
Abstract
MicroRNA (miRNA) therapeutics is increasingly being developed to either target bone-related diseases such as osteoporosis and osteoarthritis or as the basis for novel bone tissue engineering strategies. A number of miRNAs have been reported as potential osteo-therapeutics but no consensus has yet been established on the optimal target. miR-16 has been studied extensively in nonosteogenic functions and used as functionality reporter target in the development of nonviral miRNA delivery platforms. This study hypothesized that miR-16 may also play an inhibitory role in osteogenesis due to its ability to directly target Smad5 and AcvR2a. This study thus aimed to assess the potential of miR-16 inhibition to increase osteogenesis in human mesenchymal stem cells (hMSCs) using a previously established miRNA delivery platform composed of nanohydroxyapatite (nHA) particles as nonviral vectors in combination with collagen-nHA scaffolds designed specifically for bone repair. Initial results showed that antagomiR-16 delivery efficiently increased the relative levels of both putative targets and Runx2, the key transcription factor for osteogenesis, while also increasing osteocalcin levels. Furthermore, significant increases in mineral calcium deposition by hMSCs were found in both monolayer and most importantly in scaffold-based osteodifferentiation studies, ultimately demonstrating that miR-16 inhibition further enhances the therapeutic potential of a scaffold with known potential for bone repair applications and thus holds significant therapeutic potential as a novel bone tissue engineering strategy. Furthermore, we suggest that harnessing the additional functions known to miR-16 by incorporating either its enhancers or inhibitors to tissue-specific tailored scaffolds provides exciting opportunities for a diverse range of therapeutic indications.
Introduction
MicroRNAs (miRNAs) are endogenous RNA molecules involved in posttranscriptional gene regulation by both perfect and partial sequence complementarity binding to the untranslated region of messenger (m)RNA targets. Thus, miRNAs are capable of binding to multiple mRNA targets and are postulated to inhibit the protein expression of entire gene cohorts.1–5 The availability of synthetic enhancers (mimics) or inhibitors (antagomiRs) has triggered the investigation of the potential of miRNA therapeutics. Hence, miRNA therapeutics is increasingly proposed as a novel strategy to treat diseases such as osteoporosis and osteoarthritis6,7 or to provide improved bone tissue engineering strategies targeting a global market valued in 2015 at $2.5 billion.8,9 Recently, a selection of miRNAs have been collectively identified to influence osteogenesis 10 and it has also been reported that, dependent on cell types and species, particular miRNAs can adopt distinct roles and functions.11–13 Therefore, control of function specificity remains key for the application of miRNA therapeutics and a consensus regarding the optimal target remains elusive.
One strategy to control miRNA function specificity is the application of a localized and tissue-targeted delivery system. 14 Previously within our laboratory, porous collagen scaffolds have been targeted to bone tissue repair by incorporating different phases and particulate forms of hydroxyapatite (HA).15,16 These composite collagen-HA scaffolds have significant bone healing efficacy in small and large animals.17–22 These scaffolds have been further utilized as delivery platforms for plasmid (p)DNA encoding bone morphogenetic protein (BMP)-2 and vascular endothelial growth factor (VEGF)23–25 to enhance bone repair. One variant, a coll-nanoHA (nHA) scaffold, has recently proven effective in delivering reporter GAPDH-silencing miR-mimics and miR-16 inhibitors 26 —targets used to demonstrate proof of concept without therapeutic indication. However, when used to deliver an inhibitor for the known antiosteogenic miR-133a, this scaffold platform also demonstrated bone repair efficacy 27 (Mencía Castaño et al. 2015, Under Revision). In this study, we sought to answer the question as to whether the highly effective miR-16 inhibition we previously observed might provide a novel therapeutic to improve osteogenesis by human mesenchymal stem cells (hMSCs).
miR-16 has been studied extensively 28 and implicated in ∼50 different pathologies spanning from different cancer types29–33 to the progression of viral, 34 bacterial, 35 and parasitic infections 36 and the activity of the innate immune system, 37 to the function of the respiratory epithelium 38 and even to the transport of neurotransmitters such as serotonin 39 or several neurological diseases such as Parkinson's 40 or Alzheimer's. 41 miR-16 has also a well-established antiangiogenic role42,43 and has been found increased in rheumatoid arthritis patients versus healthy controls,44,45 while a downregulation was seen in odontogenic keratocysts 46 and osteosarcoma.47,48 Currently, there is no defined role for miR-16 in osteogenesis, although Eguchi et al. designed miR-16 as a stemness maintenance marker that deterred osteocyte maturation in MSC osteogenesis. 49
Interestingly, studies at the developmental 50 or genome-wide screening level 51 have pointed at two targets that could link miR-16 to osteogenesis: small mothers against decapentaplegic (Smad)-5 and activin receptor (AcvR)-2a involved, respectively, in BMP and nodal/transforming growth factor (TGF)-β/activin signaling. While both of these signaling pathways are involved in multiple developmental processes, they are also tightly related to MSC osteogenesis. A direct miR-16::Smad5 interaction has been reported at a proteomic level in HeLa cells, 51 and the implication of miR-16 in BMP signaling was shown in mouse bone cells using quantitative polymerase chain reaction (qPCR). 52 Finally, the direct miR-16::AcvR2a targeting was demonstrated in a study performed in the human hepatocarcinoma cell line HepG2 and in Xenopus embryos. 50 If miR-16 inhibits Smad5 and AcvR2a, which play a positive role in osteogenesis, we hypothesized that treatment with antagomiR-16 may in turn enhance both of these targets and thus promote osteogenesis by MSCs. This study thus aimed to elucidate the inhibitory role of miR-16 in osteogenesis and subsequently assess the potential of harnessing miR-16 inhibition with the nonviral nHA delivery system, to ultimately produce antagomiR-16-loaded collagen-nHA (coll-nHA) scaffolds with potential as novel off-the-shelf platforms for improved bone tissue engineering. Specifically, we first aimed to establish the role of miR-16 in osteogenesis via bioinformatic analysis, to then determine the ability of nHA-based antagomiR-16 delivery to hMSCs to effectively enhance the level of the two putative direct targets, Smad5 and AcvR2a, and most importantly assess the ability of this treatment to improve osteogenesis not only in the monolayer culture but also when antagomiR-16 is delivered using the coll-nHA scaffold as off-the-shelf three-dimensional (3D) platforms.
Materials and Methods
Bioinformatic analysis
Bioinformatic analysis provides detailed information about the features of putative and experimentally validated interactions of a given miRNA entry, which allows to establish a suggested function for any given miRNA. To gather knowledge regarding the hypothesis of possible role of miR-16 in osteogenesis, online databases were explored in terms of (1) identification of putative mRNA targets implicated in osteogenesis and (2) identification of validated targets for which the miRNA interaction was assessed in the context of a cellular function different to osteogenesis. A subsequent analysis was carried out to assess (3) homology in the interaction of miRNAs and their relevant direct targets across human, rat, and mouse species, as well as (4) the types of miRNA::mRNA target interaction taking place and (5) occupancy of nearby seed region by other neighboring miRNAs. All of these features were explored by searching the following peer-consulted databases available online: TargetScan, 52 microRNA.org, 53 and miRTarBase. 29
hMSC culture, expansion, and osteodifferentiation
Bone marrow aspirates from the iliac crest of healthy human volunteers were used to obtain hMSCs following stern phenotype analysis, in compliance with the procedures approved by the Clinical Research Ethics Committee at University College Hospital Galway and after obtaining written informed consent from all subjects. hMSCs were expanded in low-glucose DMEM supplemented with 10% fetal bovine serum plus 1% penicillin/streptomycin (all from Sigma-Aldrich, Ireland) until passage number 4 for experimentation. For osteodifferentiation assays, standard growth medium was supplemented with 50 μg/mL ascorbic acid-2-phosphate, 10 nM β-glycerophosphate, and 100 nM dexamethasone (all from Sigma-Aldrich). This was administered to cells every 3/4 days until the experiment endpoint. Untreated cells either in standard or osteogenic complete growth medium for the duration of the assay served as controls.
hMSC transfection with antagomiR-16 and nHA particles
To study the effect of miR-16 inhibition on the identified targets and on the ability of hMSCs to undergo osteogenic differentiation, cells were transfected with antagomiR-16 and nHA particles. To do so, 3 × 104 cells per well of a six-well plate were seeded 24 h in advance of the transfection and complexes of antagomiR and nHA particles were prepared immediately before transfection as per a previously established protocol.26,54 Briefly, a volume of 12 mM sodium phosphate solution containing 0.017% Darvan 821A (RT Vanderbilt, Norwalk,) was added to an equal volume of 20 mM calcium chloride and 0.2 μm sterile filtered to obtain nHA particles; 150 μL nHA particles were then combined with 20 nM miRIDIAN antagomiR-16 or scrambled (scr) antagomiR (Dharmacon-GE Healthcare, United Kingdom) and immediately administered per well. The supplier-designed antagomiRs are based, respectively, on the MIMAT0000069 sequence of hsa-miR-16(−5p) [UAGCAGCACGUAAAUAUUGGCG] and the MIMAT0000039 sequence of cel-miR-67 [UCACAACCUCCUAGAAAGAGUAGA].
Assessment of effective antagomiR-16 transfection, genetic manipulation, and osteogenic gene expression using quantitative real-time polymerase chain reaction
Quantitative real-time PCR (qRT-PCR) analysis was carried out to assess the effective antagomiR-16 transfection at three levels: (1) miR-16, (2) relative mRNA expression of the putative direct miRNA targets Smad5 and AcvR2a, and (3) relative mRNA expression of the key transcription factor for osteogenesis Runx2 and the osteogenesis marker osteocalcin (OCN). Total RNA extraction was done using QIAzol in combination with an miRNeasy kit (Qiagen, United Kingdom) under the manufacturer's instructions. The hsa-miR-16 TaqMan® microRNA assay was combined with the corresponding TaqMan assay kit (Biosciences, Ireland); QuantiTect Reverse Transcription kit and SYBR Green master mix (Roche, Ireland) were applied in tandem with the following validated predesigned human QuantiTect primer assays (Qiagen): Hs_SMAD5_va.1_SG, Hs_AcvR2a_1_SG, Hs_Runx2_1_SG, and Hs_BGLAP_1_SG. The scr group was set as reference to calculate relative expression, normalized to 18S ribosomal RNA, using the 2−ΔΔCt method.
Assessment of alkaline phosphatase activity as a biofunctional marker of osteogenesis
To assess the effect of antagomiR-16 delivery on hMSC osteogenesis, we first analyzed the specific biofunctional marker, that is, assessment of alkaline phosphatase (ALP) activity. To do so, the SensoLyte® pNPP Alkaline Phosphatase Assay Kit (Cambridge Bioscience, United Kingdom) was used under the manufacturer's instructions on samples harvested at 10 days post-treatment. Absorbance of the color product generated was read at 405 nm using a Varioskan Flash plate reader (Thermo Scientific, Ireland) and SkanIt® for Varioskan software. The Quant-iT PicoGreen dsDNA kit (Invitrogen, Ireland) assay was performed following the manufacturer's instructions to normalize levels of ALP activity to cell number.
Assessment of mineralization as end-stage marker of osteogenesis
To complete the assessment of the effect of antagomiR-16 delivery on hMSC osteogenesis, the presence of calcium deposits in the extracellular matrix (ECM) was studied as an endpoint marker. To visualize calcium deposits, samples were stained directly on culture plates with 2% alizarin red, which stains calcium red. Briefly, samples were fixed using 10% formalin and subjected to alizarin red staining for 5 min, following which excess staining was removed rinsing with distilled water. Microscopic imaging was carried out using an inverted microscope coupled to the LASV4.5 digital imaging system (Leica, Germany). In addition, the Calcium Liquicolor kit (Stanbio Laboratories, United Kingdom) was used under the manufacturer's instructions on samples prepared in 0.5 M HCl; absorbance of the color product generated was read at 595 nm using a Varioskan Flash plate reader as before and also normalized to cell number again using the Quant-iT PicoGreen dsDNA assay.
AntagomiR-16-loaded collagen-nHA scaffolds: preparation, hMSC culture, and mineralization analysis
coll-nHA scaffolds, 1:1 weight ratio of collagen to nHA, were prepared by freeze-drying following a protocol developed in-house, which provides scaffolds of >90% porosity with uniform HA distribution and pores of ∼100 μm. 16 Briefly, suspensions combining collagen type I (Integra Life Sciences) and freshly precipitated nHA particles were homogenized in 0.05 M acetic acid and lyophilized at a −40°C in a VirTis Genesis 25 EL freeze-dryer (Biopharma, United Kingdom). Scaffolds were sterilized by dehydrothermal treatment, cut into 8 × 4 mm discs, and crosslinked for 2 h using 14 mM 1-(3-dimethylaminopropyl)-3-ethylcarbodiimide hydrochloride (EDAC) and 5.5 mM N-hydroxysuccinimide (NHS) (both from Sigma-Aldrich). Coll-nHA scaffolds were then soak-loaded on both sides with a 150 μL total volume of blank nHA particles or complexes of 20 nM antagomiR-16 or -scr. After having loaded the blank particles or antagomiRs onto the coll-nHA scaffolds, hMSCs (passage 5) were seeded dropwise onto both sides of the scaffold at 3 × 105 cells per scaffold. For mineralization assessment, scaffolds were fixed in 10% formalin and cut in serial sections of 7 μm thickness, which were then stained with 2% alizarin red and imaged using a Nikon Eclipse 90i plus DS Ri1 with NIS Elements software (Nikon Instruments, Netherlands). Calcium deposition was also quantified, as described in the section immediately above, where calcium content corresponding to noncultured scaffolds was subtracted from all values.
Statistical analysis
Results were analyzed using one-way ANOVA plus Tukey post hoc tests (n ≥ 3). p < 0.05, p < 0.01, and p < 0.001 were considered significant, highly significant, and very highly significant differences.
Results
Bioinformatic analysis supports a negative role of miR-16 in osteogenesis
Bioinformatic analysis provided the identification of 1273 predicted hit interactions between potential mRNA targets and miR-16 in human species, nine of which were of interest for osteogenesis (Fig. 1a). These target hits were then compared with the database of validated targets, miRTarBase. More than 90% of the total predicted targets have been experimentally validated. Interestingly, among the targets validated in human cells or tissues, the screening identified seven targets of interest for osteogenesis in addition to Smad5 and AcvR2a, including COL4A2 and IGF2R, which also play a positive role in osteogenesis. Taken together, these results support our central hypothesis and suggest an inhibitory role of miR-16 in MSC osteogenesis. Bioinformatic databases were then searched to document further details of the miR-16::Smad5 and miR-16::AcvR2a interactions in terms of homology across human, rat, and mouse (Fig. 1b). First for Smad5, a double number of binding sites was predicted for the interaction in mouse, and rat context percentile scores were lower than those of both human and mice. In relation to AcvR2a, the 7mer-m8 binding type, together with context percentile score, and remaining parameters tested were maintained across species. Taken together, these data confirmed the promising potential of miR-16 to exert multiple functions related with osteogenesis across the three species analyzed in this study.
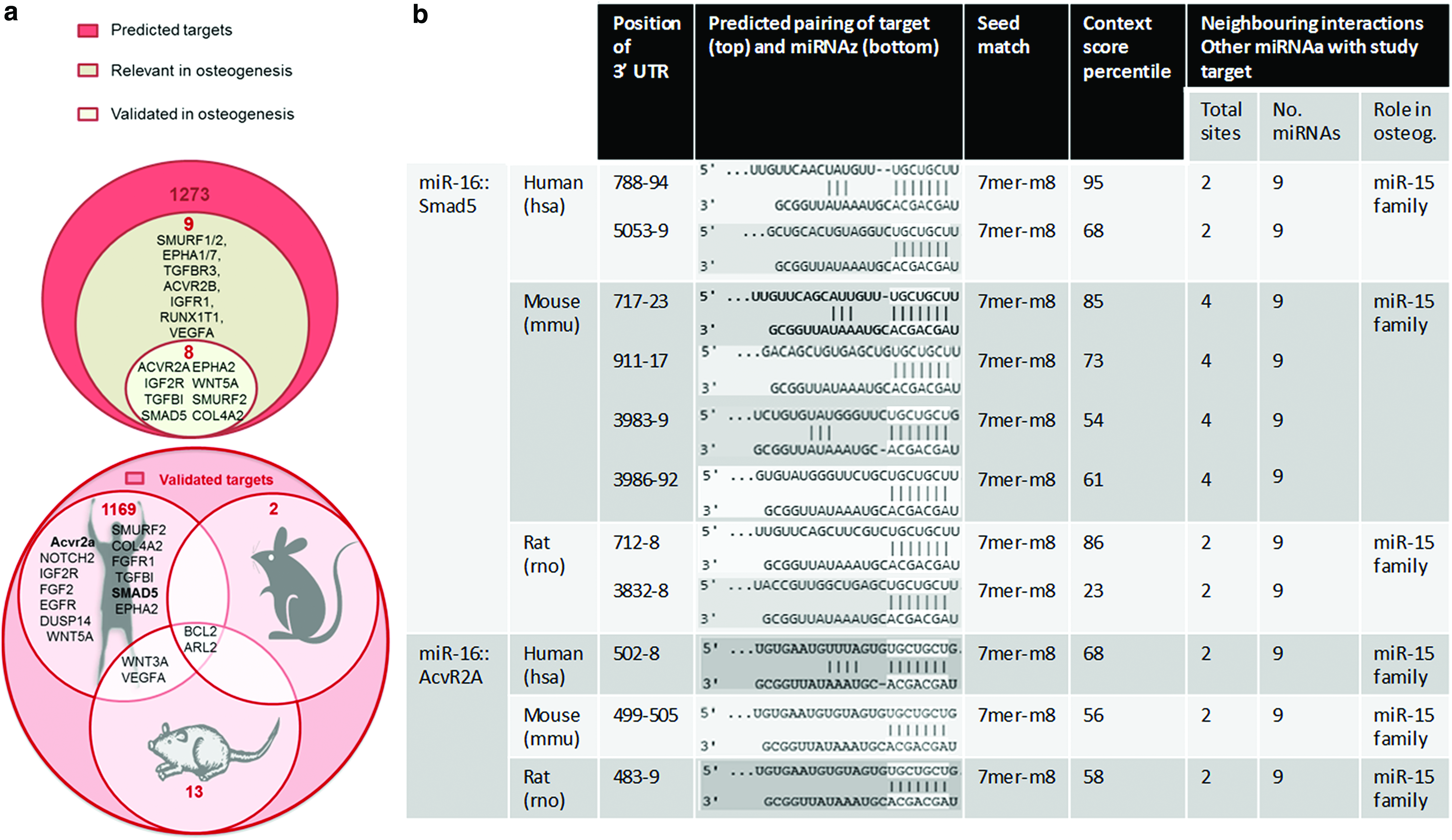
Bioinformatic exploration of miR-16.
Effective manipulation of intracellular miR-16 level in hMSC was achieved using nHA-based delivery
Manipulation of miR-16 levels with antagomiR-16 treatment during hMSC osteogenic culture proved highly efficient, yielding higher than 90% reduction of miR-16 relative expression levels (Fig. 2a). As a consequence of this efficient inhibition of miR-16, both AcvR2a and Smad5 were increased at the mRNA level when analyzed at day 7 of the osteogenic monolayer culture, achieving, respectively, a 2.54 (±0.55)-fold and 10.92 (±1.12)-fold increase over the antagomiR-scr control, respectively (Fig. 2b, c). These data thus demonstrate for the first time an underlying link between manipulation of intracellular miR-16 levels and increased positive signaling molecules in the BMP and activin signaling pathways in hMSCs undergoing osteogenesis.
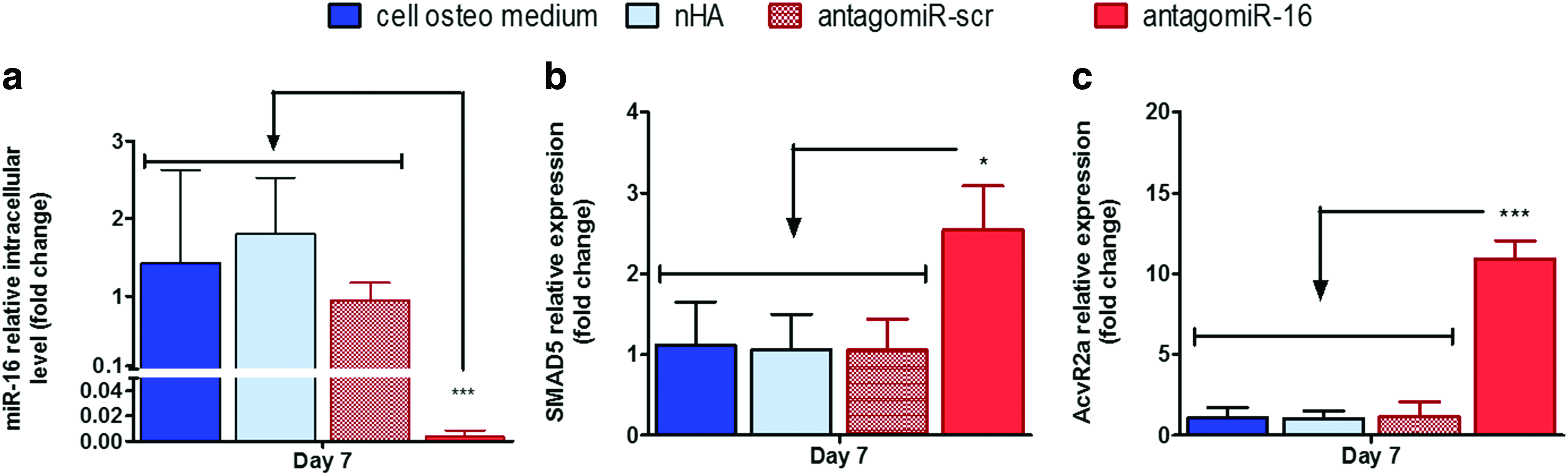
qPCR analysis of miR-16 manipulation and its effect on putative miR-16 direct targets in hMSCs after 7 days in osteogenic monolayer culture.
AntagomiR-16 treatment enhanced hMSC osteogenic gene expression and ALP activity
Having confirmed that efficient miR-16 inhibition can lead to enhanced relative levels of both Smad5 and AcvR2a, we carried out an evaluation of the downstream effect of this treatment on osteogenic differentiation of hMSCs. Initially at the 7-day time point, Runx2 and OCN mRNA expression was increased to 6.29 (±2.3)-fold and 8.19 (±1.96)-fold, respectively, in comparison with the antagomiR-16 treatment (Fig. 3a, b), supporting that inhibition of miR-16 can have a positive influence in hMSC osteogenesis at the gene level. The biofunctional assessment of osteogenesis by ALP activity quantification showed 11.32- and 8.62-fold higher levels in the antagomiR-16 treatment than in untreated cells and cells treated with blank nHA particles, respectively (Fig. 3c). These levels were also 1.22-fold higher than the antagomiR-scr group, which surprisingly enhanced ALP activity over untreated cells. In summary, this result supported the ability of antagomiR-16 treatment to enhance a panel of osteogenesis markers in hMSCs at the two time points assessed, specifically 7 and 10 days in osteogenic monolayer culture.
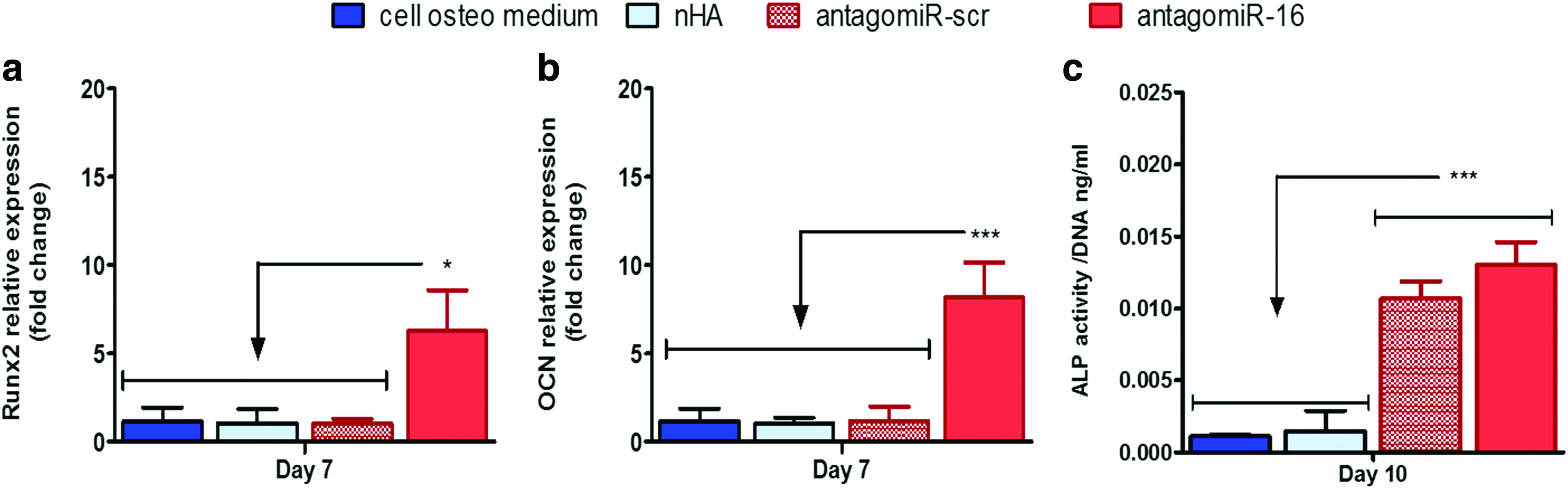
Analysis of the effect of miR-16 inhibition on osteogenic gene expression and osteogenesis bioactivity.
Mineral deposition was enhanced by antagomiR-16 treatment and antagomiR-16-loaded scaffolds
As an endpoint assessment of antagomiR-16 inhibition-mediated osteogenesis, significantly enhanced extracellular calcium deposition was demonstrated in hMSCs treated with antagomiR-16 in monolayer culture (Fig. 4a, b). Originally, alizarin red staining showed consistently abundant presence of calcium deposits in the antagomiR-16 group; in comparison, all control groups showed reduced positive staining, with deposit-free areas alternated with dispersed calcium deposits still evident in the antagomiR-scr control (Fig. 4a). Accordingly, the quantification of calcium levels deposited per cell by monolayer hMSCs demonstrated a significant 3.23-fold increase over untreated cells after antagomiR-16 delivery; while calcium deposition, relative to untreated cells, was progressively increased by the treatment with blank nHA particles or the antagomiR-scr control, this effect was largely surpassed by the antagomiR-16 treatment (Fig. 4b). More importantly, this beneficial effect translated seamlessly to the 3D microenvironment of the antagomiR-16-loaded scaffolds. Alizarin red staining of the 3D samples corroborated the prominent calcium deposition by hMSCs within the antagomiR-16-loaded scaffolds in contrast with the staining-free areas in the antagomiR-scr control scaffolds (Fig. 4d). Correspondingly, calcium quantification per cell confirmed the antagomiR-16 group being highest, at ∼2.42-fold greater than the untreated cells cultured on the antagomiR-free scaffolds. This effect indicated a significant increase versus all other groups tested and was again followed by the progressive increase, relative to untreated cells, across the remaining groups (Fig. 4e). Taken together, the results indicated the potential of antagomiR-16 treatment to enhance calcium mineralization by hMSCs, denoting clear potential of the antagomiR-16-loaded scaffold as an off-the-shelf platform for bone tissue engineering.
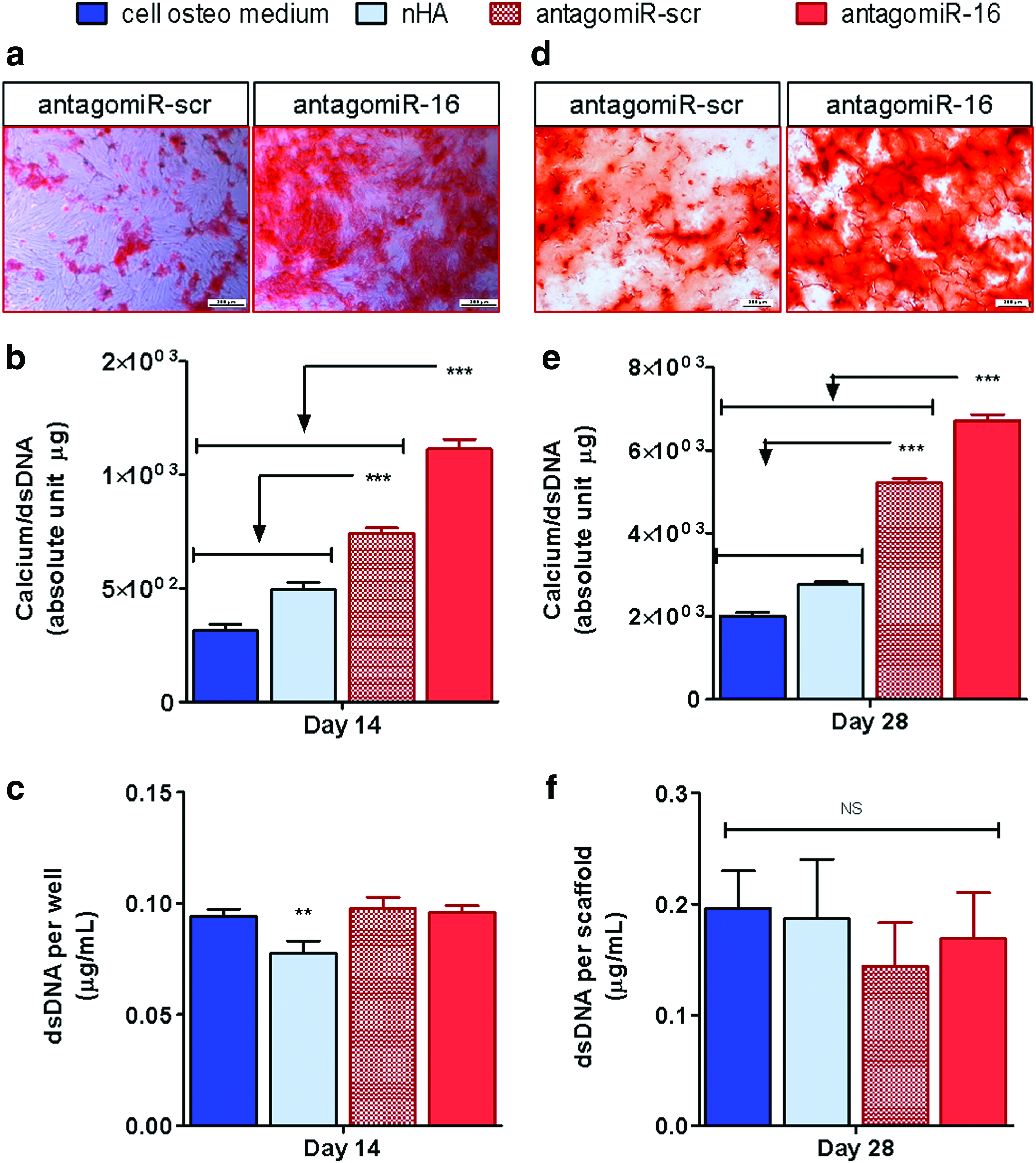
Analysis of the effect of miR-16 inhibition on mineral deposition in monolayer and 3D osteogenic culture.
Discussion
The controlled delivery of miRNAs has received much attention as novel therapeutics in tissue engineering strategies. However, translating such miRNA therapeutic strategies to the clinic has been limited by poor delivery systems and difficulties reaching consensus on best targets for any given application. This study identified for the first time a negative role of miR-16 in osteogenesis and demonstrated the potential of miR-16 inhibition as an osteotherapeutic by utilizing nHA particles as nonviral vectors in combination with coll-nHA scaffolds designed specifically for bone repair. The study identified a link between miR-16 and osteogenesis by identifying multiple putative targets through bioinformatics and then specifically focusing on two direct targets involved in BMP and activin signaling, that is, Smad5 and AcvR2a, respectively. The results demonstrated that efficient miR-16 inhibition significantly promoted Runx2 and OCN expression as well as ALP activity. Ultimately, antagomiR-16-loaded scaffolds significantly increased mineral calcium deposition by hMSCs, indicating that miR-16 inhibition holds therapeutic potential as a novel bone tissue engineering strategy.
The two putative targets of focus in this study, Smad5 and AcvR2a, were identified in the bioinformatic analysis to present robust characteristics for direct interaction with miR-16, including the presence of multiple seed sites, the high context percentile scores, and the interspecies conserved type of binding and neighboring interacting miRNAs. In addition, beyond the known functions of miR-16,28–48 the bioinformatic data reported the additional direct interaction of miR-16 with other targets such as IGF2R and COL4A2, beneficial for osteogenesis, 55 further supporting the negative role of this miRNA in osteogenesis. Following this, the single treatment with antagomiR-16 successfully reduced intracellular miR-16 levels to less than 0.1-fold over 7 days, corroborating the ability of the nonviral nHA particle system to achieve over 90% silencing functionality. 26 Subsequently, greater than twofold expression of AcvR2a and Smad5 was demonstrated versus the antagomiR-scr control at the mRNA level, indicating a robust manipulation of the direct gene targets of interest to this study. This is the first time that the interaction between miR-16 and Smad5 or AcvR2a has been assessed at the mRNA level, and this can be interpreted as of high biological relevance considering that the effects of miRNA manipulation are more easily detectable at the protein level than at the mRNA level.56,57 Moreover, it has been previously reported that antagomiR-16 treatment did not affect intracellular miR-133a levels and vice versa. 27 Hence, owing to the specific manipulation of intracellular miRNA levels with this treatment, the efficient increase in mRNA levels of the targets assessed confirmed the ability of nHA-based delivery to mediate the successful genetic manipulation of hMSC osteogenic culture at the posttranscriptional level.
The effects of miR-16 inhibition and upregulation of Smad5 and AcvR2a on downstream hMSC osteogenesis were demonstrated by the markedly increased mRNA levels of Runx2 and OCN (by >6-fold) as well as the enhanced ALP activity. Runx2 and OCN expression is routinely analyzed in osteogenesis studies, together with ALP activity, as they denote the specific commitment of MSCs to the bone-forming lineage. Changes for these genes are generally less than threefold following miRNA delivery with varied delivery vectors, miRNA doses, and cell types.58–61 However, the present study detected increases more in line with the 2.5- to 10-fold range that have been reported following the delivery of BMP2, activin-receptor like kinase (caALK)-6, or Runx2 pDNA using the popular vector polyethylenimine (PEI),62–64 which underlines the high effectivity of the nHA-based antagomiR-16 delivery to inducing robust genetic modifications. Regarding ALP activity, the remarkable ∼11-fold increase following antagomiR-16 treatment compared to untreated cells in osteogenic medium was in contrast with a limited ∼1.22-fold increase over the surprising unspecific effect of the antagomiR-scr control group. Other studies have shown that ALP activity was affected by the treatment with scr miRNAs60,61 and that a substantial (∼30%) overlapping exists with this sequence and >100 human protein-coding genes, which may account for this unexpected effect. 27 Taken together, these data confirmed the premise that the specific inhibition of miR-16 with nHA particle-based delivery can enhance a panel of markers for osteogenesis in hMSCs.
Most importantly from a bone repair perspective, a significant increase in calcium deposition with antagomiR-16 was detected at each of the culture conditions assessed: monolayer and antagomiR-loaded scaffolds. Calcium deposition in the ECM during osteogenic culture is regarded as a definite functional marker of both 2D and 3D osteogenesis assays.52,60,61,65–71 miR-16 inhibition led to ∼3.5-fold enhanced calcium deposition over untreated cells in monolayer in this study, representing a pronounced effect that surpassed the twofold effect reported for other combinations of miRNA and nonviral vectors in monolayer.71,72 In the 3D scaffold setting, a ∼2.4-fold increase in calcium deposition was found over the untreated cells cultured on antagomiR-free scaffolds. Of note, limited in vitro studies are available to interpret the effect on calcium deposition observed with the antagomiR-16-loaded scaffolds; this is due to the extended practice of testing the pretransfected cells in monolayer to directly assess in vivo the effect of combining them with the 3D biomaterials. 9 We have previously combined coll-nHA scaffolds with antagomiR-133a 27 or iterations of BMP2/VEGF pDNA23,24 and reported induced hMSC calcium deposition between 0.8-fold (80%) and ∼2-fold in vitro. Importantly, both systems later proved effective without the addition of external cells in vivo to locally transfect host cells and accelerate bone repair, underlining the high significance of the ∼2.4-fold calcium increase result of this study. In summary, these data demonstrated that miR-16 inhibition using the bone-targeted coll-nHA scaffolds presents significant potential as a novel off-the-shelf strategy for bone tissue engineering.
Importantly, due to the many physiopathological implications of miR-16,28–48 the fine-tuning of this off-the-shelf platform system to deliver either miR-16 enhancers or inhibitors and target other tissues and pathways offers exciting opportunities in the wide arena of tissue engineering; as examples, miR-16 inhibition in combination with HA-free skin-targeted collagen-based scaffolds for promoting angiogenesis may offer an alternative for enhancing wound healing, while the comparison between scaffolds activating or inhibiting miR-16 poses vast potential for in vitro modeling of multiple types of cancer.
Conclusion
Taken together, the results of this study demonstrated for the first time that miR-16 plays an inhibitory role in osteogenesis, which can be harnessed as a therapeutic and generate novel off-the-shelf platforms for improved bone tissue engineering by delivering antagomiR-16 from an efficient bone-targeted system consisting of nHA particles and coll-nHA scaffolds. We identified the inhibitory role of miR-16 in osteogenesis and demonstrated the successful ability of nHA-based antagomiR-16 delivery to hMSCs to mediate miR-16 inhibition and subsequently enhance the levels of the two direct targets of interest Smad5 and AcvR2a. Ultimately, this nonviral delivery induced the robust increase of a panel of osteogenesis markers along with hMSC-mediated mineralization. Most importantly, we demonstrated the potential of the antagomiR-16-loaded scaffolds as off-the-shelf 3D platforms to improve hMSC osteogenesis, indicating that miR-16 inhibition in combination with the nonviral delivery platform described herein holds significant therapeutic potential as a novel bone repair strategy.
Footnotes
Acknowledgments
Work funded by the European Research Council (ERC) Grant Agreement no. 239685 and 665777 and the BioAT Scholarship funded by the Programme for Research in Third Level Institutions Cycle 5 and the European Regional Development Fund under the 2007–2013 Funds Programme. The authors are thankful to Integra Lifesciences Corporation for providing the collagen for scaffold synthesis, Ms. Georgina Shaw and Dr. J. Mary Murphy at REMEDI NUI Galway for kindly providing hMSCs.
Disclosure Statement
No competing financial interests exist.