
Abstract
Osteoporosis, which is characterized by low bone mineral density and susceptibility to fracture, is caused by increased osteoclastic activity. Receptor activator of nuclear factor kappa B ligand (RANKL)/RANK signaling plays an important role in osteoclast differentiation and activation. The current treatment strategies for osteoporosis do not directly address this underlying cause and generates undesired side effects. This led to emergence of controlled delivery systems to increase drug bioavailability and efficacy specifically at the bone tissue. With better understanding of molecular pathology of bone, the use of small interfering RNA (siRNA) to inhibit translation of abnormal gene expression in cells is becoming a promising approach. In this study, we report a siRNA delivery system consisting of PEI:RANK siRNA complex entrapped in nanosized poly(lactic acid-co-glycolic acid) (PLGA) capsules intended to be used in the treatment of osteoporosis. The nanosize will enable the nanoparticles to be administered by intravenous injection. The RANK siRNA was complexed with polyethylenimine (PEI) and loaded into biodegradable PLGA nanocapsules (NCs). The PEI:RANK siRNA loaded nanocapsules significantly reduced (47%) RANK mRNA levels. The differentiation of osteoclast precursors to mature osteoclasts was significantly suppressed (∼54%). The reduction in the osteoclastic activity of the differentiated osteoclasts (55%) was found to be statistically significant. The siRNA delivery system developed in the study is planned to be tested i.v. in mouse and has the potential to be used as a novel alternative approach for the systemic treatment of osteoporosis.
Introduction
Bone tissue is responsible for the structural support and protection of other organs in the body. It produces blood cells and stores minerals. Disorders of bone, such as cancer, osteoarthritis, or osteoporosis, are therefore very important, and their effective treatments remain a challenge. 1 Osteoporosis is a skeletal disorder associated with increased osteoclast mediated bone resorption 2 resulting in reduced bone mineral density and structural failure of the bone. 3 The disease affects ∼200 million people worldwide and it is estimated that osteoporotic hip fractures will exceed 21 million by 2050. 4 The currently used drug types for the treatment of the disease are bisphosphonates, parathyroid hormone, or estrogen. However, they have limitations, including low bioavailability due to their short half-life and long-term safety concerns, 5 and it is known that the therapeutic efficacy of a drug depends on its specificity for the target and bioavailability at the target site. 6
It is proven that the high bone resorption rate in osteoporosis is due to the increased number and/or activity of osteoclasts. 2 Receptor activator of nuclear factor kappa B (RANK)/RANK ligand (RANKL) system is an important pathway involved in osteoclastogenesis. 7 RANKL which is expressed on osteoblast/stromal cells binds to its receptor RANK on osteoclast precursors. 8 This binding causes recruitment of tumor necrosis factor receptor associated Factor 6 (TRAF6) and activation of NFκB and mitogen activated protein kinase pathways and transcription factors such as c-Fos. These eventually lead to activation of NFATc1, the major transcription factor of osteoclastogenesis. 9 It regulates the expression of osteoclast specific genes such as tartrate resistant acid phosphatase (TRAP), Cathepsin K, and calcitonin receptor,9,10 and therefore, osteoclastic differentiation is initiated (Fig. 1). The RANK/RANKL pathway is under the control of osteoprotegerin (OPG), which is expressed by the osteoblastic cells. OPG acts as a decoy receptor by binding to RANKL and, thus, prevents the interaction of RANKL with RANK. 11 The inhibition of RANK/RANKL system in osteoclastogenesis is a promising approach to be used as a strategy for treatment of osteoporosis since it would eventually lead to reduced osteoclast function and therefore bone resorption.
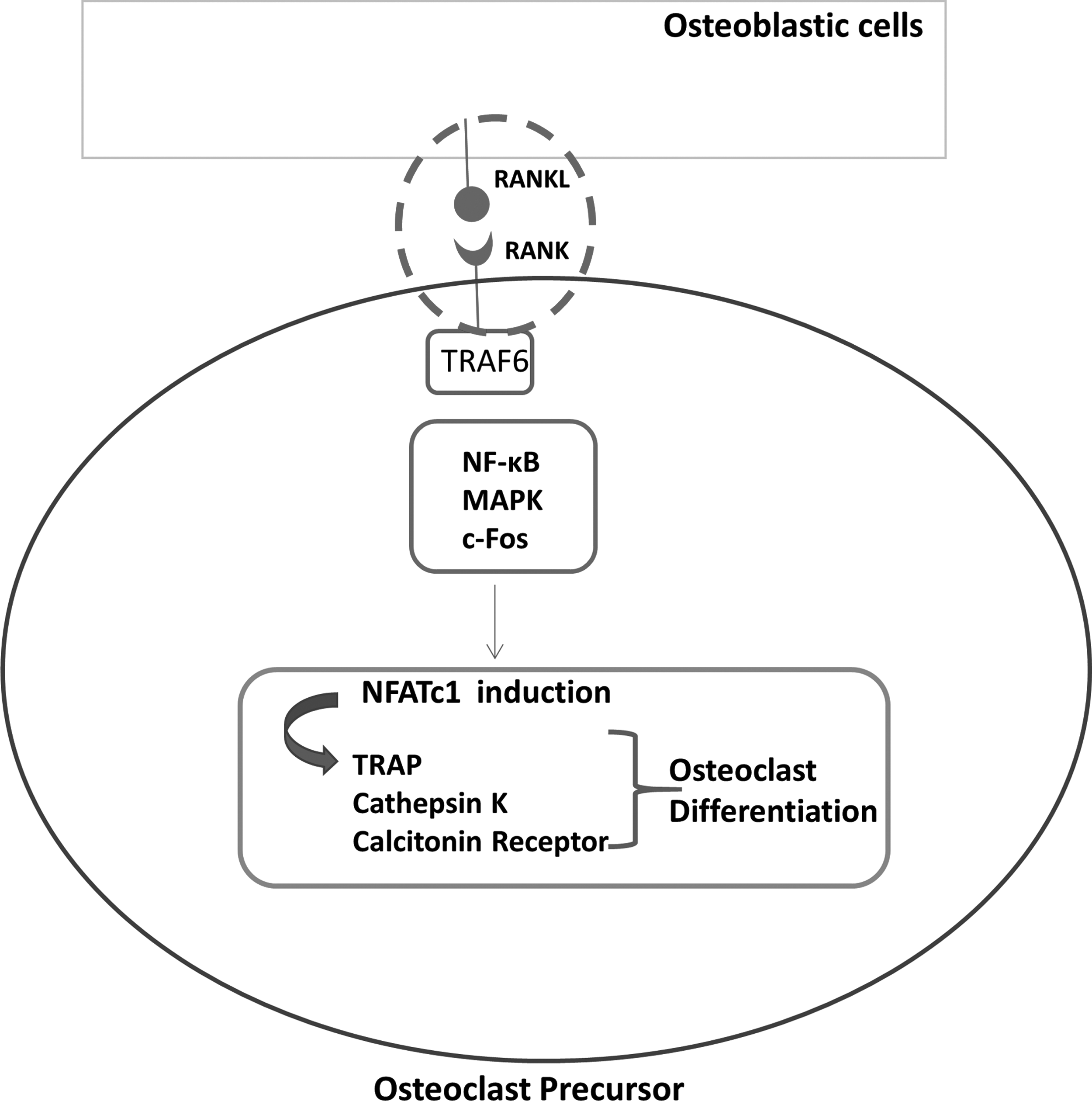
Schematic representation of intracellular signals for RANKL induced osteoclastic differentiation. MAPK, mitogen activated protein kinase; RANK, receptor activator of NFκB (nuclear factor kappa B); RANKL, receptor activator of nuclear factor kappa B ligand; TRAF, tumor necrosis factor receptor associated factor; TRAP, tartrate resistant acid phosphatase.
In recent years, with increasing understanding of the molecular background of bone pathology, new molecular targets are being tested in the search for a treatment of osteoporosis. Excessive expression of the identified molecular targets can be suppressed using RNA interference (RNAi) process. RNAi is a natural process used by the cells to suppress the production of a protein either by mRNA degradation or translation inhibition. 12 Delivery of small interfering RNA (siRNA) into the cells mimics this natural process and is used in silencing the expression of disease-causing genes. Once introduced into the cells, siRNA binds to a protein complex called RNA-induced silencing complex (RISC). With the unwinding of siRNA in RISC, the guide strand is targeted and bound to the specific mRNA with full complementarity. This binding results in the activation of the RNase H activity of the RISC and leads to degradation of the target mRNA. 13
Effective delivery of siRNA to desired tissues in vivo is challenging due to its high molecular weight, negative charge, and instability. It is rapidly degraded by nucleases in serum and excreted from the circulation by reticuloendothelial system. 14 The degradation and excretion cause lower amounts of siRNA to reach to the target cell. In addition, the efficacy of the siRNA that has reached the desired cell is further reduced by the lysosomal degradation in the cell. 15 As a result of these drawbacks, carrier systems such as liposomes and polymeric or inorganic nanoparticulate systems are being developed to protect the siRNA and increase its concentration at the desired tissues and cells. 16 There are several in vitro approaches concerning the use of siRNA as a therapeutic agent for the treatment of osteoporosis. These include the use of commercial transfection reagents17,18 and other nonviral gene carriers such as polyethyleneimine (PEI). 19 Local delivery of siRNA to the bone tissue has also been studied using scaffolds 20 or calcium phosphate cements. 21 Targeting and systemic delivery of siRNA to bone involves carriers such as liposomes22,23 or polymeric micro- or nanoparticulate systems.24,25 In the present study, we have developed a nanosized siRNA delivery system to be used in systemic treatment of osteoporosis by inhibiting the catabolic action of osteoclasts.
In our previous work, 26 we developed a nanocapsule-based siRNA delivery system, which was targeted to bone by the use of an osteotropic sequence carried within a synthetic polypeptide, elastin-like recombinamer (ELR). 27 The DNAoligo was used as a substitute for siRNA after complexation with PEI and was loaded in poly(lactic acid-co-glycolic acid) (PLGA) nanocapsules. The encapsulation efficiency of the complex in PLGA nanocapsules was 48%. The size of this system (197 ± 76 nm) was in the range of discontinuous endothelium associated with poorly structured basal membrane fenestrations in the bone marrow, increasing the potential of nanoparticles for reaching to the bone.
In the present follow-up work, we studied the efficacy of the delivery system in murine macrophage osteoclast precursor cell line, RAW264.7. We loaded the complex, PEI:RANK siRNA, into the PLGA nanocapsules and studied the internalization and toxicity of the nanocapsules and assessed the efficacy of the delivery system in inhibition of RANK mRNA levels and osteoclast differentiation. The effect of the drug delivery system on the inhibition of osteoclastic activity was also studied by pit formation assay.
Materials and Methods
Cell culture
Murine-macrophage osteoclast precursor cell line RAW264.7 was purchased from American Type Culture Collection (ATCC) and maintained in Dulbecco's modified Eagle's medium (DMEM) high glucose (Biological Industries) supplemented with 10% fetal bovine serum (Biological Industries) and 1% penicillin–streptomycin (Sigma, Germany) at 37°C in a humidified atmosphere containing 5% CO2. For the differentiation of RAW264.7 cells to osteoclasts, RAW cells were treated with 100 ng/mL RANKL (Peprotech) and 20 ng/mL M-CSF (R&D Systems) for 5 days with a medium change on day 3.
Inhibition of RANK mRNA by complexes with varying N/P ratios
The efficacy of two RANK siRNA sequences (Table 1) was studied using the HiPerFect Transfection Reagent (Qiagen, Germany). Based on the results of the study, RANK siRNA#1 was more effective than RANK siRNA#2 and, therefore, was chosen to be used in the further experiments (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
Receptor Activator of Nuclear Factor Kappa B Small Interfering RNA Sequences Used in the Study
RANK, receptor activator of nuclear factor kappa B; siRNA, small interfering RNA.
To determine the effective N/P ratio of PEI:RANK siRNA complexes that would lead to sufficient RANK mRNA silencing and were minimally toxic to cells, we tested different N/P ratios (6, 20, 25, 30, and 60) by quantitative real-time PCR (qRT-PCR) using a known amount of siRNA (375 ng/well) in 12-well culture plates. The amount of siRNA to be used was decided based on the transfection study performed by HiPerFect Transfection Reagent (Supplementary Methods). The complexes were diluted in Opti-MEM (200 μL; Thermo Fisher Scientific) and added to RAW264.7 cell suspension (1.2 × 105 cells/well). After 6 h, growth medium was added (800 μL), and gene silencing was monitored at 24, 48, and 72 h post-transfection.
Preparation of PEI: RANK siRNA loaded PLGA nanocapsules
PLGA nanocapsules were prepared by water-oil-water (w/o/w) double emulsion method as previously described with minor modifications. 28 Briefly, PLGA (50:50) (50 mg) (Corbion Purac, Netherlands) was dissolved in dichloromethane (DCM) (500 μL) into which PEI:siRNA complex (150 μL) was added and probe sonicated on ice (50 W, 30 s). The emulsion was added into PVA solution (3 mL, 4%) and sonicated. This double emulsion was mixed with another polyvinyl alcohol (PVA) solution (7.5 mL, 0.3%) and mixed vigorously for 3 h with a magnetic stirrer at room temperature to evaporate DCM. The suspension was centrifuged (18,000 rpm, 10 min), and pellet was washed twice with nuclease-free dH2O and freeze-dried for 8 h under vacuum.
Determination of cell viability
Cell viability was measured by the Alamar Blue cell viability assay. RAW264.7 cells (5 × 103) were seeded on 96-well plates (30 μL). PLGA nanocapsules (0.5 mg) were diluted in OptiMEM (30 μL) and added dropwise onto the cell suspension. The cells were incubated for 6 h, and growth medium (140 μL) was subsequently added. The cell viability was assessed for 5 days with a medium change on day 3. On the day of experiment, the medium was removed, and Alamar Blue solution (10% in colorless DMEM, 200 μL) was added onto the cells and incubated for 150 min. The optical density was measured at 570 and 595 nm with a microplate reader (SpectraMax M2; Molecular Devices), from which percent reduction and cell numbers were calculated using a calibration curve for the cell line plotted from the percent reductions and corresponding cell number.
Endocytosis of the nanocapsules by the cells
To visualize the localization of PLGA nanocapsules in cells, Nile Red (λex 552 nm, λem 636 nm; Thermo Fisher Scientific) loaded PLGA nanocapsules were prepared by dissolving Nile red dye in DCM phase (0.2 mg/mL) during nanoparticle preparation. Nile red loaded PLGA nanocapsules (6 mg in 200 μL OptiMEM) were mixed with RAW264.7 cell suspension (1.2 × 105) in an Eppendorf tube and seeded on tissue culture polystyrene (TCPS) slides (2.8 cm2). On day 2, the cells were fixed with paraformaldehyde (PFA) (4% w/v) for 15 min and permeabilized with Triton X-100. After blocking with bovine serum albumin (1% w/v in phosphate buffered saline [PBS]), the cells were incubated for 1 h, 1:50 dilution in PBS, Alexa 488-labeled phalloidin (Thermo Fisher Scientific), and DRAQ5, 30 min at 37°C, 1:1000 dilution in PBS (Cell Signaling Technologies). The internalization and localization of the PLGA nanoparticles were visualized by confocal laser scanning microscopy (CLSM) (Leica DM2500, Germany).
Inhibition of RANK mRNA in nanocapsule transfected cells
To determine the RANK mRNA inhibition, 6 mg of empty PLGA nanocapsule (NC), uncomplexed RANK siRNA loaded nanocapsule (R-NC), PEI:Scrambled siRNA loaded nanocapsule (PSc-NC), and PEI:RANK siRNA loaded nanocapsule (PR-NC) were suspended in Opti-MEM (200 μL). RAW264.7 cells (1.2 × 105 cells/well) were seeded on 12-well plates, and the nanocapsules were added dropwise onto the cell suspension (200 μL) and incubated for 6 h. Growth medium was added (800 μL), and gene silencing was monitored on days 3 and 5.
Quantitative real-time PCR
The changes in RANK mRNA were studied using qRT-PCR (Rotor-Gene Q; Qiagen) using 2−ΔΔCt relative quantification method. The primer pairs used are presented in Table 2. On days 3 and 5, RNA isolation was performed using RNeasy Micro Kit (Qiagen) according to the manufacturer's protocol. The isolated RNA samples were treated with DNase I (DNA-free™ Kit; Ambion, Invitrogen, Germany) to prevent possible DNA contamination and converted to cDNA using RevertAid First Strand cDNA Synthesis Kit (Invitrogen). qRT-PCR was performed with the cDNA samples obtained. The qRT-PCR cycling conditions for both RANK and GAPDH primers were as follows: 95°C, 5 min initial denaturation, followed by denaturation at 95°C for 5 s, annealing, and extension at 60°C for 10 s for 35 cycles. The reaction was terminated with melt curve analysis (50°C–90°C; 1°C/1 cycle).
Quantitative Real-Time PCR Primers and the Amplicon Sizes
Inhibition of osteoclast differentiation through nanocapsule transfection
The differentiation of RAW264.7 cells was analyzed by Acid Phosphatase Leukocyte Kit (TRAP) (Sigma). RAW264.7 cells (1.4 × 104) were mixed with RANKL (100 ng/mL), macrophage colony stimulating factor (M-CSF) (20 ng/mL), and PLGA nanocapsules (0.7 mg) and seeded onto TCPS slides (2.8 cm2). On day 5, the cells were fixed with PFA (4%), and TRAP enzyme was stained by TRAP kit according to manufacturer's protocol. The entire images of three TCPS slides from separate runs were obtained by Zeiss Brightfield MosaiX Protocol (Zeiss Axio Imager M2, Germany). TRAP positive multinucleated cells on the slides were counted as osteoclasts using ImageJ Analysis Software (NIH).
Inhibition of osteoclastic activity
The inhibition in osteoclastic activity was studied by the pit formation assay performed on 96-well Osteo assay surface plates (Corning). RAW264.7 cells (5 × 103, 50 μL) were mixed with RANKL (100 ng/mL), M-CSF (20 ng/mL), and PLGA nanocapsules (0.4 mg, 50 μL Opti-MEM) in 96-well Osteo assay surface plates. After complete attachment, the medium was completed to 200 μL. On day 5, the cells were treated with bleach solution (10%, 100 μL) for 5 min at room temperature. The wells were washed twice with dH2O and dried. The images of resorption pits were taken at 20× magnification by another scientist who was not involved in the study. The resorbed pit areas were quantified using 15 images from each group (n = 3) by Image J Analysis Software (NIH, USA).
Statistical analysis
All quantitative data presented in the study are expressed as mean ± standard deviation. Statistical differences among the study groups in qPCR and Alamar Blue cell viability experiments were assessed by two-way analysis of variance (ANOVA) with Tukey's post hoc analysis. The statistical differences among groups in TRAP staining and pit formation assays were assessed by one-way ANOVA with Tukey's post hoc analysis. p-Value ≤0.05 was considered to be statistically significant (GraphPad Prism v6.0).
Results
Inhibition of RANK mRNA by PEI/siRNA complexes with varying N/P ratios
In our earlier work, an N/P ratio of five was needed for complete complexation and charge neutralization of PEI:nucleic acid complexes. 26 In the present study, therefore, to find out which N/P ratio was appropriate for significant RANK mRNA inhibition, complexes with higher N/P ratios (N/P 6, 20, 25, 30, and 60) were studied, where the siRNA was in fully complexed form with PEI without PLGA encapsulation (Fig. 2). When N/P ratio was 6, no alteration in the RANK mRNA expression was observed (data not shown). A N/P ratio of 20 led to a ∼28% inhibition in RANK mRNA levels after 24 h. Inhibitory effect of PEI on RANK mRNA was observed in complexes with an N/P ratio of 25 at 48 h post-transfection as shown by PEI (dark gray bars) and PEI:Scrambled siRNA transfected cells (light gray bars) (Fig. 2).
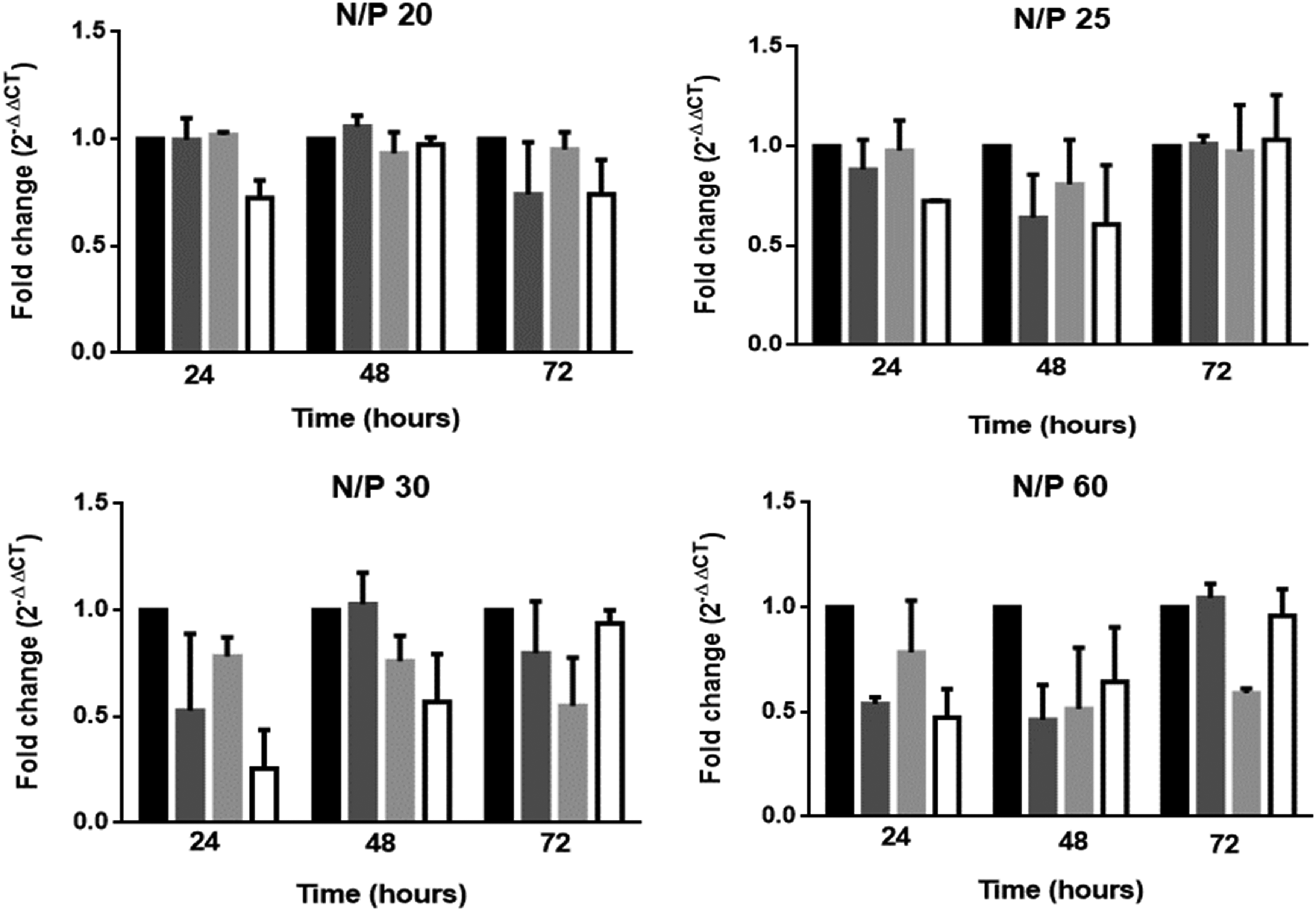
Fold change in RANK mRNA expression with N/P ratios of 20, 25, 30, and 60. Bars: black, untransfected cells; dark gray, PEI transfected cells; light gray, PEI:Scrambled siRNA transfected cells; white, PEI:RANK siRNA transfected cells (n = 2). PEI, polyethylenimine; siRNA, small interfering RNA.
During the transfection period, floating dead cells were observed when the cells were treated with N/P ratios of 30 and 60 (where the PEI concentrations were 1.5 and 3 μg/well, respectively), along with the inhibition on RANK mRNA levels probably due to the presence of high concentrations of PEI. Based on these results, N/P 20 was used in the later experiments in encapsulation in PLGA nanocapsule and determination of the efficacy of the delivery system constructed.
Effect of PLGA nanocapsules on cell viability
In this study, a possible cytotoxic effect of PLGA nanocapsules on RAW264.7 cells was studied with Alamar Blue Cell Viability assay (Fig. 3). The amount of nanocapsule to be used in these experiments was determined based on the encapsulation efficiency, which was 48% according to our previous study. 26 The results showed that PLGA nanocapsules do not significantly affect the attachment of RAW264.7 cells on the TCPS surface on day 1. Between days 1 and 5, both untransfected and empty nanocapsule-treated cells proliferated approximately with similar rates. These indicate that the PLGA nanocapsules do not significantly affect RAW264.7 cell viability.
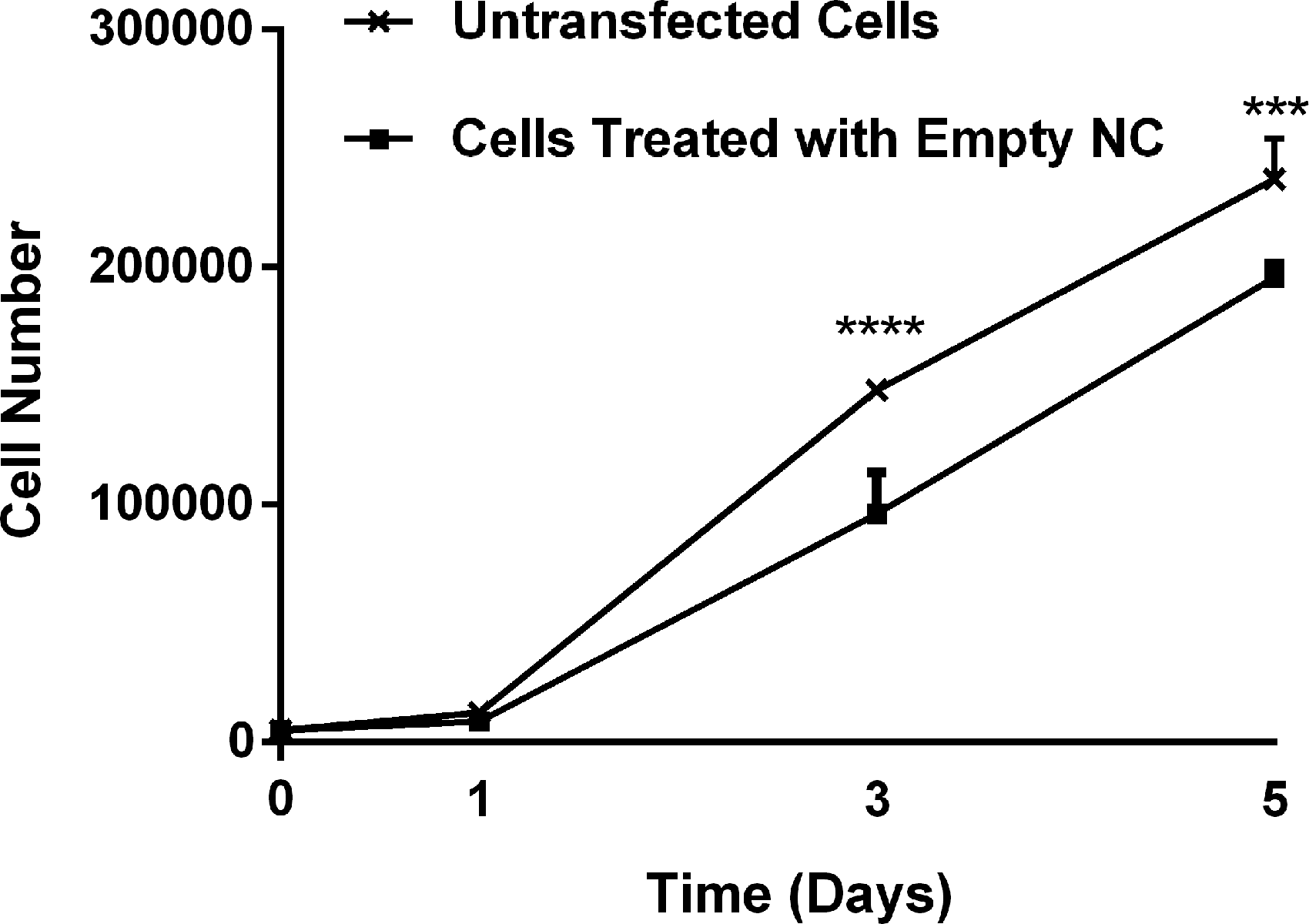
The effect of PLGA nanocapsules on the viability of RAW264.7 cells. Seeded cell density: 5 × 103 cells/well. The indicated significance values are the result from the comparison of untransfected and empty nanocapsule-transfected cells at each time point (n = 3). ***p < 0.0005, ****p < 0.0001. NC, nanocapsule; PLGA, poly(lactic acid-co-glycolic acid).
Endocytosis of the nanocapsules
CLSM was used to study the internalization of Nile Red stained PLGA nanocapsules by the RAW264.7 cells (Fig. 4). The nanocapsules were seen to have crossed the cell membranes and internalized in the cytoplasm. This is an indication of the potential of the PLGA NC to be effective in delivering their drug cargo (siRNA) into the cytoplasm.
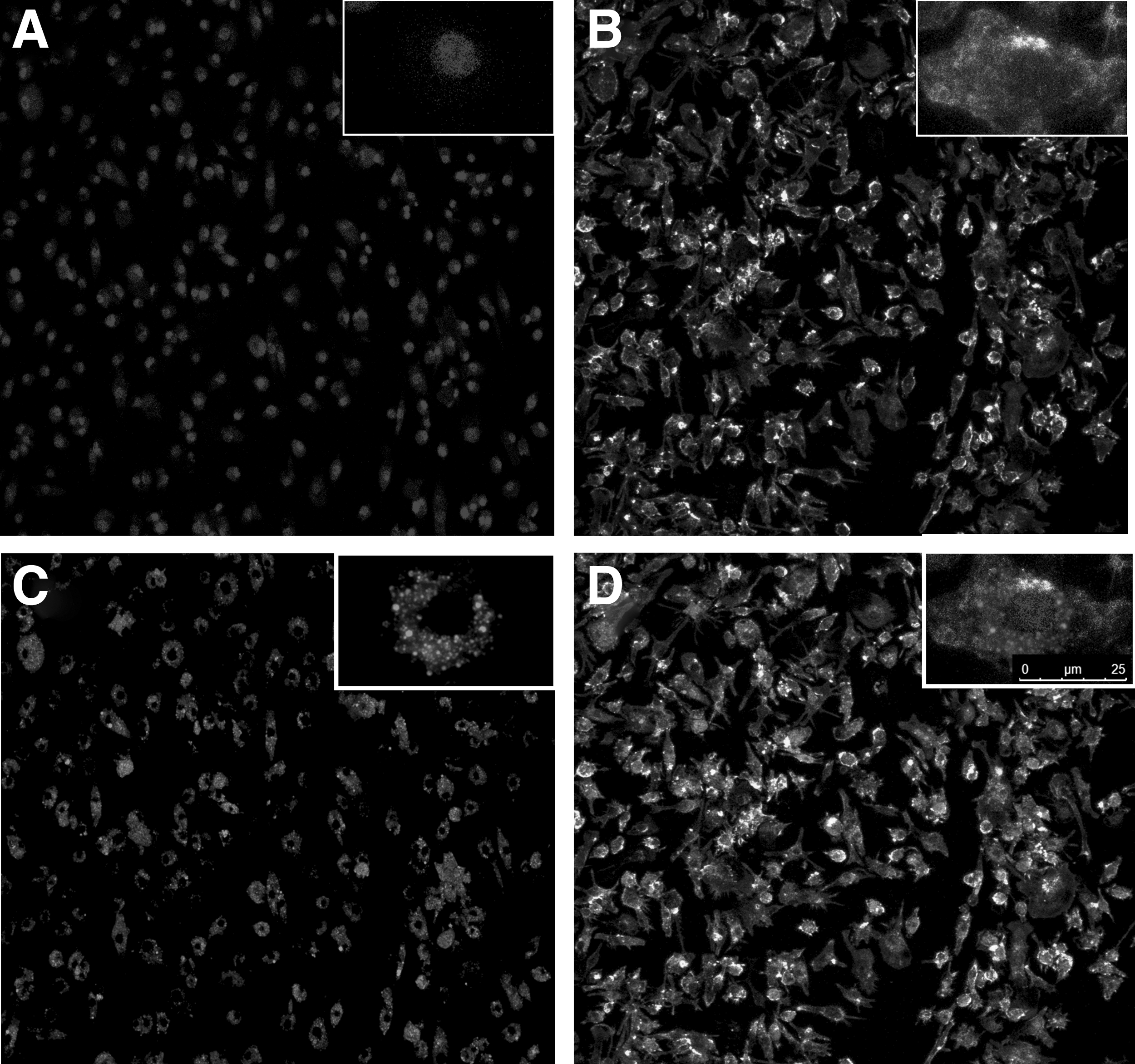
CLSM of RAW264.7 cells transfected with Nile Red-stained PLGA NP.
Inhibition of RANK mRNA by transfection with PR-NC
The effect of PEI:RANK siRNA loaded nanocapsules (PR-NCs) was investigated by qRT-PCR. The controls of the study were empty PLGA nanocapsules (NCs), uncomplexed RANK siRNA loaded nanocapsule (R-NC), and PEI:Scrambled siRNA loaded nanocapsule (PSc-NC) PLGA nanocapsules. The fold change in RANK mRNA was calculated using the NC transfected cells as the reference group (Fig. 5). The fold change of RANK mRNA was calculated to be (0.53 ± 0.10) on day 3, indicating a 47% inhibition (p < 0.05). On day 5, the fold change of RANK mRNA was 0.91 ± 0.15, which was not statistically significant in comparison to PSc-NC. Meanwhile, the R-NC did not lead to RANK mRNA inhibition at both time points (1.12 ± 0.32 on day 3 and 1.30 ± 0.21 on day 5).
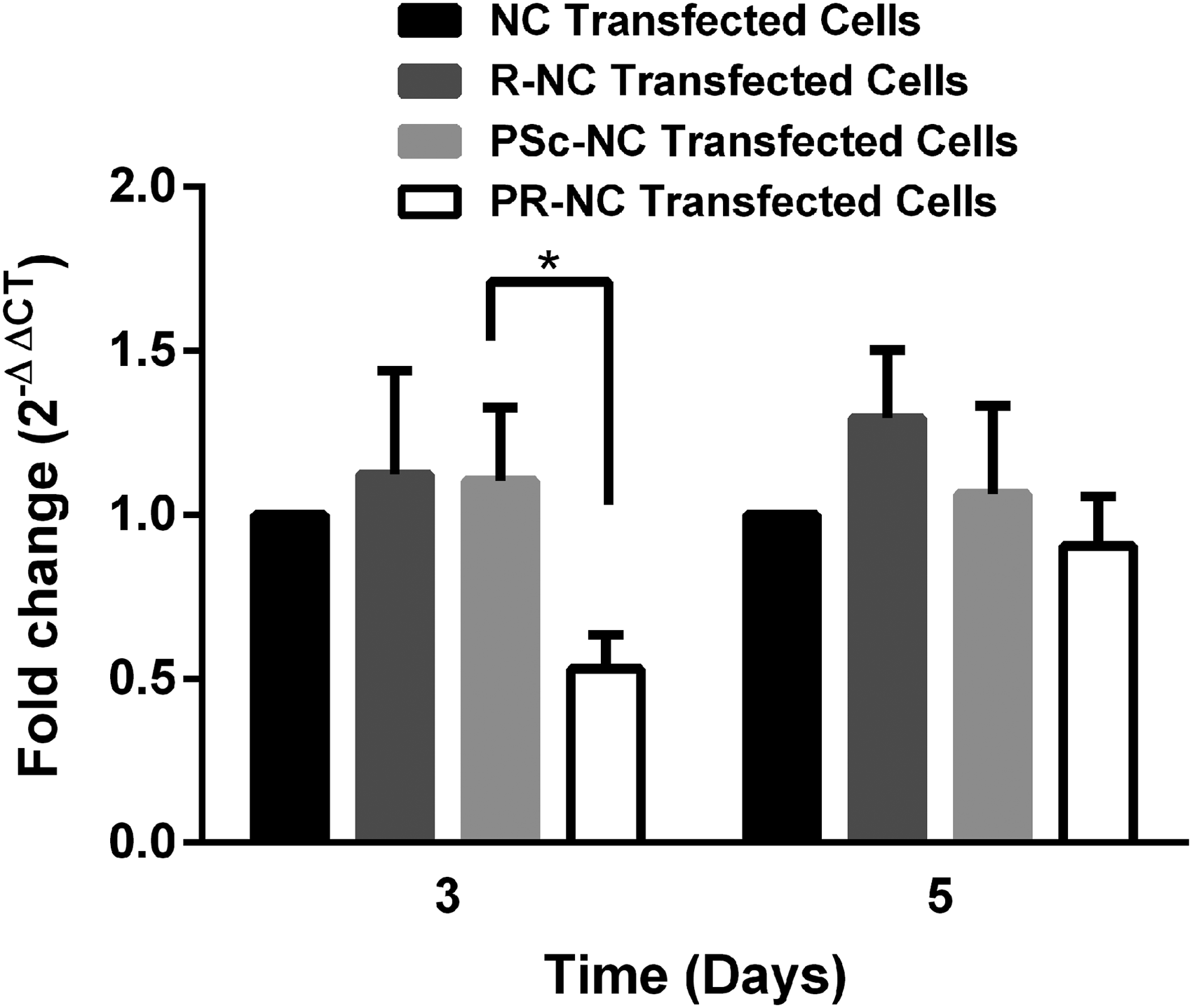
Fold change in RANK mRNA expression on days 3 and 5 post-transfection with PLGA nanocapsules carrying different cargo (n = 3). *p < 0.05. R-NC, uncomplexed RANK siRNA loaded nanocapsule; PSc-NC, PEI:Scrambled siRNA loaded nanocapsule; PR-NC, PEI:RANK siRNA loaded nanocapsule.
Inhibition of osteoclastogenesis by transfection with siRNA loaded NC
The differentiation of RAW264.7 preosteoclasts to mature osteoclasts was studied by TRAP enzyme staining after the cells are treated with three samples: NC, PSc-NC, and PR-NC along with M-CSF and RANKL for 5 days. Representative images of differentiated RAW264.7 osteoclasts on day 5 are presented in Figure 6. TRAP positive multinucleated cells on the micrographs were counted, and osteoclastogenesis was decreased to 54.7% for PR-NC transfected cells compared to PSc-NC transfected cells.
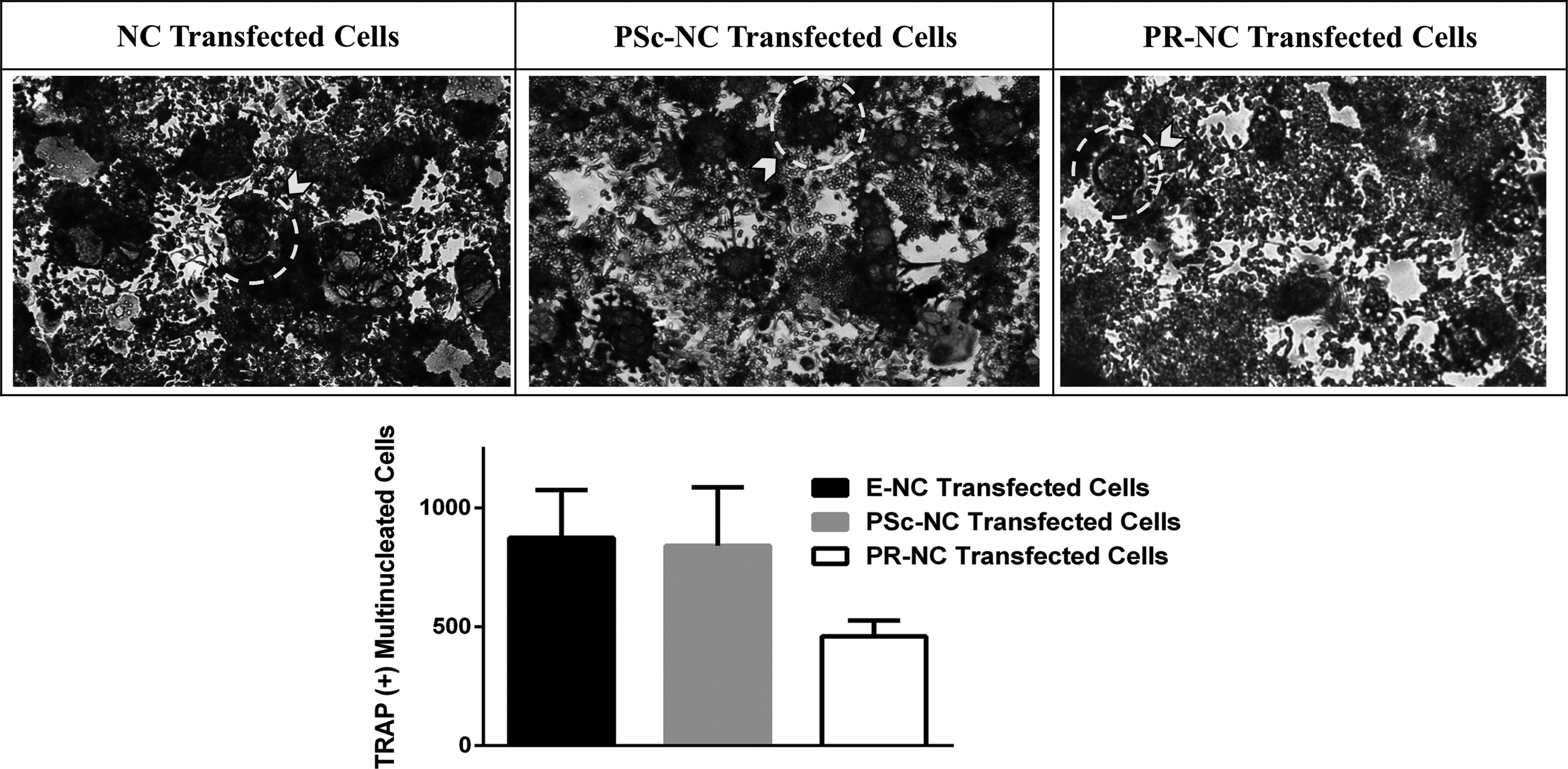
Inhibition of osteoclast differentiation assessed by TRAP staining. Top panel: representative TRAP staining images of differentiated RAW264.7 cells transfected with PLGA nanocapsules (NCs), PEI:Scrambled siRNA loaded nanocapsule (PSc-NC), and PEI:RANK siRNA loaded nanocapsule (PR-NC) on day 5. Bottom panel: quantification of TRAP (+) multinucleated osteoclasts. Dotted circles shown with arrows are examples of TRAP (+) osteoclasts in each image.
Inhibition of osteoclastic activity
The activity of the differentiated RAW264.7 was studied with pit formation assay using the Osteo assay surface 96-well plates (Corning). The total area of those resorption pits was used as a quantification method for the osteoclastic activity. The images presented in Figure 7 are the batch analysis of the 15 images obtained from each well (n = 3). As can be seen from the Figure, induction of RAW264.7 cells with RANKL resulted in formation of resorption pits (Fig. 7, untransfected cells), supporting the role of RANKL treatment to differentiate the osteoclast precursors into mature and functional osteoclasts. Treatment with NC or PSc-NC did not cause reduction in the activity of the maturated osteoclasts. When the cells were treated with PR-NC, however, the activity of osteoclasts significantly reduced, approximately by its half, compared with PSc-NC treatment.
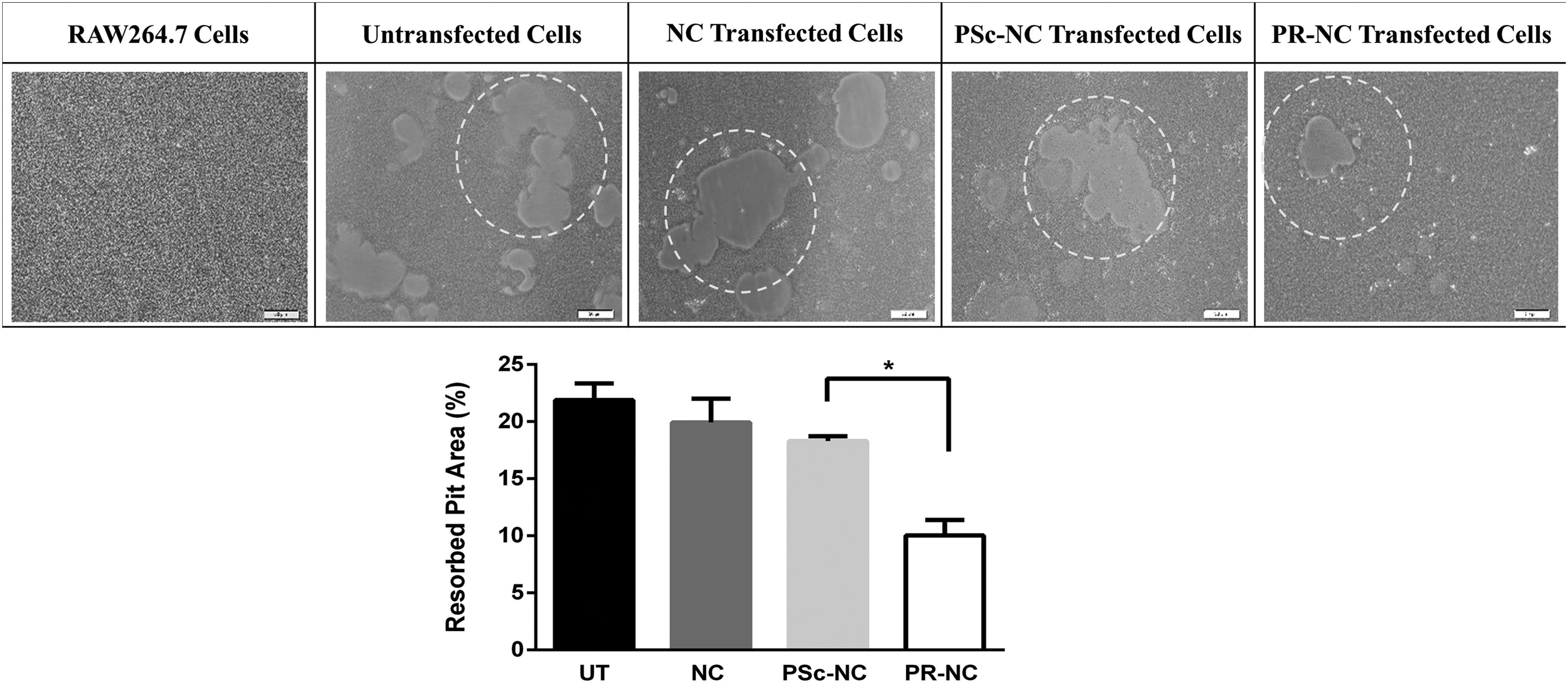
Osteoclastic activity determined by pit formation assay. Top panel: representative images of resorbed areas of undifferentiated RAW264.7 cells, untransfected cells (UT), and differentiated RAW-OC cells transfected with PLGA nanocapsules carrying different cargo. NC: nanocapsule, PSc-NC: PEI:Scrambled siRNA loaded nanocapsule, PR-NC: PEI:RANK siRNA loaded nanocapsule. Bottom panel: quantification of resorbed pit area. (n = 3). Dotted circles show examples of resorbed pits in the micrographs. *p < 0.05.
Discussion
New strategies developed for osteoporosis treatment focus on the use of RNAi approach with the biomaterials to inhibit the expression of genes involved in the pathogenesis of the disease. This study was conducted to develop a siRNA delivery system for the treatment of osteoporosis.
Initially, the inhibitory effects of two different RANK siRNA sequences were compared for their effectiveness, and the siRNA leading to most significant RANK mRNA inhibition was chosen for use in the rest of the studies (Supplementary Fig. S1). This siRNA was then complexed with PEI, and the complex was loaded into PLGA nanocapsules to be transported in a stable form. The constructed delivery systems and their controls were tested on preosteoclast cell line, RAW264.7, to compare their toxicity, nanoparticle internalization, and efficacy.
It was found that empty PLGA nanocapsules did not influence the proliferation of RAW264.7 cells (Fig. 3), even when they were internalized by the cells (Fig. 4). It is known that the internalization of nanoparticles is controlled by various factors such as size, charge, and chemical composition of the particles and the cell type, 29 and it is initiated by the attachment of the nanoparticles to the cell membrane by electrostatic interaction which is affected by the charge of the particle 30 or the presence of a targeting moiety on the capsule to interact with a cell receptor. Positively charged nanoparticles are more advantageous in binding to cells than the negatively charged carriers because of the negative charge of the cell membrane. However, positively charged molecules are more toxic to cells, in a dose dependent manner, compared to negatively charged particles. 31 In contrast, the influence of cell type on particle internalization cannot be ignored. Unlike nonphagocytic cells, phagocytic cells are known to interact more with negatively charged particles. 32 The cell type used in this study, RAW264.7, is reported to internalize the nanoparticles mainly through phagocytosis. 33 Our results show that the particles used in this study were negatively charged (zeta charge −10.96 ± 3.84) and were effectively internalized probably because of cell type (Fig. 4). Similar results were also obtained in other studies. Wang et al., showed the internalization of negatively charged PLGA microparticles by murine primary monocyte macrophages after 3 days of incubation. 34 In another study, the preferential uptake of the anionic polystyrene nanoparticles (with a size of 116 nm) by human macrophages was also shown. 35 The confocal microscopy images of the internalized particles show that they are located in the cytoplasm. This is an encouraging result for our goal since our target, the mRNA, is located in the cytoplasm where siRNA functions to inhibit the protein translation from the mRNA. In the cytoplasm, PEI:siRNA loaded PLGA nanocapsules can act as drug depots and release their content, so that siRNA could function once it is dissociated from PEI.
The release of siRNA from PEI depends on the strength of the electrostatic interactions between the positively charged amino groups of PEI (N) and the negatively charged phosphates of siRNA (P) forming the complex. The ratio of N/P of the complex is also important in the endosomal escape of the complex into the cytoplasm because mainly the free, uncomplexed PEI expands volumetrically in the endosome leading to the permeabilization of its lipid bilayer membrane. 36 The difference in the mRNA inhibition efficacies of complexes carrying the same siRNA but different PEI amount, and therefore different N/P, (Fig. 2) might be explained by this phenomenon. The excess PEI in the N/P 20 complexes might have caused higher escape from the endosomes and thus led to an efficacy higher than with the N/P 6 complexes. This result is parallel with other studies in the literature 34 where the N/P ratio of 20 was used in mRNA knockdown, which resulted in higher gene silencing compared with the silencing capabilities of complexes with lower N/P ratios such as 15 and 10.
When the N/P 20 complexes were loaded in PLGA nanocapsules, it resulted in significant RANK mRNA inhibition (47%) 3 days post-transfection (Fig. 5). On day 5, the inhibition was reduced to ∼10% which might be due to dilution of the nanocapsules, thus the siRNA, after the routine culture medium change on day 3. After medium change, further cell division decreases the amount of nanoparticles within each cell. The inhibitory effect of uncomplexed RANK siRNA loaded nanocapsules (R-NCs) was also studied, and no significant reduction in RANK mRNA was observed 3 days post-transfection. Similar results were also observed by other groups. 37 It was shown that although naked siRNA loaded PLGA microspheres caused inhibition in the first 12 h, the level of this inhibition was reduced during the 8 days of culture period. They concluded that the addition of a cationic carrier such as PEI into the microspheres could achieve long-lasting release of siRNA in the cells and thus silence the target. The absence of PEI would allow the degradation of the siRNA in the lysosomes as naked siRNA is more prone to degradation by nucleases compared with its complexed form with PEI because complexation with PEI protects the siRNA.
The effect of PR-NC on suppressing the differentiation of osteoclast precursors into mature osteoclasts was studied by staining TRAP enzyme (Fig. 6), which is the marker for mature osteoclasts.38,39 It was observed that transfection with PR-NC resulted in reduced osteoclast formation compared to transfection with PSc-NC. Furthermore, we have inhibited the osteoclastic activity by treating the cells with PR-NC (Fig. 7). It was concluded that the reduction in pit formation might be due to, at least in part, formation of fewer osteoclasts as was determined by TRAP staining, together with the reduced activity of mature osteoclast as a result of RANK inhibition.
In the literature, there are studies concerning the treatment of osteoporosis using siRNA as a therapeutic approach, and they include liposomes or polymer-based particulate systems for systemic delivery.22,25 In this study, we used PLGA nanoparticles for siRNA delivery to inhibit RANK mRNA in osteoclast precursors to suppress osteoclastic differentiation and activity, which are the major problems associated with the pathogenesis of osteoporosis. The delivery of siRNA targeting RANK on osteoclastic cells was also studied by Wang et al. 34 where they focused on local delivery of RANK siRNA to the bone using CPC cement, which was used to improve the fragility of the tissue.
Our delivery system, on the other hand, was designed to deliver its siRNA content systemically to treat osteoporosis through injection of a complex of siRNA carried in PLGA nanocapsules. The siRNA loaded nanocapsules were in the size range of the fenestrations in the bone marrow and, therefore, are expected to extravasate through the epithelium into the bone marrow. Once the nanoparticles are in the bone microenvironment, they would be internalized by osteoclast precursors, and the nanoparticles would release their PEI:RANK siRNA complex to inhibit the differentiation of the osteoclasts. In our previous study, we had shown that the PLGA nanocapsules could be successfully coated with an osteoconductive peptide, ELR (ELR-HAp), to target the nanocapsules to the bone tissue. In the current study, we did not use ELR-coated nanocapsules and studied the efficacy of the PR-NC in the inhibition of osteoclastic maturation and function. In vivo studies will be performed to study the bone targeting capability and efficacy of ELR-coated PR-NC.
Conclusion
The results of the present study show that an effective nanoparticulate siRNA delivery system was successfully developed to be used as a treatment tool against osteoporosis. The PLGA nanocapsules were not toxic and were readily internalized by RAW264.7 cells. Once internalized, they were localized in the cytoplasm, acting as drug depots to release their PEI:siRNA complex content. PLGA nanocapsules loaded with PEI:RANK siRNA complex at an N/P ratio 20 resulted in significant reduction in RANK mRNA levels in osteoclast precursor cells. The nanocapsules also caused suppression of osteoclast differentiation and activity. The constructed delivery system will be tested as a novel systemic delivery tool in the treatment of osteoporosis.
Footnotes
Acknowledgments
The authors acknowledge BIOMATEN, the Middle East Technical University Center of Excellence in Biomaterials and Tissue Engineering for the use of the facilities. Deniz Sezlev Bilecen acknowledges TUBITAK BIDEB 2211-c scholarship.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.