
Abstract
The authors developed SOX-6, 9-transfected human adipose stem cells (SOX-6, 9ASCs) to treat osteoarthritis (OA) and tested their effectiveness in arresting OA progression when injected intra-articularly (IA) in a surgically-induced OA caprine model. SOX-6, 9ASCs demonstrated similar in vitro chondrogenic potential, as determined by proteoglycan production, to ASCs treated with TGF-β2 and BMP-7 (positive control). In contrast, the messenger RNA (mRNA) expressions of COL2A1 and aggrecan were 3.5-fold (p = 0.004) and 1.4-fold (p = 0.058) higher, respectively, in SOX-6, 9ASCs than in the positive control. In vivo tracking of injected SOX-6, 9ASCs in rats demonstrated that these cells disappear from the joint cavity within 2 weeks, suggesting paracrine mode of action. In a surgically-induced goat model of OA, IA SOX-6, 9ASCs at a dose of 0.6 × 107 best preserved articular cartilage and produced significantly better macroscopic and microscopic scores than negative controls in femoral and tibial articular surfaces. In conclusion, SOX-6, 9-transfection led to in vitro chondrogenesis of ASCs comparable to that achieved by growth factor treatment. IA injection of SOX-6, 9ASCs in optimal dose reduced the progression of surgically-induced OA in goats. We suggest that SOX-6, 9ASCs offer a novel potential strategy to treat OA.
Impact Statement
We developed SOX-6, 9-transfected human adipose stem cells (SOX-6, 9ASCs) to treat osteoarthritis (OA). SOX-6, 9-transfection led to in vitro chondrogenesis of adipose stem cells (ASCs) comparable to that achieved by growth factor treatment. Intra-articular injection of SOX-6, 9-transfected ASCs reduced the progression of surgically-induced OA in goats. In vivo tracking of transfected cell suggested paracrine mode of action. We think that SOX-6, 9-transfected ASC provides a novel potential strategy for the treatment of OA.
Introduction
Osteoarthritis (OA), the most common type of arthritis, is characterized by the breakdown and subsequent loss of articular cartilage (AC). Unfortunately, current treatment modalities have several limitations. Conservative treatments, such as nonsteroidal anti-inflammatory drugs, provide symptomatic relief but do not regenerate damaged AC. 1 Microfracture is performed to mobilize endogenous bone marrow cells for tissue repair, but it actually results in the production of fibrous cartilage rather than hyaline cartilage and poor clinical outcomes. 2 Autologous chondrocyte implantation (ACI), which is used to treat focal defects, is associated with donor site morbidity and disadvantaged in the limited amount of available cells. Thus, ACI is not indicated for the diffuse involvement typical of OA. 3 Accordingly, the current standard treatment for advanced OA is joint replacement. However, an artificial joint cannot be used for a lifetime in younger patients, and eventually revision surgery is required. These limitations of current treatment methods have created a need for a cell therapy that can promote AC regeneration, alleviate symptoms, and arrest the progression of OA at an early stage.
SOX-trio (SOX-5, -6, and -9) are transcription factors that belong to the same family of regulatory molecules. 4 SOX-9 is expressed in all chondrogenic cells except hypertrophic chondrocytes 5 and binds to and activates other chondrocyte-specific enhancer elements in COL2A1, COL9A1, COL11A2, and aggrecan.5–7 SOX-5 and SOX-6 are cofactors of SOX-9 that are required for optimal chondrogenesis.5,8,9 Previous studies have reported on the use of SOX gene transfers to induce chondrogenesis in human embryonic stem cells and fibroblasts and to promote chondrogenesis in murine mesenchymal stem cells (MSCs).10–12 Moreover, adenoviral cotransfections of SOX-5, -6, and -9 induce the chondrogenic differentiations of MSCs and dermal fibroblasts that normally do not differentiate into cartilage. 13 In previous studies, we found that the electroporation-mediated transfection of SOX trio genes enhanced the in vitro chondrogenic potentials of bone marrow stem cells (BMSCs) 14 and adipose stem cells (ASCs). 15 We also reported that a porous PLGA scaffold impregnated with plasmid deoxyribose nucleic acid (DNA) containing SOX trio genes promoted in vitro chondrogenesis of ASCs and in vivo regeneration of osteochondral defects. 16 Our group had also shown that implantation of retroviral SOX trio-cotransduced ASCs enhanced cartilage repair and delayed the progression of surgically-induced OA in rats. 17 Encouraged by these results, we sought to devise a SOX gene-transfected stem cell to treat OA.
In previous studies, we transfected BMSCs and ASCs with separate SOX-5, 6, 9 plasmids at the same time.14–17 However, because it was not possible to confirm whether all three genes had entered the nucleus, we constructed a polycistronic plasmid containing all three genes from which all three genes would be transcribed simultaneously. However, due to very high cell death rates during electroporation encountered when these large plasmids (>20 kb) were transfected into human ASCs (hASCs) (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea), we switched to a two gene (SOX-6 and SOX-9) construct to improve the viability of the transfected cells. Although previous studies have reported that all three genes are required to induce chondrogenesis from nonchondrogenic cells,4,5,13 a recent study showed that functionally SOX-5 is similar to SOX-6 and that SOX-6 alone can act as a SOX-9 cofactor. 18
Therefore, in this study, we developed SOX-6, 9-transfected human ASCs (hereafter designated as SOX-6, 9ASCs) to treat OA and tested the hypothesis that they effectively prevent OA progression and are safe when administered intra-articularly (IA) in caprine models. In addition, extensive cell tracking of injected cells was performed in rats to study the cell survival and distribution in this therapy.
Materials and Methods
Construction of SOX-6, 9 minicircle parental plasmids
The parental plasmid of minicircle (MC) containing CMV (pMC.CMV-MCS-SV40 PolyA) was purchased from System Biosciences (Mountain View, CA). This system comprises a parental plasmid pMC.CMV-SOX9-T2A-SOX6-SV40 PolyA (hereafter called pMC.SOX-6, 9; size 9827 bp) from which MC.DNA vector was derived using the induction protocol described below. The coding regions of human SOX-6 and SOX-9 were amplified by polymerase chain reaction (PCR) from the NIH Mammalian Gene Collection (MGC) of complementary DNA (cDNA) Clones (Invitrogen, Carlsbad, CA) using the primers listed below (XbaI, EcoRI, and BamHI restriction sites are underlined; the 2A peptide sequence was inserted into SOX-9 reverse primer):
SOX-6 Forward: 5′-
SOX-6 Reverse: 5′-
SOX-9 Forward: 5′-
SOX-9 Reverse: 5′-
PCR products were subcloned into pGEM-T Easy vector and confirmed by DNA sequencing. After ligation of PCR products and pGEM-T Easy vector, they were inserted into empty parental plasmid vector using EcoRI/EcorI (SOX-6) and XbaI/EcoRI (SOX-9) restriction enzymes, respectively (Fig. 1A).
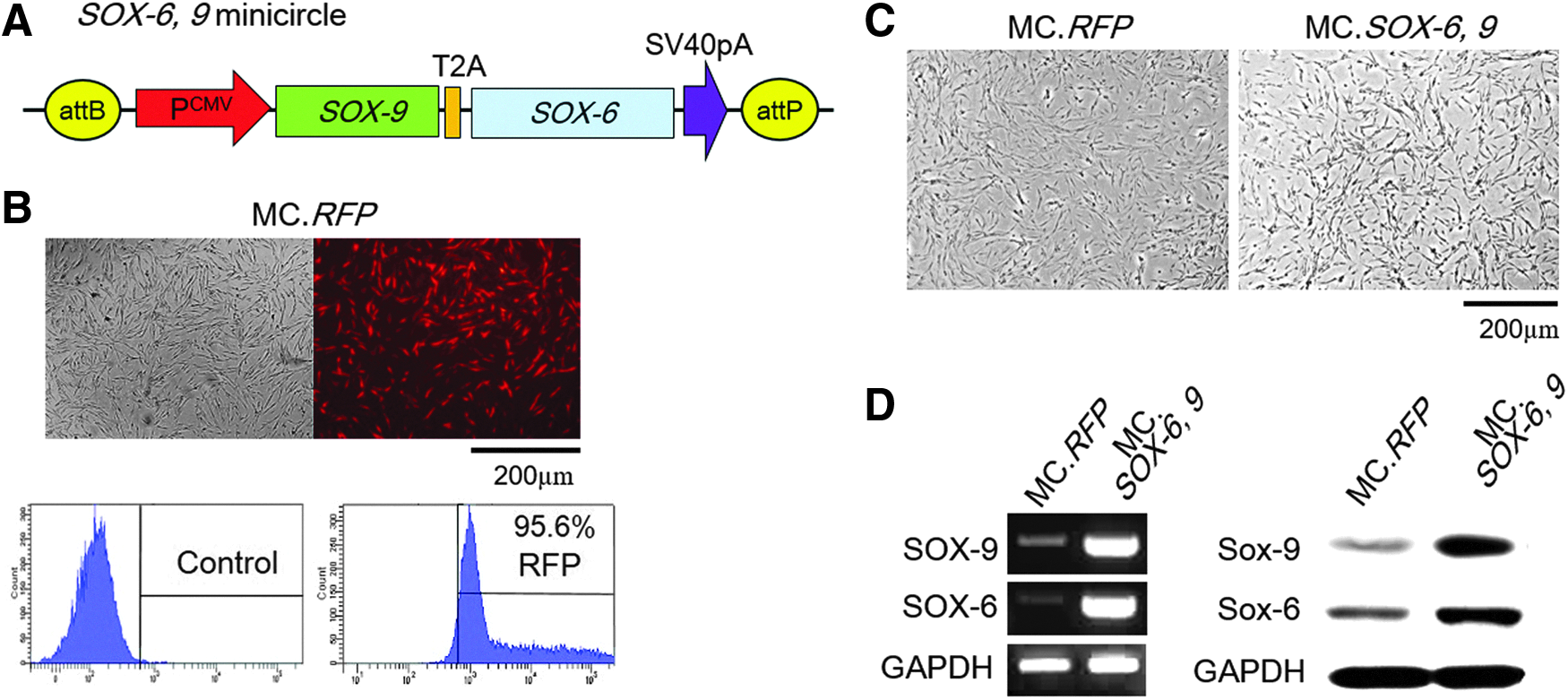
Preparation and transfection efficiency of SOX-6, 9 MC vector.
Production of MC vector
To produce MC vectors, parental plasmids were transformed into ZYCY10P3S2T Escherichia coli strain (System Biosciences).
19
Assessment of growths and inductions of MC vectors were performed according to the manufacturer's instructions and using the protocol described by Chen et al.
19
Briefly, a single colony was precultured at 37°C for 2 h in Luria-Bertani (LB) broth containing kanamycin (50 mg/mL) and then seed-cultured in Terrific broth (TB) containing kanamycin (50 mg/mL) at 30°C overnight. The next day, induction medium supplemented with 0.01%
Preparation of hASCs
hASCs were isolated from the lipoaspirates generated during elective liposuction of 3 female patients (mean age, 52 years). Protocol involving human tissue was approved by the Institutional Review Board (IRB) of Hallym University Hangang Sacred Heart Hospital (IRB 2015-001). Informed consent was obtained from all patients. hASCs were cultured and expanded as previously described 20 in Dulbecco's modified Eagle's medium (DMEM)/F12 (Gibco, Gaithersburg, MD) containing 10% fetal bovine serum (Gibco), which was changed every 48–72 h. Cells passaged five to seven times were used for transfection.
Transfection by electroporation
For transfection using the Neon transfection system (Life Technologies, Carlsbad, CA), cultured hASCs were harvested, washed with phosphate buffered saline (PBS), resuspended in resuspension buffer R (Invitrogen) at a density of 1 × 10 6 cells/100 μL, and mixed with 4 μg of MC vectors of SOX-6, 9 or red fluorescence protein (RFP). Electroporation was performed using a microporator using programs recommended by the manufacturer (1400 V, 20 ms, two pulses). After electroporation, cells were plated on a 10 cm dish and placed in a 5% CO2 incubator at 37°C. Red fluorescence from MC.RFP was visualized using a Leica DMI 6000B microscope (Leica Microsystems, Wetzlar, Germany). To measure transfection efficiency quantitatively, after 48 h of transfection, cells were harvested in 0.05% trypsin-EDTA and washed with PBS thrice. They were then analyzed using a FACSCanto II flow cytometer (BD Biosciences). hASCs electrophoresed without MC vector were used as the controls. To measure cell viabilities, harvested cells were stained with trypan blue, and then percentages of viable cells were obtained using a LUNA-FL system (Logos Biosystems, Gyeonggi-do, South Korea).
Characterization of SOX-6, 9ASCs
hASCs were analyzed for a set of accepted markers (CD34, positive for hematopoietic stem cells and endothelial cells; CD45, positive for white blood cells; HLA-DR, positive for leukocytes; and CD73, CD90, and CD105, each positive for MSCs) before and after transfection using fluorescence-activated cell sorting analysis (CytoFLEX; Beckman Coulter, Inc.) of hASCs. In untransfected hASCs, 98.1–99.8% of cells were negative for CD34, CD45, and HLA-DR, while 91.2–97.0% of cells were positive for CD73, CD90, and CD166. (Fig. 1A). Similarly for SOX-6, 9ASCs, 99.2–99.7% were negative for CD34, CD45, and HLA-DR, while 99.7–99.8% of cells were positive for CD73, CD90, and CD166 (Supplementary Fig. S2).
Chondrogenic pellet culture of transfected hASCs
To investigate the in vitro chondrogenic differentiation of hASCs, transfected cells were harvested from 10 cm dishes after 72 h of in vitro culture, and then 2.5 × 10
5
transfected hASCs were cultured in pellets under base medium (BM), which was DMEM/F-12 supplemented with 1% bovine serum albumin (w/v), 10−7 M dexamethasone, 50 μM ascorbate-2-phosphate, 50 μM
Analysis of glycosaminoglycan contents
Pellets of each treatment group were digested in papain buffer at 60°C overnight, and the digested samples were then transferred to 1.5-mL microcentrifuge tubes. DNA content was determined using the Quant-iT dsDNA Assay Kit and Qubit Fluorometer system (Invitrogen). Glycosaminoglycan (GAG) production was determined using a Blyscan Kit according to the manufacturer's instructions (Biocolor, Carrickfergus, Northern Ireland). This assay is based on the specific binding of the cationic dye 1,9-DMMB to the sulfated GAG (s-GAG) chains of proteoglycans and to protein-free s-GAG chains.
Reverse transcription-polymerase chain reaction
RNA extraction was performed using TRIzol reagent (Invitrogen) according to the manufacturer's instructions and then quantified using a nanophotometer (Implen, Inc., Westlake Village, CA). Total RNA was reverse-transcribed with Maxime RT PreMix Kit oligo (dT) primer (iNtRON Biotechnology, Gyeonggi-do, South Korea) according to the manufacturer's instructions. PCR was carried out with HotStart PCR PreMix (Bioneer, Daejeon, South Korea). Real-time PCR was performed using a SYBR® Premix Ex Taq™ (Takara Korea Biomedical, Inc., Seoul, Korea) and a LightCycler 480 system (Roche Diagnostics, Mannheim, Germany). The expressions of the SOX-9, aggrecan, and COL2A1 genes were determined. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control for the PCR amplification, and relative normalization ratios of PCR products derived from target genes were calculated using LightCycler System software (Roche, Nutley, NJ) based on delta-delta Ct method. The primer pairs used in the reactions are detailed in Table 1. Values obtained were normalized versus the negative control and expressed as fold changes. All experiments were performed in triplicate.
Polymerase Chain Reaction Primer Information
GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Western blot analysis
Proteins were extracted from cultures, electrophoresed, and transferred to nitrocellulose membranes. For total protein extraction, cells or pellets were washed twice with PBS, then homogenized by grinding, and suspended in 100 μL of RIPA lysis buffer (Invitrogen) supplemented with protease/phosphatase inhibitor cocktail (Invitrogen). For aggrecan, chondroitin sulfate-GAGs were removed by digestion with chondroitinase ABC (Sigma-Aldrich), 0.1 U per 1 μg GAG at 37°C for 4 h. Blots were probed with anti-rabbit SOX-6 (1:500; Abcam, Cambridge, United Kingdom), SOX-9 (1:500; Abcam), COL2A1 (1:200; Abcam), aggrecan (1:200; Abcam), GAPDH (1:2000; Abcam), followed by horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (1:2000; Cell Signaling Technology, Beverly, MA). GAPDH was used as the reference protein and was detected using its specific antibody (Santa Cruz Biotechnology, Inc.). This experiment was repeated on three samples from each of the three individuals.
In vivo tracking of injected SOX-6, 9ASCs in rats
Retention time of SOX-6, 9ASCs in knee joints. Three 12-week-old male SD (Sprague Dawley) rats were used for the detection of fluorescence dye-labeled cells. Harvested SOX-6, 9ASCs were labeled with CellVue® Claret Far Red Fluorescent Cell Linker Kits (Sigma-Aldrich) or Vybrant™ DiD cell-labeling solutions (Invitrogen) according to manufacturers' instructions. Labeled cells (1 × 10 6 /100 μL PBS) were injected into left and right knee joints (left: CellVue-labeled cells, right: DiD-labeled cells). After injecting fluorescence dye-labeled SOX-6, 9ASCs IA, fluorescence spectra (n = 3) were scanned using an in vivo imaging system (IVIS; Xenogen, Hopkinton, MA). Three replicate images were obtained on days 1, 2, 7, 14, 21, 28, and 42 after injection.
A 56-day IA biodistribution study was also performed using SD rats. Control rats (3 females and 3 males) were injected with 60 μL of vehicle (HypoThermosol-FRS), and experimental rats (28 females and 28 males) were injected with 1 × 10 6 of SOX-6, 9ASCs in 60 μL of vehicle. Animals were then assessed clinically and weighed on a daily basis and sacrificed on days 1, 2, 3, 7, 14, 28, or 56 (n = 8/time point). Samples of liver, heart, lung, kidney, spleen, pancreas, lymph nodes, bone marrow, brain, blood, ovaries/testes, and right knee cartilage were collected from each rat. All tissues were evaluated for the presence of human Alu or human SOX-6 cDNA by quantitative PCR (qPCR), which was performed using 3 μL of purified DNA (600 ng DNA). Standard curves were obtained from the data using simple regression analysis. Animal experiments were approved by the Animal Research and Care Committee of Chemon Institution (Chemon-16-R105: Yong-in, Korea). The primer pairs of human Alu and human SOX-6 are shown in Table 1.
IA injection of SOX-6, 9ASCs in OA-induced rats
OA was induced surgically using anterior cruciate ligament (ACL) transection in twelve 12-week-old male SD rats, according to a previous report. 1 The rats were treated with SOX-6, 9ASCs by IA injection at week 8 after OA induction. IA injection of vehicle (100 μL PBS) or 1.0 × 10 6 SOX-6, 9ASCs in 100 μL PBS was performed in the same way to be used as controls (n = 6 in each group). Rats were sacrificed for analysis after days 7 and 14. The distal femora in each group were dissected and fixed in 10% formaldehyde in PBS overnight. Then they were decalcified and sliced on a freezing microtome (CM 1850; Leica Microsystem). The sections (20 μm thick) were subjected to immunohistochemistry (IHC). The human nuclear antigen (HN; 1:100; Chemicon Temecula) primary antibody was applied overnight at 4°C followed by an hour application of secondary antibody (goat anti-mouse antibody; 1:200; Jackson ImmunoResearch Laboratories, West Grove, PA). Tissues were mounted in VECTASHIELD containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA) and analyzed under an epifluorescent microscope (Nikon, Tokyo, Japan).
In vivo efficacy of SOX-6, 9ASC injection in goats
Surgically induced caprine OA model
Goat experiments were approved by the Animal Research and Care Committee of the authors' institution (IACUC No. MCTTIACUC ASP-15-001). Thirty Korean native male black goats (DooYeol Biotech, Seoul, Korea) were used in the study. All goats were aged about 40 months and weighed 30–45 kg (mean, 37.5 kg) at the time of surgery. Animals were kept in a barn and fed nutrient pellets and hay on a daily basis. The animals were anesthetized using ketamine (2 mg/kg; Yuhan Corp., Seoul, Korea) and xylazine (0.1 mg/kg; Bayer Korea Corp., Seoul, Korea). After skin preparation, knee joints were exposed using a medial parapatellar approach. The medial meniscus was completely removed from its attachments by sharp dissection. Three left and three right knees were allocated to each of five groups, as defined below. After recovery from anesthesia, animals were allowed free movement in an unconfined environment.
Injection of SOX-6, 9ASCs into the knee joints of OA-induced goats
Animals were allocated to one of five groups (six goats in each group) at 4 months after OA induction as follows: Group 1—naive hASC group, animals were injected with 0.6 × 10 7 naive hASCs; Group 2—low-dose group, animals were injected with 0.18 × 10 7 SOX-6, 9ASCs; Group 3—mid-dose group, animals were injected with 0.6 × 10 7 SOX-6, 9ASCs; Group 4—high-dose group, animals were injected with 1.8 × 10 7 SOX-6, 9ASCs; and Group 5—negative control group, animals were injected with vehicle (PBS). Injection doses were calculated based on considerations of joint volume and cartilage sizes in goats and man. 21 All animals were sacrificed for analysis 5 months postinjection.
Autopsies and visual/histopathological examinations of treated joints
Atropine sulfate (0.1 mg/kg; Jeil Pharm., Daegu, Korea) was administered intramuscularly as a preanesthetic to prevent any respiratory distress. Animals were then anesthetized with an intraperitoneal injection of ketamine 2 mg/kg and xylazine 0.1 mg/kg and euthanized by injecting potassium chloride (2 mmol/kg; Sigma-Aldrich) intravenously. Femoral condyles and tibial plateaus were carefully and separately dissected without damaging cartilage surfaces. Harvested tissues were fixed in 10% formalin solution (Wako, Osaka, Japan) for 1 day, decalcified, and decalcified in 10% ethylenediaminetetraacetic acid in a 0.01 M phosphate buffer (pH 7.4) at 48°C for 3 weeks. After dehydration through a graded series of ethanol solutions, specimens were embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin (HE), fast green, and Safranin-O.
The Osteoarthritis Research Society International (OARSI) Scoring System for goats was used to grade the degenerative status of tissues.22,23 The system takes into account macroscopic findings of cartilage (score: 0–16) and osteophytes (score: 0–12) from 4 knee joint sites. Microscopic scoring was performed by evaluating cartilage structure (score: 0–10), chondrocyte density (score: 0–4), cell cloning (score: 0–4), interterritorial proteoglycan staining (score: 0–4), and tidemark integrity (score: 0–3).
Enzyme-linked immunosorbent assay results for inflammatory cytokines in goat synovial fluid
Synovial fluid (SF) samples were obtained from all knee joints during sacrifice and then centrifuged within 4 h at 10,000 g for 10 min and frozen at −70°C to −75°C. SF samples were also obtained from normal knee joints of six goats, which did not undergo any surgical procedure. Supernatants were assayed for goat tumor necrosis factor (TNF)-α and interleukin (IL)-1β using an Enzyme-linked Immunosorbent Assay Kit (MyBioSource, San Diego, CA). Well optical densities were read at 450 nm.
Statistical analyses
All quantitative data are expressed as the mean ± standard error of the mean of at least four independent experiments. Statistical comparisons between groups were made using one-way analysis of variance with Bonferroni's correction or using Dunnett's t test (SPSS 18.0; SPSS, Inc., Chicago, IL). Statistical significance was accepted for p-values <0.05.
Results
Construction of SOX-6, 9 MC vector
MC.RFP was used to measure transfection efficiencies achieved by electroporation because SOX-6, 9 MC vector lacks a fluorescent marker gene. FACS showed that transfection efficiency was 95.6% (Fig. 1B). Mean cell viabilities of RFP and SOX-6, 9 MC vector transfected cells were 92.67% ± 16.02 and 80.03% ± 13.83, respectively (Fig. 1C).
After transferring the SOX-6, 9 MC vector into hASCs, gene transfection was confirmed by examining changes in the messenger RNA (mRNA) expressions of SOX-6 and SOX-9. Both were elevated in SOX-6, 9ASCs than in negative controls (RFP-transfected hASCs) which showed low, endogenous gene expression of both factors. Western blot confirmed the expressional upregulations of SOX-6 and SOX-9 at the protein level (Fig. 1D).
In vitro chondrogenic potential of SOX-6, 9ASCs
hASCs underwent standard chondrogenic pellet culture. Pellet diameter sizes were statistically greater for SOX-6, 9ASCs than for hASCs treated with TGF-β2 and BMP-7 (p < 0.01, positive control) or hASCs treated with BM only (p < 0.05, negative control) (Fig. 2A). Proteoglycan productions, as measured by GAG/DNA ratio and Safranin-O staining, were comparable between SOX-6, 9ASCs and the positive control (Fig. 2B, C). In contrast, SOX-9 mRNA expression was >250-fold greater in SOX-6, 9ASCs than in the positive control. COL2A1 and aggrecan mRNA expressions were 3.5- (p = 0.004) and 1.4-fold (p = 0.058) greater, respectively, in SOX-6, 9ASCs than in the positive control (Fig. 2D). Western blot also confirmed the expressional upregulations of SOX-9, COL2A1, and aggrecan at the protein level (Fig. 2E and Supplementary Fig. S3). SOX-6, 9ASCs had comparable proteoglycan production to that of normal articular chondrocyte as observed by Safranin-O staining, while COL2A1 and aggrecan mRNA expression from SOX-6, 9ASCs tended to be higher but statistically insignificant than those from articular chondrocytes (Supplementary Fig. S4).

Chondrogenic potential of SOX-6, 9ASCs in vitro.
Retention time of SOX-6, 9ASCs in the knee joints of rats
Retention times of SOX-6, 9ASCs were investigated in rats by fluorescence imaging after IA injection. hASCs were labeled with two dyes, CellVue and DiD. The fluorescence signals of hASCs labeled with CellVue were undetectable at 7 days after injection, while those labeled with DiD produced fluorescent signals up to 28 days after injection (Fig. 3A).

Retention times and hASC biodistributions were measured after a single IA injection in SD rats.
A 56-day study of SOX-6, 9ASCs was also conducted to establish the biodistribution of cells following a single IA injection in SD rats. Analysis of PCR data showed the presence of human SOX-6 in bone marrow (in one of eight animals), lymph nodes (in one of eight animals), and heart tissues (in one of eight animals) on day 1, but not any other tissue except for right knees (the site of administration) on day 2 (data not shown). qPCR was also performed to detect the presence of human DNA from SOX-6, 9ASCs. Human Alu and SOX-6 gradually decreased until day 8 and were not detected at sites of administration on day 14 (Fig. 3B) nor was any positive signal or persistence of human Alu or SOX-6 in any other tissue or blood beyond day 2. These results indicated that injected hASCs gradually perished from joints.
To further analyze the mode of action of SOX-6, 9ASCs at arresting OA progression, we investigated whether the injected SOX-6, 9ASCs persisted in the joint cavity of OA-induced rats by detecting HN by IHC. The human cells (SOX-6, 9ASCs) were not detected in the joint at days 7 and 14 after SOX-6, 9ASC injection (Fig. 3C). These results suggest that the injected SOX-6, 9ASCs perished from the joint after probably exerting paracrine effects.
Gross findings of AC in the OA-induced goats treated with SOX -6, 9ASCs
AC statuses were assessed in both femoral and tibial sides of joint. Medial meniscectomy (MM) led to the development of OA lesions in medial knee joint compartments.
In negative control knees, only saline was injected. Three different doses of SOX-6, 9ASCs were injected, and to isolate the effect of hASCs per se, naive hASCs were also injected in one group. At 5 months after injection, OA changes, including denudation of AC and fibrillation of articular surface, were visible in the negative control. In the other groups, these changes were less pronounced, but to variable degrees, except in the high-dose group. The mid-dose group showed best AC preservation (Fig. 4A). OARSI macroscopic scores were significantly better for mid-dose SOX-6, 9ASCs than for negative controls in the femoral and tibial articular surfaces (Fig. 4B). At the time of sacrifice, joint fluid was collected and analyzed for the inflammatory cytokines IL-1β and TNF-α. TNF-α levels were significantly lower in the low-dose and mid-dose groups than in negative controls, and IL-1β levels were significantly lower in the mid-dose group than in negative controls (Fig. 4C).
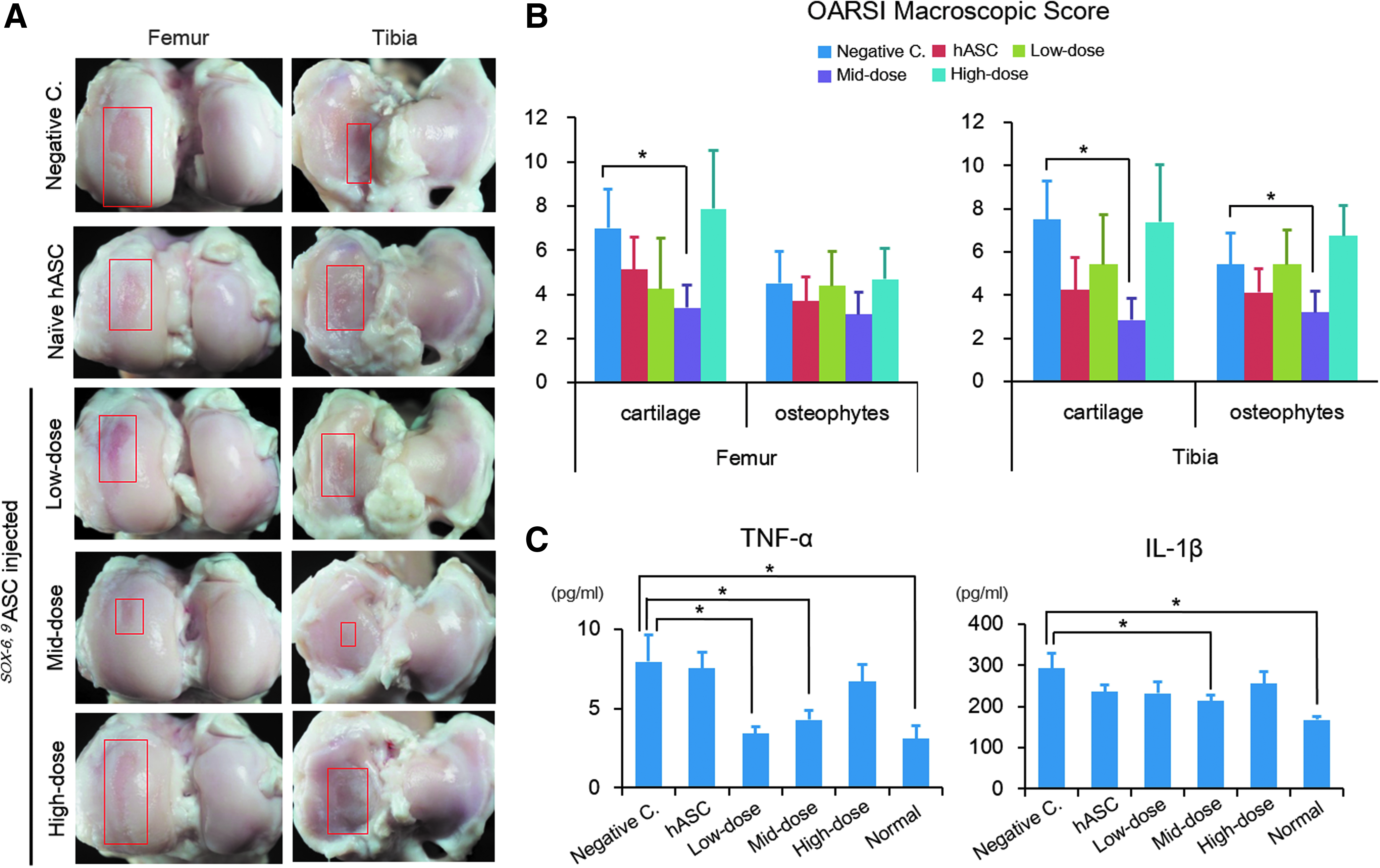
Gross findings of AC and inflammatory cytokine levels of the synovial fluids in OA-induced goats.
Histological findings of AC in the OA-induced goats treated with SOX-6, 9ASCs
H&E and Safranin-O staining was performed after decalcifying harvested goat samples. Overall results corroborated gross examination findings. Surface fibrillation and loss of proteoglycan were observed more obviously in negative controls than in the other groups except the high-dose group. The mid-dose group exhibited relatively intact smooth surface with good extracellular matrix status, showing less OA progression than the other groups (Fig. 5A and Supplementary Fig. S5). OARSI microscopic scores indicated that mid-dose group achieved best results, and these scores were significantly lower than those of negative controls regarding structure, chondrocyte density, cell cloning in femoral articular surfaces, and chondrocyte density and proteoglycan staining in tibial articular surfaces (Fig. 5B).

Histological analysis and OARSI microscopic scores of AC in the OA-induced goats. Negative controls were injected with vehicle (PBS), the hASC group was injected with 0.6 × 10
7
naive hASCs, the low-dose group was injected with 0.18 × 107 SOX-6, 9ASCs, the mid-dose group was injected with 0.6 × 10
7
SOX-6, 9ASCs, and the high-dose group was injected with 1.8 × 10
7
SOX-6, 9ASCs.
Discussion
Although nonviral gene delivery is generally less efficient than viral delivery for gene transduction, it is less immunogenic, which makes it a safer tool for gene transfer. 24 Electroporation, a physical gene delivery method, has been widely used to introduce genes into various types of cells in vitro. During this process, an electrical pulse is used to create transient pores in cell membranes, facilitating the spontaneous movement of exogenous DNA into cells.21,25
Our preliminary studies showed that transfection of SOX-6, 9 induced chondrogenesis to the same extent as the SOX-5, 6, 9 (Supplementary Fig. S1). While Yang et al. reported that single retroviral transduction of SOX-9 could induce ASC differentiation into chondrocyte-like cells, 26 our previous data clearly demonstrated that cotransduction of SOX-5, -6 together with SOX-9 led to much more effective chondrogenesis than single transfection of SOX-9 alone.14,16
Rat model was used for cell tracking because it was not possible to monitor injected cells in goats. Of note, injected cells gradually disappeared within 2 weeks of injection in knee joints of rats. IHC for HN in OA-induced rat samples also demonstrated the absence of injected SOX-6, 9ASCs. These results suggest that injected human ASCs disappear after exerting paracrine effects and that the differentiation of injected SOX-6, 9ASCs into articular chondrocytes did not occur. Given that this gene-cell therapy works in a paracrine effect mode, there is a question as to whether multiple injections are necessary to achieve best results. While most of reported cell injections to treat OA used a single injection, there is a report that multiple injections are necessary to maintain viable cells and inhibit OA progression by secretion of trophic factors. 27 Although multiple injections are likely to enhance the chondroprotective effect of treatment, increased costs and higher risks of immune reactions must also be considered. While the paracrine action of ASCs was not investigated in detail in the current OA study, it may be mediated by exosomes that reduce the production of inflammatory mediators from OA chondrocytes. 28 hASCs can also downregulate inflammatory factors (IL-1β) in the SF through paracrine pathway. 29
Reduction of IL-1β-mediated inflammation and oxidative stress may promote chondrogenesis-associated gene expressions in hASCs in OA environments. 30 Besides, when cocultured with OA chondrocytes, ASCs protect chondrocytes from degeneration associated with OA by reducing apoptosis and inducing hepatocyte growth factor. 31 In contrast, hASC-derived conditioned medium has a limited ability to decrease IL-1β, TNF-α, and matrix-degrading enzymes compared with ASCs per se. 32
The surgically-induced OA model used in the present study was established in our previous study, in which we compared MM alone with MM plus ACL transection. Contrary to expectation, MM alone more reproducibly led to the development of OA. 33 In a former study, we found that the effect of hASC xenografting to treat OA was not different between immunosuppressed and nonimmunosuppressed goats with respect to AC status and synovial inflammation, possibly due to the immune-protected status of synovial joints. 33 Good chondroprotective effects were also demonstrated with xenografting of SOX-6, 9ASCs in the current study, which would provide a supportive data for human trial in the future. At present, there is limited clinical proof of ASC therapy for OA because most studies are not well controlled and used stromal vascular fraction rather than cultured and characterized ASCs.34,35 A recent single center, randomized, double-blind, placebo-controlled, single ascending dose study showed that single injection of ASCs to OA knees led to significant symptomatic relief and arrested cartilage volume loss. 36 A systematic review of nonhomologous ASC therapies also showed improved functional, radiographic, and histological scores in OA with very few serious adverse events. 37 From our data, it is surmised that SOX-6, 9ASCs would be more effective than naive ASCs in human OA.
Overall the results obtained in the present study showed that a single injection of SOX-6, 9ASCs at a mid dose of 0.6 × 10 7 SOX-6, 9ASCs effectively arrested OA progression in surgically-induced OA and to significantly greater extents than SOX-6, 9ASCs at low dose of 0.18 × 10 7 SOX-6, 9ASCs. Interestingly, the high dose of 1.8 × 10 7 SOX-6, 9ASCs, as well as low dose, was less effective in this respect than the mid dose, indicating the presence of an optimal dose of SOX-6, 9ASCs for the treatment of OA. It is uncertain why SOX-6, 9ASCs at high dosage resulted in no effects. Melief et al. showed that ASCs secreted a higher level of cytokines (IL-6 and transforming growth factor-β1) implicated in the immunomodulation compared with BMSCs, correlating with higher metabolic activity of ASCs which may lead to imbalance of cartilage metabolism. 38 In addition, Pigott et al. reported that greater level of inflammatory cytokine was released from peripheral blood mononuclear cells when xeno-MSCs were cocultured than when auto- or allo-MSCs were cocultured. 39 We speculate that the greater level of inflammation within the synovial joint in the high-dose group than in the mid-dose group may be associated with inferior gross and histological results in the group.
While this study is a large-animal study that demonstrated effectiveness and safety, as well as mechanism of action in IA SOX-6, 9ASC therapy, there are shortcomings. First one includes inability to perform IHC in histological sections for inflammatory cytokines and enzymes related with OA pathogenesis because primary antibodies for goats were not available. In addition, to further investigate mode of action, isolation and IA injection of SOX-6, 9ASC-derived exosomes would have confirmed the pilot observation results of high-dose ASC treatment. In addition, the effect of multiple IA injection which was reported to be safe in an allogeneic equine model may be investigated in future studies. 40
In conclusion, SOX-6, 9-transfection led to in vitro chondrogenesis of ASCs comparable to that achieved by growth factor treatment. IA injection of SOX-6, 9 ASCs in optimal dose reduced the progression of surgically-induced OA in goats. We suggest that IA SOX-6, 9ASCs be considered a novel potential strategy to treat OA.
Footnotes
Acknowledgment
This study was supported by the National Research Foundation (NRF) funded by the Korean Government (2017M3A9B4064777, 2016R1C1B1008616).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.