
Abstract
Although mouse models of epithelial–mesenchymal interaction-based whole-tooth regeneration have been generated, whether this strategy can be applied to humans remains unknown and preclinical investigations using a large animal model are required. In this study, we describe the development of feasible methods for whole-tooth regeneration in a swine model. We characterized the odontogenesis of dissociated single cells from early-stage tooth germs from miniature pigs (minipigs) in vitro by tracking the morphogenesis of reassociated cell pellets in an organ culture system. Despite loss of positional information, reassociated individual dental epithelial and mesenchymal cells retained their odontogenic potential and continued to recapitulate typical dental histogenesis and morphogenesis. Based on the developmental biology of natural teeth, we allotransplanted reassociated tooth germs into the jawbones of minipigs and manipulated whole-tooth regeneration by mimicking the process of tooth development. To overcome the challenge of alloimmune rejection, bone marrow mesenchymal stem cells were systemically infused, along with local aspirin administration, during allotransplantation. Radiographic data confirmed the survival and growth of the explants in minipig jawbones. Histological analysis revealed that the explants formed well-developed tooth structures and contained normally arranged dental components, including organized dentin, cementum, periodontal membrane, dental pulp, vasculature, and nervous tissue, closely resembling those of natural teeth. In conclusion, this study achieved orthotopic whole-tooth regeneration and development in a large mammal using a cell reassociation approach, supporting the potential for regeneration of human teeth in situ.
Impact Statement
The methods developed in this study to manipulate pig tooth germ cells in vitro and in vivo provide a reference for studying whole-tooth regeneration and tooth development in large animals. Of importance, compared with conventional ectopic tooth regeneration, conducted in the omentum, subcutaneous tissues, or kidney capsule (among other locations) with low with immune reactivity in rodent models, this study achieved orthotopic regeneration and development of whole teeth in a large mammal, representing a large stride toward the realization of tooth regenerative therapy for humans with missing teeth.
Introduction
Tooth loss, caused by caries, periodontitis, tumor, trauma, or aging, is a very common disorder that causes physical and mental suffering in humans. Contemporary repair treatments, either using dentures or (currently preferred) implants, have some strict indications and several insurmountable physical or chemical limitations,1,2 such as failure to remodel with surrounding bone, which must progress continuously in natural teeth throughout an individual's lifetime. 3 Recent breakthroughs in dental regenerative bioengineering have lent significant credibility to the idea that a third dentition, after the loss of deciduous and permanent teeth, that is structurally, functionally, and aesthetically comparable with natural teeth, will be available to humans.
Despite complete loss of positional information, reassociated individual dental epithelial and mesenchymal cells from mouse embryos can continue to develop, producing tooth-like structures, when cultured in vitro, or form roots and periodontal ligament connected to developing bone, when implanted in vivo.4,5 These findings suggest that cell history is not inherent and that reassociated epithelial and mesenchymal cells from early-stage tooth germs can retain their odontogenic potential and continue to recapitulate typical dental histogenesis and morphogenesis.
Furthermore, when bioengineered tooth germs, constructed by reassociation of epithelium and mesenchymal cells, were placed in the alveolar bone in the region of a lost tooth, they could develop into fully functioning teeth in adult mice.6,7 These epithelial and mesenchymal interaction-based whole-tooth regenerative approaches in rodents, which mimic the process of tooth development, hold great promise as a strategy for developing a functional substitute for lost teeth.
However, the molecular mechanisms involved in human odontogenesis are much more complex than those of rodents and current understanding of their molecular basis remains insufficient to allow de novo tooth regeneration in humans. For example, cell source, morphological patterns, and size control, the length of time required for growth and issues of immune rejection also need to be addressed. The availability of autologous embryonic tooth germ cells for human tooth regeneration is limited and inaccessible, whereas allogeneic or xenogenic tooth germ cells may elicit immune rejection. 3 Moreover, precise control of the morphology and size of regenerated teeth has not been successfully achieved to date. 8 The long development and growth time make the study of tooth regeneration difficult and, clinically, increase the duration of tooth loss for patients. Therefore, the feasibility of de novo odontogenesis in humans is yet to be established. 9
An essential step is to move tooth regeneration strategies from rodents to a large animal model, before their introduction in humans. The miniature pig has been proven to be an attractive alternative to rodents for understanding mechanisms underlying tooth development in large mammals,10–14 and is promising for the study of human disease and preclinical therapies,15,16 owing to its many similarities to humans, including the morphology, number, and size of its teeth. In particular, it has heterodont and diphyodont dentition, which are not available in rodents.17–20
In this study, we used minipigs as animal models to investigate whole-tooth regeneration in situ using an allogeneic cell reassociation strategy. First, single epithelial and mesenchymal cells from cap stage tooth germ were dissociated and reconstituted into reassociated tooth germs. These were then cultured in a high-throughput suspension culture system, loaded with aspirin. Finally, cultured germs were transplanted into the jawbones of host minipigs pretreated using bone marrow mesenchymal stem cell (BMMSC) infusion. This pilot study establishing whole-tooth regeneration in the jawbone of a large animal model provides a reference for the study of human tooth regeneration in situ.
Materials and Methods
Animals
Pregnant minipigs and the adult host minipigs (∼1.5 years old) were obtained from the Institute of Animal Science of the Chinese Agriculture University (Beijing, China). Minipig embryos were obtained as reported previously. 18 In brief, minipig pregnancies were verified by B-type ultrasonic inspection, and staged minipig embryos were obtained by cesarean section.
Adult host minipigs were kept under conventional conditions with free access to water and a regular supply of soft food. The soft food was a mixture (4:3) of water and pig feed, which included corn powder, soybean flour, and other nutritional additives (Zhu Siliao, Beijing Keao Xieli Feed, China). Extraction of anterior and premolar teeth had minimal effect on the ability of minipigs to eat soft food. The weights of host minipigs were recorded at various stages during the study (Supplementary Table S1). All experimental animal procedures were reviewed and approved by the Animal Care and Use Committee of Capital Medical University (Permit Number: CMU-2012-x-102), and procedures were conducted in accordance with the approved guidelines.
Isolation of second deciduous incisor germs from minipigs
Second deciduous incisor germs (i2) at the cap stage were isolated from a single litter of minipig embryos, staged at embryonic day (E) 50, and pooled under stereo microscopy using an attached Olympus DP72 digital camera system (Olympus Corporation, Tokyo, Japan) (Supplementary Fig. S1 and Fig. 1A). At E50, the morphological stage of the i2 germs corresponded to the cap stage. 18

Dissociation, identification, and reconstitution of single tooth germ cells from deciduous incisor tooth germs of minipig embryos.
Dissociation and identification of single tooth germ cells
Single tooth germ cells from minipigs were obtained as previously reported.5,21 In brief, epithelium and mesenchyme isolated from second deciduous incisor germs were incubated in phosphate-buffered saline (PBS) containing Dispase II (1.2 U/mL; Sigma-Aldrich, St. Louis, MO) and DNase I (20 U/mL; Takara Bio, Shiga, Japan) for 15 min at room temperature, then separated under a stereo microscope. Then, the epithelium and mesenchyme were each dissociated into single cells in PBS supplemented with collagenase type I (3 mg/mL; Sigma, Worthington Biochemical Corp., Lakewood, NJ) and Dispase II (4 mg/mL; Sigma, Roche Diagnostics Corp., Indianapolis, IN), and filtered through a 70 μm cell strainer (BD Biosciences, San Jose, CA).
To examine the purity and origin of the mesenchymal and epithelial single cells, the percentages of cells that stained positive for cytokeratin 14 (CK14; epithelium) and vimentin (mesenchyme) (Fig. 1B) were assessed using an FACSCalibur flow cytometer (BD Biosciences, San Jose, CA) and analyzed using CellQuest software (BD Bioscience) (Fig. 1C).
For immunocytochemistry, dissociated single cells were seeded in 12-well plates (Corning Costar Corporation, NY) at a density of 2 × 105 cells per well, and cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY) supplemented with 20% fetal calf serum (Gibco) and 100 U/mL penicillin/streptomycin (Gibco). After 2–3 days, cells were washed with PBS and fixed using 4% paraformaldehyde. After fixation, cells were blocked and permeabilized for 1 h at 37°C with 5% bovine serum albumin in the presence of 0.1% Triton X-100. Subsequently, cells were incubated with the indicated primary antibody (anti-CK14, 1:100, sc-53253 or anti-vimentin, 1:100, sc-73258; both Santa Cruz, CA) overnight at 4°C. Cells were then washed with PBS and incubated for 1 h at 37°C with appropriate secondary antibodies: for anti-CK14, anti-mouse immunoglobulin G (IgG) Alexa Fluor 647 conjugate (1:1000; 4410S, Cell Signaling Technology) and for anti-vimentin, anti-mouse IgG Alexa Fluor 647 conjugate (1:100; 4408S, Cell Signaling Technology), followed by DAPI for 20 min. Cells were washed in PBS before analysis.
Reconstitution of tooth germ cells and tissue culture in a three-dimensional suspension culture system
Single cells of epithelial and mesenchymal origin were pelleted by centrifugation at 1000 rpm for 5 min. Bioengineered tooth germs were constructed, followed by the sequential injection of 0.25 μL mesenchymal and 0.25 μL epithelial single-cell pellets into a 50 μL collagen gel drop (Cellmatrix type I-A; Nitta Gelatin, Osaka, Japan) to reassociate both pellets (1:1 ratio, each pellet including ∼1 × 105 cells) and to enable compartment contact (Ep-Me) between the two pellet types, to track reciprocal interaction between epithelial and mesenchymal cells. Intact cap stage second deciduous incisor germs served as controls.
Initiation of odontogenesis by reassociated epithelial and mesenchymal cells takes 7–8 days when cultured in vitro,
21
whereas intact tooth germs, representing tissue capable of odontogenesis, do not require long-term culture; therefore, explants were cultured for 7 days (reconstituted explants) or 3 (intact explants) days in 12-well transwell plates (3493; Corning Costar Corporation), as given in Figure 1D and E, in a universal cell incubator (Memmert INCO2; Memmert, Germany) at 37°C in a 5% humidified CO2 and 21% O2 atmosphere. They were cultured in DMEM (Gibco) supplemented with 20% fetal calf serum (Gibco), 100 U/mL penicillin/streptomycin, and 0.18 mg/mL
Isolation and infusion of allogeneic BMMSCs
Allogeneic BMMSCs were obtained and expanded from minipigs, as reported previously. 22 In brief, bone marrow was collected from the femur and tibia of a 2-week-old minipig. Nucleated cells were isolated by Ficoll-Hypaque (Sigma-Aldrich) density gradient centrifugation. Mononucleated cells were incubated in minimal essential medium (α-MEM; Invitrogen), supplemented with 10% fetal calf serum (Gibco), 100 U/mL penicillin/streptomycin, and 2 mM glutamine (Invitrogen) at 37°C in a 5% humidified CO2 atmosphere. To confirm mesenchymal stem cell characteristics, flow cytometric analysis based on surface markers, including CD29, CD73, CD90, CD105, CD14, CD31, CD34, and CD45, and adipogenic or osteogenic differentiation assays were performed (Supplementary Data and Supplementary Fig. S2).
To compare the immunoregulatory potency of BMMSCs alone and a combination of BMMSC and aspirin administration, six normal minipigs with tooth germs transplanted in the maxillary bone were randomly divided into three equal groups. One group was intravenously infused with BMMSCs ( × 106/kg body weight) 23 ; another group was locally administered with aspirin (100 μg aspirin contained in gel-foam) 24 at the maxillary bone, along with the BMMSC intravenous infusion; and the third group was administered neither BMMSCs nor aspirin, as a control. The results illustrated that pigs receiving the combined BMMSC–aspirin treatment exhibited an obvious increase in CD4+ CD25+ FoxP3+ T regulatory cells (Tregs) and a reduction in CD3+ T cell numbers (Supplementary Fig. S3). Based on this finding, the BMMSC–aspirin combination was used, rather than BMMSC alone, in subsequent experiments.
For the subsequent experiment, seven new minipigs were used. Four host minipigs were infused with BMMSCs (2.68–3.61 × 106/kg) 1 day before explant transplantation, along with local administration of aspirin (Supplementary Table S2), whereas three pigs were infused with 0.9% normal saline as controls. The percentages of CD3+ T cells and CD4+ CD25+ FoxP3+ Tregs in peripheral blood were examined by flow cytometry to monitor the immunological status of the host minipigs. We obtained blood samples from the host for biochemical analysis and the results illustrated that BMMSC infusion did not significantly alter the biochemistry profile of the minipigs (Supplementary Fig. S4).
Transplantation of intact and reconstituted explants into minipig jawbones
Three months before transplantation operations, permanent maxillary anterior teeth and premolars were completely extracted from both sides of the seven host minipigs (the same minipigs as those used in the previous paragraph), and computed tomography (CT) scans were conducted before the transplantation procedure to confirm that no tooth residue was present and that no new tooth tissue had developed after extraction.
During transplantation, hosts were deeply anesthetized with a combination of 6 mg/kg ketamine chloride (H35020148; Gutian, Fujian, China) and 0.6 mg/kg xylazine (070031582; Shengda, Jilin, China) by intramuscular injection and placed in the supine position. Transplant sites were cleaned and sterilized, and then a crestal incision (∼4 cm in length) was made with a fine scalpel and a full thickness flap was elevated. The extraction area of the host maxilla was exposed and a bony hole was prepared using an MD20 Implant System (NOUVAG AG, Switzerland). A titanium nail was implanted close to each bony hole as a location marker. Next, intact second deciduous incisor germs and reassociated tooth germs were transplanted into one side of the bony hole and covered with gel-foam containing aspirin (100 μg in each implant hole). The type of explant (intact or reassociated) in each implant hole of the seven minipigs was recorded (Supplementary Table S3). Finally, the implantation area was completely covered with oral mucosa and sutured (Fig. 2). Maxillary extraction sites on the other side of the jawbones of the seven host minipigs were used as negative controls.
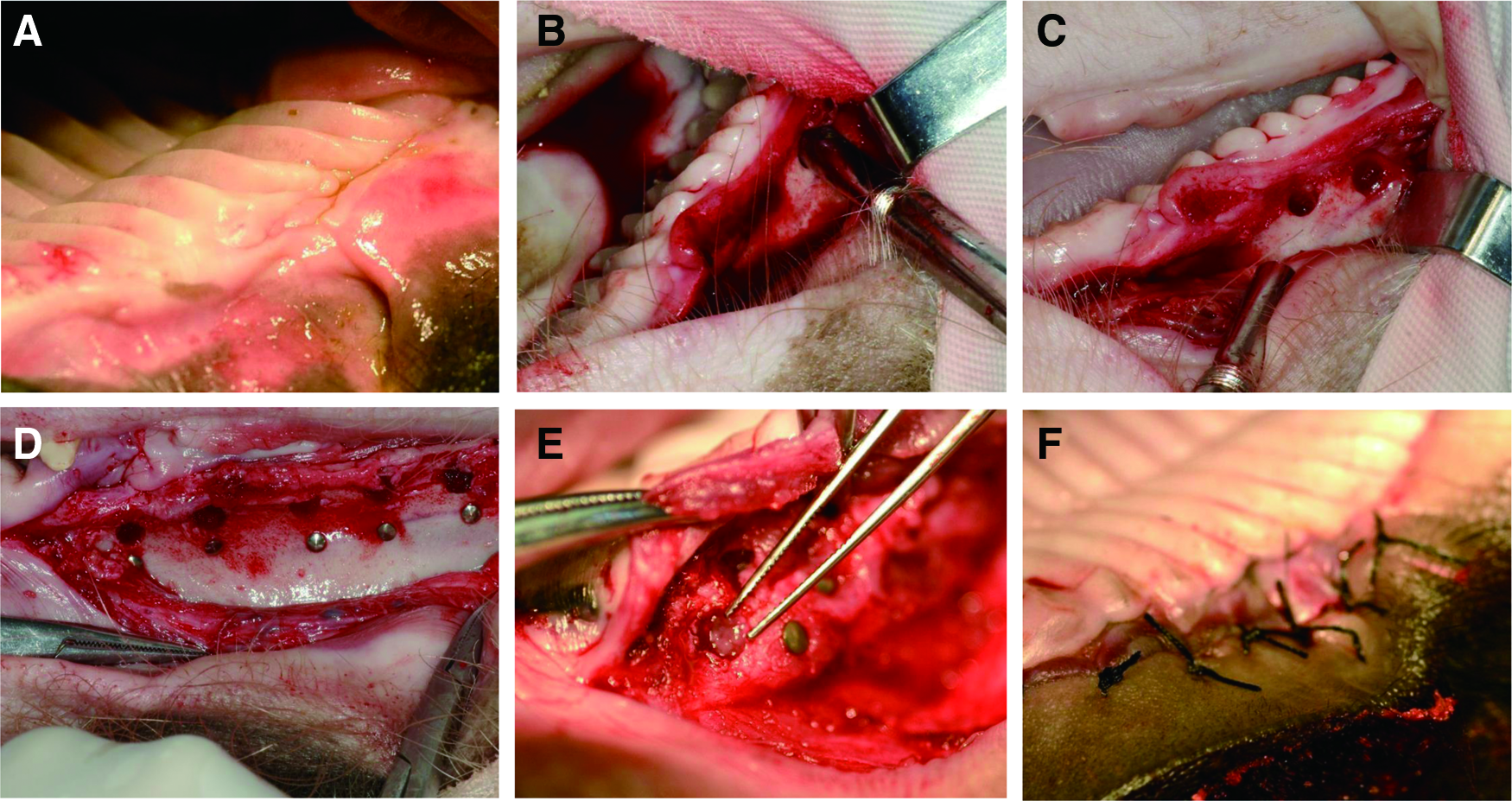
Transplantation surgical procedures of tooth germs into the minipig jawbones 3 months after extraction.
Radiographic analysis
Host minipigs were examined using 64-multidetector spiral CT (Siemens, Germany) or a CBCT device (NewTom 3G tomography; Summer, Italy) before extraction, 3 months after extraction, and on days 50 and 150 after transplantation. To generate three-dimensional (3D) anatomical conformation images of explants, CT data from day 150 posttransplantation were imported into Mimics 10.0 software (Materialise N.V., Belgium).
Histochemical and immunohistochemical analyses
For histological examination, specimens were stained with hematoxylin–eosin (H&E). To show the periodontal membrane and vascularization or innervation of the regenerated tooth, samples were immunostained with vimentin and CD31 or neurofilament, respectively. Detailed methods are provided in Supplementary Data.
Statistical analysis
Immunological data were analyzed using an independent samples t-test, with a 95% confidence interval. Values of p < 0.05 were considered significant.
Results
Single-cell dissociation from epithelium and mesenchyme
We isolated cap stage second deciduous incisor germs from a litter of minipig embryos (E50 stage) and separated the epithelium from the mesenchyme (Fig. 1A). After sequential enzymatic treatment, epithelial and mesenchymal tissues were dissociated into single cells. Epithelial cells were positive for CK14 (purity, 89.05% ± 1.66%) and mesenchymal cells were vimentin positive (purity, 87.21% ± 2.23%), suggesting that the dissociated cells contained few contaminating cells and were suitable for use in subsequent experiments (Fig. 1B, C).
Tissue culture of reassociated and intact tooth germ in vitro
To characterize the odontogenesis of single cells dissociated from minipig tooth germs, we tracked the in vitro morphogenesis of reassociated cell pellets with epithelial cell and mesenchymal cell compartment (Ep-Me) in a transwell organ culture system. Epithelial and mesenchymal cells could self-sort and self-aggregate to form epithelial and mesenchymal cell masses, respectively. Interestingly, epithelial masses could polarize and organize into well-defined dental epithelial structures surrounding the mesenchymal cell mass, like a natural tooth germ structure (Fig. 1E). These interactions were observed after 3 days of culture, and morphogenesis became apparent on days 7–8, indicating the initiation of odontogenesis.
Monitoring of immune responses
Immunological assessment of host minipigs suggested that percentage of CD3+ T cells decreased, whereas that of CD4+ CD25+ FoxP3+ Tregs exhibited a significant increase in the peripheral blood after administration of BMMSCs and aspirin, which may have contributed to the survival of the allogeneic tooth germ cells (Fig. 3).
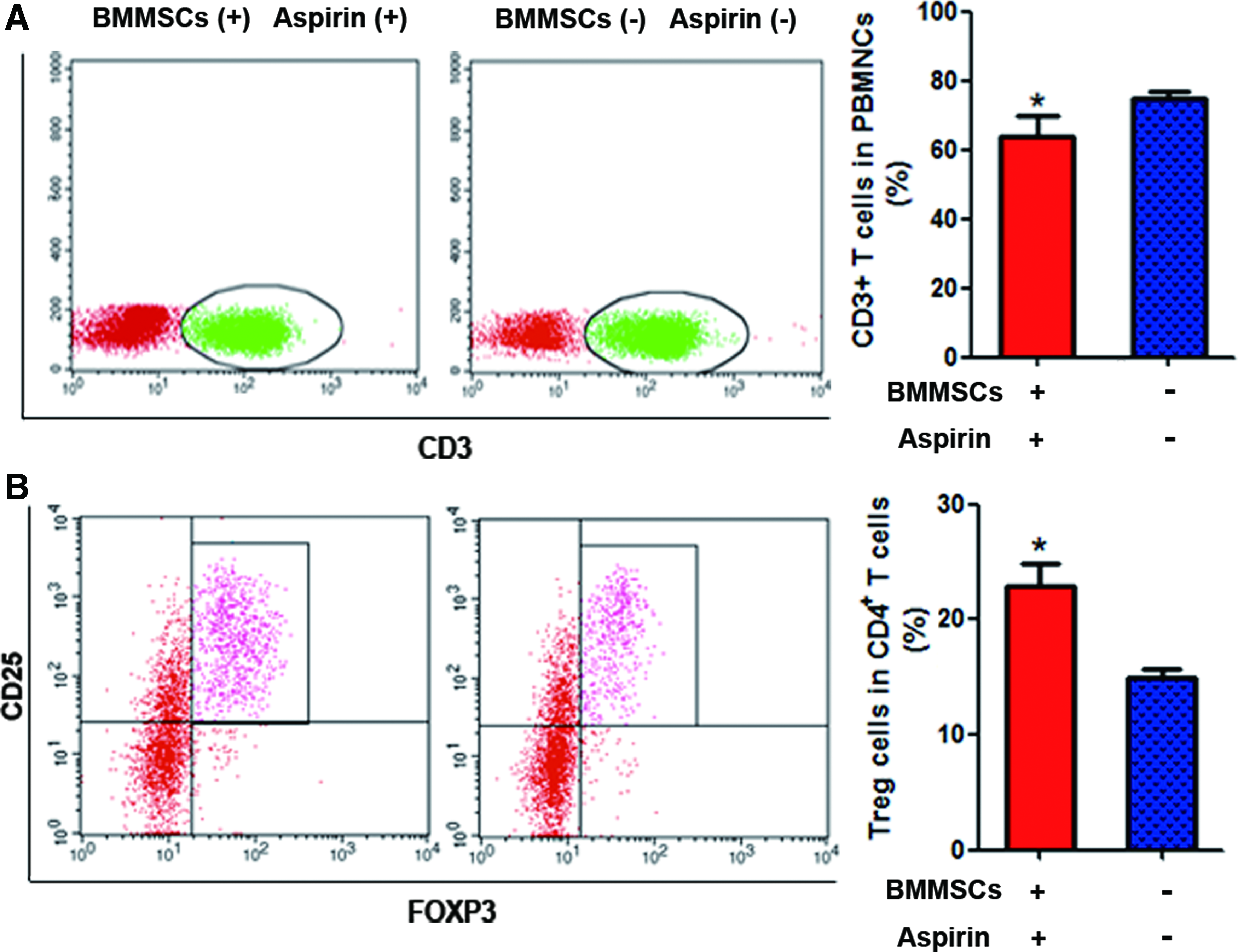
Immunological assessment of the host minipigs pretreated with or without BMMSC infusion and aspirin administration. Evaluation of CD3+ T cells in the PBMNCs and CD4+CD25+FoxP3+ T regulatory (Treg) cells in CD4+ cells of minipigs after infusion of allogeneic BMMSCs and administration of aspirin, BMMSCs (+) aspirin (+) group (n = 4), or without BMMSC infusion or aspirin administration, BMMSCs (−) aspirin (−) group (n = 3). Percentage of CD3+ T cells decreased
Radiographic analysis of the growth of reassociated and intact explants in minipig jawbones
To explore whether explants could survive and grow in the jawbone of a large animal model, we transplanted reassociated and intact tooth germs into minipig jawbones (Fig. 4A). The teeth in the canine and premolar region (Fig. 4B, preextraction) designated for transplantation had been completely removed and there were no tooth tissues in the transplantation region before the transplantation procedure (Fig. 4B, 3 months postextraction; Fig. 4E, 3 months postextraction; Fig. 4G, 3 months postextraction; Fig. 4I, 3 months postextraction). On day 50 posttransplantation, holes containing transplanted tooth germs had not been replaced by bone remodeling and were visible between the titanium nails (Fig. 4B, 50 days posttransplantation).

Radiographic analysis on the growth of reassociated and intact explants in the minipig jawbones.
By day 150 posttransplantation, three regenerated teeth had developed from reassociated explants in the four minipigs that had undergone BMMSC infusion with aspirin administration (Fig. 4B, 150 days postextraction; Fig. 4E, 150 days postextraction; Fig. 4G, 150 days postextraction). One of the developing teeth (unerupted) was observed in the posterior part of the jawbone. A calcified tissue with a shape similar to that of an anterior tooth was noted as a radio-opaque region between the third and fourth (from anterior to posterior) titanium nails in the premolar region, where a reassociated tooth germ had been transplanted. No developing teeth were detected in the anterior part of the alveolar bone in this minipig, where intact tooth germs had been transplanted (Fig. 4B, 150 days posttransplantation). In addition, there was no evidence of tooth regeneration in the alveolar bone control sites, where teeth had also been extracted and sham operations (without tooth germ transplantation) were conducted (Fig. 4B, no transplantation).
To obtain more precise images of the explants, we examined the maxilla in vitro using CBCT. Evaluation of a CBCT coronal section revealed that the cross-section of the unerupted explant was elliptical and highly mineralized, with a low-density area in its center, resembling a pulp cavity (Fig. 4C). CT scan data from day 150 posttransplantation were used to construct a 3D model of the explants in host minipigs. A 3D image of the unerupted explant revealed a tooth-like structure, with a relatively large crown and a slim and cuspate root (resembling a unicuspid, unirooted tooth) lying horizontally between the third and the fourth (from anterior to posterior) titanium nails (Fig. 4D).
Two other calcified tissue areas were detected in the anterior part of the alveolar region in two other minipigs; one resembled an erupting tooth, whereas the other had erupted (Fig. 4E–H). Radiographic data indicated that the explant had the same density and a similar shape to a natural anterior tooth. In the control group, no regenerated tooth structures were detected in any of the minipigs that had not received BMMSC and aspirin administration (Fig. 4I, J).
Overall, in this study, of four minipigs that underwent BMMSC and aspirin treatment, we successfully achieved whole-tooth regeneration in three grafts from reassociated tooth germs, and the success rate of regenerative teeth in total reassociated explants was 3/6 (50%) in this group. In contrast, no intact tooth germs and none of the three minipigs that did not receive BMMSCs and aspirin developed any regenerated teeth. More details about the explant survival rate is summarized in Table 1.
Number of Survived Teeth in Different Groups
There were only 7 minipigs available, so we divided them randomly into two groups, one group 4 and the other 3. As this study was to explore a method for tooth regeneration, in order to balance the difference and compare the explant survival rate, we transplanted an equal number of tooth germs in the two groups.
BMMSCs, bone marrow mesenchymal stem cells.
Histological analysis of the growth of reassociated and intact explants in the minipig jawbones
Histological analysis revealed that explants formed well-developed tooth structures. These contained normally arranged dental components, including organized odontoblasts, dentin, cementum, periodontal membrane, dental pulp, vasculature, and nervous tissue, very closely resembling the structure of natural teeth (Fig. 5A, B, G–I). There was no dental structure detected in sites where no tooth or intact tooth germs were transplanted in the minipigs with BMMSC and aspirin administration. Moreover, all the explants (either the reassociated or intact) failed to develop into any teeth in the minipigs without BMMSC and aspirin administration. Instead, bone regeneration was observed (Fig. 5C–F). These results demonstrate that reassociated explants with epithelial and mesenchymal cell compartmentalization possess optimal whole-tooth odontogenetic potential.
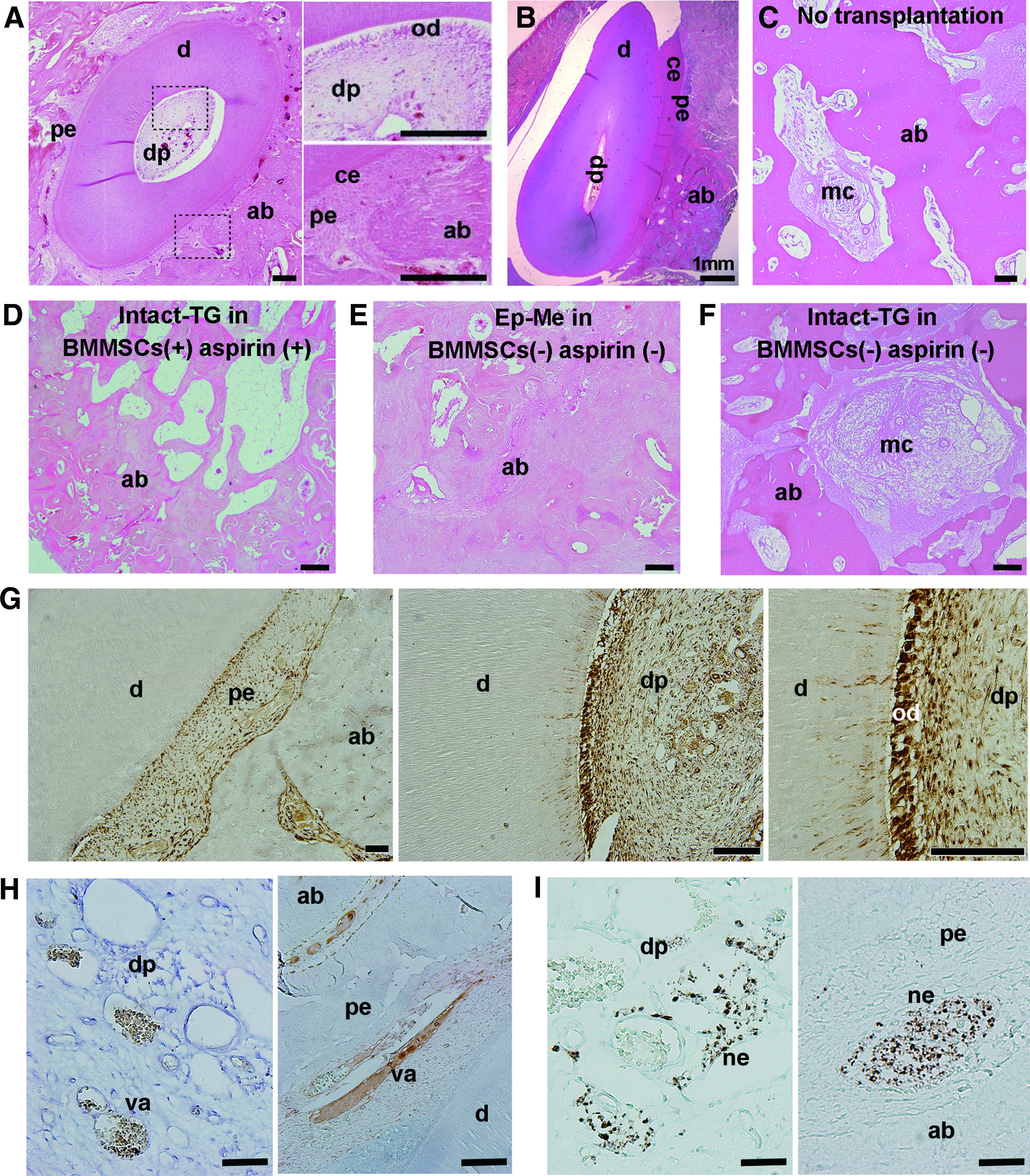
Histological analysis on the growth of reassociated and intact explants in the minipig jawbones.
Discussion
In this study, we characterized the de novo odontogenesis of minipigs in vitro and developed practical methods for whole-tooth regeneration in the jawbone of minipigs based on tooth developmental biology. Our findings illustrate the conservation of odontogenesis and feasibility of whole-tooth regeneration in a large animal model.
This successful attempt at de novo odontogenesis in minipigs indicates the achievability of developing whole teeth with correct dental components and structure from tooth germs generated from compartmentalized epithelial and mesenchymal cells. To better determine whether regenerated teeth originated from transplanted tooth germs, we extracted all the teeth from the transplantation region and verified their complete removal by CT scan 3 months after extraction. To further confirm complete tooth extraction, no tooth germ was transplanted in the contralateral side during transplantation operations. Accordingly, no tooth tissue developed in the contralateral side, indicating that teeth were indeed completely extracted. Furthermore, we also transplanted some deciduous incisor tooth germs as explants in the maxillary premolar region. Similar to molar teeth, the premolar teeth of minipigs are multicuspid and multirooted; however, the harvested tooth was unicuspid and unirooted, also supporting its origin as donor derived.
Compared with previous studies using immune-deficient mice, whole-tooth regeneration in minipigs or human faces is considerably more challenging owing to the possibility of immune rejection. In this study, three of four minipigs administered BMMSC and aspirin treatment achieved successful allogeneic cell reassociation-based whole-tooth regeneration, whereas there was no regeneration in any of three controls (no BMMSC or aspirin administration). The successful regeneration of teeth from reassociated explants may be attributable to the immunomodulatory function of aspirin and BMMSCs and the lower immunogenicity of reassociated explants, compared with intact tooth germs. BMMSCs have confirmed immunomodulatory function,25–27 and have been used to reduce transplantation rejection28–30 and treat autoimmune diseases. 31 Therefore, the BMMSCs infused into host minipigs likely functioned as a systemic immunomodulator, favoring the survival of explants during allograft.
Aspirin is reported to have important dose-dependent systemic and local effects on immunoregulation. Daily aspirin administration (50–500 mg) for several months has been used in renal allograft to inhibit cyclooxygenase activity and the nuclear factor κB pathway.32,33 Aspirin (at the mmol level)-treated dendritic cells upregulate leukocyte immunoglobulin-like receptor 3 (ILT-3) expression and induce hyporesponsiveness in responder T cells.34,35 Aspirin pretreatment (200 μg/mL) can significantly enhance the immunomodulatory function of BMMSCs by upregulating Tregs and downregulating Th17 cells. 36 Besides these systemic effects, local administration of 100 μg aspirin could significantly improve BMMSC-based bone regeneration through reduction of local interferon γ and tumor necrosis factor α concentrations. 24
Given the systemic effects of BMMSCs and local effect of aspirin, in this study, tooth germ explants were pretreated with aspirin (during culture in vitro and transplantation in vivo), along with infusion of BMMSCs to decrease the immunogenicity of explants, which induced immune tolerance in the periphery blood. The results of this study imply that allogeneic regeneration of teeth in a large animal model is feasible through suppression of host immune rejection using BMMSCs combined with aspirin; however, further studies with more comprehensive design are warranted. In particular, with the aim of enhancing their potential to induce immune tolerance and improve explant survival rates, pretreatment of BMMSCs with aspirin before their infusion should have to be explored. Moreover, to rule out the possibility that explant survival is entirely the result of the local effects of aspirin on the mesenchymal component of tooth germs, it would be beneficial to include another treatment group with local aspirin alone.
We analyzed possible reasons for the failure of the tooth regeneration from some reassociated and all intact tooth germs. Regarding reassociated tooth germs, the ratio of epithelial and mesenchymal cells used for reassociation was pivotal and may have been a major reason for the low explant survival rate. The use of a 1:1 ratio of epithelial and mesenchymal cells in reconstituted explants in this study was based on the ratio used in previous investigations.7,21 Epithelial and mesenchymal cells from swine tooth germs were reconstituted using a 1:1 ratio, and the resulting explants succeeded in regenerating whole teeth when transplanted into mouse subrenal capsules and jawbones. 21 However, other ratios of epithelial and mesenchymal cells have not been explored, and further study to determine the optimal ratio of epithelial and mesenchymal cells is required to improve the success rate.
The failure of intact tooth germs to develop may be attributable to nutritional and blood deficiency or homograft rejection. Such explants are larger; thus, it is more difficult for nutrition and blood to reach the center of intact germs. 37 Furthermore, as tissues include a substantial amount of extracellular matrix, intact tooth germs are more immunogenic than reassociated explants consisting of epithelial and mesenchymal cells.
At present, ectopic tooth regeneration in physiological niches such as in the omentum, embryonic diastema region, anterior eye chamber, subcutaneous tissues, and kidney capsule, is predominant,38,39 as those ectopic sites are rich with nutrients (or blood) and exhibit low levels of immunoreactivity. By overcoming the challenge of immunological rejection, the results presented in this study take a further major step toward achieving the orthotopic regeneration of whole teeth in large animals.
Collectively, the methods developed in this study to manipulate swine tooth germ cells in vitro and in vivo provide a reference for the investigation of whole-tooth regeneration and tooth development in large animals. More importantly, compared with previous studies of ectopic tooth regeneration in rodents, this investigation achieved orthotopic whole-tooth regeneration in a large mammal, representing another large stride forward in the realization of tooth regenerative therapy for missing teeth in humans.
Footnotes
Acknowledgments
The authors thank Dr. Xiao, J. for the helpful suggestions. This work was supported by the National Natural Science Foundation of China (91649124 to S.W., 81600883 to Z.W., and 81771032 to F.W.), Beijing Municipal Science & Technology Commission No.Z181100001718208, and Beijing Municipality Government grants (Beijing Scholar Program- PXM2018_014226_000021, PXM2018_193312_000006_0028S643_FCG, PXM2017_014226_000023, PXM2016_014226_000034, and PXM2016_014226_000006 to S. W.).
Disclosure Statement
This text is part of our actual manuscript file. The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.