
Abstract
Implantation of chondrogenically differentiated mesenchymal stromal cells (MSCs) leads to bone formation in vivo through the process of endochondral ossification. The use of allogeneic MSCs for this purpose may be a promising new approach to replace the current gold standard of bone regeneration. However, the success of using allogeneic cells depends on the interaction between the implanted cells and the host's endogenous immune cells. Th17 T cells and other CD4 helper T cell subtypes have been shown to negatively impact chondrogenesis, however, it is unclear how the interaction between these cells affects bone regeneration mediated by these cells. The aim of the current work was to assess the effect of chondrogenic MSC pellets on Th1, Th2, Th17, and regulatory T cells in vitro. Human MSCs were nonchondrogenic (−TGFβ3) and chondrogenically (+TGFβ3) differentiated for 7 or 21 days. Memory T cells (sorted from the CD4 population of peripheral blood mononuclear cells [PBMCs]), as well as total PBMCs were cocultured with allogeneic nonchondrogenic and chondrogenic MSC pellets for 3 days. Seven-day differentiated allogeneic nonchondrogenic and chondrogenic MSC pellets that were cocultured with memory T cells resulted in a significant increase in Th2 and a decrease in Th1 T cells. Furthermore, the co-culture of 21-day differentiated nonchondrogenic and chondrogenic MSC pellets with memory T cells resulted in a significant increase in Th2 and Th17 T cells, as well as a decrease in Th1 and regulatory T cells. Interleukin (IL)-6 was identified as a predominant cytokine involved in this interaction between allogeneic chondrogenically differentiated MSC pellets and memory CD4 T cells, with high levels of IL-6 being secreted in the supernatants of this cocultured condition. The findings of this study highlight the potential of chondrogenically differentiated MSC pellets to alter the ratio of Th1 and Th2 as well as Th17 and regulatory T cell subsets. Additional analysis investigating bone formation by chondrogenically differentiated MSCs in an allogeneic setting may identify a novel role of these T cell subsets in bone regeneration processes mediated by chondrogenically differentiated MSCs.
Impact statement
Allogeneic mesenchymal stromal cells (MSCs) have the potential to be an off-the-shelf treatment for bone repair. However, the lack of knowledge of the immune cells involved in this process has hampered the progression to the clinic. The current study has shown that allogeneic chondrogenic MSCs have the potential to skew the ratio of specific helper CD4 T cell subsets in vitro. This has now provided insight for future in vivo experiments to investigate the role of these T cell subsets in the early stages of bone regeneration mediated by allogeneic chondrogenic MSCs.
Introduction
Implantation of chondrogenically differentiated mesenchymal stromal cells (MSCs) leads to bone formation in vivo through the process of endochondral ossification.1–3 Before implantation, MSCs are primed with transforming growth factor beta (TGFβ) to induce chondrogenesis. Without TGFβ, these nonchondrogenic MSCs do not form bone in vivo. 4 The potential of using allogeneic MSCs for this purpose would lead to the exciting opportunity to develop an off-the-shelf treatment for bone repair. Understanding the immune cells potentially interfering with the ability of allogeneic chondrogenic MSCs to regenerate bone will determine the progression of this potential treatment to the clinic.
Previously, we have demonstrated that allogeneic nonchondrogenic and chondrogenic MSCs do not affect the proliferation of CD4 or CD8 T cells. 5 The findings of this study highlight that chondrogenically differentiated MSCs are nonimmunogenic in terms of T cell proliferation and are therefore not expected to induce detrimental immune responses in vivo. However, the specific T cell subsets involved in the process of bone regeneration mediated by chondrogenic MSCs still needs to be identified. Additionally, further understanding of the immune responses toward chondrogenic bone-forming MSCs and nonchondrogenic nonbone-forming MSCs and how these responses may differ, may identify immune cell subsets involved in bone forming processes mediated by chondrogenic MSCs.
CD4 T lymphocytes and their cytokines have been previously shown to affect bone regeneration mediated by undifferentiated allogeneic MSCs.6,7 Activation of CD4 T cells by antigen-presenting cells, such as dendritic cells, induces their differentiation from naive T cells into their effector subsets (T-helper [Th] 1, Th2, Th17, and regulatory T cells). Specific chemokine receptor markers allows the identification of the different subsets and are as follows: Th1 (CCR6−CCR4−CXCR3+), Th2 (CCR6−CCR4+CXCR3−), Th17 (CCR6+CCR4+CXCR3−), and regulatory T cells (CD4+CD25highCD127lowFoxP3+). 8 Furthermore, the CD4 T cell subsets can be further discriminated by their cytokine expression. Th1 cells are known to predominately secrete interferon gamma (IFNγ), whereas Th2 cells secrete interleukin (IL)-4, IL-5, and IL-13.9–12 IL-17A, IL-17F, and IL-22 are the signature cytokines produced by Th17 cells,13–15 whereas IL-10 has been shown to be secreted by regulatory T cells in order for them to induce their anti-inflammatory role in the immune system.16,17 Th17 cells and their related cytokines have been shown to play a role in disrupting joint homeostasis in inflammatory diseases.18,19 The combined expression of TGFβ and IL-6 results in the differentiation of Th17 cells. 20 Particularly in rheumatoid arthritis (RA), IL-17 secretion by these Th17 cells has been shown to negatively impact matrix production by chondrocytes and osteoblasts in the joint. 21 Furthermore, IL-17 has been shown to induce receptor activator of nuclear factor kappa-B ligand expression by osteoblasts leading to osteoclastogenesis and bone destruction.22–24 While Th17 cells and their associated cytokines have been shown to induce bone destruction in RA, in other inflammatory diseases such as ankylosing spondylitis, IL-17 may be involved in the induction of ectopic bone formation. 25 Therefore, it is possible that these cells and associated cytokines could be altered by chondrogenic MSCs to generate bone in vivo.
Few studies to date have investigated Th17 cells and MSC-mediated bone regeneration. In a study by Liu et al., proinflammatory T cells (including Th17 cells) inhibited bone regeneration mediated by undifferentiated MSCs, while regulatory T cells were shown to inhibit these T cell responses to allow MSC-mediated bone regeneration to occur. 6 In contrast, IL-17 has been shown to enhance osteogenic differentiation of bone marrow-derived MSCs in vitro.26–28 In vivo studies investigating the effect of IL-17 on bone repair have been somewhat contradictory, however it is evident that the effect of IL-17 depends on the origin of the cells for the bone repair.29,30 Th17 cells and other CD4 T cell subsets have been shown to negatively impact MSC-mediated chondrogenesis31–34 ; however, it is unclear how interactions between these cells may affect bone regeneration mediated by MSCs. Therefore, the main aim of this study was to assess the effect of chondrogenically differentiated MSCs on the differentiation of naive CD4+ T cells and the ratio of different T helper memory subsets in vitro. Furthermore, the effect of allogeneic nonchondrogenic (nonbone-forming) and chondrogenic (bone-forming) MSCs on these CD4+ T cell subsets was compared. The results from this in vitro study could give indications whether and which of the Th subsets are potentially involved in the early stages of bone regeneration mediated by allogeneic chondrogenic MSCs.
Materials and Methods
Isolation and expansion of human bone marrow MSCs
Human adult bone marrow MSCs (A-MSCs) were obtained from heparinized femoral-shaft marrow aspirates of patients (32–78 years) undergoing total hip arthroplasty (with informed consent after approval by Erasmus MC Medical Ethics Committee protocol METC-2004-142). Using a sterile Jamshidi needle, 5–10 mL of bone marrow was harvested into sterile 10-mL syringes containing 0.5 mL of heparin (1000 U/mL). Heparinized aspirates were seeded at a density of 30–100 × 106 mononuclear cells per T175 flask in 25 mL of standard MSC medium consisting of minimum essential medium alpha (α-MEM) (Invitrogen, Carlsbad, CA) supplemented with 10% heat-inactivated fetal calf serum (FCS; Gibco, The Netherlands, selected batch), 50 μg/mL gentamicin, 1.5 μg/mL fungizone (All Invitrogen), 1 ng/mL fibroblast growth factor-2 (Instruchemie B.V., Delfzijl, The Netherlands) and 0.1 mM of
Chondrogenic differentiation of MSCs
MSCs were detached using 0.05% trypsin and added to 15 mL polypropylene tubes (Sarstedt) at a density of 0.2 × 106 cells in 0.5 mL of standard chondrogenic medium consisting of high-glucose Dulbecco's modified Eagle's medium supplemented with 50 mg/mL gentamicin, 1.5 mg/mL fungizone, 100 mM sodium pyruvate (Sigma-Aldrich), 1:100 ITS (BD Biosciences),
Histological analysis of human MSC pellets before coculture
Nonchondrogenic and chondrogenically differentiated MSC pellets were harvested before coculture to assess the level of chondrogenesis. The pellets were washed in PBS and then fixed in 4% formalin for 1 h at room temperature. Following fixation, pellets were embedded in 3% agarose, processed, and embedded in paraffin. Sectioned slides (5 μm sections) were deparaffinized through alcohol series (xylene, 100% ethanol, 96% ethanol, 70% ethanol) and rinsed twice in distilled water. Sections were stained for 5 min with 0.04% thionine in 0.01 M aqueous sodium acetate (pH 4.5) followed by differential staining in 70% ethanol (±10 s), 96% ethanol (±30 s), and 100% ethanol (1 min).
Isolation of peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats of healthy male blood donors (Sanquin Bloedvoorziening, Rotterdam, The Netherlands). Peripheral blood was transferred into 50-mL falcon tubes and gently inverted to mix. Samples were centrifuged at 388 g for 7 min to remove the top layer of plasma. Wash medium (RPMI-1640 medium containing 1.5 μg/mL fungizone, 50 μg/mL gentamicin) was added to the remaining peripheral blood in a 1:1 dilution and samples were carefully divided into 50-mL filter falcon tubes containing Ficoll-Paque PLUS (density 1.077 g/mL; GE Healthcare, Uppsala, Sweden) and centrifuged without the brake at 690 g for 20 min. Plasma was carefully removed and discarded, and the remaining layers above the filter were transferred to a new 50-mL falcon tube and washed with medium. Samples were then centrifuged at 835 g for 7 min using the brake. The supernatant was carefully removed, and the pellet was resuspended in wash medium followed by centrifugation at 690 g for 5 min. Following three washes, cells were resuspended in RPMI-1640 medium consisting of 1% GlutaMAX (Life Technologies), 1.5 μg/mL fungizone, 50 μg/mL gentamicin, and 10% FCS (batch tested for optimal viability and survival of PBMCs) and a cell count was performed. Cells were stored in liquid nitrogen until needed for human MSC (hMSC) pellet cultures in complete RPMI-1640 medium and 10% dimethylsulfoxide.
Naive and memory T cell sort and coculture
PBMCs were thawed from liquid nitrogen and CD4 T cells were separated using the MACS CD4+ T cell Isolation Kit (Miltenyi Biotech) according to the manufacturer's instructions. Briefly, 500 × 106 PBMCs were incubated with 500 μL CD4+ MicroBeads (Miltenyi, Biotech) for 20 min in the dark at 4°C, followed by washing the cells and resuspending them in running buffer (PBS with 0.5% bovine serum albumin [Sigma] and 2 mM EDTA [Gibco]). Subsequently, magnetically labeled cells were applied on a column (LS column) and placed in a MidiMACS Separator (All Miltenyi Biotech). The CD4+ fraction was collected following removal of the column from the MidiMACS Separator. Cells were sorted based on anti-CD4 FITC (BioLegend), anti-CD45RO PerCP.Cy5 (BD Biosciences), anti-CD25 PECy7 (BioLegend) and DAPI (Life Technologies) on the BD FACS ARIA II. Unsorted PBMCs, Naive T cells, and memory T cells (all stimulated with anticluster of differentiation 3 [CD3] and anti-CD28 [1 mg/mL, 1 μL/106 cells] and a goat linker antibody [0.5 mg/mL, 2 μL/106 cells] all from BD Biosciences) were cocultured with nonchondrogenic and chondrogenically differentiated MSCs in 1.5-mL polypropylene tubes (Sarstedt) for 3 days in supplemented RPMI-1640 medium.
Characterizing lymphocytes from the coculture using flow cytometry
Lymphocytes in suspension from cocultures with nonchondrogenic and chondrogenically differentiated MSC pellets were harvested and transferred to fluorescence-activated cell sorting (FACS) tubes. Lymphocytes were washed with FACS buffer and centrifuged at 689 g for 5 min. The cells were resuspended in 25 μL of FACS buffer containing anti-CD4 FITC, anti-CD45 Pe-CF594 (BD Biosciences), anti-CD45RO PerCPCy5.5, CCR6 PE (BD Biosciences), CCR4 APC (BioLegend), CXCR3 BV421 (BioLegend), and incubated for 15 min at room temperature in the dark. Samples were washed once with FACS buffer (689 g for 5 min), followed by a wash with PBS (689 g for 5 min). Cells were incubated with a Fixable Viability Dye (eBioscience) for 30 min at 4°C. Samples were washed once with PBS followed by a 10-min incubation with 2% paraformaldehyde in PBS at room temperature in the dark. Finally, cells were washed twice with FACS buffer (689 g for 5 min) and analyzed on the BD LSRII flow cytometer. To identify regulatory T cell populations, cells were permeabilized and stained using an anti-human FoxP3 APC Staining Kit (eBioscience) and stained for CD3 (FITC), CD4 (PerCP), CD25 (PeCy7), and CD127 (PE; BD Biosciences). These samples were resuspended in 200 μL of FACSFlow and analyzed using flow cytometry.
Cytokine measurements in supernatants from coculture
Supernatants from cocultures were centrifuged at 690 g for 10 min and stored at −80°C before analysis. Levels of IL-17A, IL-17F, IL-4, IL-6, IL-10, and IFNγ were determined in the supernatants from cocultures using enzyme-linked immunosorbent assays (ELISA; All eBioscience). The measurements were performed and calculated according to the manufacturer's instructions.
Quantitative real-time reverse transcription/polymerase chain reaction
A portion of the lymphocytes (∼1 × 105 cells) from cocultures were removed, washed with PBS, and resuspended in TRIzol reagent (Thermo Scientific). Similarly, nonchondrogenic and chondrogenically differentiated hMSC pellets were removed from the coculture, washed with PBS, and crushed in TRIzol reagent. RNA was isolated from all samples using the RNeasy Mini Kit (Qiagen). cDNA was synthesized from the isolated RNA using the First-Strand cDNA Synthesis Kit (Thermo Scientific) and used for quantitative real-time polymerase chain reaction (qRT-PCR). Quantitative gene expression was determined using TaqMan Universal PCR Master Mix for specific genes within the pellets SRY (sex-determining region Y)-box 9 (SOX9), matrix metalloproteinase (MMP)-1, and MMP-13. TaqMan assays-on-demand were used to assess the quantitative gene expression of the immune-related genes CD3 (Hs00167894_m1) within both the lymphocytes and the MSC pellets from the coculture. For the genes IL-6 and IL-10, qPCR Mastermix Plus for SYBR Green I—dTTP (Eurogentec) was used. Gene expression levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and relative expression calculated using the 2-ΔCT method. All gene sequences can be found in Table 1.
Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction Gene Sequences
Statistical analysis
Statistical analysis was performed using GraphPad Prism v.5 using a Repeated measures ANOVA with Bonferroni posttest. Values are presented as mean ± standard deviation. n = 3 PBMC donors with one hMSC donor, where p < 0.05 was considered statistically significant.
Results
Both allogeneic nonchondrogenic and chondrogenic A-MSC pellets induced an increase in the percentage of Th2 cells in unsorted PBMCs
To assess the effect of nonchondrogenic and chondrogenic A-MSC pellets on the differentiation of CD4 T cell subsets, A-MSCs were differentiated with and without TGFβ3 for 7 days. Following 7 days of differentiation, both nonchondrogenic (−TGFβ3) and chondrogenically differentiated (+TGFβ3) A-MSCs were cocultured with unsorted PBMCs for 3 days. Thionine staining performed on A-MSC pellets confirmed that A-MSCs cultured in the presence of TGFβ3 were chondrogenic (Supplementary Fig. S1). Both allogeneic nonchondrogenic and chondrogenically differentiated A-MSCs induced an increase in the percentage of Th2 cells compared with PBMCs alone (21.7% ± 6.1% vs. 34.9% ± 4.4% and 34.2% ± 4.1%, respectively) (Fig. 1A (ii)). Although there was a trend toward a decrease in the percentage of Th1 cells, this was not statistically significant (16.7% ± 0.7% vs. 13.4% ± 1.8% and 13.7% ± 1.1%) (Fig. 1B (i)). No effect was found on Th17 cells cocultured with either allogeneic nonchondrogenic or chondrogenically differentiated A-MSC pellets (13.5% ± 2.1% vs. 11.2 ± 1% and 10.7% ± 3%) (Fig. 1B (iii)). CD8+ T cells were not affected in unsorted PBMCs by allogeneic nonchondrogenic or chondrogenic A-MSC pellets (Fig. 1B (iv)). Furthermore, there were no differences in the percentage of naive/memory CD4 T cells in total PBMCs with or without MSC pellet (data not shown). Importantly, there was no difference between nonchondrogenic or chondrogenically differentiated A-MSCs on the CD4+ Th subsets in unsorted PBMCs.
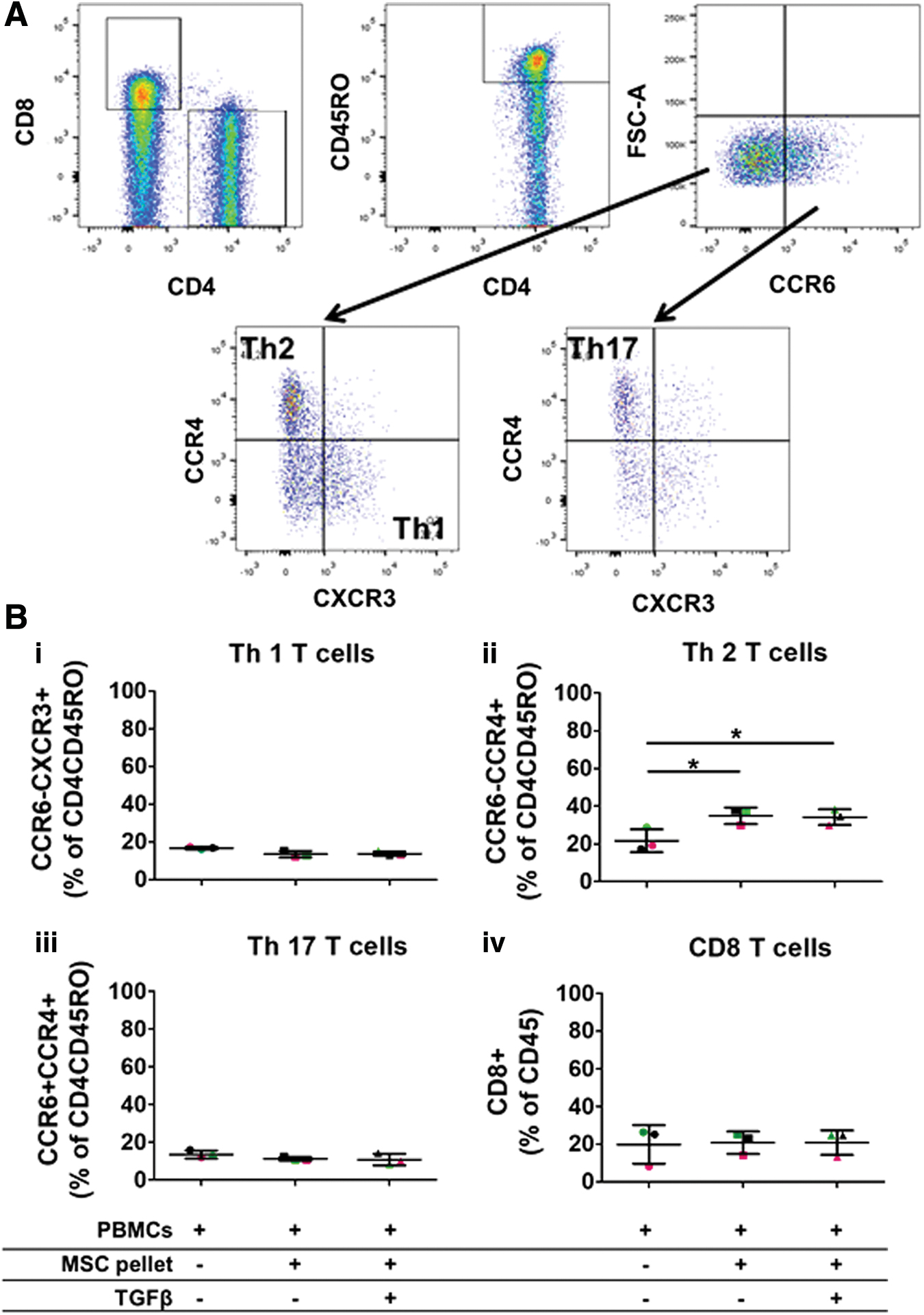
Nonchondrogenic and chondrogenically differentiated A-MSC pellets increase the percentage of Th2 cells in coculture with total PBMCs. Human A-MSCs were nonchondrogenic (−TGFβ3) and chondrogenically (+TGFβ3) differentiated for 7 days. Unsorted PBMCs were thawed and stimulated with CD3/CD28. CD3/CD28 PBMCs were cultured with 7-day nonchondrogenic and chondrogenically differentiated A-MSCs for 3 days. Following coculture, PBMCs were harvested and analyzed by flow cytometry for specific markers (CD45, CD8, CD4, CD45RO, CCR6, CCR4, and CXCR3). The gating strategy for flow cytometry analysis is depicted in
Allogeneic nonchondrogenic and chondrogenic A-MSC pellets do not affect naive CD4 T cell differentiation, but significantly change the ratio of Th1/Th2 cells in memory T cells
The effect of allogeneic chondrogenic A-MSC pellets on the differentiation of CD4 T cell subsets was further assessed using naive or memory CD4+ T cells. CD4+ T cells were sorted from PBMCs and further purified for naive (CD4+CD45RO−) or memory (CD4+CD45RO+) T cell subsets (Supplementary Fig. S2). Allogeneic nonchondrogenic and chondrogenic A-MSC pellets that were differentiated for 7 days were then cocultured with the sorted naive T cells. CD8 T cells were not detected in any of the sorted samples, demonstrating successful sorting of the CD4 T cell population (data not shown). Furthermore, memory T cells highly expressed CD45RO in comparison to naive T cells demonstrating successful sorting of both of these cell types (Supplementary Fig. S3). A trend toward a decrease in Th1 cell differentiation was found in naive T cells cultured with allogeneic nonchondrogenic and chondrogenic A-MSC pellets (17.1% ± 13.1% vs. 6% ± 3.7% and 7.7% ± 2.9%), however, this was not significant (Fig. 2A (i)). No effect on Th2 cell differentiation was found in naive T cells cocultured with allogeneic nonchondrogenic and chondrogenic A-MSC pellets (11.3% ± 2.2% vs. 17.1% ± 4.9% and 16.5% ± 5.4%) (Fig. 2A (ii)). Finally, also no significant effect was found on Th17 T cell differentiation in naive T cells cocultured with nonchondrogenic and chondrogenically differentiated A-MSCs (2.5% ± 1.1% vs. 4.6% ± 1.4% and 4.6% ± 0.4%) (Fig. 2A (iii)). Again, there was no difference between nonchondrogenic or chondrogenically differentiated A-MSCs on CD4+ T cell differentiation in naive T cells. In contrast to the effect on naive T cells, both allogeneic nonchondrogenic and chondrogenic A-MSC pellets significantly decreased the percentage of Th1 cells (16.6% ± 2.2% vs. 6.8% ± 1.7% and 9.2% ± 1.9%) and increased the percentage of Th2 cells (25.2% ± 4.6% vs. 41.6% ± 4.9% and 40.7% ± 4.2%) (Fig. 2B (i) and (ii)). Nonchondrogenic A-MSC pellets had a higher inhibitory effect on Th1 cells than chondrogenically differentiated A-MSC pellets. An increase in the percentage of Th17 cells was found in memory T cells cocultured with nonchondrogenic or chondrogenically differentiated A-MSC pellets, but this finding was not significant (6.7% ± 0.9% vs. 11% ± 3.7% and 9.8% ± 1.7%) (Fig. 2B (iii)).
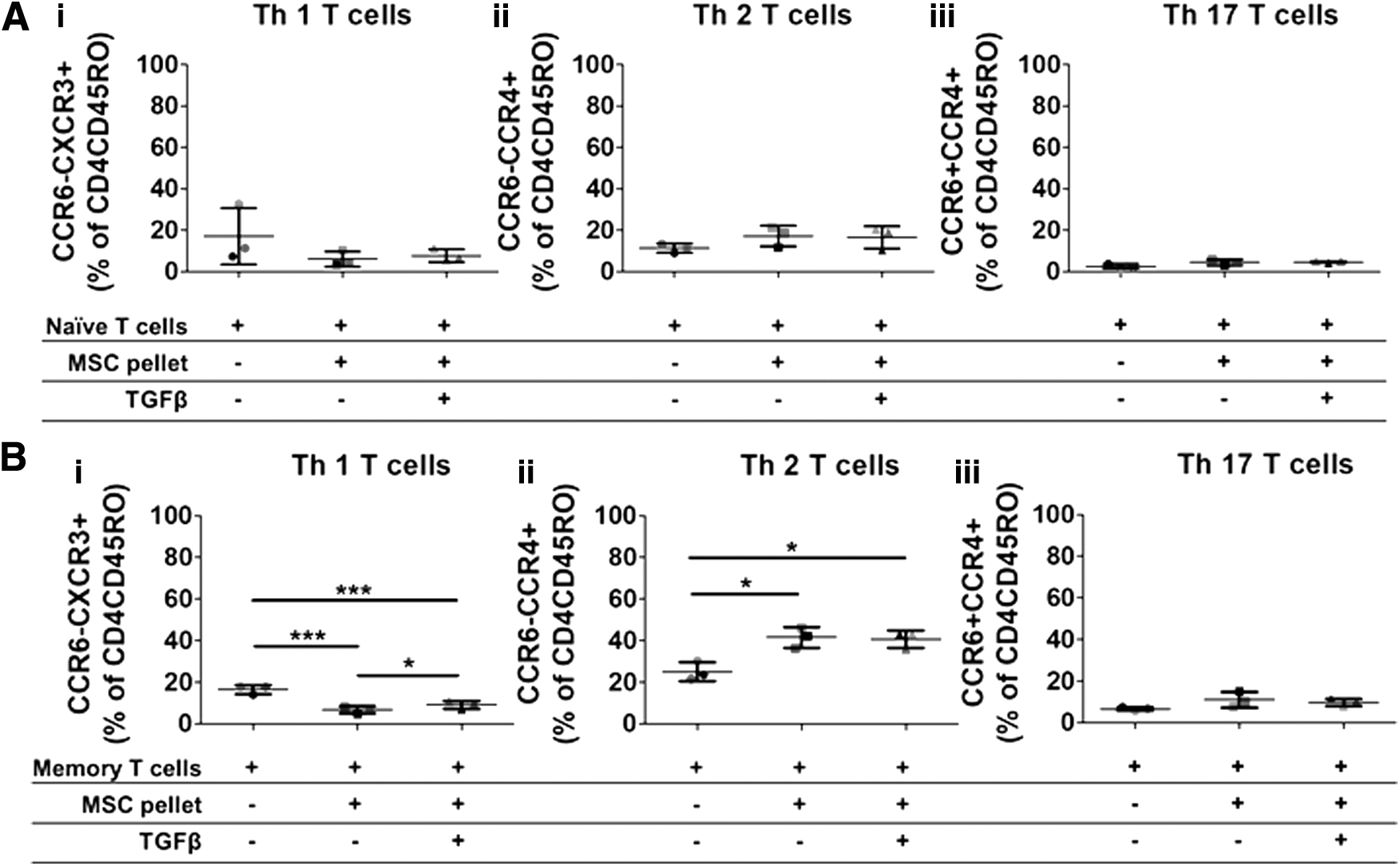
Allogeneic nonchondrogenic and chondrogenic A-MSC pellets do not affect naive CD4 T cell differentiation, but significantly change the percentage of Th1 and Th2 cells in memory T cells. Human A-MSCs were nonchondrogenic (−TGFβ3) and chondrogenically (+TGFβ3) differentiated for 7 days. Naive (CD4+CD45RO−) and memory (CD4+CD45RO+) T cell subsets were sorted from CD4 T cells and stimulated with CD3/CD28. CD3/CD28 naive or memory T cells were cultured with 7-day nonchondrogenic and chondrogenically differentiated A-MSCs for 3 days. Following coculture, naive and memory T cells were harvested and analyzed by flow cytometry for T cell-specific markers (CD45, CD8, CD4, CD45RO, CCR6, CCR4, and CXCR3). Both nonchondrogenic and chondrogenically differentiated A-MSCs had no effect on the differentiation of CD4 subsets in naïve T cells
Memory T cells infiltrate the matrix of both nonchondrogenic and chondrogenically differentiated A-MSC pellets and modulate A-MSC gene expression
To assess whether memory T cells had infiltrated the matrix of the differentiated A-MSC pellets, the A-MSC pellets were removed from the coculture, washed once with PBS, and then processed for RNA isolation. The RNA from both nonchondrogenic and chondrogenic A-MSC pellets cocultured with memory T cells was isolated and analyzed by qRT-PCR. CD3 gene expression was significantly upregulated in nonchondrogenic (40 vs. 27.6 ± 1.1 Ct value) and chondrogenically (40 vs. 27.5 ± 1.3 Ct value) differentiated A-MSC pellets (Fig. 3A), highlighting the presence of CD3-positive memory T cells in the matrix of nonchondrogenic and chondrogenic A-MSC pellets. Furthermore, the coculture of memory T cells significantly inhibited the expression of Sox 9 in chondrogenically differentiated (0.03 ± 0.01 vs. 0.01 ± 0.01) A-MSC pellets, but to a lesser extent in nonchondrogenic (0.02 ± 0.01 vs. 0.01 ± 0.003) A-MSC pellets (Fig. 3B). Gene expression levels of the matrix-degrading enzyme MMP-1 were upregulated in nonchondrogenic (0.001 ± 0.0001 vs. 0.01 ± 0.003) and chondrogenic (0.001 ± 0.0002 vs. 0.004 ± 0.001) A-MSC pellets cultured with memory T cells. MMP-3 was also upregulated in nonchondrogenic (0.002 ± 0.001 vs. 0.09 ± 0.04) and chondrogenic (0.005 ± 0.002 vs. 0.13 ± 0.05) cocultured A-MSC pellets.
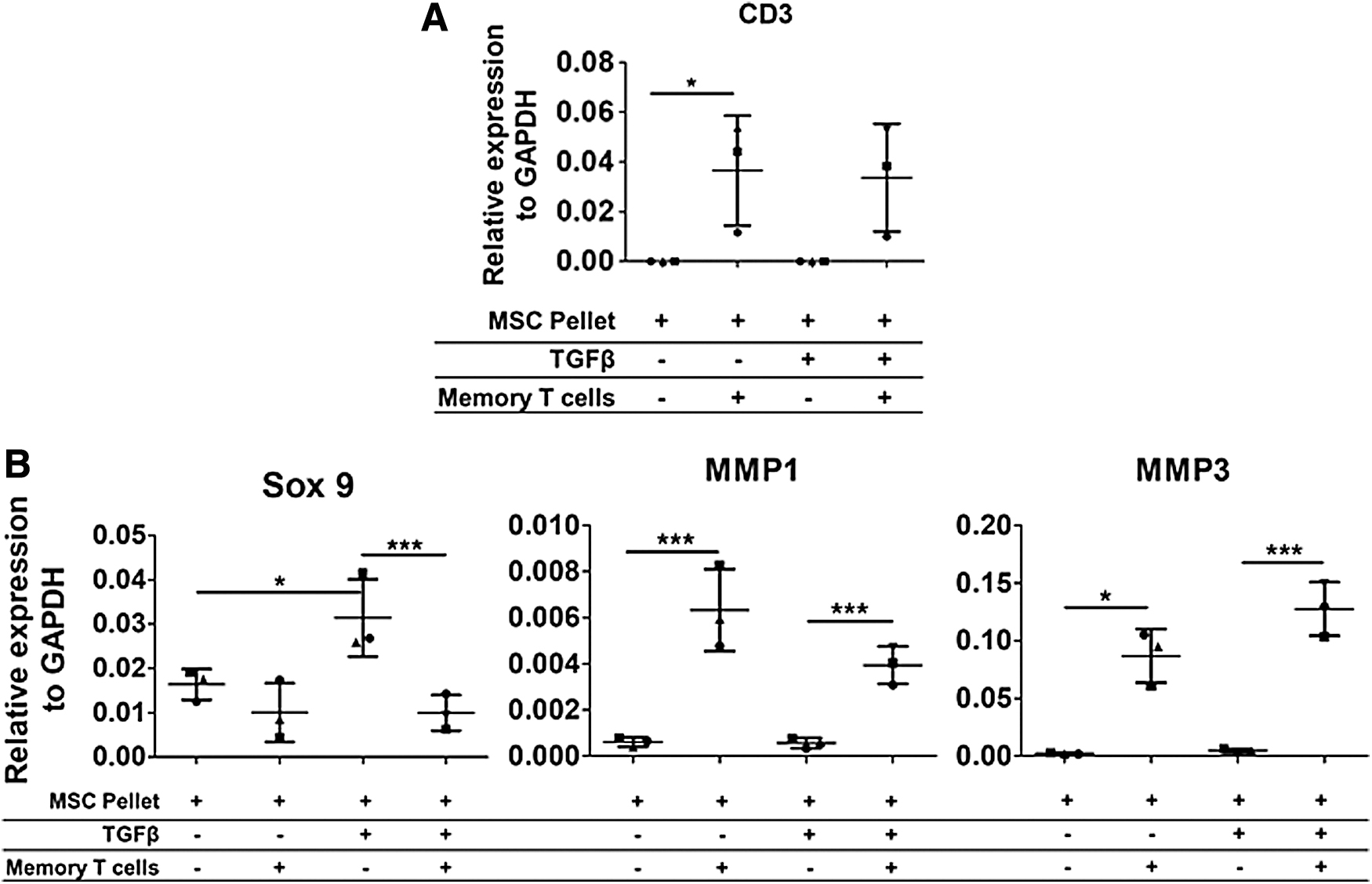
Memory T cells infiltrate the matrix of both nonchondrogenic and chondrogenically differentiated A-MSC pellets and modulate A-MSC gene expression. Memory T cells were cultured with 7-day nonchondrogenic and chondrogenically differentiated A-MSCs for 3 days. Following coculture, quantitative gene expression of CD3, Sox 9, MMP-1, and MMP-3 was analyzed within the A-MSC pellets from the coculture relative to the housekeeper (GAPDH). CD3 gene expressed increased in nonchondrogenic and chondrogenically differentiated hMSC pellets cultured with memory T cells
IL-6 is highly secreted in supernatants from cocultures of memory T cells and nonchondrogenic and chondrogenic A-MSC pellets
To assess the effect of differentiated A-MSC pellets on the secretion of CD4 T cell-specific cytokines, all supernatants were harvested after 3 days of coculture of memory T cells with nonchondrogenic and chondrogenically differentiated A-MSC pellets. Nonchondrogenic and chondrogenically differentiated A-MSC pellets had no effect on the secretion of IL-17A (Fig. 4A (i)). There was a trend toward a decrease in the secretion of IL-17F in supernatants of memory T cells with nonchondrogenic and chondrogenic hMSC pellets, however, this was not significant (Fig. 4A (ii)). IL-6 was highly secreted in cocultured supernatants only (Fig. 4A (iii)). While supernatants from nonchondrogenic and chondrogenic A-MSC pellets had low levels of IL-6 secretion, IL-6 was not detected in supernatants from memory T cells alone. There was also a trend toward a decrease in IL-10 and IFNγ secretion in coculture supernatants of memory T cells with nonchondrogenic and chondrogenic A-MSC pellets, however, again this was not significant (Fig. 4A (iv) and (v)). Furthermore, IL-4 was not detected in any supernatants (data not shown). A-MSC pellets that were cocultured with memory T cells had significantly higher gene expression levels of IL-6 compared with memory T cells from cocultures. However, no significant difference was observed between IL-6 gene expression levels in nonchondrogenic or chondrogenically differentiated MSCs (Fig. 4B). Gene expression levels of IL-10 did not significantly differ between culture conditions.
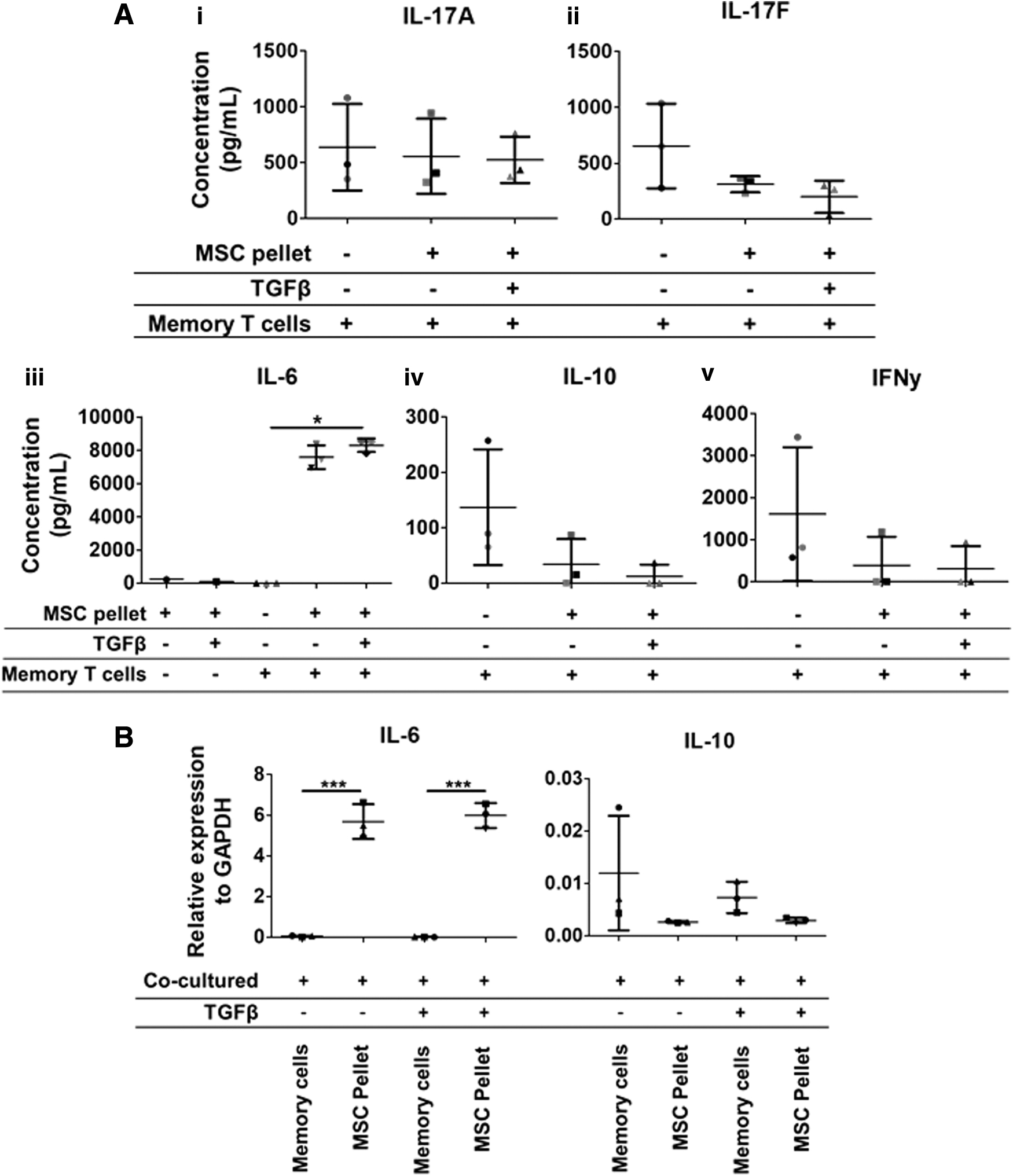
IL-6 is highly secreted in supernatants from cocultures of memory T cells and nonchondrogenic and chondrogenic A-MSC pellets. Memory T cells were cocultured with 7-day nonchondrogenic and chondrogenically differentiated A-MSCs for 3 days. The level of IL-17A, IL-17F, IL-6, IL-10, and IFNγ was analyzed in the supernatants from the coculture by ELISA. IL-6 was secreted at a significantly higher level in supernatants from cocultures of memory T cells with nonchondrogenic and chondrogenically differentiated A-MSC pellets. All other cytokines were not significantly affected
P-MSC pellets at day 21 of chondrogenesis skew Th1/Th2 as well as Th17 and regulatory T cell subsets
To confirm the change in the ratio of Th2 versus Th1 in memory T cells at a later stage of chondrogenesis, P-MSCs were differentiated with and without TGFβ3 for 21 days. Following 21 days of differentiation, both nonchondrogenic (−TGFβ3) and chondrogenically differentiated (+TGFβ3) P-MSCs were cocultured with unsorted PBMCs or sorted memory CD4+ T cells for 3 days. After 21 days of differentiation, chondrogenesis of the P-MSC donor was confirmed (Supplementary Fig. S4 and S5). A trend toward a decrease in the percentage of Th1 cells was observed in unsorted PBMCs cocultured with allogeneic nonchondrogenic and chondrogenically differentiated P-MSC pellets (26.6% ± 7.1% vs. 20.5% ± 5.1% and 19.5% ± 4.9%) (Fig. 5A (i)). A small yet significant increase was found in the percentage of Th2 cells in cocultures of unsorted PBMCs with allogeneic nonchondrogenic and chondrogenically differentiated P-MSCs (11.9% ± 2.8% vs. 15.1% ± 3.3% and 16.7% ± 4.3%) (Fig. 5A (ii)). Furthermore, a small yet significant increase in the percentage of Th17 cells was observed in unsorted PBMCs cocultured with allogeneic chondrogenically differentiated P-MSC pellets (25% ± 5.6% vs. 29.4% ± 4.5%), but this was not statistically significant with allogeneic nonchondrogenic P-MSC pellets (25 ± 5.6 vs. 27.2 ± 5.5) (Fig. 5A (iii)). Furthermore, a significant decrease in the percentage of regulatory T cells was noted in cocultures with both nonchondrogenic and chondrogenically differentiated P-MSC pellets (55.3% ± 3.3% vs. 43.3% ± 8.2% and 40.6% ± 4.6%) (Fig. 5A (iv)). The gating strategy for regulatory T cells was the same as previous studies 5 and is also shown in Supplementary Figure S3. Similar to the previous experiment, a trend toward a decrease in Th1 cells was observed in memory T cells cocultured with allogeneic nonchondrogenic and chondrogenically differentiated P-MSC pellets (15.8% ± 7.6% vs. 12.2% ± 6.5% and 11.2% ± 5.1%), whereas Th2 cell differentiation was significantly increased by both treatment conditions (35% ± 8.2% vs. 46.8% ± 11.1% and 51.4% ± 10.5%) (Fig. 5B (i) and (ii)). Furthermore, the percentage of Th17 cells was significantly increased in memory T cells cocultured with allogeneic nonchondrogenic and chondrogenically differentiated P-MSCs (27% ± 4.3% vs. 39.1% ± 5.9% and 40.2% ± 6.7%) (Fig. 5B (iii)). On the other hand, the percentage of regulatory T cells in memory T cells cocultured with nonchondrogenic and chondrogenically differentiated P-MSC pellets was significantly inhibited (30% ± 5.9% vs. 15.5% ± 4.6% and 12.7% ± 4.1%) (Fig. 5B (iv)).
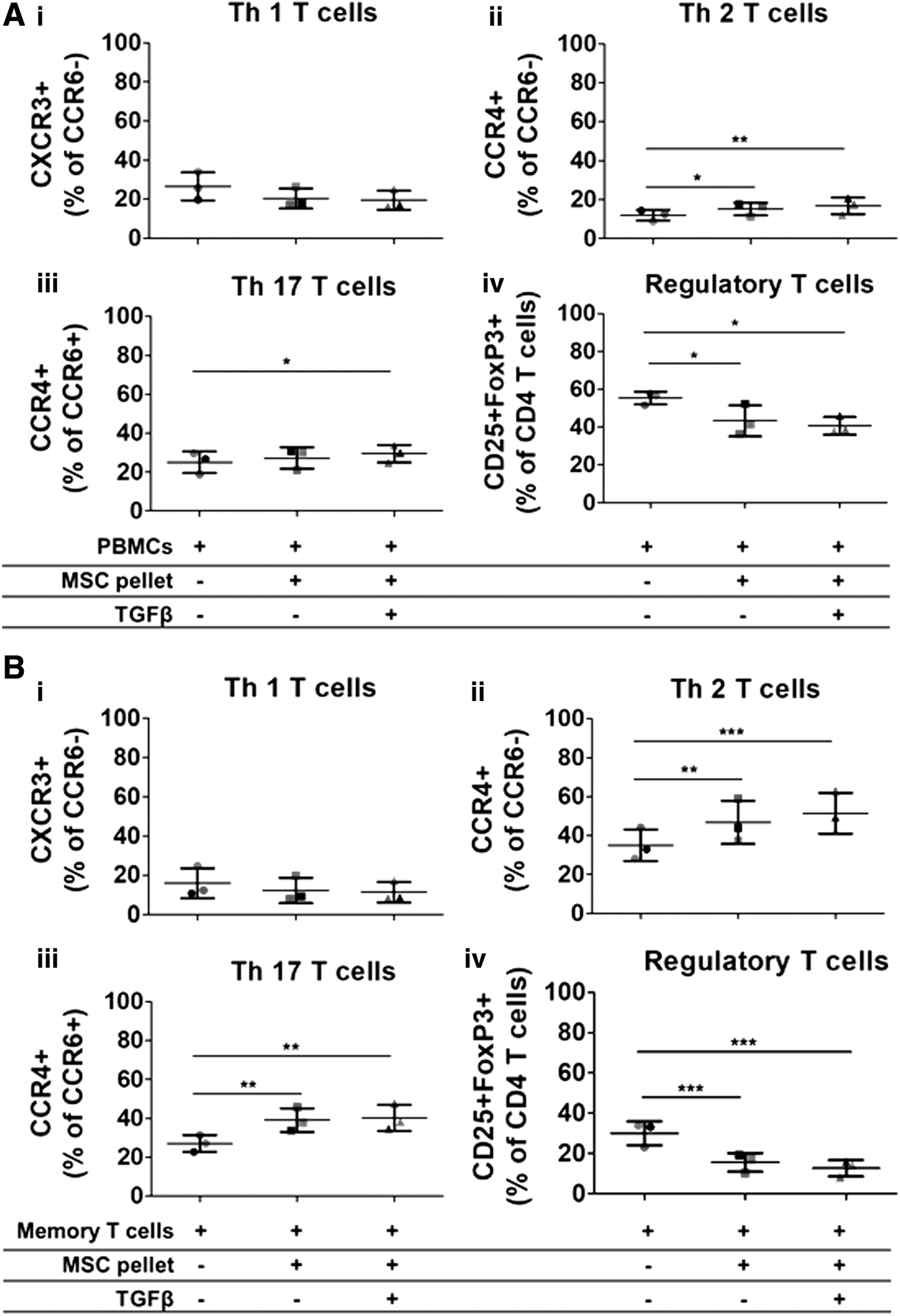
P-MSC pellets at day 21 of chondrogenesis skew Th1/Th2 as well as Th17 and regulatory T cell subsets. PBMCs or CD3/CD28-stimulated memory T cells were cultured with 21-day nonchondrogenic and chondrogenically differentiated P-MSCs for 3 days. Following coculture, PBMCs or memory T cells were harvested and analyzed by flow cytometry for T cell-specific markers (CD45, CCR6, CCR4, CXCR3, CD25, CD127, and FoxP3). Nonchondrogenic and chondrogenically differentiated P-MSCs induce an increase in the percentage of Th17 and Th2 cells in unsorted PBMCs
Chondrogenic differentiation alters cytokine secretion in coculture with memory T cells
To assess the effect of differentiated P-MSC pellets on the secretion of CD4+ T cell-specific cytokines, all supernatants were harvested after 3 days from cocultures of memory T cells with 21-day allogeneic nonchondrogenic or chondrogenically differentiated P-MSC pellets. Levels of IL-17A, IL-17F, IL-4, IL-10, and IFN-γ secreted in supernatants of memory T cells with either allogeneic nonchondrogenic or chondrogenic P-MSC pellets did not significantly differ (Fig. 6). However, coculture of memory T cells with allogeneic chondrogenically differentiated P-MSC pellets resulted in significantly higher levels of IL-6 secretion, compared with coculture with nonchondrogenically differentiated P-MSCs or cultures of memory T cells alone (Fig. 6). Interestingly, the levels of IL-6 in 21-day chondrogenic P-MSC pellets were about six times higher compared with 7-day chondrogenic A-MSC pellets cocultured with memory T cells (Fig. 4). Both nonchondrogenic and chondrogenically differentiated P-MSC pellets cultured alone did not secrete any IL-6.

Chondrogenic differentiation alters cytokine secretion in coculture with memory T cells. CD3/CD28-stimulated memory T cells were cultured with 21-day nonchondrogenic and chondrogenically differentiated P-MSCs for 3 days. The level of IL-17A, IL-17F, IL-4, IL-6, IL-10, and IFNγ was analyzed in supernatants from cocultures by ELISA. Coculture of memory T cells with allogeneic chondrogenically differentiated P-MSC pellets results in significantly higher levels of IL-6 secretion, compared with coculture with nonchondrogenically differentiated P-MSCs or cultures of memory T cells alone. n = 3 PBMC donors. Data are represented as mean ± SD. Repeated measures ANOVA with Bonferroni's posthoc test: **p < 0.01.
Discussion
CD4 T lymphocytes have been previously shown to affect bone regeneration mediated by undifferentiated MSCs. The aim of the current study was to investigate the effect of allogeneic chondrogenically differentiated MSC pellets on Th1, Th2, Th17, and regulatory T cell subsets in vitro. We have shown that allogeneic nonchondrogenic and chondrogenically differentiated MSC pellets induce changes in the percentage of memory Th1, Th2, Th17, and regulatory T cells. Furthermore, IL-6 appears to be a predominant cytokine involved in this interaction between allogeneic chondrogenically differentiated MSC pellets and CD4 T cells.
The differentiation of MSCs has been shown to affect their immunogenic properties. 36 Our group has extensively reviewed the effect of these differentiated MSCs on the immune system from various research groups. 37 Previous studies have shown that undifferentiated MSCs suppress more proinflammatory T cell subsets (Th1 and Th17).38–42 While the majority of studies do not focus on the effects of MSCs on specifically Th2 T cell differentiation, additional studies have reported that undifferentiated MSCs induce a shift in the cytokine profile in the T cell differentiation process toward anti-inflammatory Th2 cells. 43 Furthermore, in an animal model of multiple sclerosis, MSC injection into these animals was shown to induce a Th2 response but inhibit a Th1 response. 44 Therefore, MSCs seem to promote Th2 responses. While the study by Liu et al. reported an inhibition of Th1, Th2, and Th17 differentiation in their in vitro coculture system, upon infusion of the MSCs in vivo, the levels of IFNγ and TNFα were reduced but not levels of IL-4, suggesting that Th2 cells may still be induced by MSCs in vivo. 45 The fact that both nonchondrogenic and chondrogenic MSCs were found to increase Th2 cells and decrease Th1 cells suggests that, in our culture system, chondrogenically differentiated MSC pellets have similar immunomodulatory properties as undifferentiated and nonchondrogenic cells. Therefore, allogeneic chondrogenic MSCs are not expected to induce a negative alloresponse in vivo.
TGFβ priming of MSCs is required to induce successful chondrogenic differentiation of these cells. Implantation of chondrogenically differentiated MSC pellets generates bone in vivo through the process of endochondral ossification. 2 In this study, we were also interested to compare allogeneic nonchondrogenic MSC pellets, which did not receive TGFβ during the differentiation process, with allogeneic chondrogenically differentiated MSC pellets. Nonchondrogenically differentiated MSCs do not form bone when implanted in vivo. 4 Therefore, it was hypothesized that there would be a difference in the activation of specific T cell subsets by either allogeneic nonchondrogenic or chondrogenically differentiated MSCs pellets that may give more information as to which T cell subset(s) may be involved in the MSC-mediated endochondral bone formation. Our group has shown that 7-day chondrogenic priming of the MSCs is sufficient to generate bone in vivo. 46 Therefore, in the first part of this study MSC, pellets were differentiated for 7 days with and without TGFβ3 before coculture with the lymphocytes. Based on histology, at 7 days the GAG production was quite low in the A-MSC donor used and therefore there was not much difference in matrix production between nonchondrogenic and chondrogenically differentiated A-MSC pellets. This may explain why there was little difference in the differentiation of the CD4+ T cell subsets between allogeneic nonchondrogenic and chondrogenically differentiated A-MSC pellets cocultured with either unsorted PBMCs or memory T cells. The experiment was repeated with MSCs chondrogenically differentiated for 21 days before the coculture to determine whether a more chondrogenic donor would still skew the Th1/Th2 balance and whether this differs from the nonchondrogenic MSC pellet coculture. Again, an increase in the Th2 cells and a decrease in Th1 cells was observed. Regulatory T cells were also included in the analysis, where it is known that TGFβ can induce regulatory T cell differentiation and TGFβ + IL-6 can induce Th17 cell differentiation. 20 A significant increase in the percentage of Th17 cells and a decrease in the percentage of regulatory T cells in cocultures with 21-day nonchondrogenic and chondrogenically differentiated P-MSC pellets was observed in the current study. This could be due to the high production of IL-6 by the MSCs to increase the percentage of Th17s and decrease the percentage of regulatory T cells. However, there were still no major differences between cocultures of nonchondrogenic and chondrogenically differentiated MSC pellets and T cells. It is important to note that Th17 cells are not the only subtype of CD4 T cells to express CCR6 and CCR4. Th22 cells are also known to play a role during tissue inflammation and can be excluded from Th17 cells with the chemokine CCR10 and the production of the cytokine IL-22.47,48 The percentage of Th22 cells is relatively low compared with Th17 cells 49 and while this chemokine and cytokine were not included in the current study, Th22 cells cannot be excluded, in that they did not play a role in the interaction between allogeneic nonchondrogenic and chondrogenic MSC pellets. Future studies are required to further study perhaps the role of these cells in the interaction between allogeneic chondrogenic MSCs and the immune system.
Perhaps the most interesting cells altered by nonchondrogenic and chondrogenically differentiated pellets were those found within the matrix. In a previous study, CD3 immunohistochemistry was performed on MSC pellets cocultured with PBMCs. In this study, we showed that CD3 T cells can in fact infiltrate the matrix of the MSCs, which was also supported by increased CD3 gene expression in only those MSCs cocultured with PBMCs. 5 In the current study, CD3 gene expression was significantly upregulated in nonchondrogenic and chondrogenically differentiated A-MSC pellets cocultured with memory T cells. Nonchondrogenic and chondrogenic MSC pellets were negative for CD3 gene expression. This suggested that memory T cells had infiltrated the matrix of the cells. While no differences were found in the differentiation of T cells between nonchondrogenic and chondrogenic A-MSCs, perhaps these infiltrating cells are the cells where most difference would be found. It would be interesting to attempt to analyze these infiltrating cells in future studies to determine what cells invade the matrix of the MSCs and their role in the whole process of the bone regeneration.
IL-6 is known to be secreted at large quantities by MSCs.43,50 The functions of IL-6 are both pro- and anti-inflammatory. It has been reported that MSCs secrete IL-6 to prevent apoptosis and maintain not only their immunoprivileged state, but also their proliferation. 34 Furthermore, IL-6 has been shown to be an important cytokine in the rejection of allogeneic MSCs following implantation, whereby, in a study by Li et al., IL-6 prevented allogeneic leukocyte-mediated cytotoxicity of differentiated MSCs. 51 The enhanced IL-6 production by the MSCs prevented their rejection by the host to allow tissue repair and regeneration to occur. In our cocultures of memory T cells and 7-day nonchondrogenic and chondrogenically differentiated A-MSC pellets, IL-6 was found to be highly secreted in the supernatants. Interestingly, upon protein analysis, differences were now arising between the cytokine secretions of cocultures of memory T cells with allogeneic nonchondrogenic and chondrogenically differentiated P-MSC pellets. With 21-day chondrogenic P-MSC pellets, levels of IL-6 were significantly higher in cocultures with memory T cells compared with nonchondrogenic P-MSC cocultures. Furthermore, the levels of IL-6 were about six times higher in 21-day chondrogenic P-MSC pellets compared with 7-day chondrogenic A-MSC pellets. This was most likely due to the fact that the P-MSC pellets were more chondrogenic than the 7-day A-MSC pellets. The IL-6 found in the cocultures could have had a variety of purposes, including as already mentioned having an effect on the regulatory T cell/Th17 cell ratio. First, since IL-6 has been shown to be involved in allogeneic stem cell rejection, perhaps allogeneic chondrogenically differentiated MSC pellets secrete IL-6 to suppress host immune cells from initiating rejection. Previously, we have shown that chondrogenically differentiated MSC pellets are responsible for the IL-6 production and not dendritic cells. 52 We have also further confirmed this finding in the current study by demonstrating high IL-6 gene expression in MSC pellets and not memory T cells. In light of these findings, we hypothesize that chondrogenic MSC pellets are also responsible for the IL-6 production in response to these immune cells. Furthermore, IL-6 has been previously shown to induce IL-4-producing CD4 T cells. 44 IL-4 and IL-13 (both Th2-related cytokines) have been reported to be chemotactic factors for osteoblasts. 53 Whether chondrogenically differentiated MSCs induce Th2 T cell differentiation to acquire osteoprogenitors in vivo, requires additional investigation. Further studies investigating the role of IL-6 in MSC-mediated endochondral bone formation need to be completed to confirm its role in the whole process; however, these results suggest that chondrogenic MSC pellets produce IL-6 during the immune response and that this cytokine may be important in their capacity to alter T cell behavior.
In conclusion, allogeneic chondrogenically differentiated MSCs increase Th2 cells, as well as skew Th17 and regulatory T cell subsets in sorted memory CD4 T cells in vitro. The change in Th2 and Th17 cell ratio by allogeneic chondrogenic MSC pellets, highlights a potential role of Th2 or Th17 T cells in the bone formation process mediated by chondrogenic MSC pellets in vivo. Additionally, IL-6 appears to be an important cytokine in the interaction between allogeneic chondrogenic MSC pellets and memory T cells. Additional in vivo analysis investigating bone formation by chondrogenically differentiated MSCs in an allogeneic setting could confirm the involvement of these T cell subsets in the early stages of bone regeneration mediated by chondrogenically differentiated MSC pellets.
Footnotes
Author Contributions
C.H.K., E.L., and E.F. conceived and designed the study. C.H.K., P.S.A., E.F., and E.L. acquired, analyzed, and interpreted data and drafted and edited the article. N.F. and J.W.-B. were involved in acquiring experimental data, editing, and critical review of the article. E.B.W. was involved in the conception, design, and critical review of the article. P.A.J.B. was involved in the conception, design, analysis, interpretation of data, and critical review of the article. All authors revised the current article and gave final approval of the current version to be published. All authors are accountable for all aspects of the work in ensuring that questions related to the accuracy and integrity of any part of the work are appropriately investigated and resolved.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the AO Foundation, Switzerland (AOCMF-14-12F and AOCMF-15-27F).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.