
Abstract
Injectable hydrogels have the potential to enhance stem cell-based therapies by improving cell localization, retention, and survival after transplantation. The inflammatory response to both the hydrogel and the encapsulated cells is a critical aspect of this strategy, with macrophages being highly involved in the process of hydrogel remodeling, angiogenesis, and tissue regeneration. As a step toward the development of a cell-based strategy for therapeutic angiogenesis, this work compared the intramuscular injection of allogeneic rat adipose-derived stem/stromal cells (rASCs) in an in situ gelling hydrogel with the injection of the hydrogel alone and rASCs in saline in an immunocompetent rat model by immunohistochemical analysis over 4 weeks. rASCs delivered in the hydrogel were retained intramuscularly at significantly higher densities as compared with cells delivered in saline. The encapsulated rASCs modulated the inflammatory response, promoting CD68+ macrophage recruitment, with the majority of infiltrating cells expressing the M1 marker CCR7, as well as a higher fraction of CD163+ M2c macrophages surrounding the hydrogel. Furthermore, rASCs reduced the initial expression of inducible nitric oxide synthase and promoted arginase-1 expression in the infiltrating macrophages over time, consistent with a shift toward a more proregenerative phenotype. Coincident with the enhanced macrophage infiltration, significantly more CD31+ lumens were observed surrounding and within the hydrogels with rASCs at 2 and 4 weeks as compared with the hydrogels alone. Overall, these results are a promising indication that encapsulated rASCs can have immunomodulatory effects and may enhance angiogenic processes after intramuscular injection, promoting a regenerative macrophage response and blood vessel formation.
Impact Statement
Hydrogel-encapsulated mesenchymal stem/stromal cells are immunomodulating in the context of an implanted hydrogel scaffold and can have a significant effect on the fate of the material. Future design of mesenchymal stem cell delivery platforms should consider the interaction of the stem cells with resident macrophages, and the potential this interaction has on material degradation and integration.
Introduction
M
To address these challenges, in situ forming hydrogels have been widely investigated as a strategy to improve cell retention after intramuscular injection.17,18 In this approach, cells are injected in a polymer solution that crosslinks in situ to form a three-dimensional network, encapsulating the cells at the site of delivery to minimize initial cell washout, prevent anoikis, and facilitate the generation and local release of paracrine factors. 19 Toward these goals, our group has previously developed a mechanically resilient, injectable hydrogel composed of two polymers: a triblock copolymer of poly(trimethylene carbonate)-b-poly(ethylene glycol)-b-poly(trimethylene carbonate) diacrylate [PEG-(PTMC-A)2] and methacrylated glycol chitosan (GC) functionalized with an arginine-glycine-aspartic acid-containing peptide (MGC-RGD). 20 The PEG-(PTMC-A)2 component was used to modify the mechanical properties of the hydrogel to withstand the mechanical loading conditions of the lower limb, and the MGC-RGD component was selected to support cell retention, viability, and function.21–23
The purpose of this work was to evaluate the ability of the hydrogel to improve the retention of allogeneic rat adipose-derived stem/stromal cells (rASCs) after intramuscular injection, and second to characterize the in vivo response to the hydrogel, with and without rASCs, in terms of the inflammatory response and vascularization. An immune-competent, healthy rat model was used to study the inflammatory response to the material without the confounding effects of inflammation due to ischemia.
Inflammation is a fundamental response to implanted hydrogels that is closely linked to the angiogenic process, and ultimately determines the implant fate in vivo.24–26 Both inflammation and angiogenesis are governed by the activity of macrophages, through their long-term interactions with the biomaterial and their role in the recruitment and direction of endothelial cells. 27 As such, modulating the macrophage response to biomaterials is an important strategy to promote wound healing and angiogenic activity, the retention and survival of encapsulated cells, and the remodeling and eventual clearance of the material.28–32 Specifically, strategies for treating ischemic conditions have focused on balancing the responses of M1 and M2 macrophages,33–35 which represent the proinflammatory and proregenerative phenotypes along the spectrum of macrophage polarization, respectively. 36 These phenotypes are frequently characterized by surface marker expression, where M1 is associated with higher levels of chemokine receptor CCR7 expression and M2 with higher levels of scavenger receptor CD163. 33
ASCs have received increasing attention as an abundant and readily accessible source of MSCs for the treatment of ischemia not only due to their angiogenic potential6,7,9,10 but also due to their potent immunomodulatory effects. In vitro, ASCs promote a regenerative phenotype, and the production of anti-inflammatory cytokines in peripheral blood mononuclear cells 37 and macrophages.38,39 In vivo, ASCs have been associated with a more regenerative macrophage response and increased systemic anti-inflammatory cytokine levels after liver transplantation in rats, 40 as well as in diabetic mice. 39 When seeded in decellularized adipose tissue bioscaffolds and implanted in rats, rASCs promoted a shift toward an M2 macrophage phenotype, and were associated with enhanced vessel formation and healthy, host-derived soft tissue formation. 41 These properties make ASC delivery a promising strategy for modulating both the proinflammatory microenvironment within tissues impacted by chronic ischemia and the host response to implanted biomaterials.
In this study, immunohistochemical (IHC) analysis was performed at 1, 2, and 4 weeks to compare the host response to intramuscularly injected composite PEG-(PTMC-A)2 + MGC-RGD hydrogels incorporating allogeneic rASCs relative to control groups of hydrogels without rASCs and rASCs delivered in saline. Intramuscular allogeneic rASC density was semiquantitatively analyzed through PKH26 fluorescent cell tracking to determine whether the hydrogel enhanced cell retention. To assess the effects of rASCs on the macrophage response to the hydrogels, macrophage infiltration and polarization toward a “pro-inflammatory” M1 versus a “pro-regenerative” M2 phenotype were probed using a range of markers. Finally, the ability of rASCs to enhance blood vessel formation was characterized in comparison with the controls based on the density of CD31+ lumens at the site of each treatment.
Experimental
Materials
Trimethylene carbonate (TMC) was obtained from Leapchem (Hangzhou, China). HCl (1 M in diethyl ether) was obtained from Acros Organics. GC (minimum number average molecular weight = 82 kg/mol, 85% degree of deacetylation [DOD]) was obtained from Wako Chemicals, Inc. GGGG
Polymer preparation
PEG-(PTMC-A)2 was prepared as previously described 20 using 20 kg/mol PEG as an initiator and PTMC block lengths of 1.8 kg/mol. This molecular weight and PEG/PTMC ratio was selected to provide a reduced compressive modulus and enhanced toughness suitable for the intramuscular environment. 20
MGC was prepared as previously described. 23 The free amine groups of GC were methacrylate functionalized through reaction with glycidyl methacrylate. RGD peptide was N-terminal functionalized with N-acryloxysuccinimide and conjugated to MGC by Michael addition to free amine groups.
Rat adipose-derived stem cell isolation and culture
All studies involving animals followed the Canadian Council on Animal Care guidelines, and were reviewed and approved by the Animal Care Committee at Queen's University (protocols Amsden 2015-1628 and Amsden 2013-048). rASCs were isolated from the epididymal fat pad of male Wistar rats (aged 10–12 weeks) using published methods, 41 adapted from protocols for the isolation of human ASCs. 42 rASCs were plated at 5000 cells/cm 2 in complete medium, cultured at 37°C with 5% CO2, and passaged at 80% confluence. Passage 2 cells were used for the study. Single color flow cytometry analysis was performed (n = 3) using a Guava easyCyte 8HT Benchtop flow cytometer (EMD Millipore, Billerica, MA) as previously reported (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/tea). 43 Two days before intramuscular delivery, donor rASCs were labeled with the lipophilic membrane dye PKH26 according to the manufacturer's instructions, replated at their original density, and rinsed with PBS.
Intramuscular injection
Intramuscular injections were administered to healthy female Wistar rats aged 8 weeks. The rats were anesthetized with isoflurane, and given subcutaneous injections of meloxicam (2 mg/kg) as a preoperative analgesic and bupivacaine (2 mg/kg) as a local anesthetic. Three treatment groups were tested:
Gel + rASCs: rASCs (2 × 10
7
cells/mL) in hydrogel prepared from mixed solutions of 4% w/v PEG-(PTMC-A)2 + 1% w/v MGC-RGD, dissolved in PBS; (ii). Gel alone: hydrogel prepared from mixed solutions of 4% w/v PEG-(PTMC-A)2 + 1% w/v MGC-RGD, dissolved in PBS; (iii). rASCs + saline: rASCs (2 × 10
7
cells/mL) suspended in PBS.
All polymers were decontaminated by exposure to low-intensity UV light in a biological safety cabinet for 40 min before dissolving in sterile PBS. For hydrogels with rASCs, a suspension of PKH-labeled rASCs in sterile PBS was thoroughly mixed in the polymer solution. To initiate polymerization of both hydrogel groups, sterile filtered 100 mM solutions of ammonium persulfate (APS) and tetramethylethylenediamine (TEMED) were added sequentially to the polymer solutions immediately before administration, with thorough mixing between each addition, providing a final concentration of 5 mM each of APS and TEMED.
The treatments were performed by injecting a total volume of 50 μL divided evenly over two injection sites 5 mm apart centered along the length of the adductor muscle using a 25G × ½″ needle (N = 4 per treatment type, per time point). After injection, the animal was maintained under anesthesia for 5 min to prevent muscle contractions from dispersing the delivered material or cells. Rats were euthanized by CO2 overdose at 1, 2, and 4 weeks postinjection. A 15 mm long section of the adductor muscle group centered on the injection sites was excised, embedded in Cryomatrix frozen embedding resin, and snap frozen in liquid nitrogen.
IHC analysis
The characterization studies focused on IHC staining to be able to assess the spatial distribution of rASCs, macrophages, and CD31+ lumens relative to the hydrogels and surrounding muscle tissue. Frozen tissue was cryosectioned perpendicular to the muscle fiber orientation at 5 μm thickness. Sections were taken at the injection sites, as identified by the presence of PKH+ cells and/or the hydrogel.
PKH+ rASC retention
To quantify PKH+ rASC density, unfixed sections from three depths across the injection site were mounted in fluoroshield mounting medium with 4′,6-diamidino-2-phenylindole (DAPI; Abcam), and all nucleated PKH+ cells across the entire section were imaged with a Zeiss Axio Imager M1 microscope with a 20 × objective. The total number of PKH+ cells was normalized to the total muscle cross-sectional area. Identification of PKH+ rASCs was performed on unfixed, unstained tissue sections, as the process of fixation and permeabilization reduced the PKH staining fidelity.
Macrophage response and vessel formation
Sections were fixed with 4% formaldehyde, rinsed with TBS, and blocked with 5% goat serum in TBS with 0.2% Tween-20 (TBS-T) for 1 h at room temperature. The serum solution was replaced with primary antibodies diluted in TBS-T with 1% bovine serum albumin as shown in Table 1, and incubated overnight at 4°C. Anti-CD31 was applied alone; anti-CD163 and anti-CD68 were applied alone to serial sections to facilitate comparison of CD163+ and CD68+ cell densities; anti-CCR7, antiarginase-1 (Arg-1), and anti-inducible nitric oxide synthase (iNOS) were each paired with anti-CD68. After primary incubation, the sections were rinsed three times with TBS and incubated with appropriate secondary antibodies diluted in TBS-T with 1% bovine serum albumin as shown in Table 1 for 1 h at room temperature. The secondary antibody solutions were rinsed three times with TBS, and the sections were mounted with fluoroshield mounting medium with DAPI. All imaging was performed with a Zeiss Axio Imager M1 microscope.
iNOS, inducible nitric oxide synthase.
The maximum penetration depth of CD68+ cells into each hydrogel was measured perpendicular to the border from five points (10 × magnification) using Zeiss Zen software (v. 2.3). The fractional area of the gels staining CD68+ was determined by converting the images (10 × magnification) to binary and measuring the ratio of CD68+ area within the gel to total gel area using ImageJ software (v. 1.46r). To assess macrophage polarization, a minimum of four nonoverlapping images per hydrogel were analyzed (40 × magnification, centered on hydrogel border). The total number of cells that were CD68+ and CD68+ CCR7+ was counted; the number of CD163+ cells was similarly counted on images taken from serial sections to the CD68 images, and matched in terms of location at the gel border. To assess enzymatic expression, at least four images per hydrogel were used (40 × magnification, taken from areas within the hydrogel), and the total number of cells that were CD68+, CD68+ iNOS+, and CD68+ Arg-1+ was counted (ImageJ v. 1.46r and Zeiss Zen v. 2.3 software).
CD31+ vessel density was quantified using at least six nonoverlapping images from each sample (20 × magnification) at the border of the hydrogels or at the site of PKH+ cells, when delivered in saline. The minor diameter of all CD31+ lumens was measured using AxioVision software (v. 4.9.1).
Statistical analysis
All data are presented as mean ± standard deviation. Comparisons were made by two-way ANOVA. Differences between groups at given time points and differences within groups across time points were determined by Sidak's multiple comparisons test, with adjusted p-values <0.05 considered significant. Statistical analysis was performed using GraphPad Prism 7.
Results
Injection within in situ forming hydrogels improves rASC retention at the target site
Cell implantation within the hydrogels was achieved through intramuscular injection with in situ thermally initiated free radical crosslinking of a suspension of rASCs in an aqueous mixture of the prepolymers MGC-RGD and PEG-(PTMC-A)2. PKH+ rASCs delivered in the hydrogel and in saline were identified in the adductor muscle of all recipient rats at 1, 2, and 4 weeks postinjection. rASCs delivered by in situ hydrogel formation were observed predominately within the gel, located in the intramuscular space (Fig. 1a). When delivered in saline, rASCs were largely found along the subdermal fascia (Fig. 1b).

Representative images from adductor muscle sections showing PKH+ rASCs (red) delivered in the
At weeks 1, 2, and 4 significantly more rASCs were observed per adductor cross-sectional area when delivered in the hydrogel, compared with saline delivery (p = 0.009, 0.001, and 0.001, respectively) (Fig. 1c), supporting that the hydrogel improved cell retention. No significant differences in rASC density between time points were found, suggesting that any rASCs that were lost during delivery were lost within the first week.
rASCs promote macrophage recruitment and infiltration into the hydrogel
CD68 was used as a pan-macrophage marker in assessing the response to the hydrogels formed in situ with (Fig. 2a) and without (Fig. 2b) rASCs. In general, greater CD68+ cell infiltration was observed after 2 weeks in response to the hydrogels with rASCs, compared with those without, in terms of the fractional area of the hydrogel staining CD68+ (Fig. 2c) (p < 0.001) and depth of CD68+ cell infiltration (Fig. 2d) (p < 0.0001). In hydrogels with rASCs, the infiltration fraction increased significantly from weeks 1 to 2 (p < 0.05), and the infiltration depth increased significantly from weeks 2 to 4 (p < 0.0001). In contrast, in hydrogels without rASCs, the CD68+ cell infiltration fraction did not change significantly over time, and the infiltration depth decreased from week 1 compared with weeks 2 and 4 (p < 0.01 and p < 0.05, respectively).
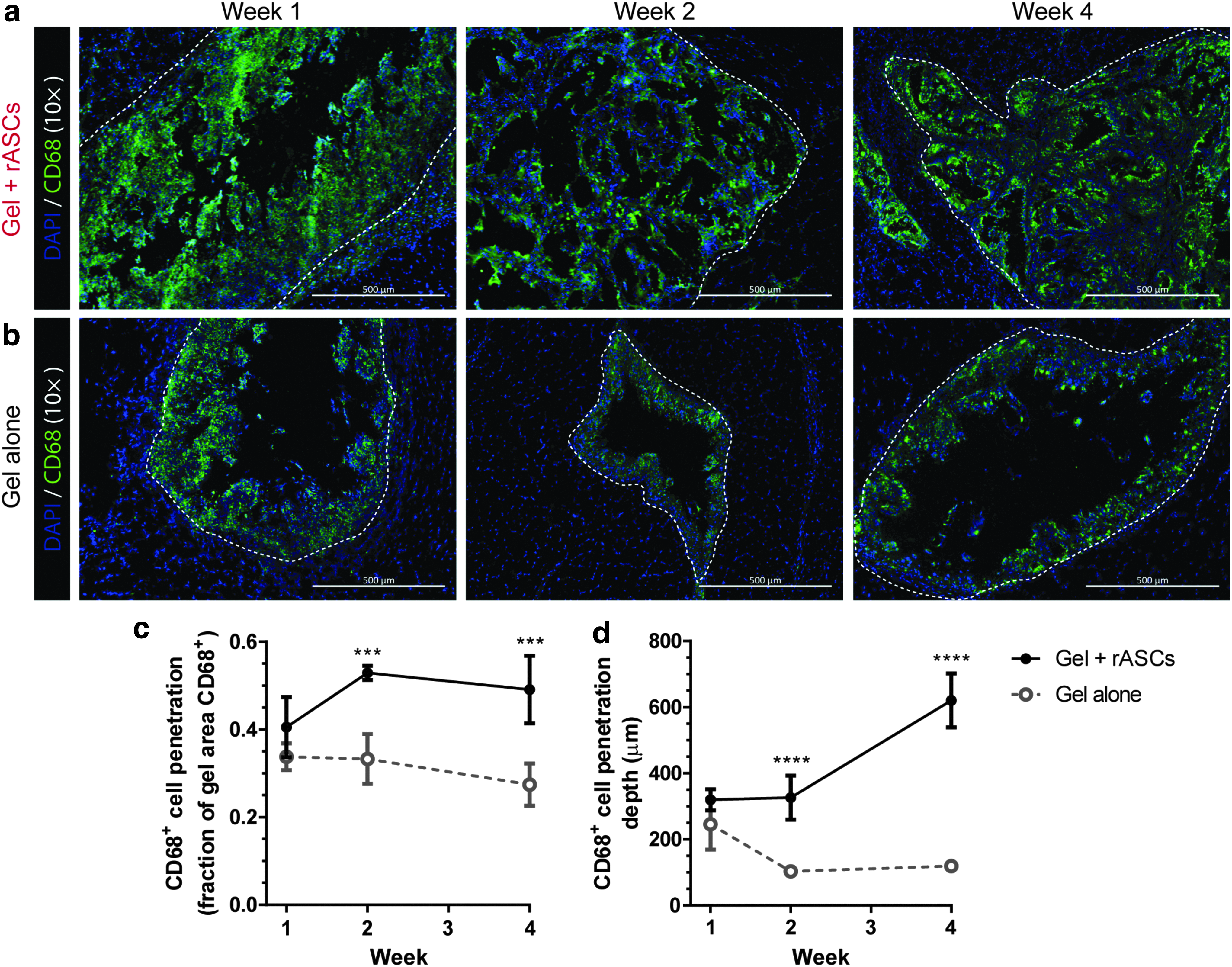
Representative images of CD68+ cell (green) penetration into the hydrogels
rASCs promote M1 macrophage infiltration and an M2c phenotype at the hydrogel periphery
The chemokine receptor CCR7 and the scavenger receptor CD163 were used as markers of M1 and M2c macrophages, 44 respectively, at the borders of the hydrogels with (Fig. 3a, b) and without (Fig. 3c, d) rASCs. Detailed images (40 × magnification) are provided in Supplementary Figures S1 and S2. In general, infiltrating cells were predominantly CD68+ CCR7+, while few peripheral CD68+ cells were CCR7+ (Supplementary Fig. S3). CD163+ cells were found at the periphery of the hydrogels, with minimal infiltration into the hydrogel.

Representative images of CCR7+ cells (yellow) and CD163+ cells (green) at the hydrogels
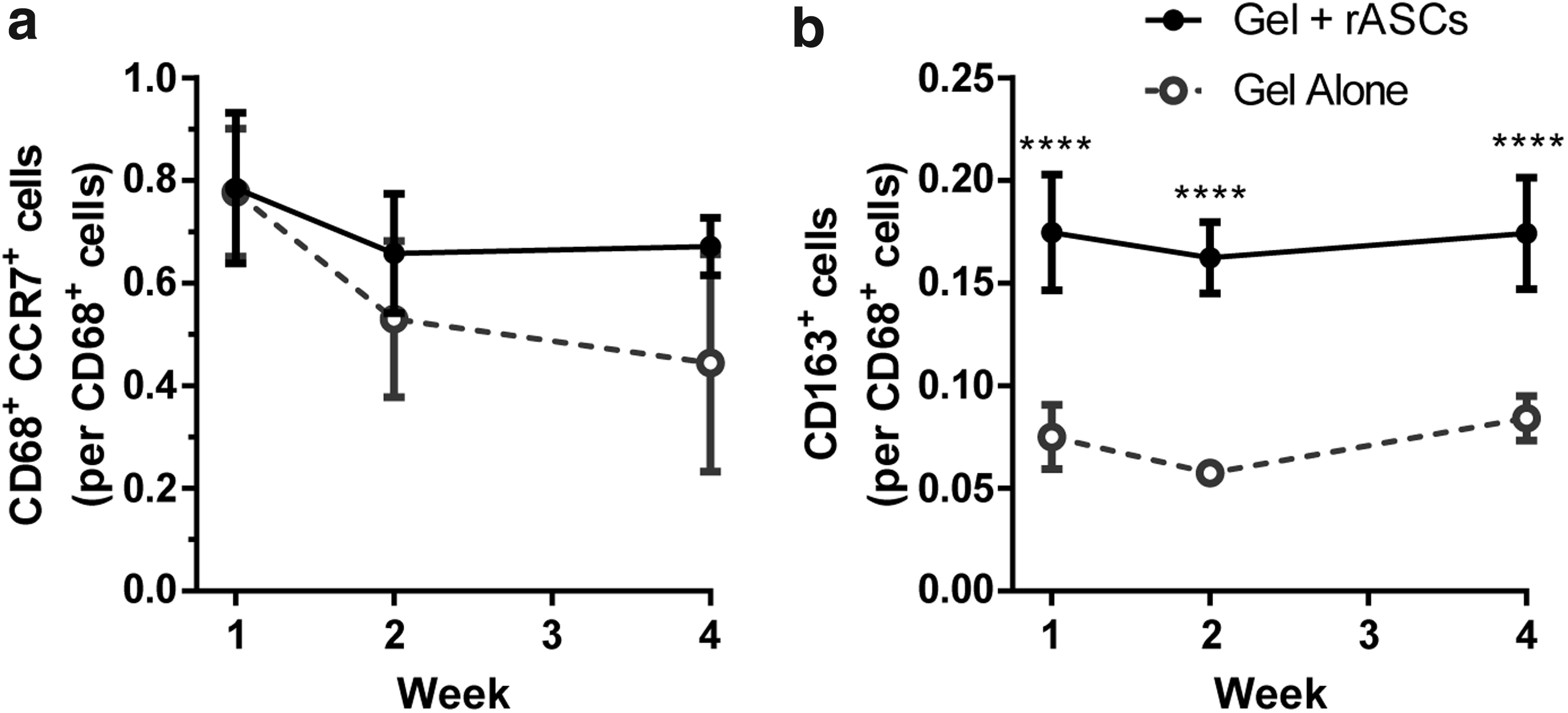
At all time points, the fraction of CD68+ cells that were CCR7+ was not significantly different between the hydrogels with and without rASCs (Fig. 4a). However, this ratio reduced significantly (p = 0.027) between weeks 1 and 4 in the hydrogels without rASCs, from ∼78% ± 12% to 44% ± 21%. As a fraction of the CD68+ cells, the relative number of CD163+ cells was significantly higher in response to the hydrogels with rASCs at all time points (Fig. 4a) (p < 0.0001). In both groups, the relative fraction remained constant over 4 weeks at 17% ± 3% and 8% ± 1% in response to the hydrogels with and without rASCs, respectively.
rASCs reduce initial iNOS activity and promote Arg-1 activity in infiltrating macrophages
Staining was performed to identify CD68+ cells expressing iNOS in the hydrogels with (Fig. 5a, b) and without (Fig. 5c, d) rASCs. In general, CD68+ iNOS+ cells were predominantly found within the gels at the advancing edge of the infiltrating CD68+ cells. Staining was also performed to identify CD68+ cells expressing Arg-1 in the hydrogels with (Fig. 6a, b) and without (Fig. 6c, d) rASCs. In general, CD68+ Arg-1+ cells were distributed throughout the infiltrating CD68+ cell population.

Representative images of iNOS+ cells (yellow) and CD68+ cells (green) infiltrating the hydrogels

Representative images of Arg-1+ cells (yellow) and CD68+ cells (green) infiltrating the hydrogels
At 1 week, a significantly higher fraction of CD68+ cells were iNOS+ in the hydrogels without rASCs versus with F7c rASCs (21.6% ± 4.7% vs. 10.9% ± 1.9%, p = 0.001) (Fig. 7a). The fraction of iNOS+ cells in the hydrogels without rASCs was significantly lower at weeks 2 and 4 compared with week 1 (p = 0.027 and p = 0.017, respectively), and not statistically different from the ratio in the hydrogels with rASCs.

At all time points, a significantly higher fraction of CD68+ cells were Arg-1+ in the hydrogels with rASCs, compared with the hydrogels without rASCs (Fig. 7b) (p < 0.05). The fraction of Arg-1+ cells increased significantly from weeks 1 to 4 in the hydrogels with rASCs (20.9% ± 6.5% to 32.7% ± 2.2%, p = 0.037), as well as in the hydrogels without rASCs (5.7% ± 1.2% to 20.4% ± 7.7%, p = 0.010).
rASCs enhance vessel density when delivered within an in situ forming hydrogel
CD31 staining was used to identify endothelial lumens at the border of the hydrogels and at the site of rASC localization for saline delivery. At 4 weeks postdelivery, CD31+ vessels were observed at the border and within the hydrogels with rASCs (Fig. 8a), whereas vessels were only observed near, but not within, the hydrogels without rASCs (Fig. 8b). Quantification of CD31+ lumen density normalized to image area (Fig. 8c) indicated that vessel density was significantly greater surrounding the hydrogels with rASCs at 2 and 4 weeks postdelivery as compared with all other groups (p < 0.0001 for all comparisons). The range in the minor lumen diameter was consistent between groups (Fig. 8d–f), centered at ∼6 μm, with no statistically significant differences in the mean diameter of each group. However, the mean vessel CD31+ lumen diameter surrounding the hydrogels with rASCs was significantly lower at weeks 2 and 4, compared with week 1 (p = 0.009 and p = 0.046, respectively), consistent with the formation of new smaller diameter vessels.

Representative images of CD31+ vessels (green) surrounding the hydrogels with
Discussion
Our understanding of the roles that macrophages play in tissue engineering is continually evolving as new evidence refines the definitions of macrophage subtypes and their activities. The field is increasingly interested in modulating this macrophage response as a strategy to promote biomaterial integration, tissue regeneration, and angiogenesis.
Chitosan, a natural polysaccharide obtained from the N-deacetylation of chitin, was used as a basis for the injectable hydrogel due to its demonstrated capacity to support encapsulated cell survival and retention in vitro.23,45 This work extends these findings by demonstrating that chitosan-based hydrogels can effectively retain encapsulated cells intramuscularly, even after 4 weeks of active muscle use. Previous studies have shown that subcutaneously implanted chitosan induced substantial macrophage recruitment, and that macrophage polarization was dependent on the DOD of the chitosan used; 85% DOD resulted in a primarily M1 response, whereas 95% DOD was associated predominantly with an M2 phenotype. 46 In this work, the composite hydrogel containing modified chitosan with a DOD of 85% similarly elicited a strong M1 macrophage response, evident in the extensive recruitment and infiltration of CD68+ CCR7+ cells. Macrophage recruitment is also influenced by the release of chitosan oligosaccharides (COSs), the degradation products of chitosan. COSs have also been demonstrated to have macrophage immunomodulatory effects, increasing Arg-1 expression in M1 macrophages, 47 and reducing the production of inflammatory factors and NO in the RAW 264.7 macrophages. 48 Thus, in this work, the enhanced macrophage infiltration observed in hydrogels with rASCs may be contributing to the production of COSs by hydrogel degradation, subsequently influencing the phenotype of the recruited macrophages.
Previous studies have demonstrated that PEG-(PTMC-A)2 hydrogels were susceptible to macrophage-mediated degradation, where specifically the incorporation of PTMC domains into a PEG-based hydrogel permitted macrophage infiltration and enhanced degradation by cholesterol esterase enzymatic activity in vitro. 49 In addition, studies of PEG diacrylate hydrogels indicate that PEG domains are susceptible to oxidative degradation through macrophage-generated reactive oxygen species. 50 Thus, the degradation of the PEG-(PTMC-A)2 copolymer was expected to be primarily enzymatically mediated, producing soluble PTMC and PEG oligomers, and 1,3-propane diol. 51 While these products are pH neutral and relatively inert, it is still likely that their release contributed to continued macrophage recruitment.
In this study, the responses of inflammatory and endothelial cells to intramuscularly injected hydrogels were highly dependent on the encapsulation of rASCs within the hydrogels. The most apparent effect of the allogeneic rASCs was the enhanced recruitment and subsequent hydrogel infiltration with CD68+ cells. Although studies with human cells have identified varying levels of CD68 expression in T cells, monocytes, fibroblasts, and fibrocytes,52–54 it would be expected that macrophages would comprise the majority of the CD68+ population in the context and time frame of this model. The rASC-enhanced recruitment of CD68+ cells was observed at 2 and 4 weeks postdelivery, when both fractional area of infiltration and penetration depth were significantly higher in the hydrogels with rASCs. In hydrogels without rASCs, macrophage penetration depth decreased after week 1; however, further studies would be required to determine if this result was due to a change in invasion behavior over time or rather macrophage-mediated surface degradation of the material. These infiltrating CD68+ macrophages were predominantly CCR7+, suggesting a primarily M1 infiltrating population. The CCR7+ fraction remained constant over time in hydrogels with rASCs but decreased in hydrogels without rASCs by week 4, suggesting that the rASCs and/or the enhanced total macrophage recruitment may be contributing to the continued M1 activation. While CCR7 is a traditional M1 marker and is most intensely expressed on M1s over other subtypes, it is important to note that not all M1s express CCR7. 33 Thus, it is possible that the infiltrating cells that were not CCR7+ may still have a proinflammatory phenotype. M1 inflammatory macrophages are capable of producing high levels of TNF-α. 44 In response to TNF-α, ASCs have been previously shown to increase production of chemotactic factors responsible for monocyte migration.55,56 Thus, it is possible that the rASCs are responding to TNF-α produced by the early inflammatory M1 macrophages, secreting chemotactic factors, and recruiting more macrophages into the hydrogel. In contrast, this interaction would have been absent at the site of hydrogels without rASCs, consistent with the reduced macrophage infiltration observed. The promotion of an M1 response by encapsulated cells has been similarly demonstrated in collagen-based scaffolds with bone marrow-derived MSCs. 57
In addition to enhancing overall macrophage recruitment, our results suggest that rASCs significantly increased the fraction of macrophages polarized toward an M2c phenotype. M2c polarization was assessed based on the CD163 marker, 33 a hemoglobin scavenger receptor associated with scavenging cellular debris and inducing intracellular signaling leading to the secretion of anti-inflammatory cytokines. 58 The fraction of M2c polarized cells was constant over 4 weeks for hydrogels both with and without rASCs, and these macrophages were predominantly found at the periphery of the hydrogels. This distribution pattern, with M1 macrophages infiltrating the hydrogels and M2 macrophages at the periphery, has been previously reported in response to subcutaneously implanted porous poly(hydroxyethyl methacrylate) hydrogels 59 and collagen-based scaffolds implanted in cranial defects. 57
M2c polarization can be stimulated through IL-10,33,44 and ASCs have previously been shown to produce IL-10, 8 as well as promote IL-10 production and CD163 expression in macrophages in vitro.37,38,60 In addition, ASCs delivered in vivo have promoted IL-10 production in obese mice, 39 and after orthotopic liver transplantation in rats. 40 Thus, the encapsulated ASCs may be enhancing the localized production of IL-10, resulting in the observed enhanced M2c polarization, although further studies would be required to verify this hypothesis.
The rASC-enhanced CD68+ cell infiltration and remodeling of the hydrogels suggest that the rASCs not only induced macrophage migration but also altered macrophage metabolism. This metabolic shift was confirmed by identifying CD68+ cells that were iNOS+ and Arg-1+. These two subsets represent the two major enzymatic pathways of arginine metabolism, and are commonly used to differentiate M1 and M2 macrophages.36,61 At week 1, a higher fraction of iNOS+ macrophages were found in the hydrogels without rASCs, suggesting that rASCs may have reduced the initial inflammatory response toward the hydrogels. In addition, the inclusion of rASCs resulted in a higher fraction of Arg-1+ macrophages at all time points, and the levels increased over time with and without rASCs. In combination with the CD163 staining results, this pattern suggests that the encapsulated rASCs promoted a transition from a more proinflammatory macrophage response to a more proregenerative Arg-1+ macrophage response. 62 This causal association is supported by the fact that, in vitro, ASCs have been shown to lower iNOS expression and increase Arg-1 expression in M1-activated macrophages, 39 and can induce unactivated macrophages toward a unique immunomodulatory phenotype with low iNOS expression and high Arg-1 expression. 38
MSC therapy for treating ischemic conditions ultimately relies on stimulating the generation of new, functional blood vessels in the affected tissue. The enhanced vessel formation observed in this nonischemic hind limb study is a promising sign for subsequent applications in an ischemic model. Compared with the hydrogels without rASCs and the sites of rASCs delivered in saline, the density of CD31+ lumens was significantly higher at the site of hydrogels with rASCs at 2 and 4 weeks postdelivery, and the average lumen diameter was smaller compared with week 1, suggesting new vessel formation. These results indicate that the effects depend on the sustained, localized concentration of rASCs, and are not simply a response to the hydrogel.
While the enhanced vessel formation is likely associated with the many angiogenic factors produced by ASCs,6–8 the fact that this process was closely linked to the progress of macrophage recruitment suggests that the macrophages may also be playing a role. While previous studies proposed that M1 macrophages were antiangiogenic and that M2 macrophages were responsible for promoting angiogenesis, 63 more recent evidence has revealed that macrophage subtypes function in a coordinated, temporally organized manner to promote vessel growth. 33 Specifically, M1 macrophages are thought to initiate angiogenesis by releasing factors known to promote an endothelial tip cell phenotype and blood vessel sprouting.33,64,65 Subsequently, M2a macrophages are thought to recruit pericytes to stabilize nascent vessels and promote anastomosis, and M2c macrophages support vessel growth through the release of matrix remodeling enzymes that facilitate vascular remodeling of the extracellular matrix. 33 In this work, the enhanced recruitment of CD68+ CCR7+ cells into the rASC-containing hydrogels observed at week 2 corresponded with the onset of increased vessel density, suggesting that endothelial cell recruitment may have been promoted by M1 macrophages. Further, rASCs promoted macrophage polarization toward the M2c phenotype, which may have facilitated the observed migration of endothelial cells and sprouting of vessels toward and into the hydrogels through the secretion of matrix remodeling factors.66,67 This activity is compared with hydrogels without rASCs, which demonstrated lower CD68+ cell recruitment and infiltration, reduced M2c polarization, and significantly lower CD31+ vessel density.
Conclusion
This work demonstrates the feasibility of using an in situ forming hydrogel to retain therapeutic MSCs in an intramuscular environment. The hydrogel itself, a composite of mechanically robust PEG-(PTMC-A)2 and cell-supportive MGC-RGD, was easily administered by injection and well retained in the intramuscular space, suggesting that the delivery strategy could be applied to a wide range of mechanically dynamic tissue environments. Compared with delivery in saline, the hydrogel improved the localization and retention of ASCs intramuscularly over 4 weeks in vivo. The presence of ASCs enhanced macrophage recruitment into the hydrogel and promoted a more proregenerative macrophage phenotype. The combined ASC and macrophage response was associated with significantly higher vessel density surrounding, and even within, the hydrogels with rASCs as compared with those without. Together, these results suggest the potential to harness the potent immunomodulatory effects of encapsulated ASCs to mediate the inflammatory response toward implanted biomaterials to promote constructive tissue remodeling and regeneration. The favorable interactions of ASCs encapsulated within the hydrogel cell delivery platform with host-derived macrophages may be applied to enhance angiogenesis in the treatment of a variety of ischemic conditions.
Footnotes
Acknowledgments
Funding for this project was provided by the Heart and Stroke Foundation of Canada Grant in Aid Program (GIA Project No. G-15-0008991). The authors would like to thank Cody Brown for his technical assistance with the flow cytometry characterization of the rASCs.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.