
Abstract
Conferring antithrombogenicity to tissue-engineered vascular grafts remains a major challenge, especially for urgent bypass grafting that excludes approaches based on expanding autologous endothelial cells (ECs) that requires weeks of cell culture. Adipose-derived stem cells (ASCs) are available from most patients in sufficient number for coronary bypass graft seeding and may be effective as allogeneic cells. We thus compared the adhesion and platelet binding of human ASCs that were shear conditioned with constant and pulsatile shear stress (SS) after seeding the cells on a biologically engineered matrix suitable for arterial grafts. A monolayer of cells was maintained up to 15 dyn/cm2 constant SS and up to 15 dyn/cm2 mean pulsatile SS for 6 days of shear flow. Platelet binding was reduced from 83% to 6% of surface area and nitric oxide production was increased 23-fold with 7.5–15 dyn/cm2 constant SS, but not pulsatile SS, relative to cells cultured statically on the matrix for 6 days. The reduction in platelet binding varied from no reduction to maximum reduction over a constant shear range of ∼2 to 4 dyn/cm2, respectively. Collectively, the study supports the potential use of ASCs to seed the luminal surface of a vascular graft made from this biologically engineered matrix to confer an antithrombogenic surface during the development of an endothelium from the seeded cells or the surrounding blood and tissue.
Introduction
C
Graft selection for CABG is crucial to the success of long-term patency. Both arteries and veins have been used as autologous CAB grafts, primarily internal mammary artery and saphenous vein. The failure of saphenous vein grafts (e.g., about 50% of saphenous vein grafts are patent 10 years after CABG 5 ) requires reoperation but may be limited by the availability of suitable autologous vessel; about 30% of patients do not have suitable vein grafts due to harvested vein for prior vascular procedures. 6 Also, harvesting saphenous grafts requires a second surgical procedure, which may not be suitable for certain patients with medical constraints, and any autologous vessel graft entails second site morbidity.
Tissue-engineered vascular grafts (TEVG) have emerged to overcome many of the major limitations of comorbidities, limited availability of autologous vessel harvest, and frequent thrombogenicity in small-diameter synthetic grafts. The success of TEVG depends on the ability to emulate mechanical properties of the native vessels with an organized extracellular matrix that possesses a confluent and functional endothelium to resist thrombosis in vivo. Because a functional endothelium is the only proven recourse for long-term thrombus prevention, achieving one on the luminal surface of a TEVG is crucial for long-term patency without sustained anticoagulant therapy. Although spontaneous endothelialization of our TEVG occurs in preclinical studies with healthy animals over several months depending on anatomical location,7–11 if it cannot be realized in patients over acceptable time frames, even with treatment of the luminal surface with capture molecules such as SDF-1,12–14 then seeding EC onto the luminal surface of the TEVG is one potential option.
The substantial immunogenicity of EC necessitates an autologous source at present. While blood outgrowth ECs are an attractive autologous source for elective CABG patients who can wait the weeks to months required for their isolation and expansion, 15 a clinically suitable source of primary EC is lacking in the case of urgent CABG (needed within 48 h to 15 days), 16 which comprises between 20% and 30% of CABG cases.17–19
Mesenchymal stem cells (MSCs) have been investigated for the treatment of vascular diseases since they can differentiate into EC.20–22 MSC can be found in human bone marrow, 23 adipose tissue, 24 or umbilical cord blood. 25 They have immunomodulatory capacity and immunosuppressive properties due to their ability to regulate the immune response system through direct cell-to-cell interactions and secretion of soluble factors 26 and have demonstrated efficacy in some studies as allogeneic cells.27,28 Recently, Krawiec et al. 29 fabricated TEVG using xenogeneic MSCs (derived from human adipose tissue from healthy donors) and showed robust and patent grafts lined with von Willebrand factor (vWF)-positive cells in a Lewis rat model after 8 weeks of implantation as well as no overt immune response. 29
Isolating MSC from adipose tissue (termed adipose-derived stem cells or ASCs) is clinically feasible, less invasive compared to isolation of MSC from bone marrow (termed BM-MSC), and robust with respect to advanced patient age and comorbidity21,30 except for diabetes. 31 Adipose tissue harvest also yields much more abundant MSC (1–10%) 32 than bone marrow harvest (0.001–0.01%), 33 which makes ASC a very attractive source for TEVG seeding before implantation if autologous sourcing proves necessary.
To use ASC for TEVG, efficient and reliable strategies to differentiate ASC to EC are needed as well as avoidance of differentiation into smooth muscle cells, osteoblasts, and chondrocytes. It was shown that chemical factors, such as vascular endothelial growth factor (VEGF), stimulated ASC to express EC markers. 21 Potent but less explored mediators of ASC differentiation are shear stress (SS) and cyclic stretching, which are relevant for CABG because of immediate exposure of the seeded cells to pulsatile blood flow. Constant SS alone has been reported to induce EC differentiation of ASC 21 as well as in combination with VEGF, 34 consistent with similar studies using BM-MSC. 35 However, no previous studies have assessed the effects of pulsatile SS on ASC differentiation nor has there been a direct demonstration of functional antithrombogenicity—as measured by reduced platelet binding in vitro—by shear-conditioned ASC. Pulsatile SS is of particular interest since this would be the shear regimen experienced immediately postimplantation of an ASC-seeded TEVG in the arterial system.
In this study, we compared the adhesion and platelet binding of human ASC shear-conditioned with constant and pulsatile SS after seeding the cells on a biologically engineered matrix that we previously used in preclinical studies with 36 and without 7 preseeded ECs. In addition, the SS range that yields reduced platelet binding was determined using a parallel plate flow chamber (PPFC) based on Usami et al., 37 wherein SS decreases linearly with axial position along the chamber. We also assessed nitric oxide (NO) production, vascular endothelial (VE)-cadherin, and surface expression of heparan sulfate proteoglycan (HSPG2) as a possible explanation for reduction in platelet binding of the shear-conditioned ASC.
Materials and Methods
Isolation and culture of ASC
Human ASCs (Lonza) were plated at 5000 cells/cm2 and cultured in expansion medium (ADSC-GM, Lonza), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C, 5% CO2. Medium was replaced every 3 to 4 days. After ASCs reached near confluency (∼90%), cells were passaged for subsequent experiments and used at passage 5.
TEVG preparation
TEVG from fibroblast-remodeled fibrin gel was prepared using our established protocols. 38 Thrombin (1.1 U/mL) and calcium chloride (5.0 mM) were added to fibrinogen (4 mg/mL) solution with suspended human dermal fibroblasts (Clonetics) in 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-buffered saline. After gelation, the samples were removed from the tubular mold and cultured on the 4 mm diameter glass mandrel in Dulbecco's modification of Eagle's medium (DMEM; Cellgro) containing 10% fetal bovine serum (FBS; EMD Millipore), 100 U/mL penicillin, 100 μg/mL streptomycin, 2 μg/mL insulin, and 50 μg/mL ascorbic acid. Medium was changed three times per week for 2 to 3 weeks to allow for initial gel compaction and associated circumferential alignment of the cell-produced matrix. Following this static culture, the grafts were mounted in pulsed flow-stretch bioreactors at 0.5 Hz pulse frequency and constant strain amplitude circumferential strain in the range 3–7% for 5–7 weeks. 38
After bioreactor maturation of the grafts, they were decellularized as previously described. 8 The constructs were rinsed with phosphate-buffered saline (PBS) for 10 min at room temperature with gentle rocking, followed by 1% sodium dodecyl sulfate (SDS; Sigma) for 6 h. Subsequently, they were rinsed with 1% Triton X-100 (Sigma) for 30 min, followed by 1 week of PBS rinses. A final treatment with deoxyribonuclease enzyme (DNase; Worthington Biochemical) in DMEM/10% FBS at 37°C, 5% CO2 for overnight incubation was used to maximize removal of residual DNA. Grafts were then washed with PBS for 3 to 4 days to continue removing any residual SDS/Triton/DNAase/cellular components and stored in PBS at 4°C until use.
SS conditioning
The decellularized grafts were cut longitudinally into 3 × 0.5 cm rectangles and embedded in 3% agarose in polycarbonate flow chamber inserts to expose the luminal surface for ASC seeding.
39
The biologically engineered matrix was pretreated with 50 μg/mL fibronectin (Sigma, F1141), and then, ASCs were seeded as a monolayer from a suspension (1.5 × 106 cells/mL) onto the mounted matrix and incubated for 24 h. After the static culture period, a subset of the samples was subjected to targeted SS for 6 days in our custom PPFCs with an inlet width of 1 cm, length of 3 cm, and height of 0.05 cm. The varying width of the chamber was adapted from Usami et al.'s study
37
to apply a linear range of SS. Endothelial growth medium (EGM-2; Lonza) was used as a source of endothelial differentiation factors, including VEGF, human basic fibroblast growth factor, and human epithelial growth factor. A pulse dampener was placed right after the peristaltic pump to create constant SS or removed to create pulsatile SS at 0.5 Hz. The Womersley number (Wo) is a dimensionless number measuring the ratio of inertial forces to viscous forces.
where h = chamber gap height,
Platelet adhesion assay
Human platelets (Red Cross) suspended in platelet-rich plasma were centrifuged at 1300 g for 10 min to yield the platelet pellets. Pellets were suspended in “acid citrate dextrose” buffer (100 mM trisodium citrate, 10 mM citric acid, and 136 mM glucose, pH 6.5). The platelets were centrifuged again at 1300 g for 10 min and suspended in Tyrode's solution (134 mM sodium chloride, 12 mM sodium bicarbonate, 2.9 mM potassium chloride, 0.34 mM sodium phosphate monobasic, 1 mM magnesium chloride, 10 mM HEPES, and 5.5 mM glucose, pH 7.4). Samples were incubated in suspended platelets for 2 h at 37°C. After incubation, they were rinsed with Tyrode's solution two to three times. The samples were fixed with 1% paraformaldehyde for 30 min at 4°C followed by 3 × 5 min PBS rinses.
Immunostaining
The samples were permeabilized by 0.1% Triton X-100 (Sigma) and blocked with 5% normal donkey serum at room temperature. They were incubated with primary antibodies CD41 for glycoprotein IIB/IIIA (Abcam, ab11024, 1:400), HSPG2 (Abcam, ab26265, 1:100), and VE-cadherin (Abcam, ab33168, 1:200) overnight at 4°C. Species-matched secondary (Cy3- or Cy5-conjugated, 1:200) antibody (Jackson Immunoresearch) and Alexa Fluor 488 phalloidin (Life Technologies, 1:400) were added to the samples. Cell nuclei were stained with Hoechst 33342 (Invitrogen, H3570, 1:10,000).
Nitric oxide assay
Medium samples were collected from each of the flow conditions and frozen at −80°C. Using a Fluorometric Assay Kit (Abcam, ab65327), nitrite in the samples was converted to nitrate during 3-h incubation. Enhancer, DAN probe, and sodium hydroxide were added to the converted nitrate and was measured at 360/450 nm (excitation/emission).
Alignment of ASC
Cell alignment on the matrix was evaluated postflow by phalloidin staining and analyzed using an ImageJ orientation plugin 40 to determine the orientation angle relative to the flow (SS) direction.
Statistical analysis
Results are expressed as mean ± standard deviation with at least three repeated experiments (indicated by “n”). Statistical significances were evaluated by one-way analysis of variance for multiple groups or one sample t-test (for cell orientation), compared to a theoretical value of 1, with a probability value of p < 0.05 considered statistically significant.
Results
Linear SS parallel plate flow chamber design validation
Assessing different levels of uniform SS for endothelial differentiation of ASC in vitro in a conventional PPFC would be time consuming and require many samples to be run in separate experiments. The PPFC of Usami et al., 37 wherein SS varies linearly with axial position in a prescribed region, was modified to study the effects of selected ranges of constant laminar SS on ASC preseeded on biologically engineered matrix embedded in agarose within an insert that mounts flush with the flow surface. 39 Figure 1A shows a top view of the chamber and the contour of the sidewalls needed to achieve a region of linear dependence of SS (a conventional PPFC uses sidewalls that have a constant separate distance along the chamber axis). The resulting SS profile along the chamber axis for constant laminar flow from left to right for a fixed volumetric flow rate computed using COMSOL Multiphysics is shown in Figure 1B. The SS is highest near the inlet and then decreases linearly with distance from 15 to 7.5 dyn/cm2 over a 3 cm length in the central region (where a rectangular sample is located) until it becomes nonlinear with distance toward the outlet. The decrease in SS by a factor of two occurs for any chosen inlet flow rate given the chamber design, a feature that was also used in this study. As shown in Supplementary Figure S1 (Supplementary Data are available online at www.liebertpub.com/tea), at any axial position in the boxed region, the predicted SS is uniform over ∼90% of the width, and only cells within that domain were assessed.
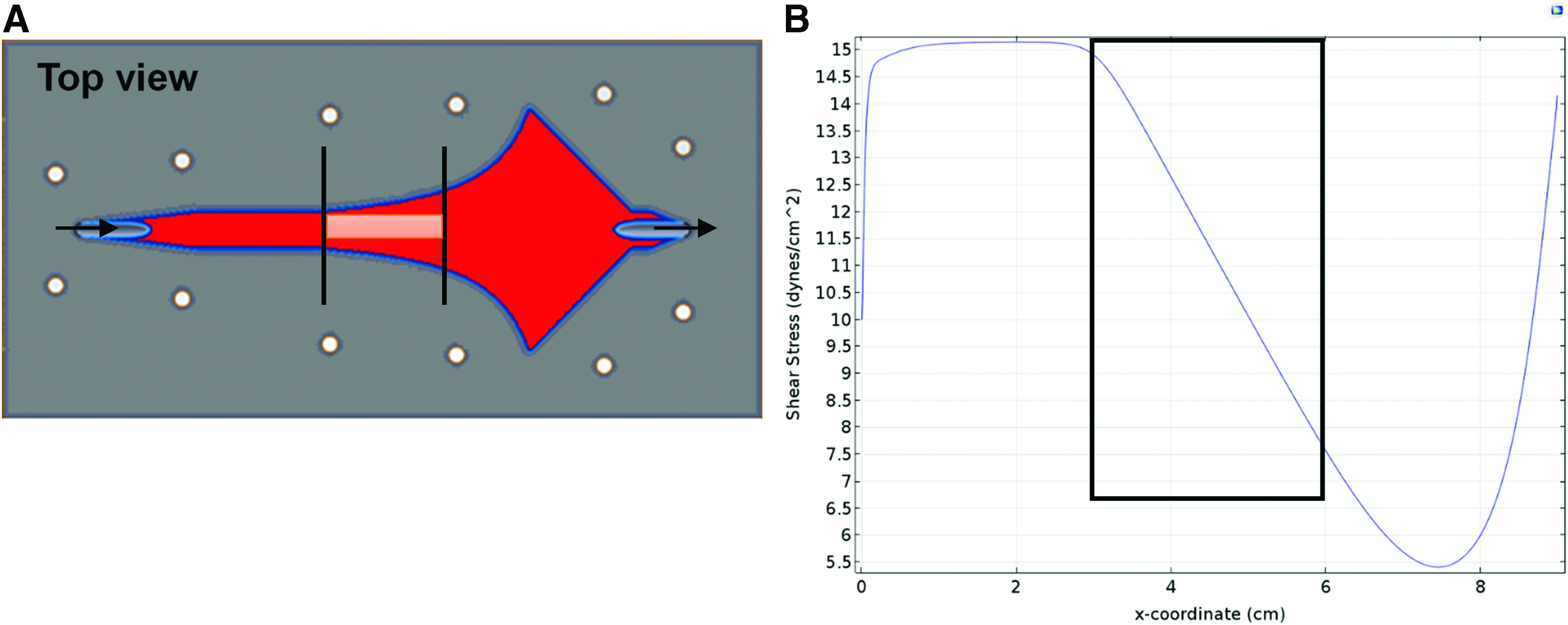
Top view
ASC retention and platelet binding post-shear conditioning
ASCs seeded onto slabs of the biologically engineered matrix were mounted flush onto one plate of the PPFC, cultured statically for 1 day, and then subjected to constant or pulsatile SS in a ramp over 2 days to a target value and held for 4 days. Samples were harvested and assayed for retention via staining with Hoechst and for platelet binding via immunostaining for glycoprotein IIB/IIIA. Figure 2A shows that the unseeded matrix led to extensive platelet binding, as expected, given it is composed of extracellular matrix (Supplementary Fig. S2), including collagen, fibronectin, and remnant fibrin,7,36 known to bind platelets. It was found that platelet binding was reduced following ASC for static culture the entire 7 days (Fig. 2B), but the reduction was greater for ASC cultured for 6 days under constant SS for the range 7.5–15 dyn/cm2, during which time a monolayer of ASC was maintained (Fig. 2C, D). There was no dependence on the magnitude of constant SS over the range 7.5–15 dyn/cm2 (Supplementary Fig. S3), over which a monolayer of ASC was maintained (Supplementary Fig. S4), and there was no difference in the reduction using constant and pulsatile SS of mean 15 dyn/cm2 (Fig. 2E; see Supplementary Fig. S5 for pulsatile waveform). Interestingly, there was a graded reduction in platelet binding over the range 2 to 4 dyn/cm2 of constant SS (Fig. 3A) with a difference across this range (Fig. 3B), over which a monolayer of ASC was again maintained (Fig. 3C). The platelet binding measured at 2 dyn/cm2 was approximately the value measured for static incubation (Fig. 2E) and the value measured at 4 dyn/cm2 approached that measured in the range 7.5–15 dyn/cm2 (Supplementary Fig. S3), indicating 4 dyn/cm2 of ASC shear conditioning to be an approximate threshold for maximal reduction of platelet binding to the ASC.
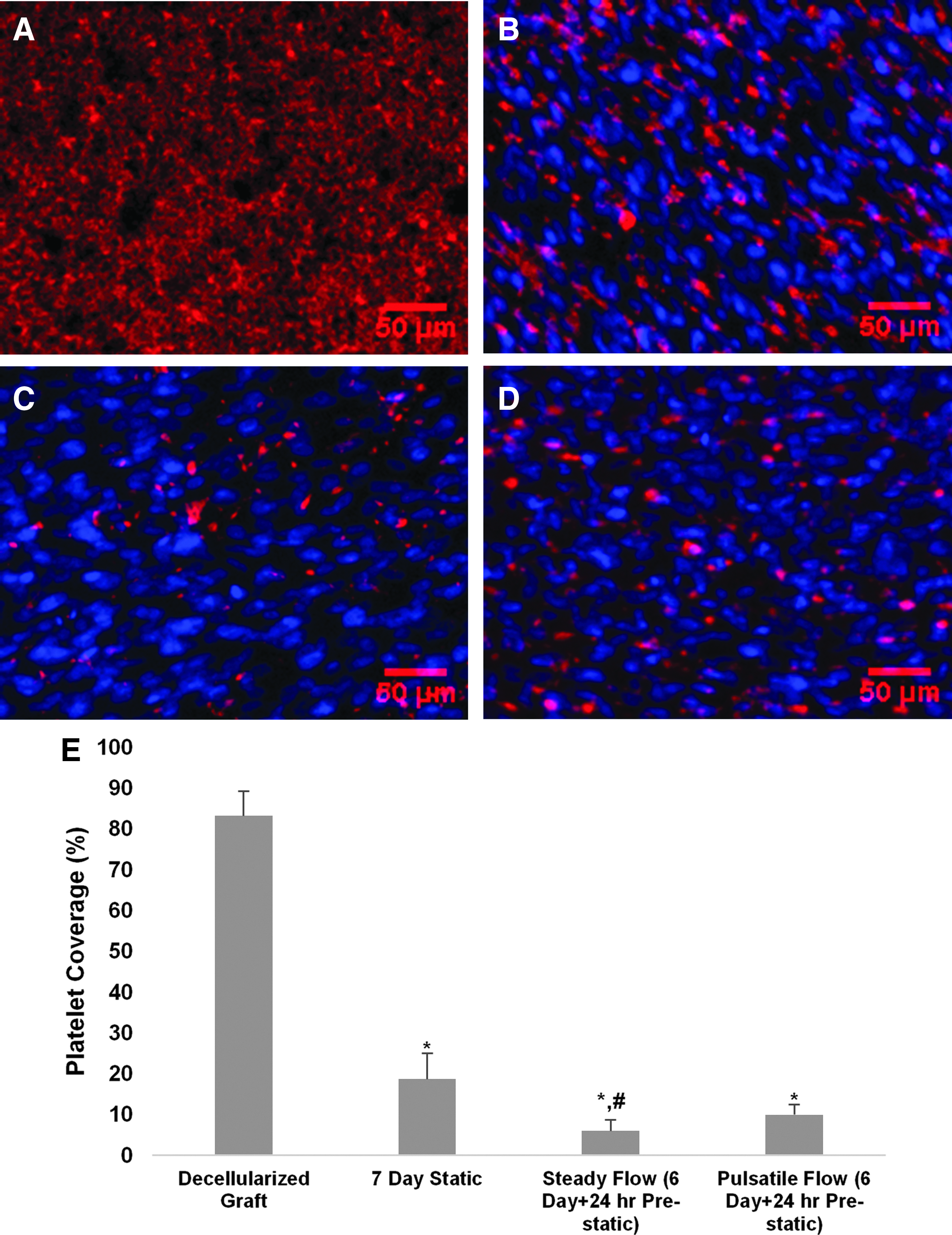
Reduced platelet binding of TEVG seeded with shear conditioned ASC.
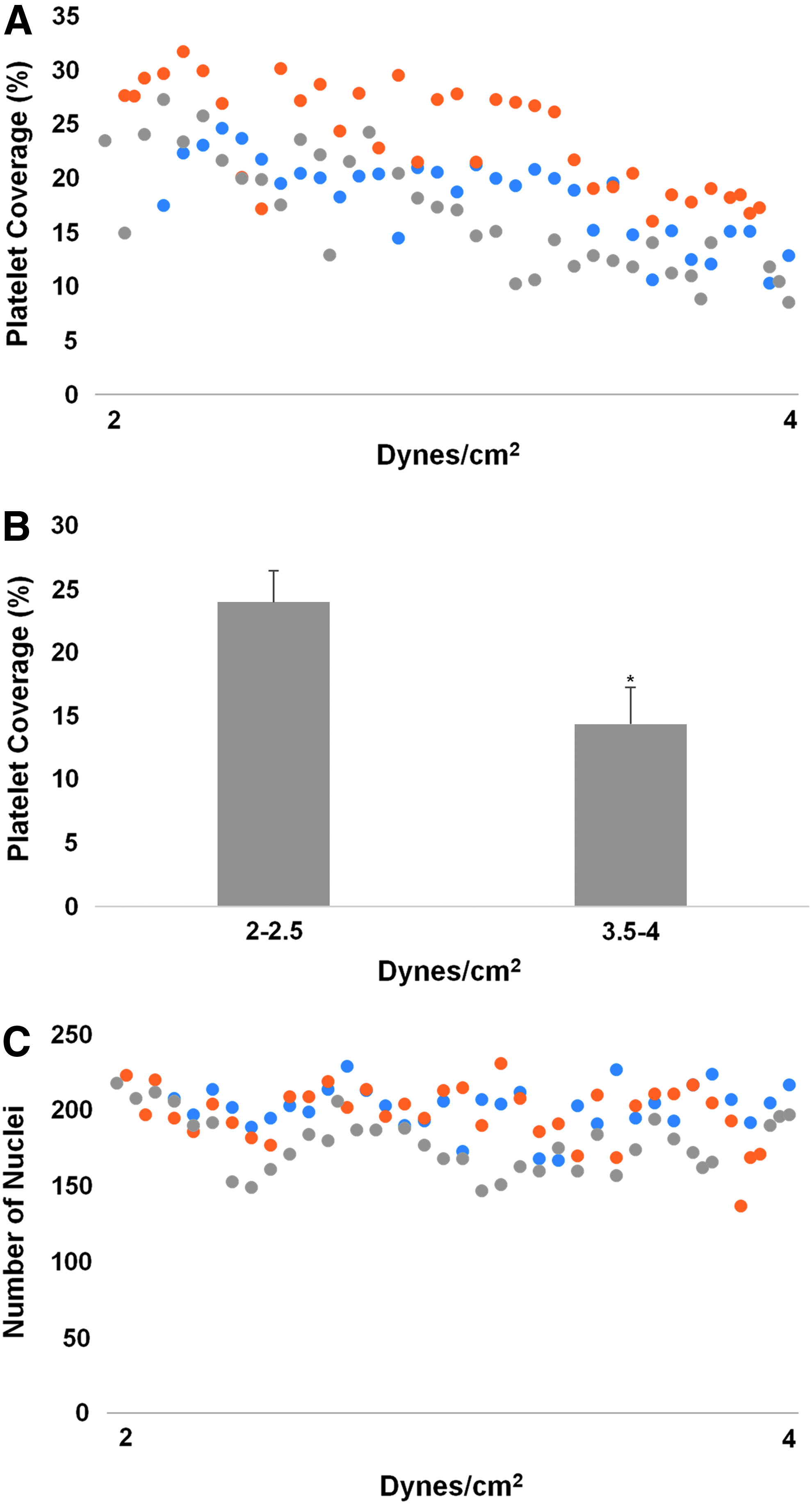
Platelet binding at low constant SS.
ASC production of NO post-shear conditioning
NO production is a hallmark of the endothelium, and NO is well known to be a platelet inhibitor (as well as vasodilator), so its production in these experiments was measured. Medium collected from the chambers for the same three ASC-seeding groups presented in Figure 2E was analyzed for NO via conversion of its natural degradation product nitrite to nitrate. Figure 4 shows that there was dramatic 23-fold increase of NO produced in the cases of constant shear conditioning relative to static culture for the entire 7-day incubation. Interestingly, there was also an NO production increase resulting from constant shear relative to pulsatile shear. There was no apparent upregulation of HSPG2 or VE-cadherin resulting from shear conditioning based on immunostaining (Supplementary Fig. S6).
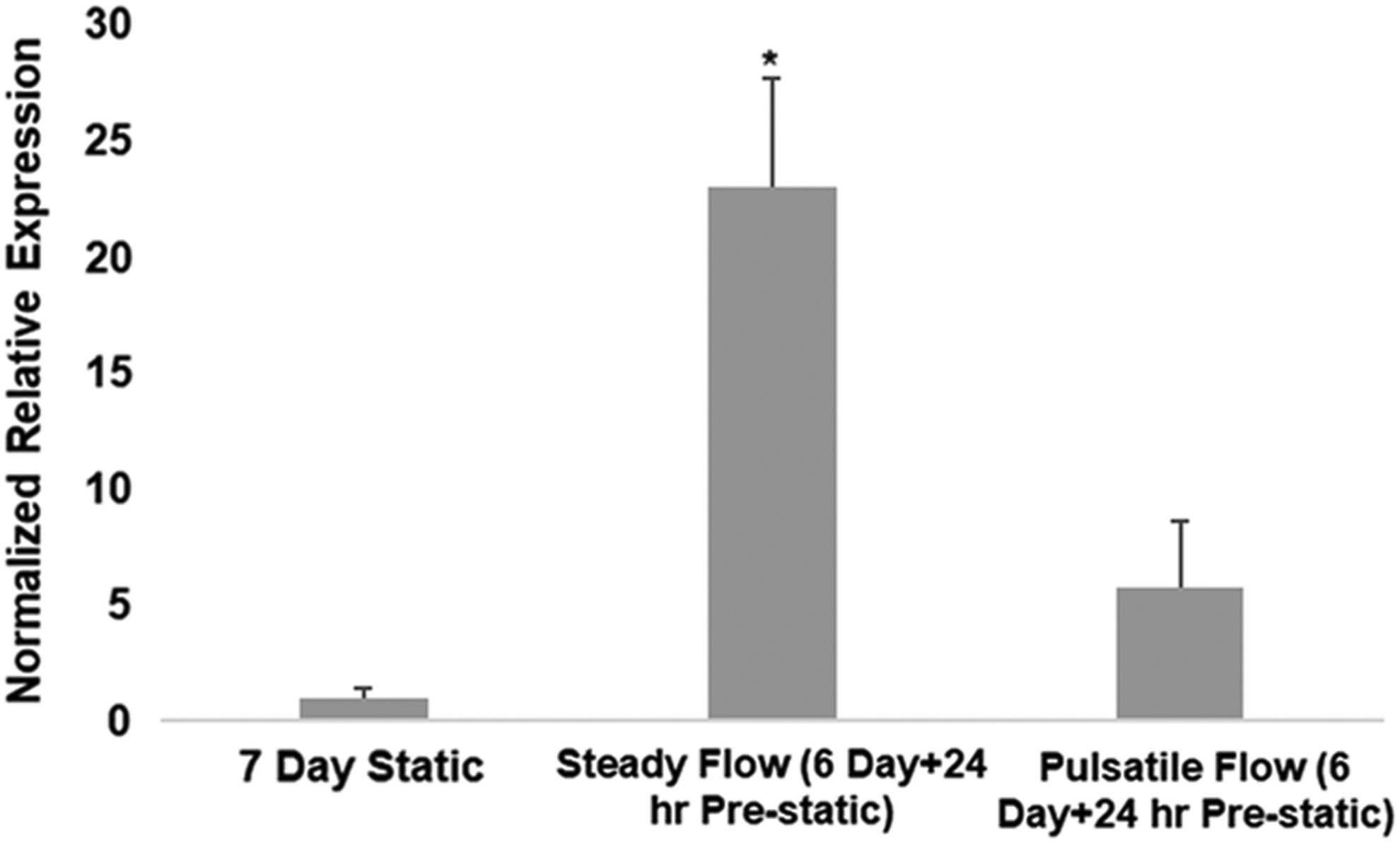
Nitric oxide production of shear conditioned ASC. *p < 0.001 vs. 7 day static and pulsatile flow (n = 4).
ASC orientation post-shear conditioning
ASC orientation was measured as <cosθ> from F-actin-stained stress fibers for the same ASC-seeding groups presented in Figure 2E is shown in Figure 5. Only the static culture group had a value of <cosθ> different from the flow direction, with both constant and pulsatile shear conditioning leading to ASC alignment with the flow direction (<cosθ> =1). The value of <cosθ> for the static culture group was consistent with random orientation (<cosθ> = 0.5).
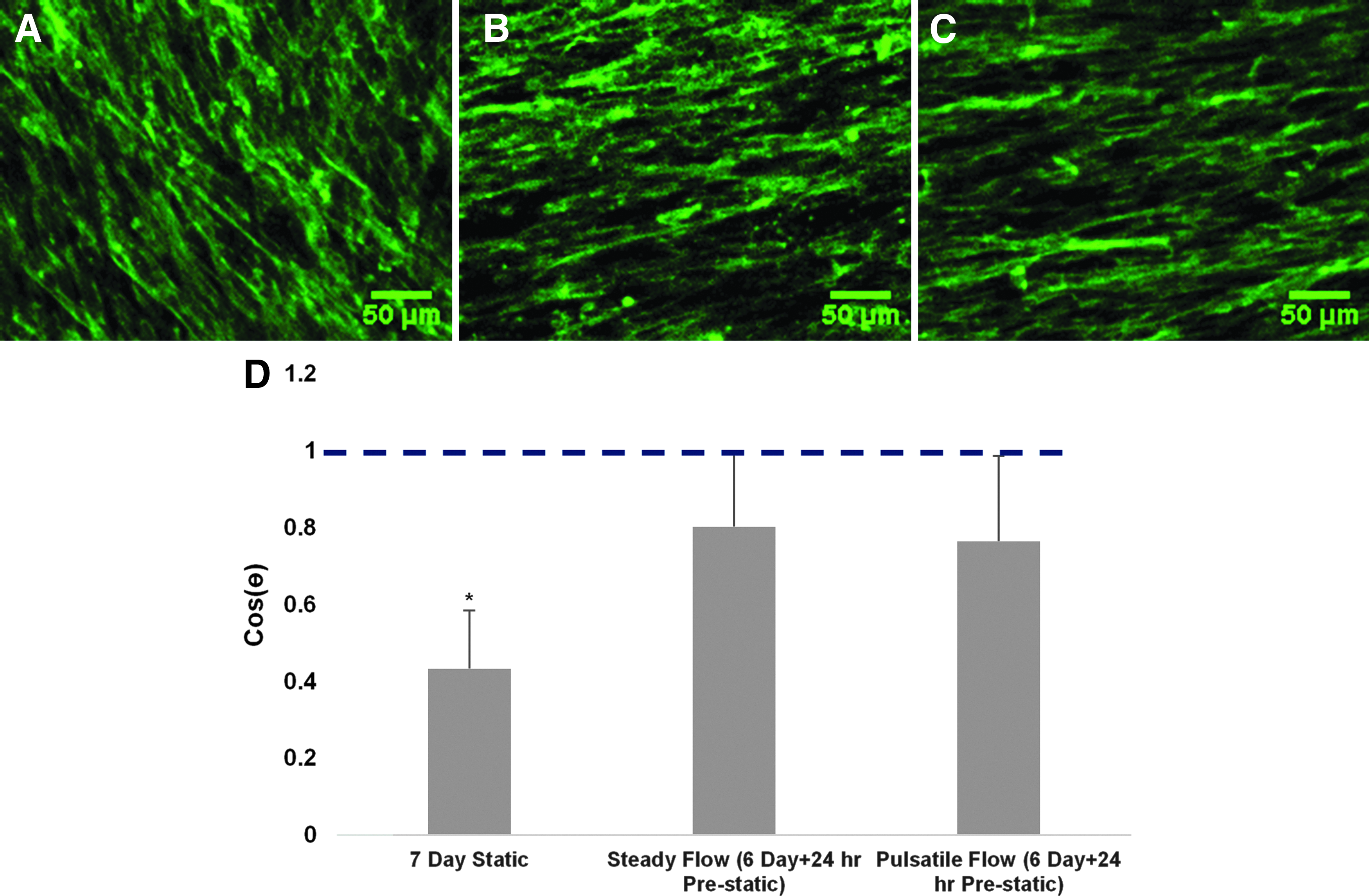
Alignment (F-actin) of ASC in response to SS.
Discussion
The motivation for this study was to ascertain whether short-term shear flow conditioning in vitro could be used to drive ASC differentiation toward an antithrombogenic phenotype postseeding on biologically engineered matrix as a strategy to confer rapid hemocompatibility for urgent CABG applications, using tubes of this matrix as a TEVG. Previous studies have indicated that shear conditioning of ASC on various synthetic substrata has resulted in the expression of endothelial markers and function consistent with increased antithrombogenicity, namely NO production. 21 However, a more definitive assessment of antithrombogenicity has not been reported, and no assessment of ASC on a biologically engineered matrix has been reported. Our TEVG—a decellularized tissue tube grown in vitro from dermal fibroblasts in a sacrificial fibrin scaffold—has been shown to yield physiological burst pressure and stiffness7,38 and confer excellent retention of ECs both in vitro and in vivo.36,39 It also endothelialized spontaneously over time in preclinical studies using healthy animals.7–9 It was thus considered a relevant matrix to use for these experiments performed to assess whether short-term shear flow conditioning could result in decreased platelet binding to the seeded ASC as well as increased NO production, and whether any such effects depended on the shear flow being constant or pulsatile.
The linear shear flow PPFC conceived by Usami et al. 37 was very advantageous for these studies. Not only did it allow for assessment of how ASC retention on the matrix varied with SS without repeated experimentation as would be required using a conventional constant shear flow PPFC but it also allowed for determination of a threshold constant SS, above which a further reduction in platelet binding did not result. A previous study 41 experimentally validated the linear shear flow predicted by Usami et al. 37 and used computational fluid dynamics to demonstrate the quasi-steady solution (where the surface SS is given by the steady-state solution at the instantaneous flow rate, to good approximation) for sufficiently small (<1) Womersley number, in their case 0.75 and in our case 0.35.
The reduction in platelet binding over the range 2 to 4 dyn/cm2 cannot be attributed to loss of an ASC monolayer and exposure of the underlying matrix, as a monolayer was observed with Hoechst staining, and indicates a differentiation toward an antithrombogenic phenotype. No further reduction of platelet binding was observed beyond a constant SS of ∼4 dyn/cm2, indicating a threshold for differentiation. Surprisingly, there was no benefit of pulsatile shear conditioning over constant shear comparable to the time-averaged value, at least for the waveform used. These observations provide some gross clues to the biophysical mechanism of shear mechanotransduction. 42 For example, if nuclear deformation determines the ASC differentiation leading to the observed platelet binding inhibition, these results suggest that either the deformations resulting from the steady and pulsatile waveform used are no different, or, more likely, the resulting differences are inconsequential. Further clues might be obtained from testing other frequencies and amplitudes that can be related to dynamics of second messenger dynamics and cytoskeletal/nuclear strains, respectively.
Low-magnitude laminar SS (e.g., 0.3–2.7 dyn/cm2) has been shown to induce BM-MSC/ASC toward osteogenic cells rather than EC,43–45 whereas laminar arterial SS (e.g., 15 or 20 dyn/cm2) has been reported to increase messenger RNA (mRNA) and protein expression of endothelial markers in BM-MSC.46,47 These reports are consistent with our finding that the reduction in platelet binding became more pronounced as laminar SS increased to 4 dyn/cm2 and was independent of SS magnitude over the range of 7.5–15 dyn/cm2. The relatively low value of the threshold in our system for apparent endothelial differentiation (i.e., 4 vs. 15 or 20 dyn/cm2) may reflect a difference in the outcome measure used (platelet binding reduction vs. EC marker expression), environmental conditions (biological matrix vs. synthetic substrate), or MSC type (ASC vs. BM-MSC).
Endothelial NO is well known to inhibit platelet activation, adhesion, and aggregation, and a deficiency of bioactive NO is associated with arterial thrombosis. 48 The marked 23-fold increase in NO produced by the ASC after constant shear conditioning relative to static culture may explain the observed reduction in platelet binding and is consistent with ASC differentiation toward an EC phenotype, although there are other endothelial mechanisms platelet binding inhibition that might also contribute to this reduction involving, for example, prostacyclin and HSPG2. The 23-fold increase we observed was substantially greater than the 3.6-fold increase reported by Zhang et al., 21 but that can be attributed to any number of differences in the study conditions, including shear duration (2 vs. 6 days), SS platform (orbital vs. laminar), and substrate (decellularized human saphenous vein precoated with fibronectin vs. biologically engineered matrix precoated with fibronectin).
While elevated NO production is consistent with endothelial differentiation, we did not detect immunostaining for VE-cadherin and HSPG2, which was shown indirectly using heparinase to be the molecular mechanism leading to a reduction of platelet binding in shear-conditioned BM-MSC isolated from bone marrow. 49 Thus, despite the increased NO production and reduced platelet binding, whether the shear-conditioned ASCs were differentiating toward an endothelial phenotype in this study is an open question. Although Zhang et al. 21 demonstrated that SS increased NO production and upregulated mRNA levels of both VE-cadherin and vWF in ASC, Bassaneze et al. 50 did not detect upregulation of any endothelial mRNA levels (e.g., vWF) in ASC after exposure to SS of 10 dyn/cm2 for 96 h, even though NO production was increased. These different responses of ASC to SS exposure could be due to different substrates and different magnitude and duration of SS.
Cell alignment with flow under constant and pulsatile shear flow in the normal arterial range is a hallmark of ECs seeded on a variety of substrates and has also been reported for ASC after 2 days of constant shear conditioning. 21 Our observation of ASC alignment with flow after 6 days of both constant and pulsatile shear conditioning is consistent with these results and with ASC differentiation toward an EC phenotype.
Any number of substrate properties can influence cell differentiation, including chemical composition, 51 mechanical stiffness, 52 and topography. 53 While our biologically engineered matrix resembles vascular wall tissue in all three respects, it is not a chemically defined synthetic matrix and its composition, being cell-produced, is complex. In a previous study seeding ECs onto the lumen of similarly prepared TEVG, we observed faint luminal staining for fibronectin 36 that may play an important role in ASC adhesion and mechanotransduction, even though the bulk matrix is predominantly collagen.
The similarities and differences of BM-MSC and ASC have been reviewed 54 and remained the subject of investigation, but this study further confirms the potential of the more abundant ASCs, with appropriate preconditioning, as a clinically attractive source of autologous cells that can be used to confer hemocompatibility to TEVG and other implants into the cardiovascular system. A central question that remains, however, is the extent of preconditioning necessary to ensure further EC differentiation, or at least maintenance of antithrombogenicity, postimplantation.
Footnotes
Acknowledgments
The authors gratefully acknowledge technical assistance provided by Zeeshan Syedain, Lee Meier, Susan Saunders, Sandy Johnson, and Naomi Ferguson, and funding provided by Regenerative Medicine Minnesota Award No. 2015 1872 (to R.T.T.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.