
Abstract
Although skeletal muscle is capable of complete recovery after an injury, specific situations require support or acceleration of this process, such as in the elderly and athletes, respectively. Skeletal muscle regeneration is due to muscle stem cells (MuSCs) that undergo adult myogenesis, a process sustained by MuSC environment. Although recognized as important, extracellular matrix (ECM) has been overlooked in this process. Matrix-based therapy aims at improving ECM remodeling to support tissue repair. In this context, we investigated the properties of a single injection of the clinical grade glycosaminoglycan mimetics RGTA® (ReGeneraTing Agents) on skeletal muscle regeneration in a context compatible with a clinical application, that is, 3 days after the injury. Our results show that RGTA-treated muscles showed an increase of the number of myonuclei in regenerating myofibers and an increase of the capillarization of the new myofibers. In vitro experiments showed that RGTA directly acts on MuSCs by stimulating their fusion into myotubes and on endothelial cells by stimulating the formation and maturation of vessels in a 3D culture setup. These results indicate that a single administration of RGTA in regenerating muscle stimulated both myogenesis and angiogenesis, thus accelerating skeletal muscle regeneration.
Impact Statement
Although highly powerful in normal condition, postinjury skeletal muscle regeneration is less efficient in some situations, such as obese, elderly, or resting people. In other context, such as high-performance sport, skeletal muscle regeneration must be shortened but in a way ensuring a full functional recovery. In this context, our results show that a single injection of the clinical grade glycosaminoglycan mimetics RGTA® (ReGeneraTing Agents), in a context compatible with a clinical application, that is, 3 days after the injury, is beneficial for skeletal muscle regeneration, through the stimulation of both myogenesis and angiogenesis.
Introduction
Adult skeletal muscle is highly adaptable to the physiological demand. Moreover, skeletal muscle is capable of complete recovery after an injury, thanks to the activation of muscle stem cells (MuSCs) that activate, expand, differentiate, and finally fuse to form new functional myofibers. 1 While historical investigations have focused on MuSCs, which are indispensable for muscle regeneration, more recent studies have evidenced that their environment is equally important for the efficacy of the regeneration process.2,3 MuSC environment encompasses several cell types as well as the extracellular matrix (ECM), which has been overlooked in the regeneration process. Indeed, much attention has focused on the pathological development of excessive ECM, which is fibrosis, 4 but the physiological roles of ECM remodeling remains poorly understood. The mechanisms by which ECM components support skeletal muscle regeneration are still largely unknown, due to technical limitations for the study of the large and insoluble ECM proteins and glycoproteins. Nevertheless, attempts have been made to manipulate skeletal muscle ECM to improve muscle repair, notably through the use of ECM scaffolds to restore volumetric muscle loss. 5
Matrix-based therapy aims at improving ECM remodeling to support tissue repair. In this context, components such as RGTA® for ReGeneraTing Agents include polymeric molecules that bind growth factors and cytokines and modulate their activity. Glycosaminoglycan mimetics mimic their capacities to sequester and present soluble factors to their receptors.6,7 RGTA are resistant to enzymatic degradation, and replace heparan sulfates (HS) at heparan-binding sites in the matrix, allowing the ECM scaffold and growth factors to reposition in the cellular microenvironment. 7 Among the latest developed RGTA are dextran polymers, harboring controlled sulfated or controlled sulfated carboxymethyl and acetylated groups and acting as an HS mimetic7,8 that has proven to improve repair of various tissues, including skin, 9 bone, 10 tendon, 11 or periodontal tissue, 12 with no toxicity. 13
First developed RGTA were tested in mechanical models of skeletal muscle injury, in healthy rats, where they were injected at the time of the damage. Results have shown an acceleration of the regeneration process assessed by the kinetics of myosin heavy chain expression. 14 Injection of RGTA was also tested in the model of hindlimb critical ischemia that induces muscle injury, where it was also injected at the time of the damage. Results showed a protective effect of RGTA on myofibers, and an improvement of the muscle repair process, mainly due to an increase of surviving myofibers. 15
A clinical grade RGTA was more recently used in the same context of hindlimb ischemia. 16 It was shown that RGTA increases the number of myonuclei, indicative of beneficial effect on myogenesis, as well as an increase of vessel density, indicative of a beneficial effect on angiogenesis. 16 Here again, the administration was concomitant to the injury, preventing to discriminate the effects of RGTA on cell survival versus on tissue repair. Moreover, administration of a therapeutic agent at the time of injury is not translatable to the clinical practice.
To dissect RGTA effects on skeletal muscle regeneration, we used a defined RGTA (namely OTR4133 with carboxymethyl sulfate and acetate groups) in a model of toxic injury that targets the whole muscle (on the contrary of mechanical injuries that differentially affects the various parts of the muscle), and which is highly reproducible and well established in terms of kinetics of the biological processes involved (i.e., myogenesis, angiogenesis, inflammation, ECM remodeling). The first phase is the inflammatory phase during which inflammatory cells invade the damaged muscle, clean the tissue debris, and stimulate MuSC expansion.1–3 At the time of the resolution of inflammation (days 2–3), the recovery phase begins, MuSCs exit the cell cycle, proliferate (days 3–4 postinjury), and commit into terminal myogenic differentiation and fuse to form new myofibers (days 6–14). 17 During the recovery phase, the formation of new vessels is observed, since angiogenesis is coupled with myogenesis. 18
In the present study, we identified the effects of a single injection of RGTA at the time of recovery during skeletal muscle regeneration and evaluated the outcomes on regeneration efficiency. Data showed that during the recovery phase, RGTA administration stimulated MuSC fusion and angiogenesis, accelerating muscle regeneration.
Materials and Methods
Muscle injury
Adult 9-week-old C57BL/6J male mice were used according to the French legislation. Injury was induced by injecting 50 μL of cardiotoxin (Latoxan, 12 μM) in each tibialis anterior muscle as previously described. 17 RGTA (GAG mimetic ([OTR-4133], a close molecule to OTR4131, 11 provided by OTR3 Company, Paris, France), was injected into a regenerating muscle 3 days after cardiotoxin injury at 1 μg/10 μL/muscle (control muscle was injected with 10 μL of phosphate-buffered saline [PBS]/muscle). Mice were euthanized 8 and 28 days after injury and muscles were harvested, frozen in liquid nitrogen-precooled isopentane, and stored at −80°C. Ten micrometer-thick cryosections were prepared for histological analysis.
Histological staining
Muscle sections were stained with Hematoxylin/Eosin to evaluate the overall regeneration process. Sudan Black staining was made to stain lipids: sections were dehydrated in ethanol 70% for 10 s, stained in Sudan Black solution (Sigma-Aldrich) for 2 h, counterstained with Hemalun (Sigma-Aldrich) for 1 min, and mounted with Fluoromount-G Mounting Medium (Interchim). The entire muscle section was scanned with an Axio Scan Z1 microscope (Zeiss) at 20 × objective connected to a three CCD HV-F 2025 color camera (Hitachi). The stained area was quantified using ImageJ software and was represented as a percentage of the entire muscle section area.
Histological immunolabeling
Muscle cryosections were treated with primary antibodies as in Latroche et al. 18 for the detection of laminin (No. L9393; Sigma-Aldrich) and of CD31 (Abcam, ab28364) revealed with Cy3- or FITC-conjugated antibodies (Jackson ImmunoResearch, Inc.). Nuclei were labeled with Hoechst (Sigma-Aldrich). For laminin immunolabeling, the entire cryosections were automatically scanned at ×10 objective using an Axio Observer.Z1 (Zeiss) connected to a CoolSNAP HQ2 CCD Camera (Photometrics). The image of the whole cryosection was automatically reconstituted in MetaMorph Software. Cross-sectional area (CSA) was measured using a macro developed under ImageJ software. 19 For CD31 evaluation, about 10–12 pictures were recorded at 20 × objective. The number of vessels (CD31-positive structures exhibiting a nucleus) and the number of myofibers were counted.
MuSC culture
MuSCs were extracted from mouse muscle hindlimbs from 3 to 5 weeks of age as described previously. 17 Cells were cultured on 2-well Permanox Chambers (Nunc Lab-Tek) coated with Matrigel Matrix Growth Factor Reduced (Corning, 356231) (1/10). For proliferation assay, MuSCs were seeded at 10,000 cell/cm2 in proliferation medium (DMEM/F12 [Gibco], 20% fetal bovine serum [Gibco], and 2% Ultroser™ G [Pall, Inc.]) for 6 h, washed three times with PBS, and were incubated in proliferation medium containing RGTA (10 μg/mL) or not for 1 day at 37°C. Immunolabeling for Ki67 (Abcam, ab15580) was performed as described in Theret et al. 17 For fusion assay, MuSCs were seeded at 30,000 cell/cm2 in proliferating medium for 6 h; then they were switched to differentiation medium (DMEM/F12, 2% horse serum [Gibco]) containing RGTA (10 μg/mL) or not for 3 days at 37°C. Phalloidin staining (Sigma-Aldrich, P5282) was used to label actin and Hoechst to stain nuclei. Pictures were recorded using a Zeiss Observer Z1 microscope connected to a Coolsnap HQ (Photometrics) camera at 20 × magnification. About six to eight randomly chosen fields were counted. The number of ki67-positive cells was expressed as a % of total nuclei. The number of nuclei per cell was counted, defining several classes of myotubes. Fusion index was calculated as the number of nuclei within myotubes upon the total number of nuclei.
3D angiogenesis culture assay
The 3D culture was carried out as previously described. 18 Briefly, human umbilical vein endothelial cells were infected with lenti-green fluorescent protein (GFP), were seeded on cytodex beads, and embedded into a fibrinogen solution. The cells/fibrinogen solution was allowed to clot and form a gel in eight-well glass Lab-Tek. When set, EndoGro medium containing RGTA (10 μg/mL) or not was added on top of the gel and was changed every 2 days. After 6 days, cultures were captured using a Zeiss Observer Z1 microscope connected to a Coolsnap HQ (Photometrics) camera at 10× objective. Analysis was done with ImageJ software as described in Latroche et al. 18
Statistical analyses
Results are expressed as mean ± standard error of the mean. Statistical analyses, such as t-tests and analysis of variance (ANOVA), are as described in the figure legends. The number of experiments is indicated in the figure legends.
Results and Discussion
RGTA was in situ administrated 3 days postinjury. In this way, the effects of RGTA were analyzed on the recovery phase of muscle regeneration that takes place after the inflammatory phase is resolved. 20 Days 8 and 28 postinjury were chosen to evaluate the direct impact and the long-lasting effects of RGTA on muscle regeneration, respectively. RGTA shows a dose-dependent effect on tissue repair with a bell-shaped curve. 7 Pharmacokinetic studies showed a durable accumulation (8 days) of RGTA at sites of injury/regeneration when administrated once at the time of the injury, either locally or systemically, 21 suggesting that a single injection could be optimal. The dose chosen was adapted from previous studies and results from OTR3. 16
RGTA is beneficial to skeletal muscle regeneration by increasing muscle cell fusion and angiogenesis
Cardiotoxin-injected tibialis anterior muscles presented a homogenous regeneration pattern indicating that the puncture made at day 3 postinjury (of either saline or RGTA) did not induce a supplementary locally lasting injury (Fig. 1A). At day 8 after injury, there was no difference in weight between the RGTA-treated and control muscles (Fig. 2A). Laminin immunostaining, to label the basal membrane surrounding each myofiber, was made to analyze the number and size of myofibers (Fig. 1B). Neither the total number of myofibers (Fig. 2A), nor the size of the myofibers (Fig. 2C, D) was changed. However, our analysis showed that RGTA treatment induced an important increase of the number of myonuclei per myofiber (+145%) (Fig. 2E) that leaded to the increased number of myofibers presenting four or more nuclei on the muscle sections (Fig. 2F). This result suggests that RGTA treatment increased myogenic cell fusion at the time of the formation of new myofibers during muscle regeneration.
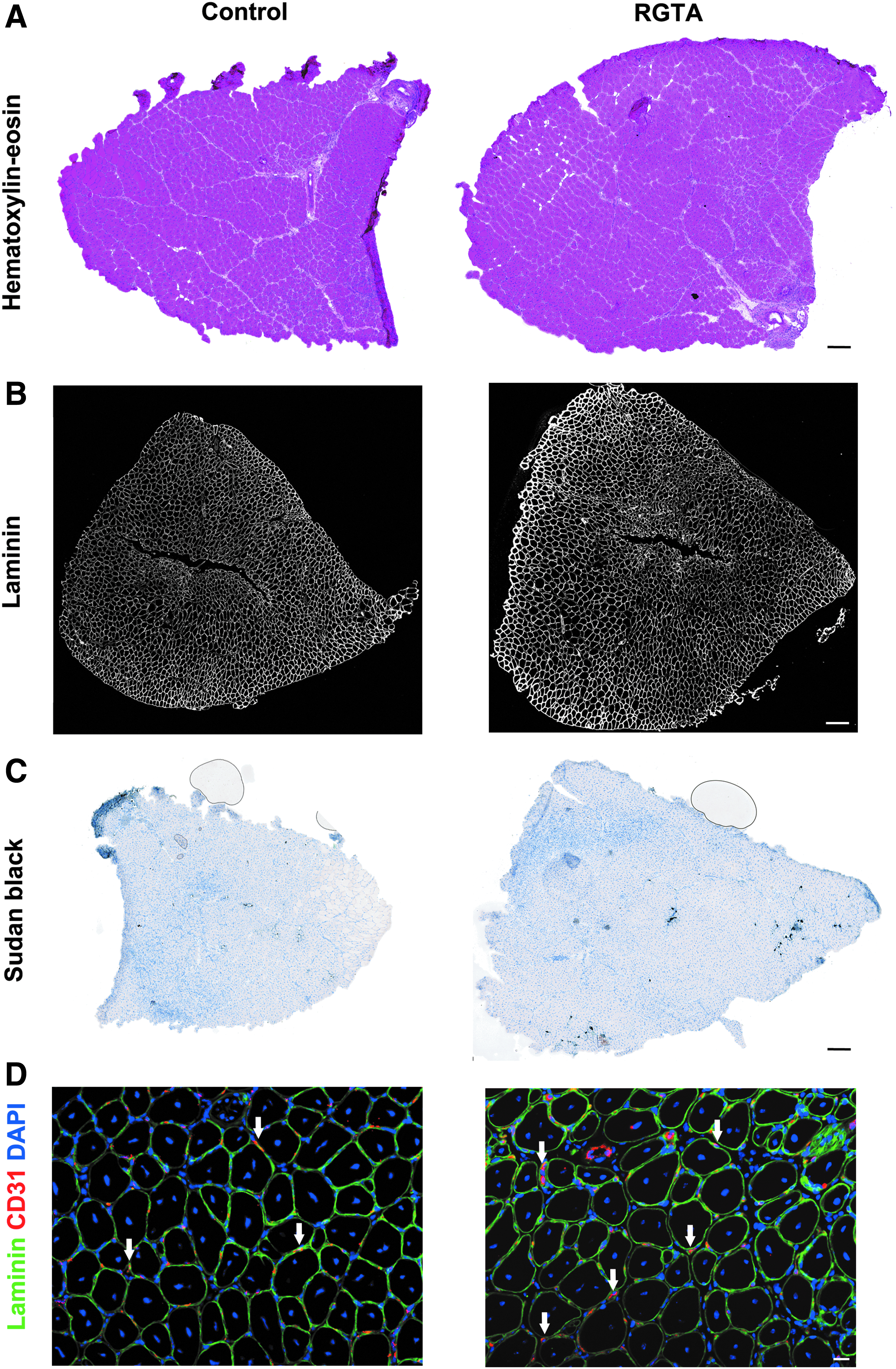
Analysis of the effects of RGTA® on skeletal muscle regeneration 8 days postinjury. RGTA (1 μg/muscle) was i.m. injected 3 days after cardiotoxin injury and muscles were recovered 5 days later.
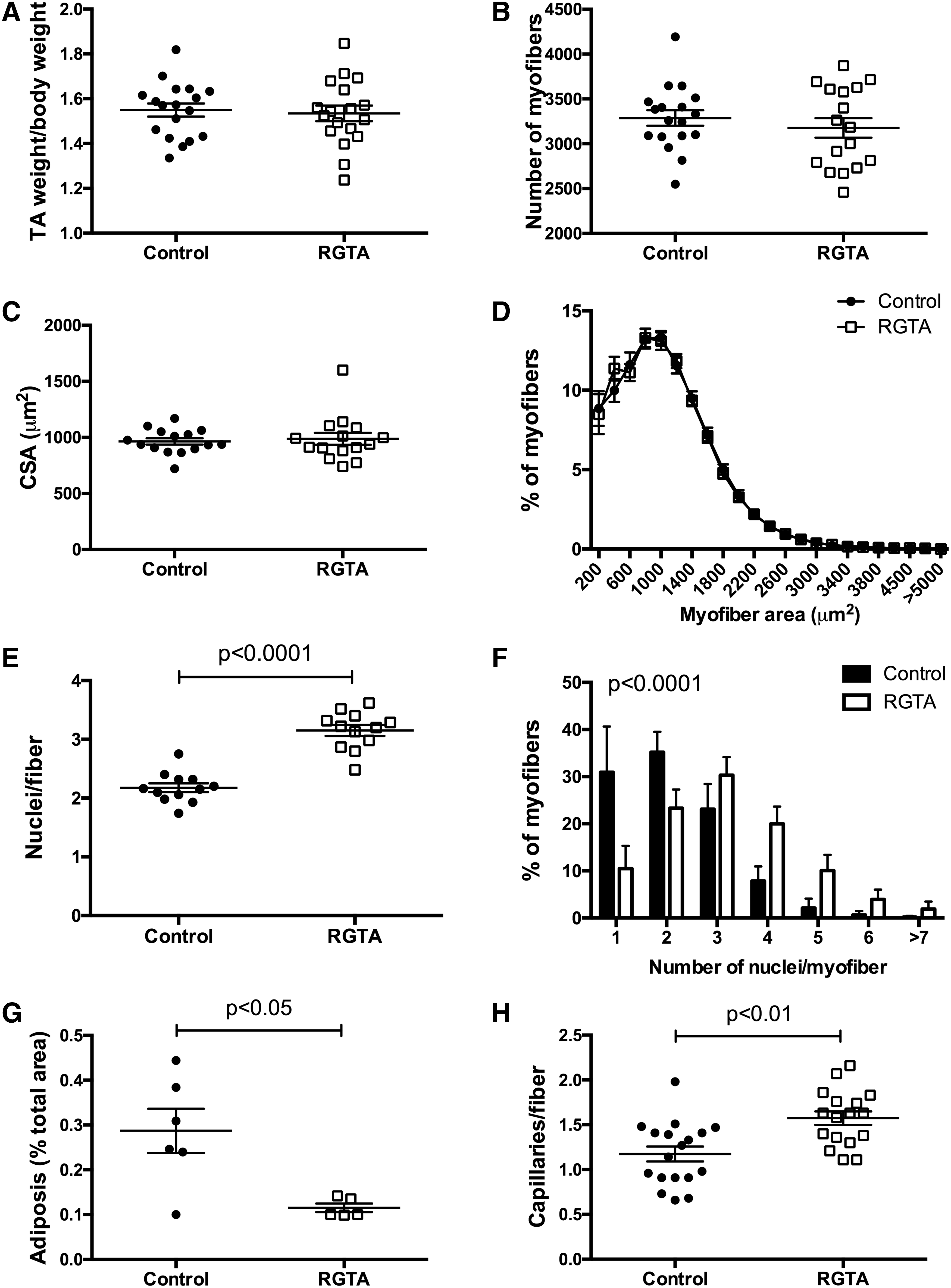
Effect of RGTA on skeletal muscle regeneration 8 days postinjury. RGTA (1 μg/muscle) was i.m. injected 3 days after cardiotoxin injury and muscles were recovered 5 days later.
Sudan Black staining indicated that the area covered by lipids decreased by 60% in RGTA-treated muscles (Figs. 1C and 2G). Lipid accumulation in regenerating muscle is a hallmark of defective regeneration, adipocytes deriving from fibroadipogenic precursors (FAPs). 22 However, the level of lipid staining was very low even in the control muscles (0.3% of the total muscle area) suggesting no major change in FAP homeostasis. Finally, immunostaining for CD31 (expressed by endothelial cells) was performed to analyze the capillarization of myofibers. The number of capillaries/myofiber was increased by 134% in RGTA-treated muscles as compared with the control muscles (Figs. 1D and 2H), indicating that RGTA increased angiogenesis during muscle regeneration.
Similar analyses were performed on day 28 after the injury to evaluate the long-lasting effects of RGTA treatment (Fig. 3). Muscle weight (Fig. 4A), myofiber number (Fig. 4B), and size of the myofibers (Fig. 4C, D) were not changed by RGTA treatment. The mean number of nuclei in myofibers was normalized between the two conditions (Fig. 4E), while the distribution of the myofibers according to their number of myonuclei was still statistically different, indicating that myofibers with high numbers of nuclei were more numerous in RGTA-treated muscles than in control muscles (Fig. 4F). Adiposis staining was very low at day 28 postinjury with no difference between control and RGTA-treated groups (Figs. 3C and 4G). On the contrary, capillarization of the myofibers was still 123% higher in RGTA-treated muscles than in control muscles (Figs. 3D and 4H).

Analysis of the effects of RGTA on skeletal muscle regeneration 28 days postinjury. RGTA (1 μg/muscle) was i.m. injected 3 days after cardiotoxin injury and muscles were recovered 25 days later.

Effect of RGTA on skeletal muscle regeneration 28 days postinjury. RGTA (1 μg/muscle) was i.m. injected 3 days after cardiotoxin injury and muscles were recovered 25 days later.
Altogether, these results indicate that RGTA administration at the time of the resolution of inflammation supported the recovery processes and accelerated muscle regeneration. A previous study reported an increased size of generating myofibers upon RGTA treatment, the mimetic was administrated at the time of the injury (which was ischemia/denervation), and the treatment was associated with a decrease of cell damage. 15 In a similar critical hindlimb ischemia, where the first damage is vascular, RGTA administration at the time of injury increased the density of myonuclei and that of vessels 8 days after ischemia. 16 Our in vivo results indicate a long-lasting effect of RGTA on muscle cell fusion and on angiogenesis during muscle regeneration.
RGTA directly acts on MuSCs and endothelial cells to stimulate myogenesis and angiogenesis in vitro
Upon activation after muscle injury (i.e., plating in culture), MuSCs first proliferate to expand, then exit the cell cycle to commit into terminal differentiation to fuse together to form multinucleated myotubes, which prefigure the myofibers in vitro. MuSC proliferation and differentiation/fusion are regulated by opposite molecular mechanisms. 1 To confirm the above in vivo observations at the cell level in vitro, culture experiments were performed in the presence of RGTA. In vitro and in vivo comparisons are difficult to make since the accessibility of the cells to the compound in vivo is difficult to assess. We therefore tested a range of RGTA concentrations to investigate its effect on MuSC fusion, since the major outcome observed in vivo was an increase of the number of myonuclei per myofiber. Myotubes were identified as structures containing several nuclei after staining for the ubiquitous cytoskeletal component actin (Fig. 5A, B).

Effect of RGTA on in vitro myogenesis and angiogenesis.
Our results show that a concentration of 10 μg/mL of RGTA produced the most consistent effect on several primary cells (data not shown) and was chosen. In accordance with the increased number of myonuclei in regenerating myofibers in RGTA-treated muscles in vivo, RGTA stimulated MuSC fusion (+145%), dramatically increasing the appearance of myotubes exhibiting high number of nuclei (Fig. 5A, B). Consistently, RGTA induced a decrease of the proliferation of MuSCs (−19%) (Fig. 5C, D) that was evaluated after immunostaining for ki67 that labels cycling cells. These results show that RGTA directly promotes MuSC fusion, in accordance with the in vivo results. Previous studies, using either cell lines or rat primary cells, showed that the effects of RGTA are dose dependent, and for the efficient concentrations, promotes both cell growth and fusion that were analyzed in the same experiment.23–25 RGTA proliferative effect was mainly driven through its potentiating effect of fibroblast growth factor basic (FGFb) on MuSCs. 24
Similarly, RGTA was shown to increase angiogenesis through the potentiation of vascular endothelial growth factor (VEGF) properties. 26 We took advantage of a 3D angiogenesis model in which GFP-labeled human endothelial cells are allowed to form new vessels that further lumenize, indicative of their differentiation and the maturation of the newly formed capillaries. 18 Using this device, we showed that RGTA increased (+168%) the length of capillaries that formed (Fig. 4E, F). Moreover, RGTA triggered the maturation of the sprouting vessels, as assessed by a huge increase of their lumenization (+248%) (Fig. 5G). It is to note that in the RGTA condition, more nuclei are visible within the gel (Fig. 5F), indicative of a stimulating effect of RGTA on endothelial cell proliferation, which was previously reported.26,27 These results show the direct proangiogenic properties of RGTA on endothelial cells, in accordance with the increased capillarization observed in the regenerating muscle in vivo upon RGTA treatment.
In conclusion, our study shows that the clinical grade RGTA is effective at accelerating muscle regeneration when locally administered at the onset of the regenerating phase, a clinically compatible time point. We showed in vitro that the main properties of RGTA are a stimulating effect on MuSC fusion that is required for the formation of new myofibers and on the formation and maturation of new blood vessels. These direct and specific effects of RGTA on MuSCs and endothelial cells accelerate myogenesis and angiogenesis during muscle regeneration.
As a natural HS, RGTA hold a strong binding capacity for soluble factors providing their stability and their protection against enzymatic degradation. 8 Growth factors known to interact with HS include molecules such as FGF, VEGF or transforming growth factor beta (TGFβ), which have been involved in both myogenesis and angiogenesis. 1 Thus RGTA compounds may be of interest for accelerating muscle regeneration or healing in specific conditions, for example in athletes, for which this process needs to be shortened.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by ANR-15-ASTR-0013-01 Myo-MECa, supported by Agence National de la Recherche, and Direction Générale des Armées.