
Abstract
Post-traumatic and focal cartilage defects of the knee affect over 3 million Americans annually. Autologous cell-based cartilage repair, for example, autologous chondrocyte implantation, is limited by the need for ex vivo chondrocyte expansion and donor site morbidity. Mesenchymal stem cells (MSCs), owing to their relative ease of isolation, higher replication activity, and chondrogenic potential, represent an alternative reparative cell type. Platelet-rich plasma (PRP) is an autologous, growth factor-rich biologic preparation that has recently received increasing attention and use as a therapeutic adjunct for the treatment of degenerative joint diseases, and there is evidence suggesting that PRP acts by promoting stem cell proliferation and tissue healing. In this study, we have examined the effect of PRP treatment on chondrogenic differentiation of adult human MSCs derived from infrapatellar fat pad-adipose stem cells (IFP-ASCs) and bone marrow (BM-MSCs). Both cell types were placed in high-density pellet culture and hydrogel-encapsulated culture under chondrogenic conditions. Our results showed that PRP did not improve IFP-ASC or BM-MSC chondrogenesis. In general, chondrogenesis was inhibited with increasing PRP concentrations and duration of exposure, on the basis of histological, biochemical, and gene expression analyses. Taken together, these findings suggest that although PRP is reported to be beneficial in terms of pain relief and joint function improvement, its mechanism of action is unlikely to directly involve enhancement of MSC-mediated hyaline cartilage formation.
Introduction
C
Although both ACI and MACI have shown promise, challenges still persist. Chondrocyte yield from non-weight-bearing cartilage is low, requiring ex vivo cell expansion, with an accompanying loss of the chondrogenic phenotype. 8 As a result, the patient must undergo two surgical procedures and the quality of cartilage repair remains inconsistent. Identification of alternative cell sources and application of improved chondroinductive conditions are needed to overcome these challenges, permitting a single-step procedure with robust cartilage formation.
Adult tissue-derived mesenchymal stem cells (MSCs) are capable of undergoing chondrogenic differentiation and have traditionally been isolated from the iliac crest, an invasive procedure that yields a relatively low percentage of MSCs. Alternatively, adipose tissue contains a higher percentage of MSCs, is abundantly available, and can be harvested through a minimally invasive lipoaspiration procedure. 9 However, MSCs derived from adipose tissue, adipose stem cells (ASCs), have been reported as less chondrogenic than MSCs derived from bone marrow (BM-MSCs).10,11
Recently, the infrapatellar fat pad (IFP), readily accessible intraoperatively during arthroscopic and open knee surgeries, was examined as an alternative source of ASCs (IFP-ASCs), which were shown to possess a greater chondrogenic potential than subcutaneous ASCs (SQ-ASCs). 12 Specifically, it was reported that IFP-ASCs were superior to SQ-ASCs in terms of sulfated glycosaminoglycan (GAG) deposition and chondrogenic gene expression, with higher gene expression levels of aggrecan, SOX9, and collagen type II, and lower levels of collagen types I and X. However, the total cell number of IFP-ASCs is more limited compared to SQ-ASCs due to smaller tissue volume. Therefore, ex vivo culture expansion of IFP-ASCs would likely be required to obtain a sufficient cell number for implantation.
Alternatively, a strategy that promotes in situ proliferation and chondrogenic differentiation of IFP-ASCs could provide a single-step treatment for chondral defects. Platelet-rich plasma (PRP), derived from whole blood, contains a large number of growth factors, and has been shown to promote stem cell proliferation and neotissue formation in cartilage defects. 13 In vivo, a number of studies have reported pain reduction and improvement of joint function with PRP treatment, 14 while others have found no improvement in cartilage formation.15,16 The effect of PRP on chondrogenic differentiation of MSCs in vitro is also equivocal,17,18 with recent findings suggesting that the effects are dependent on PRP concentration. 19 The purpose of this study was to evaluate the effect of different PRP concentrations and exposure durations on proliferation and in vitro chondrogenic differentiation of IFP-ASCs and BM-MSCs when cultured as pellets or within a 3D hydrogel biomaterial scaffold.
Materials and Methods
Materials and reagents
All chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO), unless indicated otherwise.
PRP releasate preparation
PRP releasate was prepared according to an adapted protocol. 20 In brief, anticoagulated human whole blood (Central Blood Bank, Pittsburgh, PA) was distributed into 15 mL conical tubes and centrifuged at 480g for 20 min. Pelleted red blood cells were discarded and the supernatant (including the buffy coat) was aspirated and concentrated to obtain a platelet concentration of 1 × 106 platelets/μL. To activate platelets, concentrated CaCl2 was added to a final concentration of 22.8 mM, followed by incubation at 37°C overnight. After another centrifugation at 2000g for 10 min, the fibrin clot was discarded and the supernatant, referred to as PRP releasate, was stored at −20°C until future use (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). According to the Platelet Activation White blood cells (PAW) classification system, the PRP releasate prepared here was categorized as P3-X-Aα. 21 Before use in experiments, equal volumes of PRP releasate from three donors were pooled.
Cell isolation
All adult human cells were isolated according to Institutional Review Board (IRB) approved protocols (University of Washington and University of Pittsburgh).
Human IFP-ASCs were isolated from IFPs of three different donors undergoing total knee arthroplasty (48-year-old female, 59-year-old male, and 65-year-old male). The tissue was digested with 1 mg/mL type I collagenase (Worthington Biochemical, Lakewood, NJ) and 1 mg/mL trypsin (Invitrogen, Carlsbad, CA) for 3 h at 37°C in an orbital shaker at 150 rpm. Cells were filtered through a 40 μm cell strainer (ThermoFisher, Pittsburgh, PA), pelleted by centrifugation at 1200 rpm for 5 min, and expanded in growth medium (GM: Dulbecco's modified Eagle's medium [DMEM; Invitrogen], 10% v/v fetal bovine serum [FBS; Gemini Bio Product, West Sacramento, CA] and 1% v/v antibiotic–antimycotic [Invitrogen]).
Human subcutaneous ASCs (SQ-ASCs) were isolated from lipoaspirate-derived fat tissue of three donors (34-year-old male, 38-year-old female, and 49-year-old female) using an automated cell isolation system (Icellator®, Tissue Genesis, Honolulu, HI). Cells were pelleted by centrifugation and culture-expanded in GM.
Human BM-MSCs were isolated from the bone marrow of three different patients undergoing arthroplasty (65-year-old male, 67-year-old male, and 68-year-old male). In brief, the trabecular bone marrow was rinsed with Minimum Essential Medium alpha (Invitrogen) containing 1% v/v antibiotic–antimycotic and filtered through 40 μm strainers. Cells were pelleted by centrifugation and culture-expanded in GM.
Cell characterization
The presence of surface markers, including CD31, CD34, CD44, CD45, CD73, CD90, and CD105, on SQ-ASCs and IFP-ASCs was assessed by flow cytometry using appropriate monoclonal antibodies (mouse anti-human antibodies; BD Biosciences, San Jose, CA). Detailed information on the antibodies used is provided in Supplementary Table S1.
Colony formation assay
Colony formation was analyzed by crystal violet staining. In brief, cells were cultured as monolayers on 10-cm Petri dishes (ThermoFisher, Pittsburgh, PA) at a seeding density of 100 cells per dish in GM for 14 days. On day 14, monolayer cultures were stained with 0.5% crystal violet (Sigma-Aldrich) in 100% methanol (ThermoFisher) for 10 min before imaging.
Differentiation assays
Monolayer cultures
Tri-lineage differentiation of SQ-ASCs and IFP-ASCs was analyzed via monolayer culture in, respectively: (1) chondrogenic medium (DMEM, 1% v/v antibiotic–antimycotic, 1× insulin–transferrin–selenium [ITS; ThermoFisher #41400045], 0.1 μM dexamethasone, 40 μg/mL proline, 50 μg/mL ascorbic acid, and 10 ng/mL recombinant human transforming growth factor-β3 [TGF-β3] [PeproTech #100-36E, Rocky Hill, NJ]); (2) osteogenic medium (DMEM, 10% v/v FBS, 1% v/v antibiotic–antimycotic, 0.1 μM dexamethasone, 10 mM β-glycerophosphate, and 50 μg/mL ascorbic acid); and (3) adipogenic medium (DMEM, 10% v/v FBS, 1% v/v antibiotic–antimycotic, 1× ITS, 1 μM dexamethasone, and 0.5 mM 3-isobutyl-1-methylxanthine). On day 21, the cultures were stained with Alcian Blue, Alizarin Red, and Oil Red O, using standard histological staining protocols, to assess chondrogenesis, osteogenesis, and adipogenesis, respectively.
High-density pellet culture
Chondrogenesis was also examined under conditions that mimic mesenchymal condensation during developmental cartilage formation as high-density pellet cultures. 2.5 × 105 IFP-ASCs or BM-MSCs were pelleted by centrifugation, and then maintained in serum-free chondrogenic medium. The effect of PRP on chondrogenesis was measured by varying PRP releasate concentration or exposure duration as follows: (1) pellets were cultured for 21 days in chondrogenic medium, which was supplemented with 10% v/v PRP releasate for the initial 0, 1, 3, 7, or 21 days (referred to as 1-, 3-, 7-, or 21-day pulse, respectively); and (2) pellets were cultured for 21 days in chondrogenic medium, which was supplemented with PRP releasate at 0, 1, 5, 10, or 20% v/v for the first 24 h. The 24-h exposure to PRP was intended to mimic the transient bolus of growth factors provided by a single intra-articular injection of PRP, as commonly used in clinical practice. 22 Culture medium was regularly changed three times a week, except for medium changes dictated by a particular PRP exposure duration indicated above.
3D hydrogel culture
We next examined the effect of PRP on MSC chondrogenesis in a 3D context more relevant to the translational application of MSCs in cartilage repair-based cartilage repair. MSCs were encapsulated in photocrosslinkable hydrogels previously shown to support robust chondrogenesis. 23 Visible-light-activated methacrylated gelatin (mGL)/methacrylated hyaluronic acid (mHA) hydrogels were fabricated as described previously. 24 In brief, 15 g of gelatin (Sigma-Aldrich) was dissolved in 500 mL H2O at 37°C, and 15 mL of methacrylic anhydride (Sigma-Aldrich) was added dropwise. The mixture was incubated with shaking at 150 rpm for 24 h, then dialyzed against water for 4 days, and mGL lyophilized for storage. The photoinitiator, lithium phenyl-2,4,6-trimethylbenzoylphosphinate (LAP), was prepared as described by Fairbanks et al. 25 For mHA, 5 g of sodium hyaluronate (66 kDa, Lifecore Biomedical, Chaska, MN) was dissolved in 500 mL H2O at 37°C, and 15 mL of methacrylic anhydride was added dropwise. At the end of every hour in the first 5 h, pH was adjusted to 7. The mixture was then incubated with shaking at 150 rpm for 24 h, dialyzed against water for 4 days, and lyophilized for storage.
For hydrogel fabrication, mGL and mHA were dissolved in Hank's Balanced Salt Solution at final concentrations of 9% and 1% (w/v), respectively. After adjustment of pH to 7.4 with 10 N NaOH, 1% antibiotic-antimycotic and 0.15% w/v LAP were added. IFP-ASC or BM-MSC pellets were resuspended in mGL/mHA solution at a final concentration of 15 × 106 cells/mL, and 50 μL of the suspension was transferred to a cylindrical mold measuring 5 mm diameter by 2 mm depth. The cell/monomer mixture was subjected to photoillumination (395 nm) for 90 s, forming a cell encapsulated mGL/mHA hydrogel. Constructs were cultured in Phenol Red-free chondrogenic medium supplemented with different PRP releasate concentrations (0, 1, 5, 10, and 20% v/v) for 21 days. Culture medium was changed twice a week.
Gene expression analysis
Both pellet and hydrogel cultures were collected on day 21, homogenized in QIAzol (Qiagen, Hilden, Germany), and total RNA isolated with RNeasy Plus Mini Kit (Qiagen). Gene expression levels of chondrogenesis markers (collagen type II, COL2; aggrecan, ACAN) and hypertrophy markers (collagen type X, COL10; matrix metalloproteinase 13, MMP13) were assessed by real-time reverse transcription polymerase chain reaction (RT-PCR) and normalized to that of 18S ribosomal RNA (gene primers sequences shown in Supplementary Table S2). Relative fold changes in mRNA levels were calculated using the ΔΔCt method.
Histological analysis
Pellets and hydrogels collected on culture day 21 were fixed with 4% phosphate-buffered paraformaldehyde, dehydrated, and embedded in paraffin. Histological sections (8 μm) were stained with Safranin O/Fast Green and Alcian Blue/Nuclear Fast Red for sulfated GAG. For collagen type II immunohistochemistry, deparaffinized sections were washed with 0.02% bovine serum albumin and incubated with 0.1 U/mL chondroitinase ABC and 250 U/mL hyaluronidase for antigen retrieval, followed by blocking with 3% H2O2 in 1% horse serum. Sections were then incubated with rabbit anti-human collagen type II (Abcam #ab34712, Cambridge, United Kingdom) at 4°C overnight, and then with biotinylated anti-rabbit secondary antibody. Immunolabeling was detected using the Vector ABC reagent (Vector, Burlingame, CA) and visualized with VIP Kit (Vector), with hematoxylin QS (Vector) as the counterstain.
Biochemical analyses
Pellet cultures were digested overnight at 65°C in a digestion buffer (pH 6.0) containing 2% (v/v) of a commercially available papain solution (from Papaya latex, Sigma-Aldrich, containing 15–30 mg protein/mL with ≥16 units/mg), 0.1 M sodium acetate, 10 mM cysteine HCl, and 50 mM EDTA. Sulfated GAG content was measured with the Blyscan GAG Assay (Biocolor, Carrickfergus, United Kingdom) according to the manufacturer's instructions, with chondroitin sulfate as a standard. GAG content was normalized to DNA content determined with PicoGreen Assay (Invitrogen).
Statistical analysis
Values of gene expression levels, GAG contents, and DNA contents are shown as mean ± standard deviation. Three to six independent trials were performed for each experiment. Statistical difference from control (No PRP group) was analyzed by one-way ANOVA and Dunnett's multiple comparisons using GraphPad Prism 7 (GraphPad Software, Inc., CA), with significance at p < 0.05.
Results
Comparison of IFP-ASCs and SQ-ASCs
In this study, ASCs obtained from both the IFP-ASCs and subcutaneous lipoaspirate (SQ-ASCs) were used. Both cell populations were confirmed by flow cytometry to exhibit MSC-associated surface markers. Positive expression of cell surface markers typical of adult MSCs, such as those derived from BM-MSCs, including CD44, CD73, CD90, and CD105, was detected, with negligible expression of epithelial and hematopoietic markers, including CD31, CD34, and CD45 (Fig. 1A). In addition, IFP-ASCs and SQ-ASCs were able to form colonies and differentiate into adipogenic, osteogenic, and chondrogenic lineages (representative images shown for IFP-ASCs in Fig. 1B). The chondrogenic capacity of ASCs isolated from the IFP-ASCs and subcutaneous lipoaspirate (SQ-ASCs) was next compared. Pellet cultures consisting of 2.5 × 105 IFP-ASCs or 2.5 × 105 SQ-ASCs each were maintained in chondrogenic medium. At culture day 21, the RT-PCR analysis showed that gene expression level of aggrecan was significantly higher in the IFP-ASC pellets than in the SQ-ASCs pellets, while no difference was observed in gene expression of MMP13 (Fig. 1C). Trends toward higher collagen type II and collagen type X expression in IFP-ASCs were also seen, but did not reach statistical significance. Taken together, these results indicate the preferred chondrogenic ability of IFP-ASCs over SQ-ASCs and their relevance as a candidate cell type for cell-based cartilage repair therapy.

Characteristics of monolayer cultured IFP-ASCs.
Effect of PRP treatment on chondrogenesis in pellet cultures
Gross histological observation revealed that the overall influence of PRP exposure was the inhibition of cartilage matrix deposition in IFP-ASC pellet cultures, as shown in Supplementary Figures S2, S3. Specifically, increasing PRP concentrations or duration individually, as well as additively, reduced chondrogenesis (Supplementary Figs. S2, S3). To examine the specific effects of PRP, we profiled gene expression, analyzed the biochemical composition, and carried out histological and immunohistochemical examination of pellet cultures during the process of IFP-ASC chondrogenesis, as a function of PRP concentration and treatment duration.
Varying duration of PRP treatment
Pellet cultures of BM-MSCs and IFP-ASCs maintained in chondrogenic medium were supplemented with 10% v/v PRP releasate for varying time periods during the 21-day culture. As shown in Figure 2, the magnitude of the PRP effect, that is, inhibition of chondrogenesis, depended on the duration of treatment. Increasing the duration of PRP exposure resulted in decreased expression of COL2, ACAN, and COL10, with negligible effects on MMP13 expression, in IFP-ASCs (Fig. 2A). Varying duration of PRP treatment did not affect cell number, on the basis of DNA content, in either IFP-ASC or BM-MSC pellets; however, increasing PRP treatment time resulted in decreasing GAG/DNA content in IFP-ASCs, but not in BM-MSCs (Figs. 2B, 3A). This effect was supported by histological staining, as increasing duration of PRP supplementation led to decreasing deposition of cartilage-specific extracellular matrix in IFP-ASC pellets, including sulfated GAG and collagen type II (Fig. 2C). As a result, the size of the pellet decreased as PRP concentration or treatment duration increased. In BM-MSC pellets, some decrease in cartilage extracellular matrix production was also observed, although the inhibitory effect of PRP was not as strong (Fig. 3B).
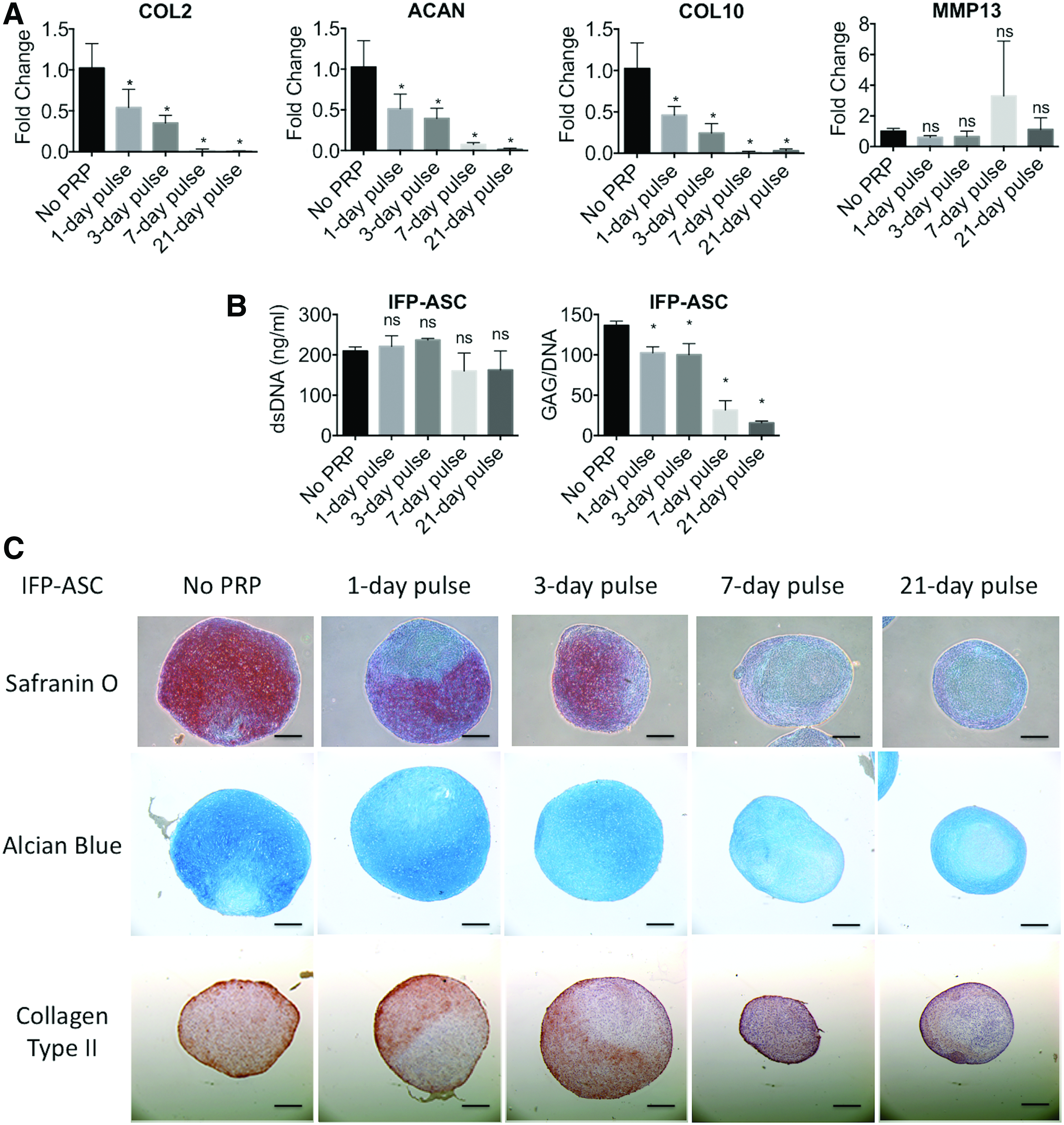
Effect of treatment with 10% PRP releasate for varying durations on gene expression, DNA and GAG content, and histology of IFP-ASCs undergoing chondrogenic differentiation as pellet cultures.

Effect of treatment with 10% PRP for varying durations on DNA and GAG content, and histology of BM-MSCs undergoing chondrogenic differentiation as pellet cultures.
Varying PRP concentration
For this study, transient PRP exposure, as in clinical intra-articular injections, 22 was simulated by exposing pellet cultures to PRP releasate supplementation only for the first 24 h of the 21-day culture period. In IFP-ASC pellets, gene expression profile remained relatively unchanged among the treatment groups exposed to different PRP concentrations (Fig. 4A), except that at low, 1% PRP, increased cartilage matrix production and collagen type II deposition was seen, compared to cultures treated with higher PRP concentrations (Fig. 4B). (Note: The first panel [No PRP] and the fourth panel [10% PRP, 1-day pulse] are repeated from Fig. 2C) In BM-MSC cultures, gene expression of COL2, ACAN, and MMP13 remained unchanged upon treatment with different PRP concentrations for 1 day; however, the hypertrophy associated gene, COL10, was significantly upregulated upon treatment with 20% PRP (Fig. 5A). Matrix sulfated GAG staining with Safranin O and Alcian Blue appeared similar in the various groups, except for 20% PRP, which showed slightly reduced staining intensity (Fig. 5B).
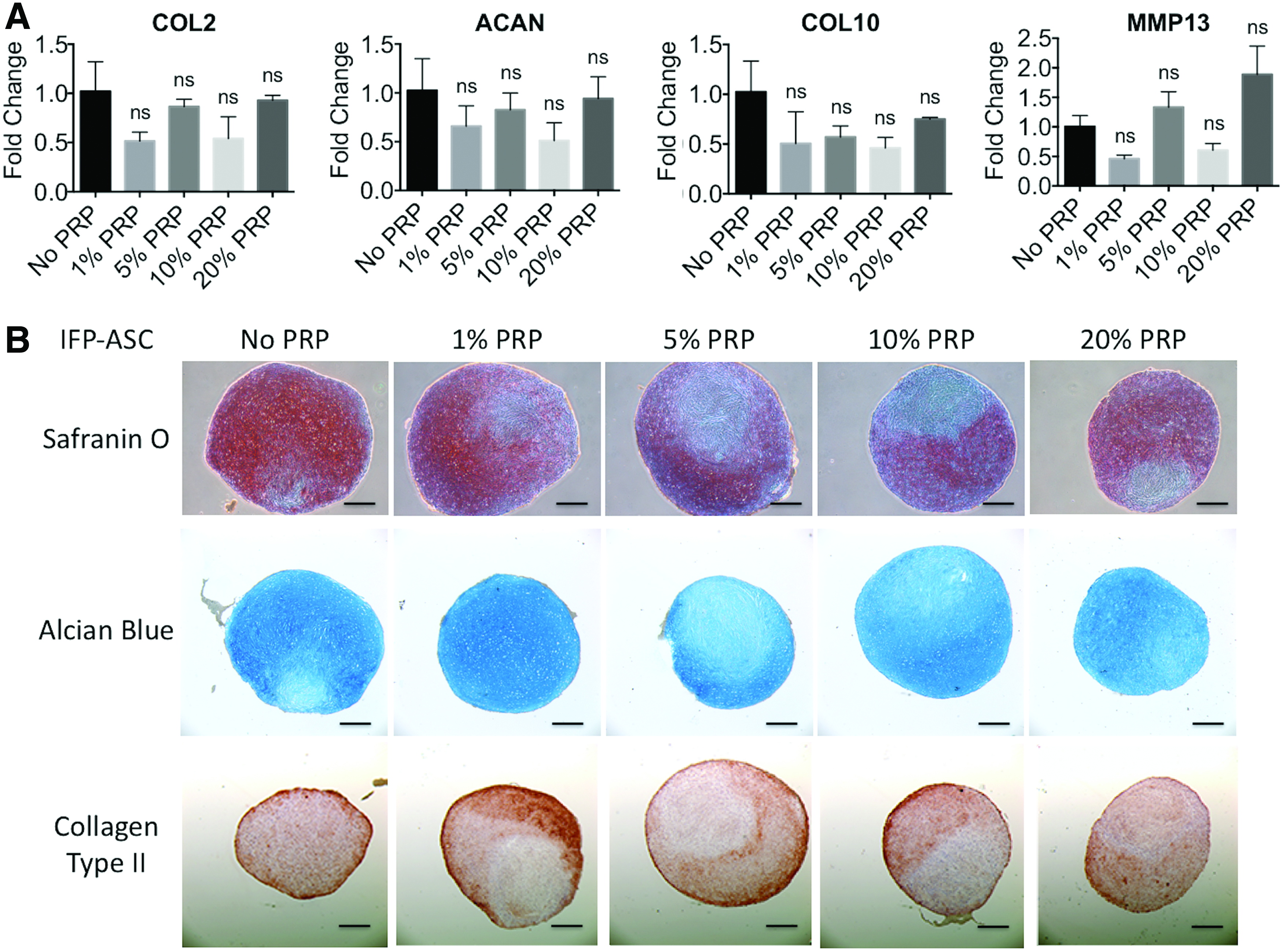
Effect of 1-day pulse PRP treatment at varying concentrations on gene expression and histology of IFP-ASCs undergoing chondrogenesis as pellet cultures.

Effect of 1-day pulse PRP treatment at varying concentrations on gene expression and histology of BM-MSCs undergoing chondrogenesis as pellet cultures.
Effect of PRP on chondrogenesis of MSCs in 3D hydrogel cultures
Given the preferred practice of encapsulating MSCs in 3D scaffolds for cartilage tissue engineering and repair, we next examined the effect of PRP exposure on chondrogenesis of MSCs in a 3D context, specifically by seeding cells in a photocrosslinkable hydrogel. IFP-ASCs were encapsulated in mGL/mHA hydrogels and cultured for 21 days in chondrogenic medium, with PRP supplementation at different concentrations. Our results showed that, with increasing PRP concentrations, the relative expression of COL2, ACAN, COL10, and MMP13 was reduced (Fig. 6A). In comparison, BM-MSCs in hydrogel cultures showed significant reduction only in COL2 and ACAN gene expression (Fig. 7A). Alcian Blue histological staining showed that PRP treatment inhibited pericellular GAG deposition (Figs. 6B, 7B).

Effect of 21-day PRP treatment at varying concentrations on gene expression and histology of IFP-ASCs undergoing chondrogenesis as 3D hydrogel cultures.

Effect of 21-day PRP treatment at varying concentrations on gene expression and histology of BM-MSCs undergoing chondrogenesis as 3D hydrogel cultures.
Discussion
PRP has received considerable recent attention as a potential therapeutic agent for tissue repair, including joint tissues; however, whether PRP enhances the formation of hyaline cartilage remains controversial. While some studies have reported pain reduction and improvement of joint function with PRP treatment, 14 others have reported no improvement of cartilage repair in in vivo models.13,15,16,26,27 In this study, our results clearly showed that PRP did not enhance chondrogenesis of IFP-ASCs or BM-MSCs in vitro.
For adult stem cell therapy, the BM-MSC is considered the gold-standard candidate cell type, by virtue of the large body of information currently available on its biological activities and responses.17,28,29 However, the biological benefits of PRP treatment on BM-MSCs, for the purpose of tissue repair and regeneration, are not well understood and remain controversial. For example, Haleem et al. reported a case study that all patients showed improvement 12 months after treatment of autologous BM-MSCs along with fibrin-containing PRP. 28 However, the advantage of PRP treatment was not observed in a porcine model. Betsch et al. reported that PRP alone or BM-MSCs alone significantly improved healing of osteochondral defects 26-week after surgery, but the combination of PRP and BM-MSCs did not further enhance wound healing. 30 In studying the efficacy of combining PRP and BM-MSCs, do Amaral et al. reported that higher concentration of PRP actually impaired chondrogenesis of BM-MSCs in pellet cultures. 19
In this study, we investigated the effect of PRP on BM-MSCs in both pellet and 3D hydrogel cultures, the latter to better recapitulate the microenvironment of MSCs encapsulated in 3D biomaterial scaffolds for clinical tissue repair applications. In both types of cultures, treatment dose and durations were varied to assess biologically optimal conditions. At 10% PRP, which approximates PRP concentrations used for intra-articular injections, 31 varying treatment durations (from 1-day pulse to 21-day pulse continuous treatment) did not yield significantly different results in pellet cultures, although PRP treatment for the first 24 or 72 h seemed to enhance GAG deposition. In hydrogel cultures, PRP treatment highly downregulated the expression of collagen type II and aggrecan, confirming that PRP does not enhance chondrogenesis in BM-MSC. While some studies have reported that PRP promoted cell proliferation of BM-MSCs, 32 we did not observe any PRP-mediated increase in DNA content in any group. In support, Krüger et al. also reported similar findings, namely that PRP cotreatment downregulated gene expression of aggrecan and collagen type II in TGF-β3-treated pellet cultures of subchondral bone progenitor cells. 33
IFP-ASCs, on the contrary, have not been extensively investigated in terms of their chondrogenic activity. Given the easy access to the IFP, for example via arthroscopic procedures, IFP-ASCs would present a more readily available cell source than BM-MSCs for point-of-care applications. We have therefore performed detailed characterization of the IFP-ASCs, in comparison to BM-MSCs. Our results showed that adipose tissue-derived stem cells expressed high levels of surface epitopes commonly detected on BM-MSCs, including CD44, CD73, CD90, and CD105, with negligible expression of surface markers for epithelial and hematopoietic cells, such as CD31, CD34, and CD45, consistent with reported findings.12,34 Interestingly, we also observed that IFP-ASCs performed better in terms of chondrogenesis than SQ-ASCs.12,35
The chondrogenic activity of IFP-ASCs maintained in 3D biomaterial scaffolds has been studied, and cell proliferation and cartilage matrix deposition have been reported for cultures in fibrin, agarose, collagen-hyaluronic acid, and polylactic acid hydrogel in vitro.36–40 At present, only one study has investigated the combination of IFP-ASCs and PRP. In this Level IV case study, 18 osteoarthritis patients were treated with autologous IFP-ASCs and 3 mL of PRP as scaffold via intra-articular injections. 41 Two-year follow-up showed promising results, including pain reduction and joint function restoration. However, in contrast to the potentially therapeutic effects reported by these authors, our in vitro analysis of the effects of PRP exposure on IFP-ASCs showed that PRP impairs chondrogenesis of IFP-ASCs in both pellet and 3D hydrogel cultures. These contrasting findings merit some discussion regarding the potential biological benefits of using PRP to treat joint cartilage-related pathologies.
A major complication in assessing the clinical benefit of PRP is the high variability of PRP preparations and the different tissue or disease targets. PRP preparation generally involves a two-step centrifugation: the first step fractionates platelet-poor plasma, buffy coat, and red blood cells; and the second step generates either leukocyte-containing PRP or pure PRP, depending on which portion is taken. 42 In our preparation, we collected both the buffy coat and platelet-poor plasma, resulting in leukocyte-containing PRP, a practice that is used in the majority of clinical practices. In view of these variabilities in PRP preparation, we propose that the following parameters and caveats must be taken into account in considering the efficacy of PRP treatment.
(a) PRP concentration and duration. Our results strongly suggest that PRP, when applied for cartilage-related treatments, must be used at low concentration and for short duration. Results from do Amaral et al. also showed that 2.5% PRP was better than 10% PRP applied in a collagen/hyaluronic acid scaffold in vitro. 19 Other studies have also suggested that higher PRP concentration was not better for chondrogenesis.43–45 Regarding the duration of PRP treatment, Park et al. reported a continuous increase of bone morphogenetic protein 7 and vascular endothelial growth factor (VEGF), both involved in bone remodeling, in rabbit chondrocyte cultures exposed to continuous presence of PRP for 10 days. 46 Taken together with our findings reported in this study, longer duration of PRP treatment very likely will lead to hypertrophy and bone formation.
(b) Platelet number. In our study, we resuspended platelets at 1.0 × 106 platelets/μL before calcium activation. Based on the PAW Classification System, our PRP releasate is categorized as P3 (0.75–1.25 × 106 platelets/μL). 21 Of relevance, a recent study showed that higher platelet number (fivefold to whole blood) diminished procollagen expression in anterior cruciate ligament fibroblasts. 47 Taken together, these findings suggest that higher platelet number is not necessarily higher performing for chondrogenesis.
(c) Platelet activation. In our PRP preparation, exogenous calcium chloride was added for platelet activation, since it has been shown to be more effective and inexpensive than exogenous bovine thrombin. 48 Calcium facilitates activation of endogenous thrombin in plasma to cleave fibrinogen into fibrin clots, which are typically removed, resulting in PRP releasate. Dohan Ehrenfest et al. summarized current protocols for pure PRP, leukocyte-rich PRP, pure platelet-rich fibrin (PRF), and leukocyte-rich PRF based on the speed and time of centrifugation and portion collected after centrifugation 42 ; however, a direct comparison between the four preparations in terms of cartilage repair was not performed. In the operating room, no exogenous platelet activation is required before PRP injections because collagen in the body acts as platelet agonist and activates platelets.
(d) Leukocyte number. As mentioned above, leukocyte-containing PRP was used in this study. Rios et al. showed that after lipopolysaccharide treatment, leukocyte-rich PRP released more growth factors and anti-inflammatory cytokines than pure PRP in healthy cartilage explants. 49 In a clinical trial, patients treated with leukocyte-containing PRP suffered from more swelling and pain when compared with leukocyte-reduced PRP group. 26 However, a prospective study of multiple clinical trials showed no significant difference between leukocyte-poor PRP and leukocyte-rich PRP on cartilage repair. 50 It is possible that inflammatory cytokines released from leukocytes induce inflammation (swelling and pain) in the early stage, but in the long term, leukocytes do not affect cartilage repair.
One of the limitations in this study is lack of information on the nature of the growth factor and cytokine profiles in the PRP preparation, which vary in different preparations depending on the methods of platelet activation. 48 Specifically, the levels of chondrogenesis-promoting factors, such as members of the TGF-β family, may regulate the extent of cartilage formation. Other factors, including VEGF and epidermal growth factor (EGF), may also impair the chondro-inductive effects of PRP. Matsumoto et al. showed that VEGF-transduced, muscle-derived stem cells performed poorly in chondrogenesis in a rat model.51,52 When combined with soluble VEGF receptor 1 (sFlt1), pellet cultures of the sFlt1-PRP-treated group showed more collagen type II deposition. 53 EGF receptor ligand that was identified in osteoarthritis cartilage promoted chondrocyte catabolic activity, but inhibited anabolic activity in a mouse model. 54 These findings suggest that VEGF and/or EGF, known components of PRP, may negatively impact the activities of other chondro-inductive components of PRP; thus, specific depletion of such antichondrogenic factors from PRP may enhance the prochondrogenesis effect of PRP on MSCs.
Another limitation of our study is that a PRP releasate pooled from several donors was used, to examine the overall effects of dosage and treatment duration on MSC chondrogenesis for a given PRP preparation. It is noteworthy that Mazzocca et al. reported that platelet and white blood cell numbers vary between individuals and even within an individual when blood is drawn repetitively over a day. 55 More extensive future studies using a statistically significant number of PRP preparations from different donors are clearly needed to examine and correlate the biological activities with the variables noted above.
Conclusion
In 21-day IFP-ASC chondrogenic pellet cultures, high PRP concentrations delivered over days 7–21 increased catabolic gene expression, decreased chondrogenic gene expression, and decreased matrix proteoglycan deposition, compared to TGF-β+/PRP− controls. Short duration and low concentration PRP treatment (1-day pulse, 1% PRP) resulted in increased cartilage matrix production and collagen type II deposition, compared with longer durations and higher concentrations. Furthermore, in 21-day 3D hydrogel cultures, PRP treatment highly downregulated chondrogenic gene expression and decreased proteoglycan deposition, compared with TGF-β+/PRP− controls. These results showed that high concentration PRP treatment for prolonged period impairs MSC chondrogenesis in vitro. Understanding the effects of PRP on MSC chondrogenesis is necessary to further develop strategies for the application of PRP in combination with cell-based therapy for the repair of focal cartilage defects. For example, autologous MSCs encapsulated in an autologous PRP-containing biocompatible hydrogel may be injected into the defect site, resulting in optimal MSC chondrogenic differentiation and production of cartilage extracellular matrix and the formation of functional neocartilage.
Footnotes
Acknowledgment
The authors thank Dr. Paul Manner, University of Washington, for providing clinical tissue samples, Jian Tan for the assistance in BM-MSC isolation and tri-lineage differentiation, Lynda Guzik for the help in flow cytometry, and Hang Lin for the preparation of mGL and hyaluronic acid. This work is supported, in part, by the U.S. Department of Defense (W81XWH-14-2-0003). J.L. is a recipient of Graduate Fellowship from the International Foundation for Ethical Research (IFER).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.