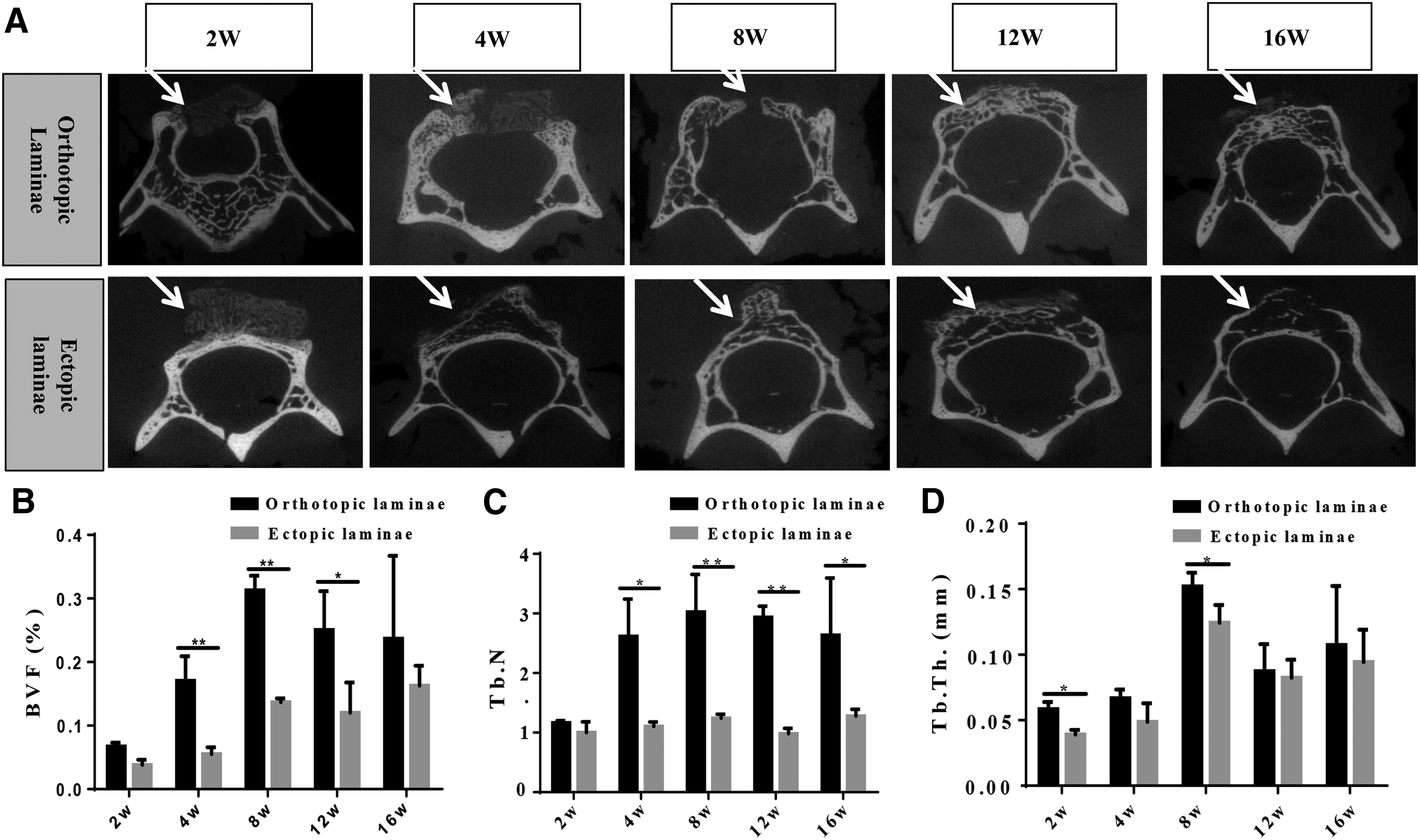
BVF, Tb.Th, and Tb.N increased continually in the first 8 weeks in both groups and their values in the orthotopic lamina group were higher than those in the ectopic lamina group (p < 0.05). In the 12th week, their values remained stable, and the Tb.N values in the orthotopic lamina group was higher than those in the ectopic lamina group (p < 0.05). In the 16th week, their values increased slightly, and the Tb.N values in the orthotopic lamina group were still higher than those in the ectopic lamina group (p < 0.05) (Fig. 4B–D).
3D system of coordinates was constructed with the cross-section of the spinal canal as the X-axis, coronal plane as the Y-axis, and the sagittal plane as the Z-axis as shown in Figure 5C. Tb.N values of the three different axes at different time points were analyzed. In the orthotopic lamina group, the Tb.N values on the X-axis and Y-axis were significantly higher than those on the Z-axis at 2, 4, and 8 weeks postimplantation (p < 0.05) (Fig. 5A). The differences of the three axes for the ectopic lamina group were less significant than the orthotopic lamina group (Fig. 5B).
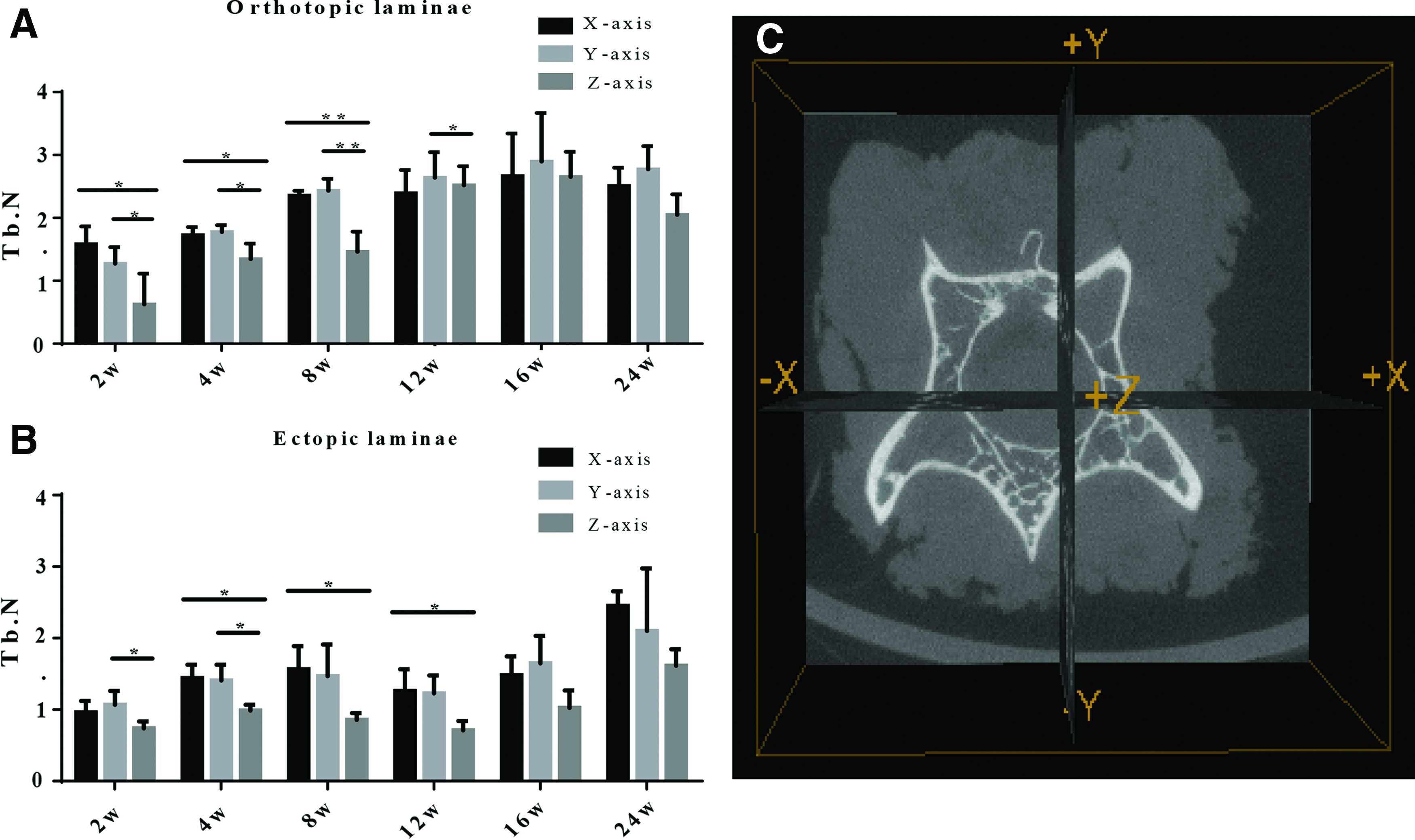
Comparison of Tb.N of three different axes in the orthotopic group
Histological and immunohistochemical analysis
H&E staining results showed that both groups experienced scaffold degradation and trabecular formation during the first 12 weeks, during which the trabeculae were disorganized and unsystematically arranged. In the 16th week, the trabeculae became more orderly and organized (Fig. 6A).

Immunohistochemical staining results showed that, in the orthotopic lamina group, the optical density (OD) values of BMP-2 increased in the first 8 weeks and then declined gradually. In the ectopic lamina group, the OD values of BMP-2 increased for the first 12 weeks and then declined. The OD values of BMP-2 in the orthotopic lamina group were significantly higher than those in the ectopic lamina group in the 4th and 8th weeks postimplantation (p < 0.05) (Fig. 6B–D).
qRT-PCR analysis
In the orthotopic lamina group, RUNX2 messenger RNA (mRNA) expression levels increased during the first 8 weeks, peaked at 8 weeks, and then decreased. While, in the ectopic lamina group, RUNX2 mRNA expression levels peaked in the 12th week and decreased. RUNX2 mRNA levels in the orthotopic lamina group were significantly higher than those in the ectopic lamina group in the 4th, 8th, and 12th weeks (p < 0.05) (Fig. 6E). The mRNA expression levels of BMP2 in both groups showed the similar trend as those of RUNX2, and the BMP2 mRNA expression levels in orthotopic lamina group were significantly higher than those of ectopic lamina group in the 2nd, 4th, and 8th weeks (p < 0.05) (Fig. 6F). OPN mRNA expression increased in the first 12 weeks in both groups and then it began to decline in the 16th week in the ectopic lamina group. The OPN mRNA expression levels in the orthotopic lamina group were higher than those in the ectopic lamina group in the 4th, 8th, and 16th weeks (p < 0.05) (Fig. 6G).
TNF-β and IL-6 mRNA expression levels increased during the first 4 weeks and decreased greatly from the 8th week. There was no significant difference between the orthotopic and ectopic lamina groups (p > 0.05) (Fig. 6H, I).
Discussion
Tissue engineering techniques have been successfully applied to reconstruct the artificial vertebral laminae to avoid the failed back surgery syndrome.7,9 In this study, we continued to investigate the roles of biological and mechanical factors underlying the formation of de novo laminae.
In the beginning of study design, we meant to construct an animal model only exposed to the mechanical stimulation of CSFP and free of the biological stimulation of bone defect, but the proper animal model could not be constructed. The reasons are as follows. The method of constructing animal model by removing the native laminae and placing rigid insulating film could guarantee the similar biological stimulation of bone defect, but the removal of native laminae could also release the CSFP stress and no isolation film could absolutely seal it, considering the mechanotransduction property and fluidity of tissue fluid.17,18 In addition, the use of rigid film would also have the potential threat of spine compression, which severely undermined the original intention of our study to prevent spine compression. Thus, we chose the current animal model to guarantee the difference of CSFP stress at the sacrifice of biological stimulation of bone defect. It is well known that the biological stimulation of cytokines released from the bone defect would impact the viability of engineered bone grafts.11,19,20 Both groups had bone defect and were influenced by its biological stimulation. As a compromise, we designed a comparison study and investigated both the biological and mechanical stimulations in this study. To assure the similar biological stimulation in both groups, we enlarged the cancellous bone defect in the ectopic lamina group to the size of 10 × 4 mm, similar to that of orthotopic lamina group (10 × 2 × 2 mm).
In the ectopic lamina group, the de novo laminae were exposed only to the biological stimulation of bone defect and free of the mechanical stimulation of CSFP stress, thereby its de novo bone indices suggesting the impact of biological stimulations on the bone formation of artificial laminae. While in the orthotopic lamina group, the de novo laminae were exposed to both the biological stimulation of bone defect and the mechanical stimulation of CSFP stress, suggesting the impact of biological and mechanical stimulations. Furthermore, the difference of de novo bone indices between orthotopic and ectopic lamina groups suggested the impact of mechanical stimulation of CSFP stress.
After the removal of lamina or spinous process, the bone ends can release a series of inflammatory and neovascularization factors that would provide the initial impetus for the bone formation.10,11 IL-6 and TGF-β were inflammatory cytokines released from the bone defect and played a key role in the migration, proliferation, and differentiation of MSCs and osteoblasts, acting as the initial impetus for the bone formation.10,11,21 In this study, we determined their mRNA expression levels and found that there was no significant difference between two groups, indicating the similar biological stimulation in both groups. Their expression levels increased in the 2nd and 4th weeks and declined greatly in the 8th week, suggesting that they mainly took effect in the early stage of bone formation. Correspondingly, the mRNA expression levels of osteogenic genes increased and had no significant difference in both groups in the 2nd week, indicating the similar impact of biological stimulation in this stage. Consistently, the cancellous microstructure parameters of both groups also showed little difference in the 2nd week, indicating the similar bone formation during this stage.
Mechanical stimulation is not required for initial bone formation, but bone formation is enhanced by the mechanical loads placed on bones at the late stage.16,22–24 In this study, the difference of osteogenic indices between the orthotopic and ectopic lamina groups was used to evaluate the impact of mechanical stimulation on the osteogenesis of de novo laminae. We found that the mRNA expression levels of RUNX2, BMP2, and OPN in the orthotopic lamina group were higher than those in the ectopic lamina group in the 4th, 8th, and 12th weeks. The values of BVF, Tb.Th, and Tb.N in the orthotopic lamina group were higher than those in the ectopic lamina group in the 4th, 8th, and 12th weeks, consistent with the expression trends of osteogenic genes. In the 12th week, complete de novo laminae formed in the orthotopic lamina group, as shown by the micro-CT scanning and H&E staining. This suggested that the mechanical stimulation of CSFP began to take effect in the 4th week and sustained throughout the whole process of lamina formation. In this stage, the mechanical stimulation began to take the predominant part, promoting the osteogenesis of de novo laminae, while the impact of biological stimulation gradually faded out as shown by the low expression levels of osteogenic genes in the ectopic lamina group.
We also found the Tb.N of the X-axis and Y-axis was higher compared with the Z-axis in the orthotopic lamina group, whereas the difference in the ectopic lamina group was less significant. The X- and Y-axis planes directly bear the brunt of CSFP and reflect the impact of CSFP, and the directions withstanding CSFP form more trabeculae than the direction shielding CSFP, consistent with the Wolff's law.
Studies22,23 have shown that fluid shear stress can regulate the proliferation and differentiation of osteoblasts, promoting the secretion of OPN through activation of MAPK signal pathways and the secretion of RhoA, ROCK II, Runx2, Osteocalcin, and ALP. Pulsation pressure can also activate the Akt and MAPK signal pathways to regulate the proliferation, differentiation, and migration of osteoblasts. 25 The distinct signaling pathways involved could be the possible explanation why the expression trends of RUNX2 and BMP2 were different from those of OPN, since the RUNX2 and BMP2 are the downstream molecules of Wnt signaling pathway. CSFP stress could promote the expression levels of RUNX2, BMP2, and OPN, eventually leading to phenotypic differences manifested as Tb.N, Tb.Th, and BV/TV.
In the ectopic lamina group, there was a rapid increase in the expression levels of osteogenic genes in the 12th week, while the expression levels of IL-6 and TGF-β decreased greatly. It is well known that fracture healing is commonly composed of four phases as follows: (1) inflammation phase, (2) soft callus formation (cartilage form), (3) hard callus formation (woven bone), and (4) bone remodeling (lamellar bone). In the 12th week, the callus formation phase finished, and the bone remodeling stage began. In this stage, the woven bone was reabsorbed, and the lamellar bone was formed, as also shown by the Figures 4A and 6A. The bone remodeling needs the coordinated activities of osteoblast and osteoclast.26,27 This explains the increase of osteogenic genes in the 12th week in the ectopic lamina group.
In conclusion, the biological stimulation of bone defect initiated the early onset osteogenesis, and CSFP promoted the osteogenesis of de novo laminae during the reconstruction of de novo laminae.
Footnotes
Acknowledgments
The study was supported by National Natural Science Foundation of China (Grant No. H0606); by Natural Science Foundation of Minhang District, Shanghai (Grant No. 81672179); and by the Foundation of Shanghai Municipal Commission of Health and Family Planning (Grant Nos. 20134305 and 201440509).
Disclosure Statement
No competing financial interests exist.