
Abstract
Anisotropic alginate hydrogels with microchannels can provide a substrate for cotransplanted cells and for axons in the injured spinal cord by physically guiding regenerating axons across a lesion. However, alginate gels alone only result in modest axonal growth responses. To determine whether modification of negatively charged alginate hydrogels with positively charged polyamino acids (poly-
Impact Statement
Axonal bridging across a lesion in the injured spinal cord requires a growth substrate and guidance cues. Using alginate hydrogels with capillary channels we show that poly-
Introduction
S
Among different biomaterials, hydrogels have many intriguing features that can be adjusted to mimic key features of spinal tissue and its extracellular matrix (ECM), including physical (stiffness, elasticity, directionality) and chemical parameters (biodegradability, in situ self-assembly, surface modification). Having a defined three-dimensional (3D) microstructure, an implanted scaffold can provide physical and directional guidance for regenerating axons and endogenous or cografted cells, thereby supporting tissue reorganization. Scaffold integration and fibroglial scarring nonetheless remain substantial hurdles for biomaterial-based approaches.
We and others have previously demonstrated that anisotropic capillary hydrogels made of the heteromeric polysaccharide alginate facilitate linear neurite extension in vitro and rostrocaudal axonal growth in vivo.5–8
Other studies have suggested neuroprotective effects of alginate transplants to the spinal cord.
9
Because alginate itself is—at physiological pH levels—negatively charged, cell attachment/survival as well as neurite outgrowth are limited.
10
Consequently, chemical or electrostatic modification may be essential to generate a more permissive surface. A combination of alginate with positively charged poly-
In addition to the surface charge of hydrogels, ECM, and in particular activation of integrin receptors, plays a crucial role in cell attachment and survival as well as neurite growth.13,14 Given the importance of pathways downstream from integrin receptors in axonal regeneration15,16 and improved axon growth by laminin-derived peptides coupled to biomaterials,17–19 coating of alginates with laminin in addition to neutralization of surface charges might further improve regenerative responses.
Besides biomaterials, cell transplantation either alone or in combination with other axonal growth-promoting approaches has aimed to enhance regeneration and functional recovery after SCI. Some of the cells investigated such as bone marrow stromal cells (BMSCs)20,21 and Schwann cells22,23 do not fully integrate into the spinal cord, but remain separated from the host spinal parenchyma frequently with distinct borders. In contrast, neural stem cell-derived immature neurons or astroglia have the capacity to generate adult CNS parenchyma that might be able to better restore tissue homeostasis. In combination with biomaterials, these cells might be able to improve integration of hydrogel implants and thereby enhance regenerative growth.
Of particular interest are astrocytes due to their homeostatic and supportive character during development.24–26
Astrocytes are marked by a high degree of heterogeneity and plasticity and are essential in many processes in the healthy and injured CNS. Following a CNS insult, reactive astrocytes exhibit molecular, cellular, and functional changes that are regulated by context- and severity-dependent signaling events also occurring during astrocyte differentiation and maturation.27,28 During the initial stages of reactive astrogliosis, the astroglial scar physically restricts the lesion and therefore prevents spreading of neuroinflammation and necrosis, whereas in the chronic phase, it may hinder axonal regrowth.
29
However, networks of intermingled astrocytic protrusions can also serve as physical guidance cues for growing axons around spinal lesion sites30–32
similar to radial glia during cortical or spinal cord development.33,34 Therefore, aligned astrocytes within biomaterials might be able to provide directional growth support. Previous studies have shown that neonatal cortical astrocytes can stably align along a protein pattern on the surface of collagen hydrogels in vitro.
35
In addition, astrocytes can attach in an oriented fashion to anisotropic poly-
The current studies therefore investigated whether modification of alginate-based hydrogels with polyamino acids can mask the negative surface charge of alginate hydrogels, and thereby introduce a permissive biological signal. In addition, we analyzed whether surface-coated alginate-based anisotropic capillary hydrogels in combination with syngeneic neonatal astrocytes provide a growth-permissive cellular substrate for in vivo axonal regeneration. Our results indicate that hydrogels can be stably modified by peptides, enhancing the regrowth of fibers into hydrogel channels in vivo. Grafted GFP-transgenic astrocytes survive within the scaffolds supporting integration into the lesion site and further enhance axonal growth.
Materials and Methods
Fabrication of alginate-based capillary hydrogels
Anisotropic alginate-based hydrogels were manufactured as previously described with slight modifications.5–7,37 Briefly, sodium alginate (Pronova UP MVG; FMC Biopolymer AS d/b/a NovaMatrix, Sandvika, Norway) with a guluronic acid content of 70% and dynamic viscosity of 211 mPa · s (10 g/L, 20°C) was dissolved in purified deionized water (dH2O) at a concentration of 20 g/L and overlaid with 20 mL of an aqueous solution of 1 M Sr(NO3)2. Following gelation at 10°C for at least 36 h, hydrogel blocks were washed in dH2O, chemically stabilized with the crosslinker hexamethylene di-isocyanate, immersed in acidic solution to remove complexed Sr2+ ions, and finally neutralized in dH2O. In vitro stability was tested by incubation of cylindrical hydrogel specimens (5 mm diameter, 2 mm thick) in phosphate-buffered saline (PBS) at 37°C for up to 12 weeks. No disintegration of the alginate polymer or the capillary structure could be observed. Hydrogel homogeneity was confirmed by light microscope analysis after ruthenium red staining of sections cut on a microtome (VT1000S; Leica Biosystems, Nussloch, Germany).
For cell culture experiments, capillary-free hydrogels were cut into slices (1 cm × 1 cm, 300 μm thick) using a microtome. For in vivo studies, cuboid implants (2 mm height, 2 mm length, 1.3 mm width) with anisotropic capillaries parallel to the 2-mm-long edge were cut. Homogeneity of the gel matrix as well as shape and orientation of the capillaries were confirmed by light microscopy for each implant (4 × objective, BX53; Olympus) and before implantation using a surgical stereomicroscope as previously described.7,8 Hydrogels were stored in 70% ethanol (EtOH) at 4°C and washed 3 × (1 h each) in sterile dH2O before further use. Two different batches of hydrogels were used in in vivo experiments evaluating the effect of peptide coating and astrocyte seeding with channel diameters of 39 ± 1.6 μm and 50.2 ± 2.1 μm, respectively. Analysis of the second batch showed a mean density of 89 channels/mm 2 and a free channel volume of 20%. The different channel diameters used in the two sets of experiments did not influence outcomes (see section “Results”: similar number of neurite growth into channels coated with PLO/laminin).
Surface coating of alginate-based hydrogels
Poly-
Isolation of neonatal cortical astrocytes from Fischer 344 rats
Animal experiments were carried out in accordance with German guidelines for animal care and in accordance with the European Union Directive (2010/63/EU) or approved by the Institutional Animal Care and Use Committee. Animals were anesthetized by intraperitoneal injection (5 mL/kg) of a mixture of ketamine (62.5 mg/kg; Bremer Pharma, Warburg, Germany), xylazine (3.175 mg/kg; CP-Pharma, Burgdorf, Germany), and acepromazine (0.625 mg/kg; Sanofi-Ceva GmbH, Düsseldorf, Germany) in 0.9% (v/w) NaCl and killed by decapitation.
Astrocytes were isolated from cortices of neonatal (P1) GFP-transgenic Fischer 344 (F344) rats. Following brain dissection, hemispheres were separated and transferred into 30-mm cell culture dishes containing 1 mL ice-cold Dulbecco's modified eagle medium (DMEM) (Life Technologies). Excessive noncortical brain tissue, meninges, and blood vessels were carefully removed from cortical tissue under a dissection microscope. Cortices were mechanically dissociated using thin forceps and enzymatically digested by adding 500 μL of a 1:1 mixture of Collagenase XI (1200 U/mg; Sigma-Aldrich, in Hank's-buffered salt solution (HBSS; Life Technologies)) and Dispase I (4 U/mg in dH2O; Worthington Biochemical Corp., Lakewood, NJ) for 45 min at 37°C, 5% CO2 under permanent mechanical agitation (125 rpm). Afterward, cells were further dissociated by adding 1 mL prewarmed culture medium (DMEM/5% fetal bovine serum [FBS]/0.5% penicillin/streptomycin [10,000 U/mL each]/1%
Two days later, cells were washed once with DPBS to remove cellular debris and nonadherent cells. Medium was changed 3 × a week until cells reached confluency. To remove contaminating cells like neurons, microglia, and fibroblasts, 150 μL of a 10 mg/mL β-
Dorsal root ganglias (DRGs) were isolated from adult GFP-transgenic F344 rats. Animals were anesthetized and killed as described above. Spinal columns were dissected and cut at the rostral and caudal end to expose the vertebral canal. The spinal cord was ejected by injecting 20 mL ice-cold HBSS into the caudal end of the vertebral canal with a 10-mL syringe. The vertebral column was cut in half longitudinally along the rostrocaudal axis and DRGs were carefully dissected by pulling the ganglia out of the bone cavity and cutting off the nerve roots. DRGs were cut in half and collected in ice-cold Hibernate A medium (Invitrogen, Carlsbad, CA). Afterward, DRGs were washed once in ice-cold HBSS and digested by incubation in 500 μL of a 1:1 mixture of Collagenase XI (1200 U/mg in HBSS) and Dispase I (4 U/mg in dH2O) at 37°C for 30 min. Cells were washed twice with 1 × DMEM/F12 (Life Technologies) with 10% FBS and mechanically dissociated by triturating with a flame-polished glass Pasteur pipette about 15 times. Pieces of meninges and tissue debris were allowed to sink and cells were immediately used for in vitro experiments.
Quantification of coating efficiency
For in vitro experiments, neonatal astrocytes of passage 2 and freshly isolated DRG cells were used. For short-term experiments, hydrogel slices were placed into six-well cell culture plates (Greiner Bio-One) containing 4 mL prewarmed culture medium (see above). In total, 4 × 10 4 astrocytes (passage 2) were plated onto uncoated control gels (n = 7), gels with PLO (n = 6) or PLO/laminin coating (n = 6). Medium was carefully changed after 2 and 4 days. After 7 days, cells were fixed with 4% ice-cold paraformaldehyde (PFA) in 0.1 M phosphate buffer (PB), pH 7.4 for 30 min and washed 3 × 10 min with 0.1 M tris-buffered saline (TBS). DRG-derived cells were seeded on uncoated control gels (n = 7) and gels with PLO (n = 6) or PLO/laminin (n = 6) coating and fixed with 4% ice-cold PFA in 0.1 M PB, pH 7.4, after 48 h.
To assess long-term bioactivity of the surface-bound proteins, PLO or PLO/laminin-coated hydrogels were incubated for 24 h in 10 mL DRG cell culture medium (DMEM/F12/2% B27 supplement/0.5% penicillin/streptomycin [10,000 U/mL each]/1%
For immunolabeling, hydrogel slices were carefully transferred into 24-well cell culture plates (Greiner Bio-One) using blunt forceps. Unspecific antibody-binding sites were blocked and the tissue permeabilized with TBS/5% donkey serum/0.25% Triton X-100 for 1 h at room temperature (RT). Primary antibodies diluted in TBS/1% donkey serum/0.25% Triton X-100 were added overnight at 4°C. Afterward, samples were washed 3 × 10 min in TBS/1% donkey serum, incubated with Alexa-fluorophore-conjugated donkey secondary antibodies (1:300; Life Technologies) together with 4′, 6-diamidino-2-phenylindole (DAPI, 0.25 μg/mL; Thermo Fisher Scientific) in TBS/1% donkey serum for 2.5 h at RT followed by three washing steps with TBS. Finally, hydrogels were mounted onto microscope glass slides and sealed with Fluoromount G (Southern Biotechnology, Birmingham, AL). The following primary antibodies were used: goat anti-GFP for GFP-transgenic cells (1:1000; Rockland, Limerick, PA), rabbit anti-GFAP for astrocytes (1:1000; Dako), and mouse anti-β-III-tubulin for neurons (1:1000; Promega). Cell densities of all cultures and neuron density of DRG cultures were assessed by counting DAPI-labeled nuclei or β-III-tubulin-labeled neurons in a randomly selected field of 1 mm 2 at 100 × magnification on a fluorescence microscope (Olympus BX53) on each gel. Neurite outgrowth was assessed by manually tracing the longest β-III-tubulin-labeled neurite of each neuron using the NeuronJ plug-in for ImageJ. All analyses were carried out by an observer blinded to group identity.
To analyze whether differences in cell adhesion underlie the observed differences in cell numbers, postnatal GFP-transgenic astrocytes (1.5 × 10 4 ) were seeded on 7 × 7 mm uncoated, PLO- or PLO/laminin-coated 300 μm thick hydrogel slices after three washes in PBS and fixed after 3 h (n = 3 each). After immunolabeling for GFP, slices were imaged at 200 × magnification using an inverted epifluorescent microscope (Olympus; CKX53) with attached camera (Infinity 3-1URM) and the cell number was quantified in five randomly selected fields/slice using ImageJ. To determine whether differences in cell death contribute to differences in cell numbers, wild-type astrocytes seeded on hydrogels were cultivated for 2 days and stained with a Live/Dead Cell Assay Kit (1 μM calcein AM/4 μM nuclear dye; Fisher Scientific) according to the manufacturer's instructions. Cells were immediately imaged and live (green) and dead (red) cells were counted. Assays were done in triplicate.
To analyze the distribution of laminin coating, uncoated, PLO-coated and PLO/laminin-coated gel cuboids (1.3 mm × 2 mm × 2 mm) were washed, fixed with 4% PFA, and immunolabeled for laminin (1:300; Sigma, Alexa 488-conjugated secondary antibody; 1:1000). Images were taken on an inverted fluorescent microscope (Olympus; CKX53) with attached camera (Infinity 3-1URM) using the same exposure time for all images at the same magnification. In addition, potential differences of concentration-dependent biological effects of laminin at the edge and center in PLO/laminin-coated hydrogel cuboids were analyzed. Freshly PLO/laminin-coated and uncoated unfixed hydrogel cuboids (n = 2) were cut with a tissue chopper (McBain) (300 μm thickness), the outermost slices were discarded and GFP+ astrocytes (2 × 10 3 cells) were seeded immediately onto the hydrogel slices (5–6 slices) in cell culture medium. After 48 h, hydrogels and attached cells were fixed, immunolabeled for GFP, and the size of cells found at the indicated distance from the edge (100–1000 μm) of the hydrogel was measured. Data are represented as the mean of all cells measured at a certain distance.
Surgical procedures
A total of 40 adult female F344 rats (12–14 weeks, >150g; Charles River) were used for in vivo studies. National guidelines and the European Union Directive (EU/2010/63) were strictly followed for all surgical procedures and animal care. Animals were housed in standard cages with free access to water and food. Animals underwent a C5/6 lateral hemisection lesion as previously described.7,8 Briefly, animals were anesthetized with a mixture (2.5 mL/kg) of ketamine (62.5 mg/kg), xylazine (3.175 mg/kg), and acepromazine (0.625 mg/kg) diluted in sterile 0.9% NaCl. A laminectomy at cervical level 5 (C5) was performed. For lesioning, a rostrocaudal incision was made into the dura along the midline and a block of spinal cord tissue (∼2 mm in length) was removed unilaterally using microscissors and microaspiration without further damaging the dura overlying the lesion cavity. Hydrogel scaffolds were implanted immediately postinjury into the lesion cavity using a surgical microscope to ensure that the channels were oriented parallel to the rostrocaudal axis of the spinal cord. Care was taken to minimize any deformation of the hydrogel during implantation.
To evaluate the impact of peptide/protein coating in Experiment 1, a total of 24 animals received either uncoated control hydrogels (n = 9) or hydrogels coated with PLO/laminin (n = 15) immediately after lesioning. To evaluate the effect of astrocyte seeding into hydrogel channels, a total of 10 animals received PLO/laminin-coated scaffolds seeded with neonatal GFP+ astrocytes, whereas PLO/laminin-coated scaffolds without cells served as control (n = 6).
The dura was covered with a thin dried agarose film (1% UltraPure™ agarose in sterile dH2O; Life Technologies) and sealed with fibrin glue (2 μL fibrinogen, 100 mg/mL and 2 μL thrombin, 400 U/mL; Sigma-Aldrich). Muscle layers were sutured and the skin was stapled. Postoperatively, animals received subcutaneous injections of buprenorphine-HCl (0.03–0.05 mg/kg; Reckitt Benckiser, Slough, United Kingdom) twice a day for 2 days for pain relief and ampicillin (50 mg/kg; Ratiopharm GmbH, Ulm, Germany) as needed. Carprofen (4–5 mg/kg; Pfizer, Inc., New York, NY) was administered subsequently once daily as needed. Animals were fed with 2 mL high-energy nutrition drink (Fresubin; Fresenius Kabi) up to three times per day until their body weight stabilized.
Tissue processing
Four weeks postimplantation, rats were deeply anesthetized (see above) and transcardially perfused with 0.1 M PBS followed by 4% PFA in 0.1 M PB, at pH 7.4. Spinal cords were carefully dissected and subsequently postfixed in 4% PFA/0.1 M PB at RT for 1 h. Tissue was cryoprotected in 30% sucrose/0.1 M PB at 4°C for 2 days and serially cut into 30-μm-thick horizontal sections on a cryostat (Leica Biosystems). Sections were directly mounted onto glass slides (Thermo Fisher Scientific), with every 14th section placed on the same slide. For immunohistochemistry, tissue sections were labeled as described above using DAPI (0.25 μg/mL), rabbit anti-GFAP for astroglial cells (1:1000; Dako), goat anti-rabbit GFP for cells isolated from GFP-transgenic rats (1:1000; Rockland), mouse anti-βIII-tubulin for axons (1:1000; Promega), rabbit anti-serotonin for descending raphespinal axons (5-HT; 1:2000; Immunostar), rabbit anti-von Willebrand factor for endothelial cells and blood vessels (1:1000; Sigma-Aldrich), and Alexa-fluorophore-conjugated donkey secondary antibodies (Alexa-488, Alexa 594; 1:300; Life Technologies). Sections were dried and coverslipped with Fluoromount G (Southern Biotech). Sections were imaged using a XC30 camera connected to a BX53 microscope (Olympus) or a FluoView1000 confocal microscope (BX61; Olympus).
Quantification of cell-filled channels, axon number, astroglia, and microglia
To quantify host cell migration into the scaffolds, the percentage of channels that were densely filled with DAPI+ nuclei was quantified at a virtual line perpendicular to the rostrocaudal orientation of channels, 100 and 500 μm from the rostral and caudal edge of the hydrogel in every seventh section at 400 × magnification under epifluorescent illumination (Olympus; BX53). Rostral and caudal areas were quantified separately as there is independent axon penetration from both sides. Distances of 100 and 500 μm were chosen to represent growth over short distances and more central location in the hydrogel channels, respectively. The same distances were measured in our previous studies making the current data comparable.7,8 For astrocyte-seeded scaffolds, survival of grafted GFP-transgenic astrocytes was assessed by quantifying the percentage of DAPI+/GFP+ channels at 100 and 500 μm from the rostral and caudal edge of the hydrogel at 400 × magnification. Channels containing only sparse, isolated DAPI-labeled nuclei and/or GFP-labeled cells were not considered positive. In addition, the total channel area filled with GFP-labeled cells was analyzed in the rostral half of the hydrogel (0–500 and 500–1000 μm) at 100 × magnification in all animals implanted with astrocyte-filled channels.
Similarly, β-III-tubulin-labeled as well as 5-HT-labeled axons within scaffold channels were analyzed using epifluorescent illumination at 200 × magnification. In all channels, neurites crossing virtual lines perpendicular to the channel axis at 100 and 500 μm from the rostral and caudal edge of the hydrogel were counted in every seventh section. For each animal, mean values from 100 and 500 μm of the rostral and caudal site were averaged for β-III-tubulin-labeled axons and analyzed separately for 5-HT-labeled axons. Results of all animals within each treatment group at each location (uncoated vs. PLO/laminin coating or PLO/laminin coating vs. PLO/laminin coating with neonatal astrocytes) were averaged for comparisons. Channel and axon quantification was performed while blinded to group identity.
The number of axons per mm
2
hydrogel was calculated using the following equation:
For quantification of GFAP and Iba1 labeling density, images of fluorescent-labeled serial spinal cord sections, rostral and caudal, to the hydrogel implantation site and of the uninjured contralateral side were taken at 200 × magnification (250 × 250 μm). Using a threshold that represents the labeling density on the uninjured side, the percent labeling density was quantified in animals that received uncoated and PLO/laminin-coated hydrogels.
Statistical analysis
All data were analyzed by an observer blinded to group identity. Data are expressed as mean ± standard error of the mean. Statistical analysis was performed using Prism 6 software (GraphPad Software, Inc., La Jolla, CA) and a significance criterion of p < 0.05 was used. Groups were compared by one-way analysis of variance (ANOVA) followed by Tukey's post hoc test or two-way ANOVA followed by Sidak's post hoc tests.
Results
Coating efficiency of differently charged polypeptides
Cell adhesion and biocompatibility of biomaterials are influenced by surface charges and the presence of ECM. To determine whether peptide/ECM coating would affect cell survival/adhesion and neurite growth in alginate scaffolds, we first explored whether polyamino acids and peptides can be stably bound to alginate-based hydrogels. The linear polysaccharide alginate consists of homopolymeric as well as alternating blocks of α-
Cell adhesion and viability on surface-coated alginate-based hydrogels in vitro
To examine the influence of polyamino acid and peptide coating on cell attachment and survival, PLO or PLO in combination with the ECM component laminin were bound to 300-μm-thick hydrogel slices, whereas uncoated hydrogels served as controls (Fig. 1). Neural cells (astrocytes or DRG neurons with satellite cells) were seeded immediately following coating and short washes of the hydrogel slices. Seven days after cell seeding, the number of GFP-transgenic neonatal cortical astrocytes was 10-fold higher on coated versus uncoated hydrogels (Fig. 1E–H; p < 0.0001), although laminin coating did not provide any additional effect (Fig. 1H). Notably, in addition to differences in cell number, clear differences in cell morphology were observed in response to coating. On uncoated and PLO-coated hydrogels, neonatal astrocytes were predominantly rounded with small somata bearing no processes. With additional laminin coating, astrocytes had larger somata and developed a bi- or tripolar morphology with longer processes. Similarly, adult DRG neurons and DRG-derived glia showed pronounced cell numbers on surface-coated hydrogels (Fig. 1I–L; p < 0.0001). Neurite growth was virtually absent on uncoated controls and only sporadically observed on PLO-coated hydrogels, whereas PLO/laminin coating elicited substantial neurite outgrowth (quantified in Fig. 2). Cells did not invade the hydrogel walls but were only found on the surface. To determine whether cell attachment and/or survival were influenced by PLO/laminin coating, astrocytes were fixed 3 h or 2 days after cell seeding. Quantification of cell numbers (GFP+ cells) shortly after seeding indicated a significantly higher number of cells attaching to PLO/laminin-coated gels (Fig. 1I). In addition, a smaller percentage of dead cells was detected on PLO/laminin-coated hydrogel slices after 2 days of cultivation, although this difference did not reach significance (Fig. 1J; ANOVA p = 0.28). Thus, binding of PLO and especially the combination of PLO and laminin to alginate hydrogels improves cell attachment and to a smaller degree survival, and increases neurite extension.
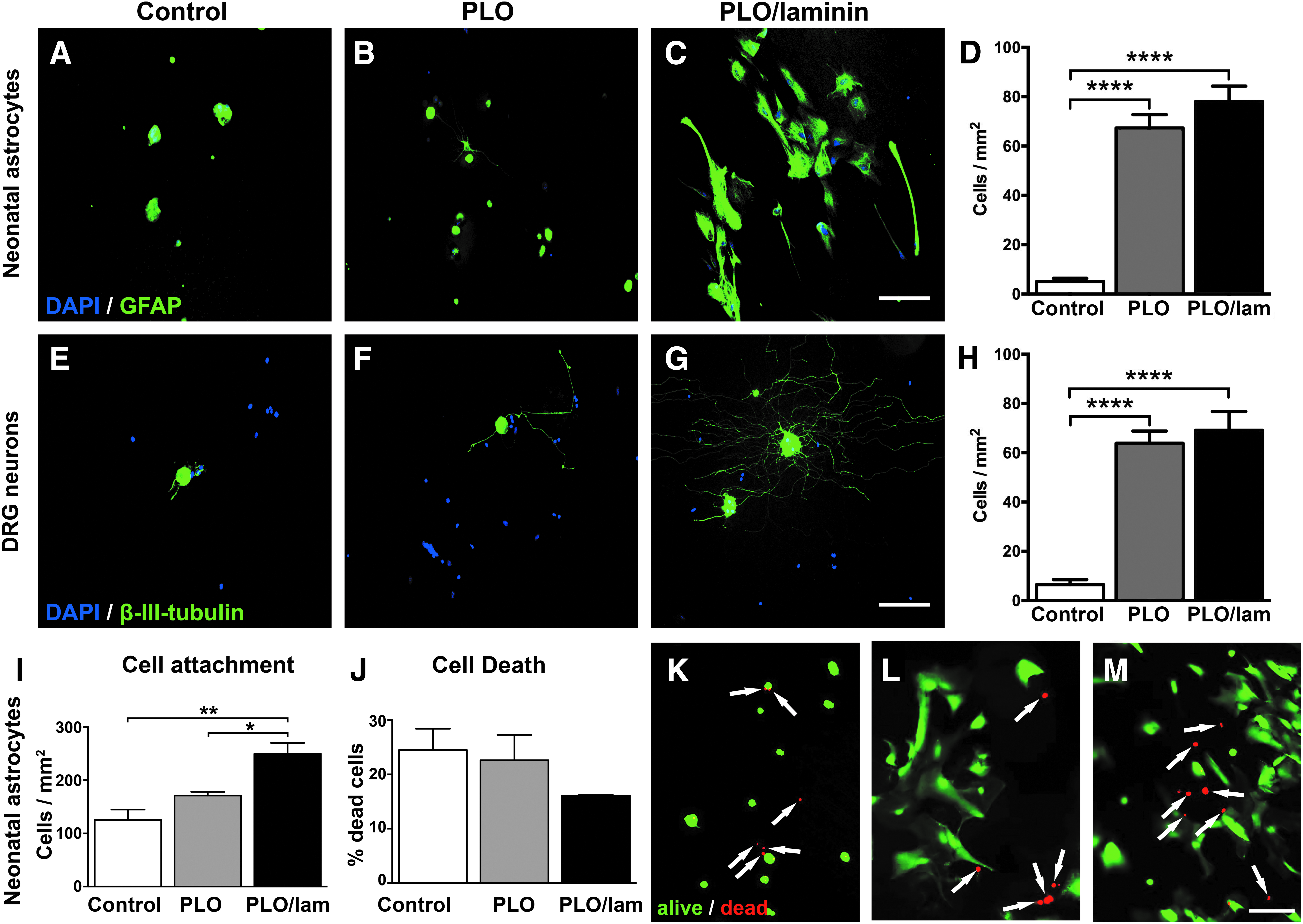
Cell adhesion and viability on surface-modified alginate-based hydrogels.
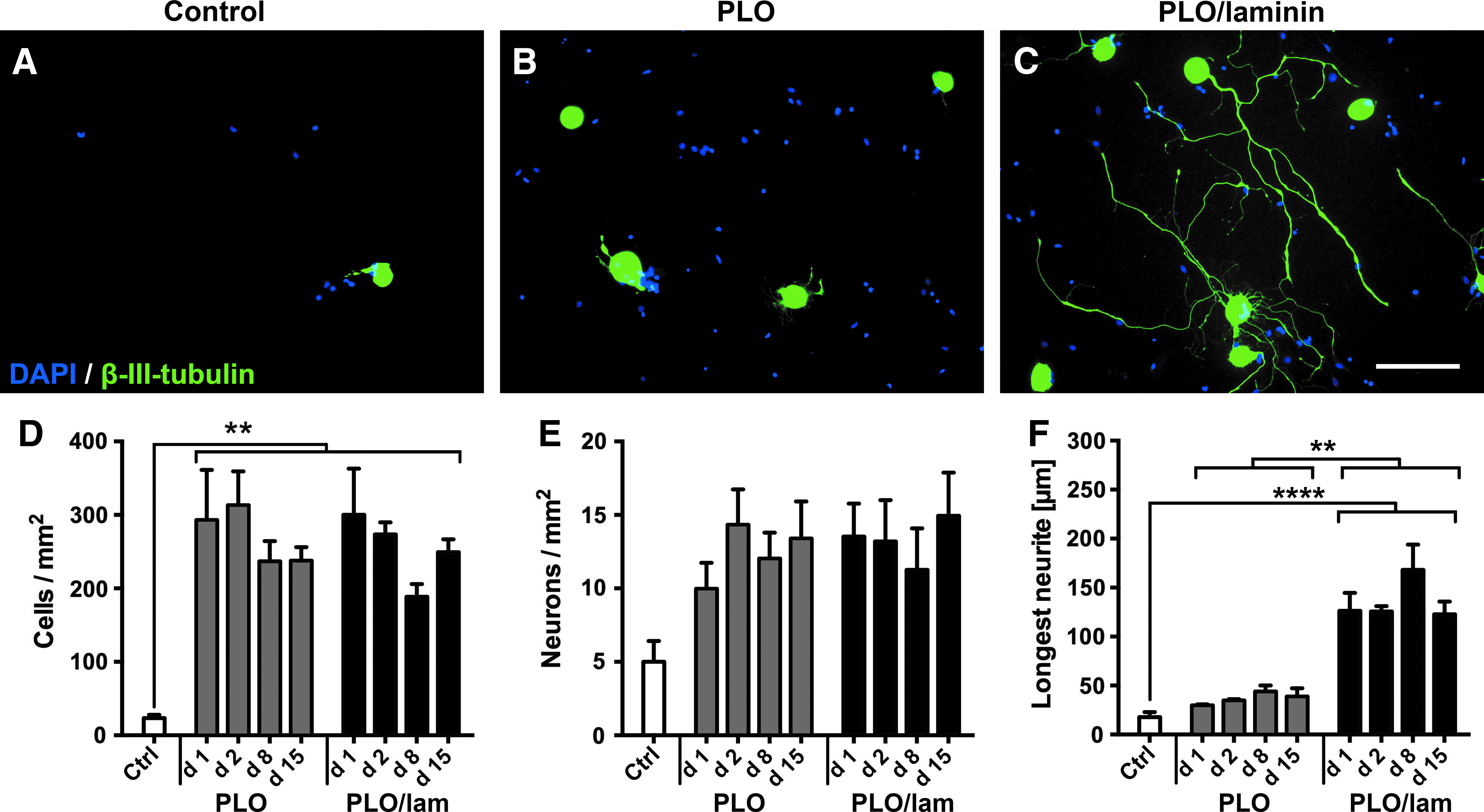
Long-term stability of alginate polypeptide/protein coating agents.
Long-term retention and bioactivity of surface-coated proteins in vitro
In vivo experiments with surface-coated scaffolds require stability and bioactivity of peptide/protein coating for longer periods than the short-term experiments described above. We therefore examined alginate hydrogels that were coated with PLO or PLO/laminin and incubated under physiological conditions (37°C, 5% CO2 in DPBS) up to 15 days, including repeated daily washing steps before cells isolated from adult rat DRGs were plated onto the hydrogels (Fig. 2A–C). At all time points, the number of cells (DAPI+ nuclei) attached to PLO-coated and PLO/laminin-coated hydrogels was considerably higher than the number of cells found on uncoated controls (Fig. 2D). This was also true for β-III-tubulin-labeled neurons, but these differences did not reach statistical significance (ANOVA p = 0.076; Fig. 2E). Importantly, there was no significant difference in the number of attached cells that were seeded 1 or 15 days after hydrogel coating. In addition to cell attachment and survival, surface coating also affected DRG neurite outgrowth. When measuring the longest neurite 48 h after cell seeding (Fig. 2F), a slight increase in neurite length was apparent on PLO-coated hydrogels. The addition of laminin dramatically increased the average neurite length a further three to four-fold (ANOVA p < 0.0001, Tukey's post hoc test p < 0.01 comparing PLO to PLO/laminin; p < 0.0001 comparing control to PLO/laminin). Again, this effect was independent of the time that hydrogels were used to after coating. Thus, surface modification with PLO alone or in combination with laminin is stable retaining full bioactivity for at least 2 weeks in vitro.
Integration and host cell infiltration of surface-coated hydrogel scaffolds in vivo
For in vivo experiments, alginate hydrogel cubes were coated with PLO/laminin. Immunolabeling for laminin indicated higher immunoreactivity at the hydrogel edges, but also a distinct signal along the hydrogel channels that was still present at the center of the hydrogel (Supplementary Fig. S3A–D). To determine whether the laminin detected in the center of the gel was sufficient for biological effects, uncoated and PLO/laminin-coated hydrogel cubes (1.3 mm × 2 mm × 2 mm) were freshly sectioned and seeded with GFP+ astrocytes. Quantification of astroglial cell size at different distances from the edge of the gel indicated an overall increased cell size by PLO/laminin coating compared with uncoated controls with no difference across different distances from the edge. Taken together, the concentration gradient observed by immunolabeling does not appear to be biologically relevant at least not for the tested cells.
To characterize cellular responses in vivo, PLO/laminin-coated alginate hydrogels, which induced superior neurite growth in vitro, or uncoated hydrogels (control) were implanted into a unilateral C5/6 spinal hemisection immediately postinjury. Four weeks postimplantation, scaffolds showed no signs of degradation, remained in rostrocaudal orientation, and the capillary 3D microarchitecture remained intact (Fig. 3), consistent with previous studies.7,8 Implants were tightly integrated into the surrounding host parenchyma without excessive hypercellularity or cavitation.
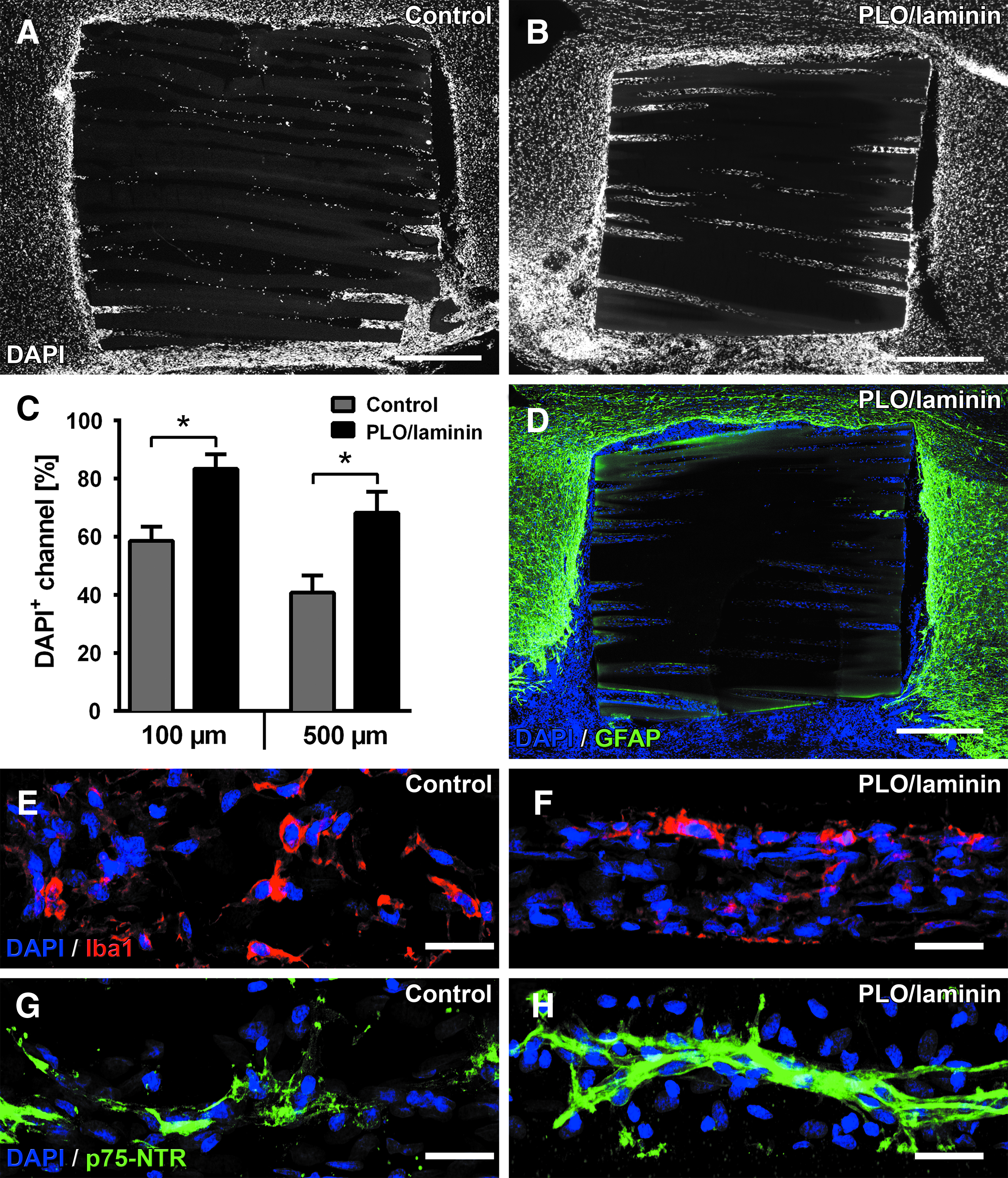
Integration and host cell infiltration into hydrogel channels. After 4 weeks, hydrogel scaffolds remain structurally intact and integrate tightly into the lesion site without excessive cavitation. DAPI labeling of nuclei (white) reveals
Nuclear staining with DAPI indicated host cell migration toward and infiltration into the hydrogel channels but not into the channel walls (Fig. 3A, B). Quantification of DAPI+ nuclei in hydrogel areas close to host tissue (100 μm) as well as at more central regions (500 μm) of the channels revealed a significantly higher percentage of cell filling in PLO/laminin-coated scaffolds compared with uncoated controls (p < 0.05; Fig. 3C). While infiltrating cells densely clustered at the channel entrances, they were sparser at central regions of control scaffolds. In contrast, host cells filled broad portions of the channel lumen in surface-coated scaffolds. A small area of hypercellularity was present at the host–graft interface surrounded by GFAP+ astrocytes independent of coating conditions (Fig. 3D). Interestingly, GFAP-labeled host astrocytes rarely infiltrated the host–graft interface and did not extend processes into the channels. Within the hydrogel scaffolds, immunolabeling identified the vast majority of infiltrating cells in both groups as Iba1+ cells (microglia/macrophages) (Fig. 3E, F) and some Schwann cells (Fig. 3G, H). To exclude the possibility that the increased cell filling is due to increased glial activation by PLO/laminin, microglial and astroglial labeling was also measured in areas surrounding the scaffolds. No difference in GFAP and Iba1-labeling density was observed between both groups (Supplementary Fig. S4).
In vivo neurite growth into channels of coated hydrogels
To evaluate direct or indirect influences of surface coating on host axonal growth in vivo, the number of β-III-tubulin-labeled neurites entering and extending within the hydrogel channels was quantified (Fig. 4). In control as well as in PLO/laminin-coated scaffolds, axons entered the scaffolds from both the rostral and caudal host parenchyma (Fig. 4A, B). Hardly any neurites were found in channels without infiltrating host cells and the number of neurites per cell-filled (DAPI+) channel did not differ between implants coated with PLO/laminin and uncoated controls (Fig. 4C) at channel entrances (100 μm) or inside the hydrogel implant (500 μm). In both groups, fewer neurites per cell-filled channel were found with increasing distance from the host–graft interface and ∼50% less neurite growth was found within the central regions of the implants.
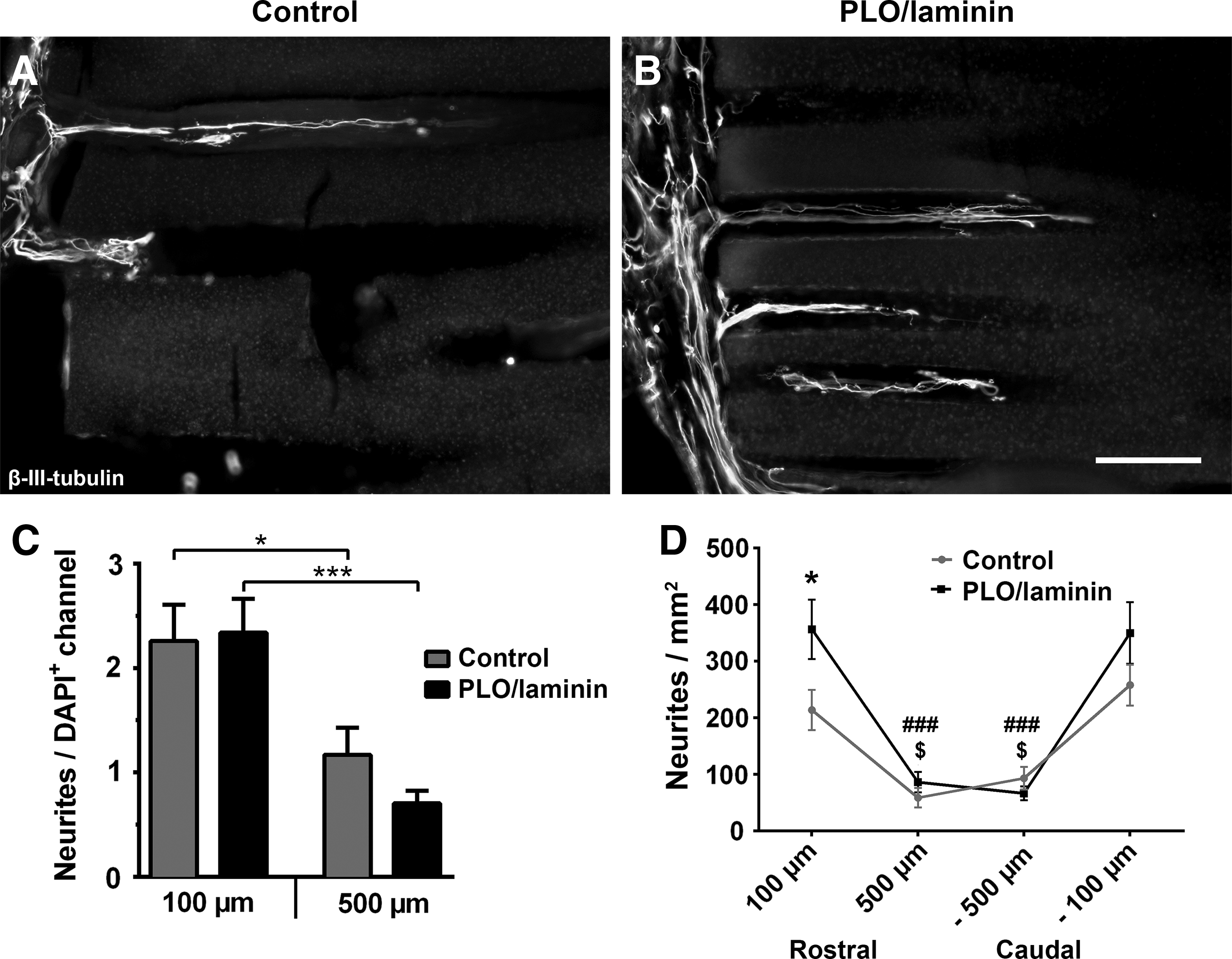
Neurite growth into microchannels of coated hydrogel scaffolds.
However, when examining the total number of neurites, significantly more neurites extended into PLO/laminin-coated scaffolds (Fig. 4D) at the rostral entry zones (p < 0.05) consistent with the higher number of cell-filled channels in this group (Fig. 3). Taken together, these data suggest that PLO/laminin coating promotes significantly higher host cell infiltration and thereby more neurite penetration into the hydrogels, but this is insufficient to sustain axonal growth for longer distances into or across the channels.
Seeding of neonatal astrocytes into surface-coated hydrogels
Even in alginate hydrogels coated with PLO/laminin, virtually no astrocytes or astrocytic processes were detected within the channels (Fig. 3D). To assess whether astroglia would provide a superior cellular substrate compared with infiltrating host cells, improve scaffold integration and increase axonal regeneration within the scaffold, we seeded syngeneic cortical astrocytes isolated from neonatal (P1) GFP-transgenic F344 rats into the channels of PLO/laminin-coated hydrogels immediately before implantation. Cell-filled scaffolds and PLO/laminin-coated scaffolds without cells as control were implanted immediately after lesioning the spinal cord. Inspection of scaffolds before transplantation showed an even distribution of astrocytes in cell-filled channels (Supplementary Fig. S5). Four weeks postinjury, scaffolds of both groups remained intact, were oriented rostrocaudally, and tightly integrated, particularly scaffolds seeded with astrocytes (Figs. 5A, B, and 6).
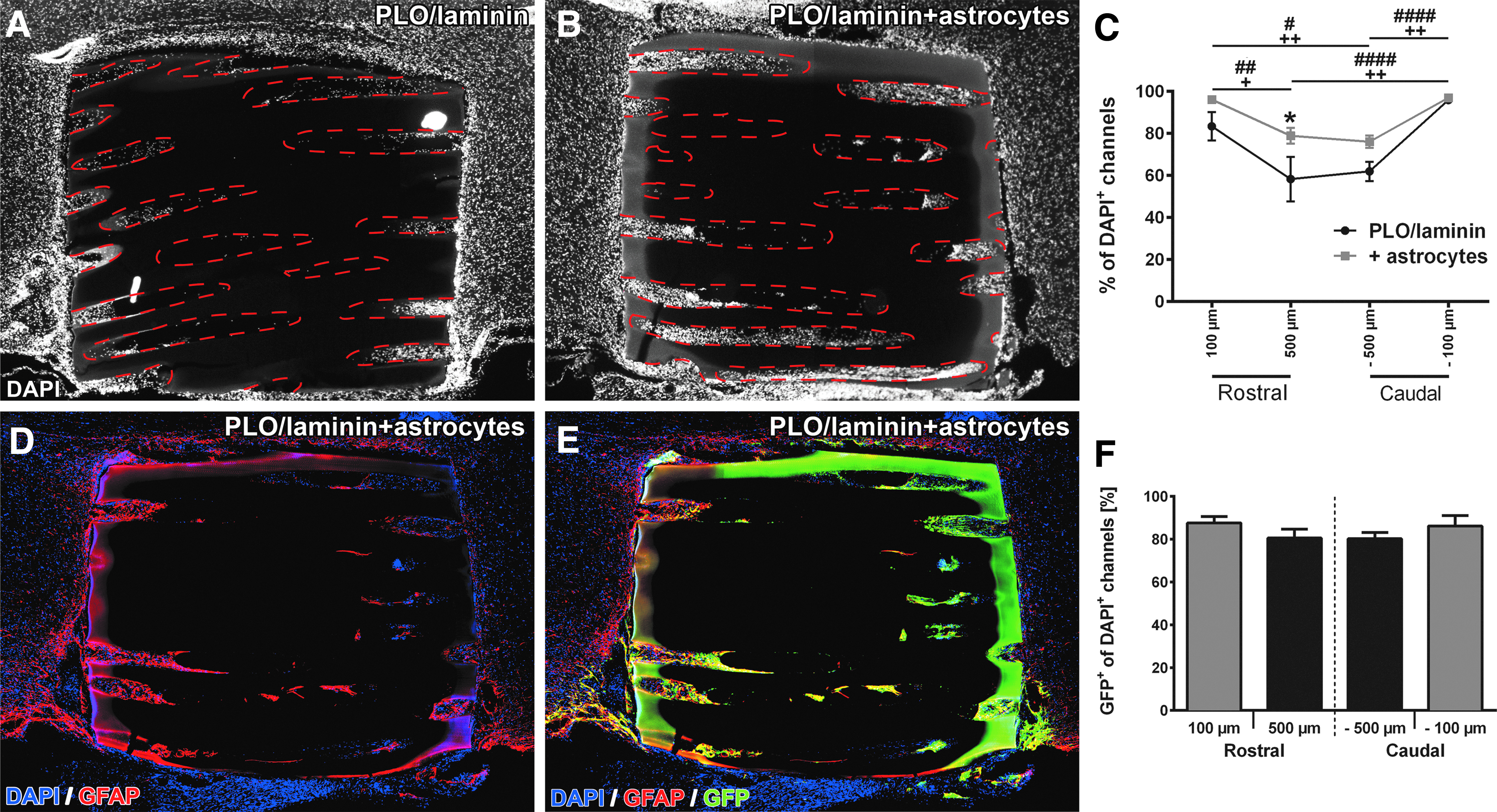
Host cell infiltration and survival of grafted neonatal astrocytes. Scaffolds in both groups remain intact and are tightly integrated into the host parenchyma 4 weeks postgrafting. DAPI-labeled nuclei (white) show host cell infiltration into
Integration and host–graft interaction of astrocyte-seeded hydrogel scaffolds. At the
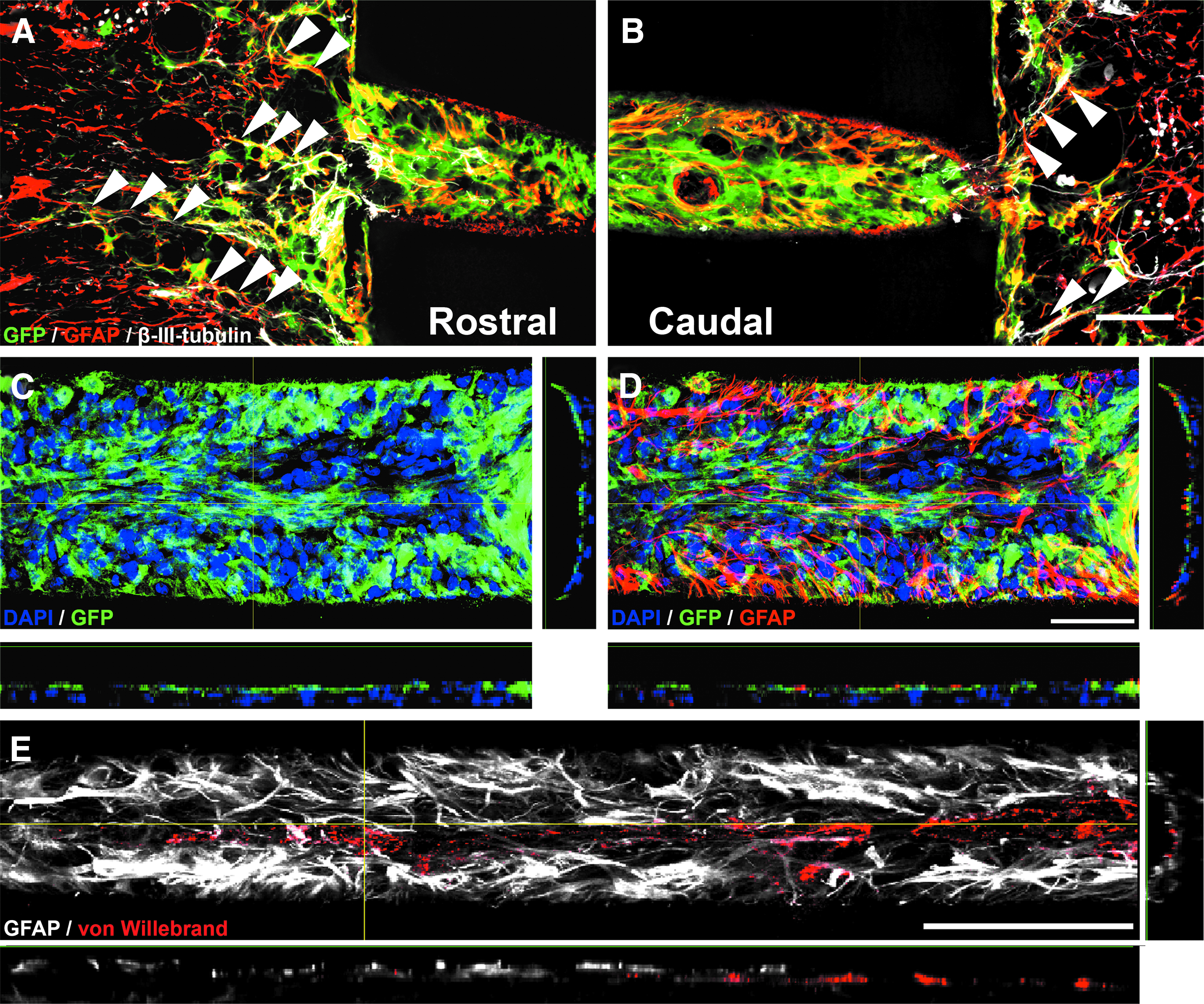
Quantification of DAPI-labeled nuclei indicated a decline in cell-filled channels at the central locations in both groups (Fig. 5C, two-way ANOVA p < 0.0001 for distance) confirming our data from the first set of experiments. In addition, an overall higher percentage of cell-filled channels was apparent in hydrogels seeded with astrocytes in particular at the central locations (Fig. 5C; two-way ANOVA p < 0.001 for group differences; p < 0.05 for 500 μm distance). Assessing survival of grafted cells (percentage of GFP+ cell-filled channels), the vast majority of cell-filled channels contained GFP+ astrocytes (Figs. 5D–F and 6C, D). At all distances, 80–85% of all cell-filled channels showed GFP labeling. At the graft–host interface, GFP+/GFAP+ double-labeled cells filled the capillary lumen in a dense network-like fashion (Fig. 6A, B) and grafted cells did not migrate extensively into the host parenchyma. However, grafted astrocytes filled the host–graft interface and their processes were tight-knit with processes of host astrocytes serving as a cellular substrate for host axons extending into the channels (Fig. 6A, B, arrowheads). Toward the hydrogel center, the density of the GFP+/GFAP+ network declined, and GFP+ cells were occasionally organized in clusters (Fig. 5E). The total GFP+ channel area was 76.3 ± 3.2% in the rostral portion of the alginate gel (0–500 μm) and slightly declined to 53.7 ± 3.1% in the more central portion of the hydrogel (500–1000 μm; unpaired t-test p < 0.001). Endothelial cells and possibly thin blood vessels also entered and extended along astrocyte-containing channels (Fig. 6E). Besides grafted astrocytes and endothelial cells, more than 80% of all GFP+ channels displayed immunoreactivity for Iba1 identifying GFP-negative cells mainly as macrophages/microglia (data not shown) similar to our previous observations (Fig. 3).
Neurite growth into astrocyte-seeded capillary hydrogels
To analyze axonal growth responses, β-III-tubulin-labeled fibers were quantified at a distance of 100 and 500 μm from the rostral and caudal edge of PLO/laminin-coated hydrogels and PLO/laminin-coated hydrogels filled with astrocytes (Fig. 7). Overall, more neurites per DAPI+ channel were present in animals implanted with astrocyte-containing hydrogels, but this difference did not reach significance (Fig. 7C). As 10–20% of channels in animals that received astrocyte-filled scaffolds only contained host cells (DAPI+/GFP−; Fig. 5), we were able to determine whether extending neurites preferred astrocyte-containing channels versus channels containing only host cells within the same alginate scaffold. Interestingly, in animals grafted with astrocyte-seeded scaffolds, the number of neurites/channel was much higher in channels that contained astrocytes (DAPI+/GFP+) compared with cell-filled channels devoid of astrocytes (DAPI+/GFP−). More than 86% of all axons in these animals were found in channels containing GFP+ astrocytes. This was true for all distances examined indicating that astrocytes generated a preferable environment for regenerating axons. In animals with and without astrocyte seeding, neurite counts per channel declined from the rostral to the central region of the scaffold (Fig. 7C). Quantifying the entire channel lumen at the same distances, neurite density was overall higher in astrocyte-filled scaffolds (two-way ANOVA p < 0.001 for group differences), but this difference only reached significance at 100 μm within the rostral half of the scaffold 4 weeks postimplantation (Fig. 7D). Again, a decline in the number of neurites from rostral and caudal ends toward the center was found in both groups.
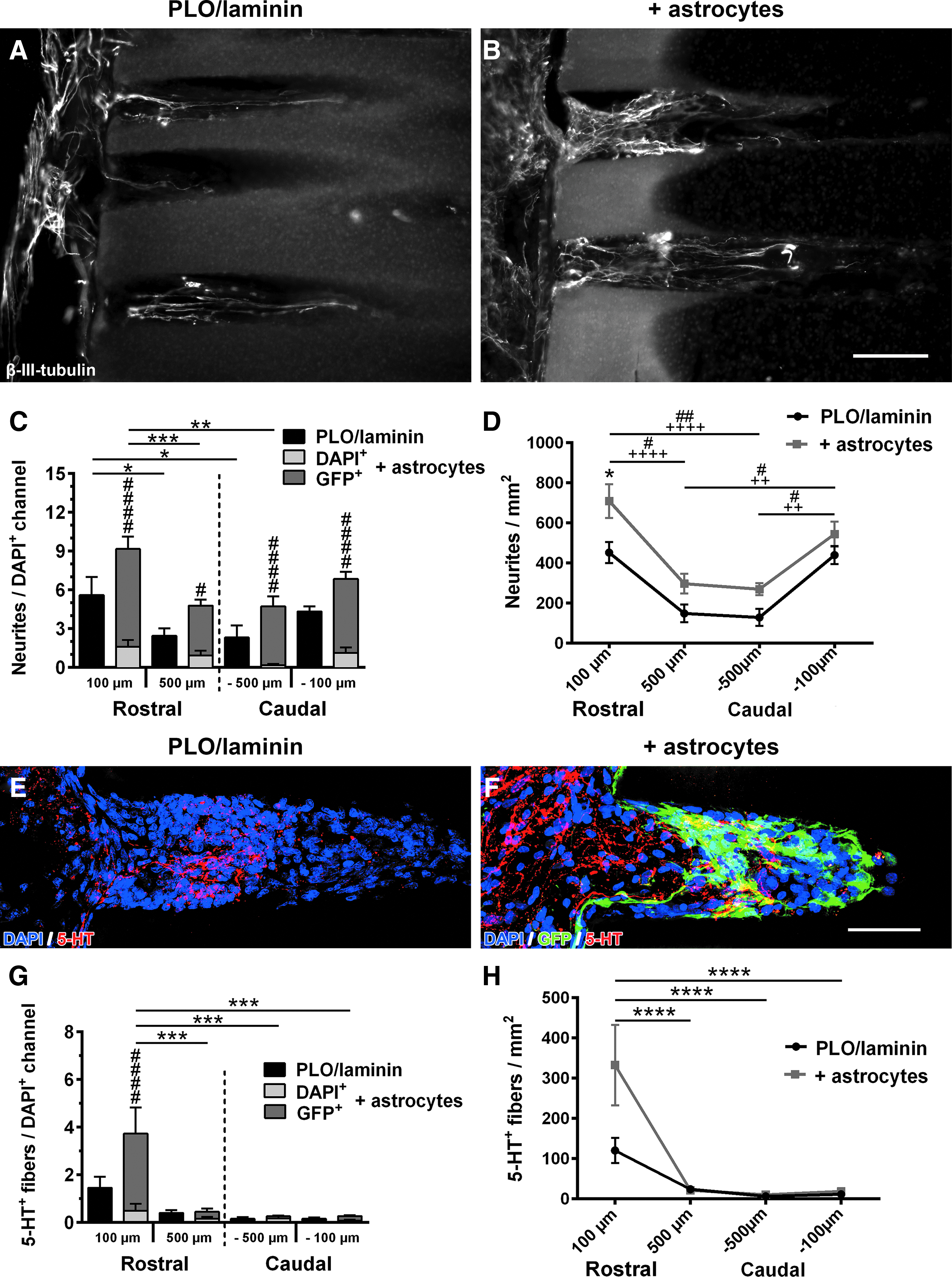
Neurite growth into microchannels of astrocyte-seeded hydrogel scaffolds.
To assess whether descending fibers contributed to neurite growth into the hydrogel scaffolds, raphespinal axons were labeled with an antibody for serotonin (5-HT) (Fig. 7E, F). Descending 5-HT+ fibers extended toward the injury site and entered the rostral end of the scaffolds in both groups (Fig. 7E, F). In astrocyte-seeded scaffolds, serotonergic fibers extended beyond areas with dense GFP labeling indicating that grafted astrocytes did not provide a barrier for axonal extension (Fig. 7E). The largest number of 5-HT+ axons was found at the rostral entry zone followed by a dramatic decrease toward the central and caudal part of the scaffold. Importantly, serotonergic fibers preferred astrocyte-containing channels over channels containing only infiltrated host cells (Fig. 7G), consistent with data evaluating overall neurite growth (Fig. 7C) (Rostral 100 μm, p < 0.0001). The same result was observed when the amount of 5-HT+ fiber growth in the entire scaffold was evaluated: on average, nearly 3 × as many raphespinal fibers entered the scaffold in animals with astrocyte seeding compared with controls, but this difference did not reach significance due to high variability, and the number of serotonergic axons dramatically declined toward the central and caudal regions of the scaffolds (p < 0.0001) (Fig. 7H).
In summary, seeding of neonatal astrocytes into alginate-based hydrogels improved scaffold integration into the surrounding host parenchyma. While seeded astrocytes only slightly increased axonal entry into the scaffold, regrowing axons preferred an environment containing grafted astrocytes over infiltrating host cells within scaffold channels.
Discussion
The present study demonstrates that the surface of alginate hydrogels can be efficiently modified by positively charged peptides, enhancing cell adhesion and neurite growth in vitro, and host cell infiltration and neurite extension in vivo. In addition, neonatal astrocytes seeded into PLO/laminin-coated hydrogel scaffolds bridged the interface between host parenchyma adjacent to the scaffold and the scaffold channels, thereby further promoting short-distance axonal growth after SCI.
Protein coating of alginate surfaces is highly charge dependent. At physiological pH positively charged polyamino acids, such as PDL and PLO, electrostatically bind to the alginate backbone in large quantities. In contrast, negatively charged PDGa hardly attaches to the negatively charged alginate (Supplementary Fig. S2). As a negatively charged surface is detrimental for cell attachment and neurite outgrowth, 10 we chose the synthetic polyamino acid PLO to mask the negative charge of the alginate backbone. PLO greatly increased the number of neonatal astrocytes, as well as DRG neurons and glia attaching and surviving on alginate in vitro. As expected, addition of laminin, an integrin receptor ligand, further promoted substrate permissiveness 41 indicated by cellular spread, the development of cell processes from astrocytes, and DRG neurite outgrowth (Fig. 1). In vitro time course studies also indicated that the biological activity of PLO/laminin coating remains stable for at least 2 weeks (Fig. 2). Although peptide and protein degradation might be faster in vivo as proteases such as matrix metalloproteinases are upregulated in astrocytes, neurons, and macrophages after SCI,42,43 more cell-filled channels were found in PLO/laminin-coated hydrogels compared with uncoated scaffolds 4 weeks postimplantation (Fig. 3C). Thus, the PLO/laminin coating appears to remain biologically active for a period of time sufficiently long to promote cell migration throughout the entire scaffold.
In contrast, neurite growth was only increased in the most rostral areas by PLO/laminin coating (Fig. 4D). Because the number of neurites per cell-filled channel did not differ between groups (Fig. 4C), it is likely that the overall higher neurite density in PLO/laminin-coated hydrogels is due to a higher number of cell-filled channels rather than a direct influence of PLO/laminin on extending axons. Indeed, axons were never found in channels devoid of cells, and axons primarily extended in the central portion of each channel instead of directly contacting channel walls (Fig. 4A, B). Taken together, increased in vivo axon growth into PLO/laminin-coated scaffolds seems to be indirectly influenced by increased cell migration and survival within the implant, including Schwann cells and possibly microglia/macrophages. The growth-stimulating effect of host cells on axons in the scaffold, however, seems to substantially decrease with increasing distance from the host parenchyma. A shift in cellular composition from growth promoting toward a more inhibitory phenotype over time or distance might underlie the decrease in neurite numbers per cell-filled channel toward the hydrogel center. The lower concentration of laminin in the center of the hydrogel might also have contributed to this observation. In addition, the limited intrinsic growth capacity of injured neurons, astrogliosis, and inflammatory responses around the implant prevent more extended axonal regeneration without additional stimuli, such as growth factors, or modification of the neuronal regenerative competence or ECM. Compared with previous experiments using scaffolds that were filled with naive BMSCs (278 ± 64 neurites/mm 2 at 100 μm and 105 ± 32 neurites/mm 2 at 500 μm; 7 ), the number of axons in PLO/laminin-coated hydrogels was slightly higher in the current studies (Figs. 4D and 7D) indicating that efficient ECM modification of alginate has effects that are at least as supportive as grafting of non-neural cells into alginate scaffolds.
Previously, we have shown that BMSCs and Schwann cells survive and fill uncoated alginate channels in vivo,7,8 but more sensitive cell types, such as astrocytes are likely to benefit from surface coating as indicated by our in vitro studies (Figs. 1 and 2). In contrast to grafts of Schwann cells and BMSCs, which frequently result in clearly defined boundaries between host and grafted cells, grafting of CNS glia would be expected to enhance integration of biomaterials into the lesioned CNS environment. Indeed, astrocyte-seeded scaffolds appear to be better integrated into the host spinal cord as grafted GFP+ cells infiltrated the host–graft interface intermingling and blending with surrounding host astrocytes. As a result, a continuous astroglial tissue bridge formed guiding neurites into the microchannels (Fig. 6). This is remarkably different from previous studies with alginate or agarose scaffolds implanted without cells or with BMSCs that are surrounded by an area of hypercellularity, which partially consists of infiltrating fibroblast-like cells.7,44 Further evidence for the proregenerative capacity of neonatal cortical astrocytes in alginate channels comes from the analysis of the number of neurites per cell-filled channel. Within animals that received scaffolds filled with astrocytes, a clear preference of neurites extending into astrocyte-filled channels versus channels containing only host cells was observed. This was true when examined with a general neurite marker (β-III-tubulin; Fig. 7C) and labeling for serotonergic axons (Fig. 7G). In addition, an overall larger number of neurites was found in alginates filled with astrocytes (Fig. 7D).
Astrocytes, specifically neonatal astrocytes, secrete a myriad of factors, including neurotrophic factors, ECM components, proteases/protease inhibitors, cytokines, and chemokines mediating tissue regeneration/reorganization and neuroprotection as well as supporting axonal regeneration.27,29,45 The preference of neurites for astrocyte-containing channels indicates that, at least shortly after grafting, astrocytes were growth permissive and not growth inhibitory. These findings are consistent with earlier studies showing that grafted neonatal astrocytes can promote sensory axon regeneration across the dorsal root entry zone,26,46 support axon regeneration in combination with chondroitinase ABC, 47 and corticospinal axon growth after genetic depletion of phosphatase and tensin homolog (PTEN). 48
Although astrocyte-filled hydrogels promoted axon growth, this effect was mostly restricted to the rostral and caudal end of astrocyte-filled scaffolds and serotonergic axons failed to re-enter the caudal spinal parenchyma. The decline in the number of axons, including descending serotonergic axons might be due to the slightly lower percentage of astroglia-filled channels in the central regions of the hydrogels. Notably, astrocytes were transplanted in the absence of any supportive fibrin matrix or any growth factors used in other transplantation studies.49,50 Addition of such a matrix or a growth factor cocktail may further increase survival of the grafted cells in particular in the more centrally located regions of the alginate gel. Alternatively, a time-dependent shift in the properties of grafted astrocytes from a growth-permissive to a growth-inhibitory phenotype might occur in vivo. The astrocytic secretome is highly adaptive, affected by the lesion microenvironment51,52 and activated microglia can induce a neurotoxic phenotype in cortical astrocytes through secretion of interleukin-1α, tumor necrosis factor α, and Clq. The exact molecular mechanism behind the observed astrocyte-mediated neurotoxicity nonetheless remains elusive. 53 The origin of grafted astrocytes may also be of importance for longer distance axon growth. It is possible that spinal, instead of cortical astrocytes, will mediate more unbranched rostrocaudal growth of ascending and descending spinal tracts if they preserve their rostrocaudal identity following cell isolation and grafting.
Besides an indirect effect on astrocytes, the most prominent cell type at the lesion site, namely microglia and macrophages, significantly impacts axonal regeneration by releasing inflammatory cytokines and expressing CSPGs that can be detrimental for axonal regeneration.29,54,55 Furthermore, an initially hypoxic and hypoglycemic environment in the central region of the alginate scaffold may cause cell death and axonal dieback. This might be particularly relevant at early time points as we observed many endothelial cells and possibly blood vessels within the scaffolds at longer times (4 weeks) postinjury. Seeded astrocytes were in close contact with von Willebrand factor-labeled endothelial cells/blood vessels and seemed to envelope them with GFAP+ protrusions. As the blood–spinal cord barrier requires astroglia and is disrupted after SCI,56–58 the observed association of grafted astrocytes and endothelial cells/blood vessels might allow for the long-term re-establishment of a blood–spinal cord barrier within hydrogel channels. Further studies are needed to determine whether grafted astrocytes can improve the reformation of a functional blood–spinal cord barrier and whether manipulations that promote and accelerate the formation of blood vessels in the central region of alginate channels influence axonal regeneration.
Taken together, the current study shows a beneficial effect of PLO and laminin coating of alginate hydrogels affecting cell attachment, cell migration, and some improvement of axonal growth, which seems to be primarily due to increased cell infiltration. By providing an astroglial substrate within the scaffold, a glial continuity from host parenchyma into alginate channels can be established supporting the integration of alginate scaffolds and further enhancing axon growth. However, a combination with other growth-promoting strategies is necessary to increase the distance of neurite extension.
Footnotes
Acknowledgments
Supported by grants from the Deutsche Forschungsgemeinschaft (BL414/3-1 to A.B. and N.W. and MU2787/3-1 to R.M.), the Indiana University Health–Indiana University School of Medicine Strategic Research Initiative, and the Morton Cure Paralysis Fund to A.B., the Indiana State Department of Health (Indiana Spinal Cord and Brain Injury Research Fund) to A.B. and P.K., and a Heinz Götze Memorial Fellowship to S.L. The authors would like to thank the Missouri Rat Resource Center for providing GFP-transgenic rats.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.