
Abstract
To obtain stable outcomes in regenerative medicine, the quality of cells for transplantation is of great importance. Cellular stress potentially results in the release of damage-associated molecular patterns (DAMPs) and activates immunological responses, affecting the outcome of transplanted tissue. In this study, we intentionally prepared necrotic chondrocytes that would gradually die and release DAMPs and investigated how the maturation of tissue-engineered cartilage was affected. Necrotic chondrocytes were prepared by a conventional heat-treatment method, by which their viability started to decrease after 24 h. When tissue-engineered cartilage containing necrotic chondrocytes was subcutaneously transplanted into C57BL/6J mice, accumulation of cartilage matrix was decreased compared to the control. Meanwhile, immunohistochemical staining demonstrated that localization of macrophages and neutrophils was more apparent in the constructs of necrotic chondrocytes, suggesting that DAMPs from necrotic chondrocytes could prompt migration of more immune cells. Two-dimensional electrophoresis and mass spectrometry identified prelamin as a significant biomolecule released from necrotic chondrocytes. Also, when prelamin was added to a culture of RAW264, Inos and Il1b were increased in accordance with the content of added prelamin. It was suggested that DAMPs from dying chondrocytes could induce inflammatory properties in surrounding macrophages, impairing the maturation of tissue-engineered cartilage. In conclusion, maturation of tissue-engineered cartilage was hampered when less viable chondrocytes releasing DAMPs were included.
Impact statement
In regenerative medicine, the quality of cells is of great importance to secure clinical safety. During culture, damage of cells could occur, if not critical enough to cause immediate cell death, but still inducing a less viable status. Damage-associated molecular patterns (DAMPs) are released from necrotic cells, but their influence in regenerative medicine has yet to be clarified. In this study, we elucidated how DAMPs from chondrocytes could affect the maturation of tissue-engineered cartilage. Also, possible DAMPs from necrotic chondrocytes were comprehensively analyzed, and prelamin was identified as a significant molecule, which may serve for detecting the existence of necrotic chondrocytes.
Introduction
Cartilage is present in the nose, ear, trachea, and joints, serving to maintain morphological form and support activities of daily life. Since cartilage lacks blood vessels and has poor capacity of healing, transplantation of cartilaginous tissues can be a reasonable choice for damaged cartilage. However, autologous cartilage is mostly unavailable for that purpose due to limited autograft availability and donor site morbidity, necessitating the development of cartilage regeneration. Autologous chondrocyte implantation (ACI) in humans was first reported in 1994, in which autologous chondrocytes from a healthy nonweight-bearing site in a knee joint were expanded in vitro, and then transplanted into defective sites. 1 Since then, the indications for ACI have been expanded to the oral and maxillofacial region, such as nasal augmentation and ear reconstruction.2–4 Our group has developed tissue-engineered cartilage with rigidity and three-dimensional (3D) morphology, consisting of autologous chondrocytes and a biodegradable polymer scaffold, and applied it in clinical research in patients with nasal deformity associated with cleft lip and palate (JPRN-UMIN000005472). 2
It is widely accepted that cellular stress results in the release of damage-associated molecular patterns (DAMPs), which are sensed by the receptors of innate immune cells, activating immunological responses. 5 DAMPs are (1) exposed on the plasma membrane of stressed cells, or (2) actively secreted by stressed cells or cells in the early stage of apoptosis, or (3) passively released into the extracellular environment from dying cells with plasma membrane rupture, or (4) released from damaged extracellular matrix.5–7 Protein DAMPs include intracellular proteins such as heat-shock proteins, high-mobility group box 1 (HMGB1), and extracellular matrix generated by tissue injury, while nonprotein DAMPs include ATP, uric acid, heparin sulfate, RNA, and DNA. 8 These DAMPs are sensed by innate immune effectors, including the family of toll-like receptors (TLRs) and the NOD-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome complex. TLRs exhibit the most diverse repertoire of DAMP ligands among all innate immune receptors, controlling potent downstream inflammatory mediators, such as interleukin (IL)-1, IL-6, IL-8, tumor necrosis factor (TNF)-β, and IL-12. 9 Meanwhile, the NLRP3 inflammasome regulates caspase-1 activation, which is required for the secretion of cytokines such as IL-1β and IL-18.10,11
In case of regenerative medicine with cell transplantation, the procedure occasionally includes a process of cell culture and making constructs, potentially inducing damage of cells and release of DAMPs. DAMPs could vary greatly depending on the type of cells and injured tissue,12,13 and their details and effects in cartilage regenerative medicine have yet to be clarified. Activation of inflammatory responses by DAMPs be destructive to tissue-engineered cartilage, given there is an association between inflammation and cartilage degradation in joint disease, such as rheumatoid arthritis and osteoarthritis. 14 In rheumatoid arthritis, blood cells and inflammatory mediators migrate into the joints, resulting in synovial hyperplasia. As a result of this process, both the synovial membrane and other joint structures, cartilage and bone, are damaged. 15 Also, the invasion of cartilage by proinflammatory cells leads to degradation of type II collagen by matrix metalloproteinases and other enzymes, which are produced by synovial cells and chondrocytes when stimulated by inflammatory cytokines. 16
In the present study, we investigated how less viable chondrocytes could affect the outcome in transplantation of tissue-engineered cartilage and also examined the details of DAMPs released from chondrocytes.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium:nutrient Mixture F-12 (DMEM/F12), penicillin-streptomycin solution, and trypsin-EDTA solution were from Sigma Chemical Co. (MO). Collagenase from Clostridium histolyticum was from Wako Pure Chemical Industries (Osaka, Japan). Insulin was from MP Biomedicals (Irvine, CA), and FGF-2 was from Kaken Pharmaceutical Co., Ltd. (Tokyo, Japan). Other materials included 3% atelocollagen implants from Koken Co., Ltd. (Tokyo, Japan) and poly-L-lactic acid (PLLA) porous scaffolds from KRI (Kyoto, Japan). Anti-F4/80 antibody (Cl: A3-1) from Abcam (Cambridge, UK), antineutrophil antibody (Cl: NIMP-R14) from GeneTex (Irvine, CA), anti-type I collagen antibody from LSL (Tokyo, Japan), anti-type II collagen antibody from LSL, antiproliferating cell nuclear antigen (PCNA) antibody from Abcam, anti-lamin A/C antibody (N-18) from Santa Cruz Biotechnology (Dallas, TX), anti-beta actin antibody from Sigma-Aldrich (MO, US), horseradish peroxidase-conjugated secondary antibody from Promega (MI), and recombinant prelamin-A from Prospec-Tany Technogene Ltd. (Rehovot, Israel) were purchased. Mammalian protein extraction reagent (M-PER) from Pierce Biotechnology (MA) and ECL from Amersham Biosciences (Buckinghamshire, UK) were used. The biotinylated secondary antibody, VECTASTAIN Elite ABC Kit, and Peroxidase Substrate kit DAB were all from Vector Laboratories (CA). Isogen from Nippon Gene (Tokyo, Japan), PrimeScript Reverse Transcriptase from Takara Shuzo (Shiga, Japan), and Platinum SYBR Green qPCR SuperMix from Invitrogen (CA) were used. C57Bl6/J mice were obtained from Nisseizai (Tokyo, Japan).
Cell culture
Mouse auricular chondrocytes were obtained and cultured as described previously. 17 In brief, the ear and external ear canal of mice were digested with 0.15% collagenase solution for 8 h, and the isolated chondrocytes were cultured in DMEM/F12 supplemented with 5% fetal bovine serum (FBS), 5 μg/mL insulin, and 100 ng/mL FGF-2. 18 All the procedures for the present experiments were approved by the Institutional Committee for animal research of the University of Tokyo Hospital. To prepare gradually dying chondrocytes (necrotic cells), a culture dish of chondrocytes in P2 was placed in an oven at 55°C for 0.5, 1, or 1.5 h and washed with phosphate-buffered saline (PBS) twice. As control cells (normal cells), a culture dish of chondrocytes in P2 was left in an incubator and then washed with PBS in the same manner as for necrotic cells (n = 4). For analysis of the time-course change of viability and proliferation, necrotic and normal cells were kept cultured in the same medium described above. Total cell number and dead cell number in dishes were counted using an automated cell counter (NucleoCounter®) from ChemoMetec A/S (Allerod, Denmark) (n = 6). To detect uric acid content in the medium, the medium of chondrocytes heat-treated at 55°C for 0.5 h was collected just after and 1, 2, 3, 6, and 24 h after heat-treatment. Collected samples were analyzed using a Uric Acid Assay Kit from BioAssay Systems (Hayward, CA) (n = 3).
For the culture of 3D pellets, chondrocytes in 1% atelocollagen gel were placed in a 15-mL conical tube (2 × 105 cells/20 μL) and incubated at 37°C/5% CO2 for 2 h. After being added to 2 mL of DMEM/F12, 3D pellets were cultured for 7 days. When examining the effects of prelamin on chondrocytes, recombinant prelamin protein was added in the medium.
The mouse macrophage-like cell line RAW264 was purchased from the Riken Cell Bank (Tsukuba, Japan), and expanded in MEM supplemented with 10% FBS and 0.1 mM non-essential amino acids (NEAA). When RAW264 reached subconfluence, the medium was changed to MEM supplemented with 1% FBS and 0.1 mM NEAA containing recombinant prelamin protein.
RNA isolation and real-time reverse transcription polymerase chain reaction
Total RNA of chondrocytes or RAW264 was isolated with Isogen, and then reverse-transcribed with reverse transcriptase and random hexamers according to the manufacturer's protocol. Gene expression was detected by real-time reverse transcription polymerase chain reaction (real-time RT-PCR) using the standard SYBR Green method with a 7500 fast real-time PCR system (Applied BioSystems, CA) (n = 3). Standard templates were produced according to a previous study. 19 Sequences of primers were 5′-GAAACCCGAGGTATGCTTGA-3′ and 5′-GGGTCCCTCGACTCCTACAT-3′ for type I collagen αI chain (Col1a1); 5′-GGCAAAGATGGCTCTAATGG-3′ and 5′-CGTCGTGCTGTCTCAAGGTA-3′ for type II collagen αI chain (Col2a1); 5′-CACCTTGGAGTTCACCCAGT-3′ and 5′-ACCACTCGTACTTGGGATGC-3′ for inducible nitric oxide synthase (Inos); 5′-AGTTGCCTTCTTGGGACTGA-3′ and 5′-TCCACGATTTCCCAGAGAAC-3′ for Il6; and 5′-AACTTTGGCATTGTGGAAGG-3′ and 5′-ACACATTGGGGGTAGGAAC-3′ for glyceraldehyde-3-phosphate dehydrogenase (Gapdh). Settings for thermal cycling were adjusted according to the manufacturer's protocols. Transcript levels were normalized to that of Gapdh. Relative ratio was calculated by dividing the value of each sample by that of the control sample.
Transplantation of tissue-engineered cartilage in mice
PLLA scaffolds of 4 × 4 × 3 mm3 were sterilized in 70% ethanol before use. Then, 1% atelocollagen gel, which was diluted from original 3% atelocollagen gel with DMEM/F12, was used as a cell suspension buffer to retain the chondrocytes in the scaffolds efficiently. To make engineered constructs, chondrocytes were heat-treated for 0.5 h (necrotic) and control chondrocytes (normal) were suspended in 1% atelocollagen gel (2 × 107 cells/200 μL) and applied to PLLA scaffolds. The constructs were incubated at 37°C/5% CO2 for 2 h before transplantation. C57BL/6J mice were anesthetized by intraperitoneal injection of sodium pentobarbital (50 mg/kg). A small incision was made on the back in the midline, and the constructs were inserted subcutaneously. After 2 weeks, the constructs were harvested and cut into equal parts (n = 6); one of them was used for mechanical and biochemical analysis, and the other part was harvested and fixed in 4% paraformaldehyde for 3 h, embedded in paraffin, and cut into 8-μm sections for histological and immunohistochemical analysis. All the procedures for the present experiments were approved by the Institutional Committee for animal research of the University of Tokyo Hospital.
Mechanical properties
The mechanical properties of tissue-engineered cartilage were analyzed using a Venustron tactile sensor (Axiom, Fukushima, Japan). 20 Under computer control, the motor-driven sensor unit automatically presses down on objects up to 0.5 mm in depth from the surface and provides the compression strength and the decrease in resonant frequency. Young's modulus was calculated from the compression strength and the frequency decrease using the software Venus 42 (Axiom) (n = 6).
Glycosaminoglycan measurement
Following the measurement of Young's modulus of tissue-engineered cartilage, the constructs were cut into small pieces with scissors, and suspended in10 mg/mL pepsin, 0.05 M acetic acid. After digestion at 4°C for 48 h, 1 mg/mL pancreatic elastase, 0.1 mM Tris, 0.02 M NaCl, and 5 mM CaCl2 (pH 7.8–8.0) were added, and the samples were kept at 4°C overnight. The samples were then centrifuged at 9100 g for 5 min, and sulfated glycosaminoglycan (sGAG) content in the supernatant was measured using a Blyscan Glycosaminoglycan Assay Kit (Biocolor Ltd., County Antrim, UK). The spectrophotometrical absorbance of the mixture was measured at a wavelength of 656 nm.
Histological and immunohistochemical staining
The sections were stained with toluidine blue to detect proteoglycan and with hematoxylin and eosin (HE staining). The sections were also used for immunohistochemical staining for COL1, COL2, F4/80, neutrophils, and PCNA according to previous studies.21,22 In brief, the sections were deparaffinized in xylene, and hydrated with step-down concentrations of ethanol. After antigen retrieval, the sections were treated with 10% normal serum for 10 min at room temperature to reduce nonspecific binding of antibodies. Incubation with primary antibodies (anti-F4/80 antibody, 1:100 dilution for 1 h at 37°C; anti-PCNA antibody, 1:500 dilution for overnight at 4°C; other antibodies, 1:200 dilution for 2 h at room temperature) was carried out. Then, the sections were incubated with biotinylated secondary antibodies for 30 min at room temperature. Consecutive steps for the visualization of positive reactions were conducted using a VECTASTAIN ABC Kit and Peroxidase Substrate Kit DAB according to the manufacturer's instructions. Subsequently, the sections were counterstained with hematoxylin or methyl green. Regarding the sections for immunohistochemical staining for PCNA, five different fields were randomly selected from each section, and the numbers of positive cells were counted. Four sections from different animals were used in each group, and the mean value was used to estimate cell proliferation in tissue-engineered cartilage.
Intraperitoneal injection of chondrocytes
Normal and necrotic chondrocytes were prepared according to the protocol described above, and a total of 2 × 106 chondrocytes in 2 mL Hank's buffered saline were intraperitoneally administered to C57BL/6J mice, respectively. At 6 days, 5 mL Hank's buffered saline was injected into the peritoneal space, and then collected after gentle pumping of the abdomen for 3 min. The collected solution was placed in RPMI containing 10% FBS, and attached cells were collected and counted (n = 3).
Two-dimensional gel electrophoresis and mass spectrometry
The medium of necrotic and normal chondrocytes was collected at 3 days, and the supernatant was recovered by centrifugation at 20,000 g for 30 min at 4°C. The sample was enriched by centrifugal filtration (5 kDa cutoff), and dissolved in a solution containing 5 M urea, 2 M thiourea, 2% CHAPS, 2% SB3-10, and 1% DTT. For two-dimensional (2D) electrophoresis, 60 μg protein was dissolved in 0.2% Pharmalyte, broad range pH 3.0–10.0, and applied to an 18-cm, pH 3–10 nonlinear Immobiline DryStrip (GE Health Care, IL). Isoelectric focusing was performed on a CoolPhoreStar IPG-IEF Type-P system from Anatech Co., Ltd. (Tokyo, Japan). Two-dimensional migration was carried out on a 20 × 21-cm 9–18% acrylamide concentration gradient gel at a constant voltage of 80 V for 17 h. The gel was scanned with a Molecular Imager FX scanner (Bio-Rad, CA), and the images were analyzed using ImageMaster 2D Platinum (GE Health Care). The total volume (%) of matched spots in two gels (from normal chondrocytes and necrotic chondrocytes) was measured, and the spots were listed according to the following criteria: (1) total percentage volume of matched spots in two gels was more than 0.2 and (2) spot ratio of matched spots in two gels was more than 2 (Necrotic > Normal). Two-dimensional electrophoresis was performed twice, and the consistency of the distribution of protein spots was checked. Using one of the 2D electrophoresis gels, the spots with the highest ratio were excised. The proteins of the spots were identified by mass spectrometry.
Western blotting
Necrotic and normal chondrocytes were treated with M-PER based on a protocol provided by the company, and the supernatants were used for western blotting to detect the expression of lamin A/C according to previous studies.19,23 In brief, equal amounts (10 μg) of samples were applied to SDS-PAGE and transferred onto polyvinylidene difluoride membranes. Rabbit polyclonal antibodies against lamin A/C and actin were used as primary antibodies (anti-lamin A/C antibody, 1:50 dilution for 2 h at room temperature; antiactin antibody, 1:500 dilution for 2 h at room temperature). The membrane was incubated with horseradish peroxidase-conjugated secondary antibody for 30 min, and immunoreactive proteins were visualized by ECL.
Statistics
Data are expressed as mean ± standard deviation. Statistical significance was evaluated using Dunnett test for multiple comparisons. A value of p < 0.05 was considered to denote statistical significance.
Results
Time-course change of viability of necrotic chondrocytes
To examine how less viable chondrocytes could affect the maturation of tissue-engineered cartilage, we first investigated the conditions for producing necrotic chondrocytes that would gradually die and release DAMPs. To obtain such chondrocytes, we adopted a conventional method of heating culture dishes. 24 Chondrocytes heat-treated at 55°C for 0.5 h showed similar viability compared to the nontreated control (0 h) at first (Fig. 1A). However, the viability of heat-treated chondrocytes started to decrease after 24 h, dropping to around 60% by 7 days (Fig. 1B). Regarding cell proliferation, heat-treated chondrocytes (0.5 h) did not proliferate with time as did nontreated control (0 h) (Fig. 1C). When observed microscopically, heat-treatment induced more irregular and shrunken chondrocytes at 5 days (Fig. 1D). On the contrary, chondrocytes treated for a longer duration (1 and 1.5 h) showed markedly decreased viability. To examine whether the treated chondrocytes were releasing DAMPs, the content of uric acid, a known DAMP, was measured. In the medium of chondrocytes heat-treated for 0.5 h, uric acid gradually increased up to 24 h (Fig. 1E). As for gene expression, heat-treatment for 0.5 h increased expression of type I collagen (Col1). Meanwhile, expression of type II collagen (Col2) was not so affected (Fig. 1F), indicating that heat-treatment for 0.5 h did not affect chondrogenic properties very severely. Since heat-treatment at 55°C for 0.5 h could induce gradually dying and DAMPs-releasing chondrocytes, we adopted this condition to obtain necrotic chondrocytes for the following experiments.
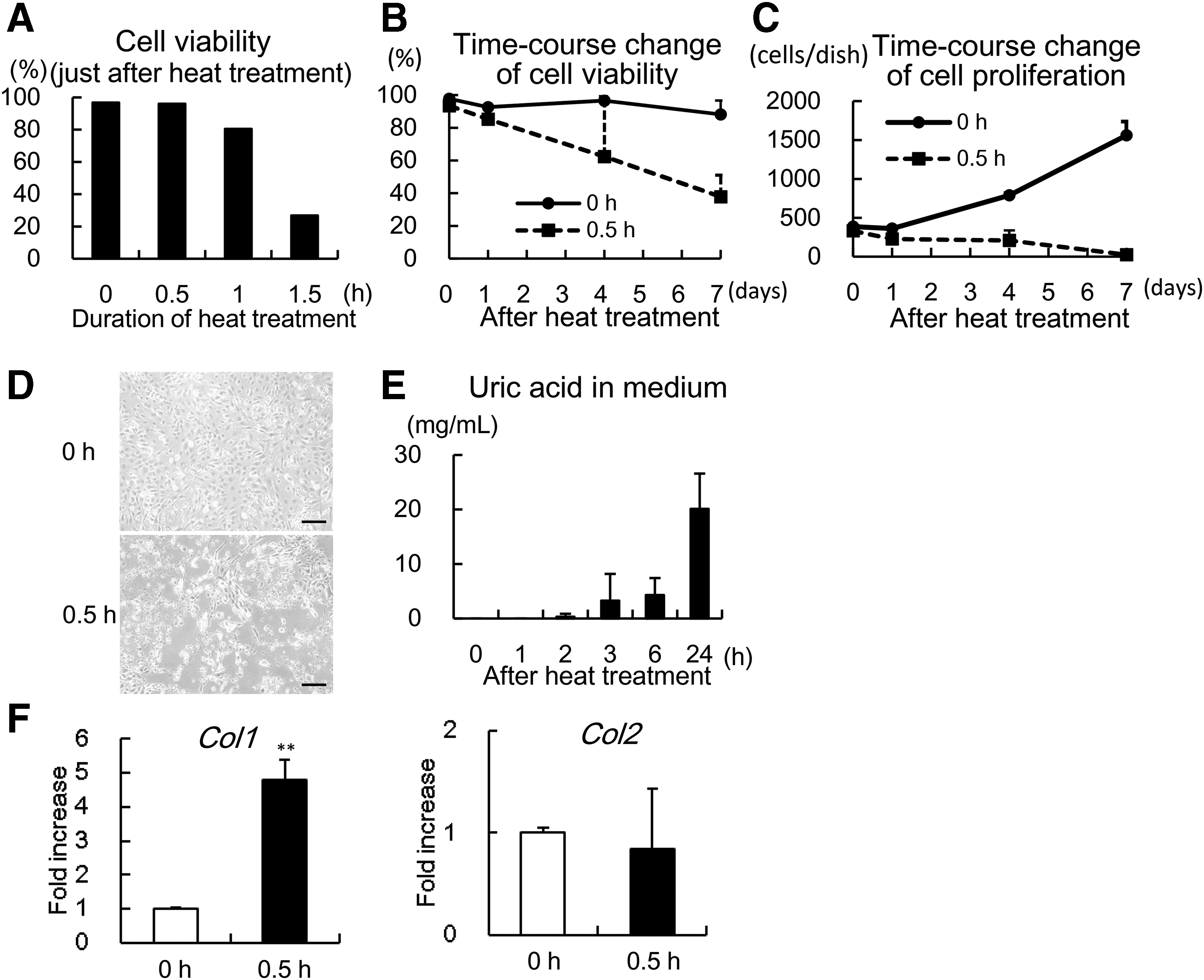
Time-course change of viability of heat-treated chondrocytes.
Transplantation of tissue-engineered cartilage with necrotic chondrocytes
To examine how necrotic chondrocytes could affect the maturation of tissue-engineered cartilage, we made tissue-engineered cartilage using normal and necrotic chondrocytes and transplanted it subcutaneously into C57BL/6J mouse. On toluidine-blue staining, both constructs demonstrated accumulation of cartilage matrix, recognized as metachromatic areas, although the amount was obviously less in the constructs of necrotic chondrocytes (Fig. 2A, TB). On HE staining, in the constructs of normal chondrocytes, we could detect cells with a round, chondrocyte-like appearance, constituting cartilaginous tissue, which were less apparent in the constructs of necrotic chondrocytes (Fig. 2A, HE). Immunohistochemical staining for COL1 and COL2 also demonstrated decreased expression of COL1 and COL2 in the constructs of necrotic chondrocytes (Fig. 2A, COL1 and COL2). Also, the content of sGAG and the value of Young's modulus were significantly decreased in the constructs of necrotic chondrocytes (Fig. 2B, C). These data indicated decreased maturation of tissue-engineered cartilage containing necrotic chondrocytes.
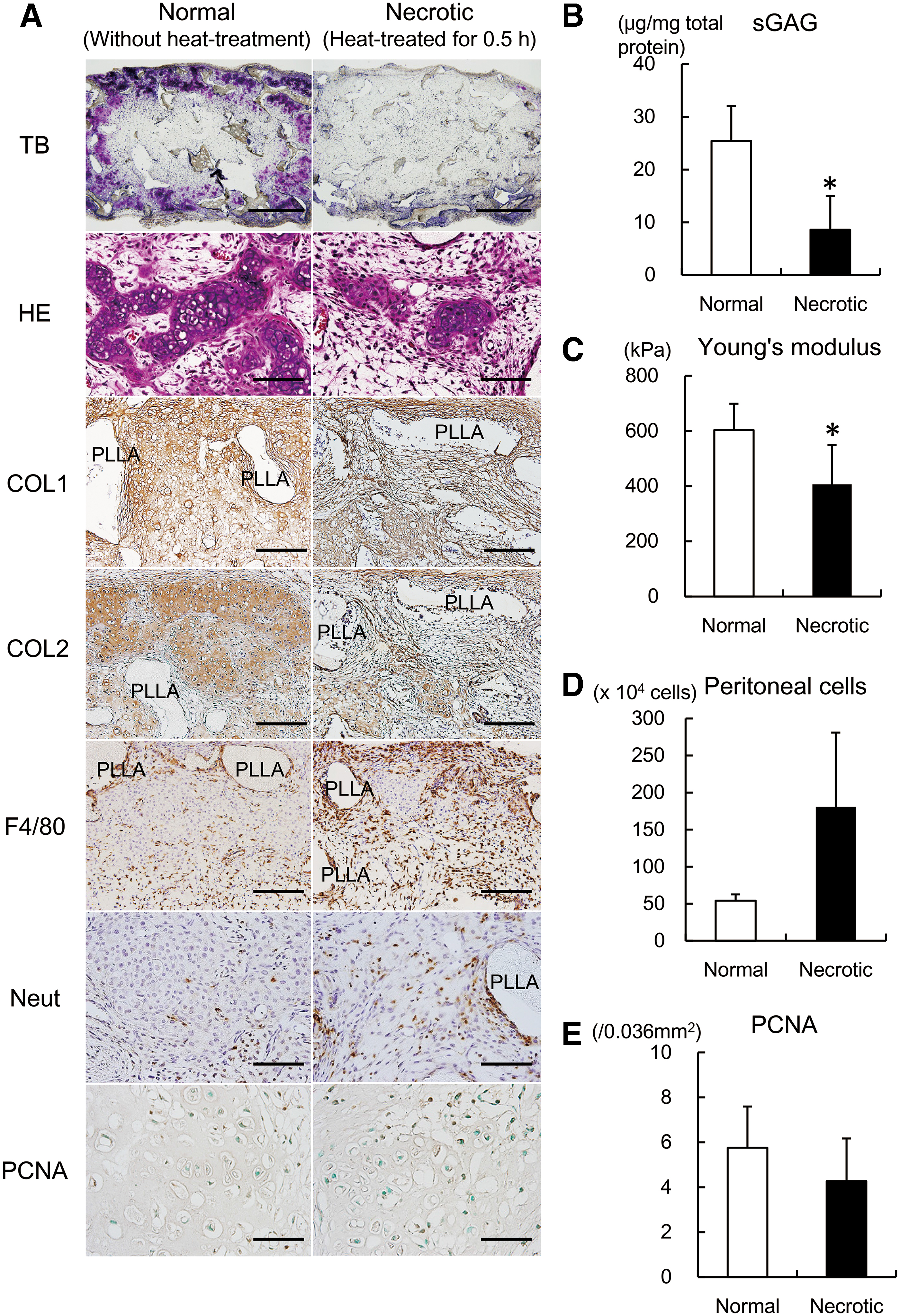
Transplantation of tissue-engineered cartilage using necrotic chondrocytes.
Since the necrotic chondrocytes prepared in the present study were gradually dying within days, DAMPs released from the chondrocytes could trigger immunological reactions after transplantation. We investigated the localization of innate immune cells at 2 weeks after transplantation. Immunopositivity of F4/80 (a marker for macrophages) and neutrophils was much more apparent in the constructs of necrotic chondrocytes (Fig. 2A, F4/80 and Neut). Also, when necrotic chondrocytes were intraperitoneally administered to C57BL/6J mice, the number of accumulated cells in the peritoneal space was higher (Fig. 2D), suggesting that necrotic chondrocytes could be a stronger stimulant for the migration of immune cells to a local site. Meanwhile, the immunopositivity of PCNA was not so apparent and did not differ significantly between both types of constructs (Fig. 2A, PCNA and E).
Identification of damage-associated molecular patterns released from necrotic chondrocytes
Since DAMPs released from cells could vary depending on the type of cells, we next identified the factors released from necrotic chondrocytes. The culture medium of normal and necrotic chondrocytes was collected at 3 days and analyzed by 2D electrophoresis and mass spectrometry. Of the spots recognized in 2D electrophoresis (Fig. 3A), there were 59 spots with a difference in ratio (necrotic/normal >2.0) (Fig. 3B). Then, the spots of gels with the highest ratio were analyzed by mass spectrometry and were identified as prelamin, while the second highest was vimentin (Fig. 3C). Prelamin generally undergoes a process of modifications, including farnesylation, endoproteolytic trimming, and methylation,25–27 and then the last 15 amino acids are clipped off before mature lamin A is released. Since anti-prelamin antibodies were commercially unavailable, we instead utilized anti-lamin A/C antibodies to confirm the expression by western blot analysis (Fig. 3D). The expression of lamin A/C was most evident at 76 kDa in necrotic chondrocytes at 5 days, while slight expression was also detected in normal chondrocytes. On the contrary, severely necrotic chondrocytes, which were heat-treated for 0.75 h, did not show any distinct expression of lamin A/C.
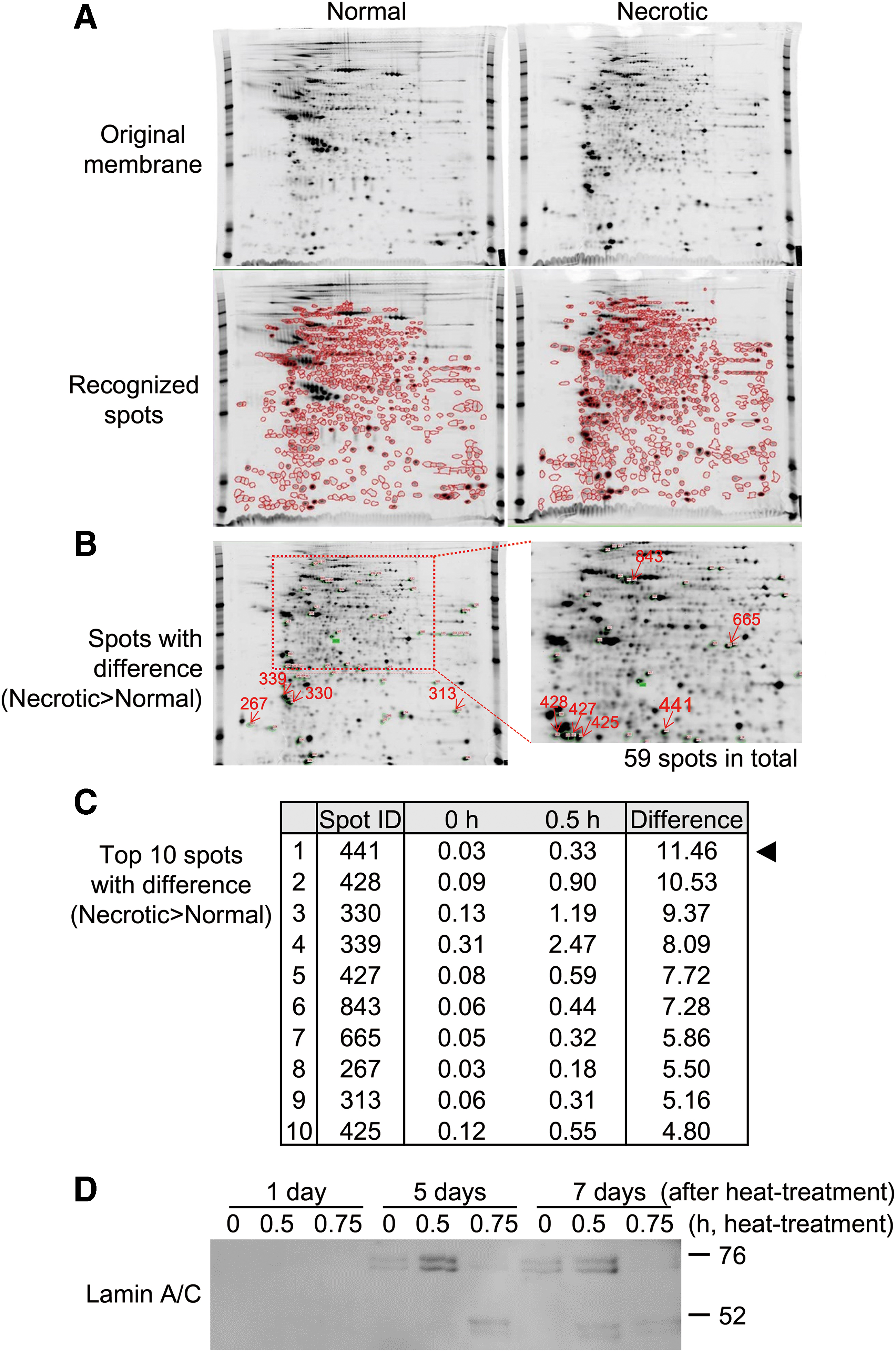
Identification of DAMPs released from necrotic chondrocytes. Culture medium of chondrocytes without heat-treatment (Normal) and with heat-treatment for 0.5 h (Necrotic) were collected at 3 days, and analyzed by 2D electrophoresis and mass spectrometry.
Effects of prelamin on macrophages and chondrocytes
We next examined the immunological effects of prelamin released from necrotic chondrocytes in tissue-engineered cartilage. Since our previous studies demonstrated that host-derived cells in tissue-engineered cartilage were mostly macrophages, 17 we added prelamin in the culture of RAW264, a murine macrophage-like cell. Regarding the expression of markers for inflammatory macrophages, the expression of Inos and Il1b in RAW264 was increased in accordance with the content of added prelamin up to a concentration of 40 ng/mL on day 1, although their expression had reversed to the original level by day 3 (Fig. 4A). It was suggested that prelamin released from dying chondrocytes could induce inflammatory properties in surrounding macrophages, impairing the maturation of tissue-engineered cartilage. We also added prelamin in the culture of chondrocytes in pellets to examine the effects of prelamin on chondrocytes. The expression of Col2 in chondrocytes decreased when prelamin was added at higher concentrations, while that of Col1 increased. It was suggested that higher concentrations of prelamin may be directly detrimental to the maturation of chondrocytes (Fig. 4B).
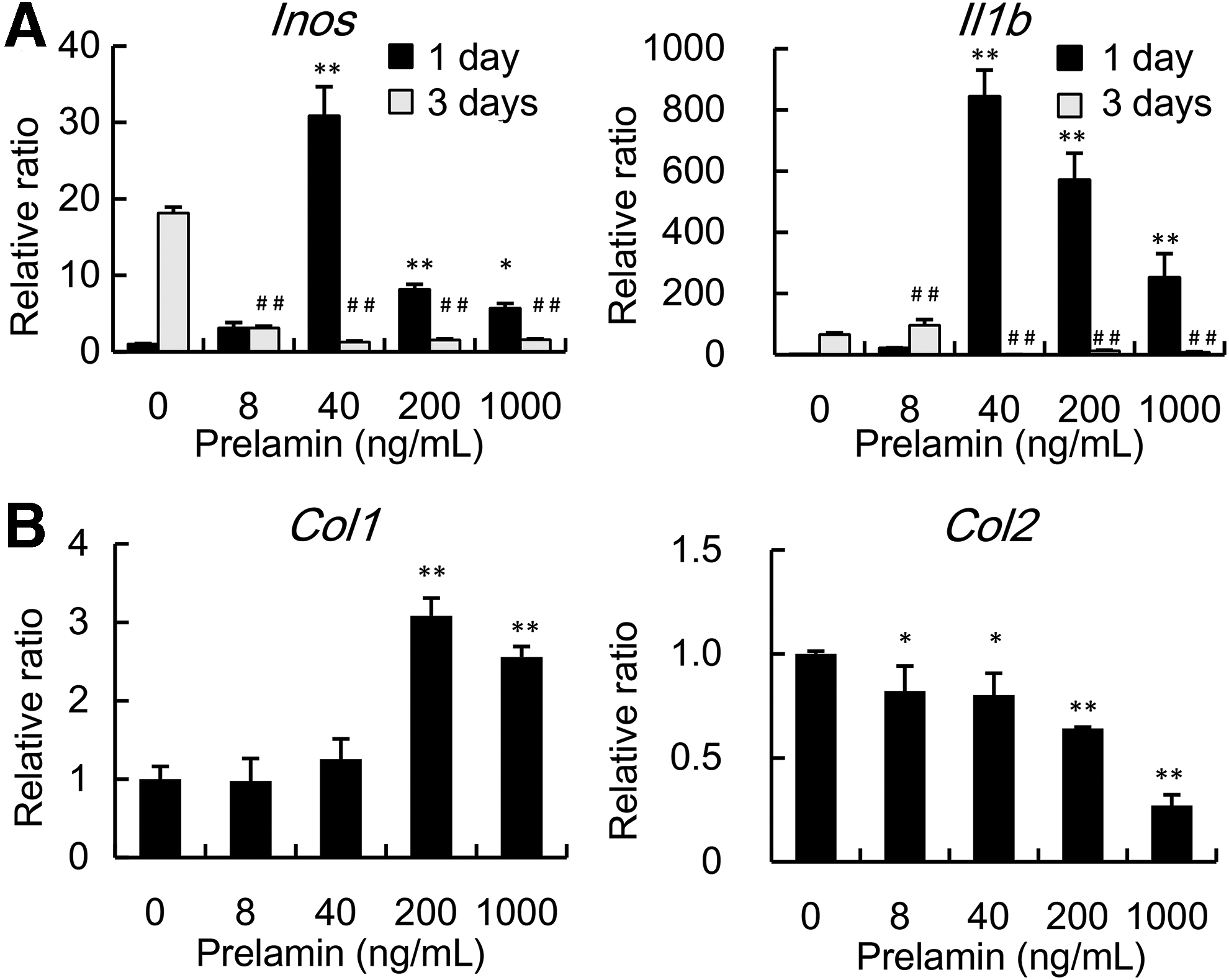
Effects of prelamin on macrophages.
Discussion
In this study, we examined the maturation and immunological response of tissue-engineered cartilage in mice when less viable chondrocytes were used for transplantation. To do so, we intentionally damaged chondrocytes with heat-intervention to prepare necrotic chondrocytes, some of which were gradually dying and releasing DAMPs. Constructs containing necrotic chondrocytes showed less maturation of tissue-engineered cartilage, with more localization of immune cells. Also, molecules released from necrotic chondrocytes were comprehensively analyzed, and prelamin was identified as a significant molecule.
Regarding the decreased maturation of tissue-engineered cartilage containing necrotic chondrocytes, we have considered the cause from several aspects. The heat damage of chondrocytes was not so severe as to induce immediate cell death; rather, it triggered gradual cell death after 24 h. Therefore, it is reasonable to think that some necrotic chondrocytes in tissue-engineered cartilage underwent cell death after transplantation, being unable to contribute to the formation of tissue-engineered cartilage. Meanwhile, the properties of chondrocytes are also altered during heat-intervention, which were evaluated by the expression of Col1 and Col2. In general, chondrocytes lose their ability to secrete cartilage matrix during monolayer culture and show increased expression of COL1 (dedifferentiation), while their secretory ability and expression of COL2 are restored once transplanted in vivo (redifferentiation). 28 Since chondrocytes after heat-treatment showed enhanced expression of Col1, their properties might have shifted to a more dedifferentiated state, resulting in decreased maturation of tissue-engineered cartilage. Another major reason for decreased maturation in constructs containing necrotic chondrocytes could be the immunological response caused by DAMP molecules. In our present study, macrophages and neutrophils were more localized in constructs containing necrotic chondrocytes at 2 weeks after transplantation, suggesting that necrotic chondrocytes in tissue-engineered cartilage released DAMP molecules and activated an immune response, affecting the outcome of tissue-engineered cartilage.
Many molecules have been reported as DAMPs, and they can vary depending on the type of cells and injured tissue. Regarding DAMPs from necrotic chondrocytes, we identified one as prelamin, whose release was 11-fold higher in necrotic chondrocytes. Lamins are a family of nuclear proteins that belong to the intermediate filaments29,30 and are primarily localized near the inner side of the nuclear membrane. There are two types of lamins; type A (lamin A and C) and type B (lamin B). Lamin A is induced from prelamin during a process of farnesylation, endoproteolysis, and methylation. Regarding the immunological function of prelamin, it increased the expression of inflammatory cytokines such as Inos and Il1b in RAW264 in this study. Indeed, another study demonstrated an increase of lamin A/C in human articular chondrocytes during lipopolysaccharide-induced inflammatory change. 31 Meanwhile, recent studies have implicated the involvement of lamins in aging-related inflammation. 32 In patients with Hutchinson-Gilford progeria syndrome (HGPS), an exceptionally rare disorder resembling premature aging, 33 accumulation of prelamin A isoforms induces NFκB activation, triggering the secretion of IL-6, CXCL1, and TNF-α. Chondrocytes heat-treated for 0.75 h failed to express lamin A/C in our present study (Fig. 3D). While chondrocytes heat-treated for 0.5 h showed a gradual decrease in viability, those treated for 0.75 h reached a state of cell death much faster by 1 day. Therefore, it can be construed that prelamin is released from cells gradually dying, but not from dead cells. Regarding the effects of prelamin, the expression of Inos and Il1b in RAW264 increased in accordance with the content of added prelamin up to a concentration of 40 ng/mL on day 1. Since the expression of these inflammatory mediators was rather suppressed at higher concentrations of prelamin, there must be an optimal condition for prelamin to increase inflammatory cytokines in RAW264. Also, the increased expression of Inos and Il1b in RAW264 by prelamin dropped by day 3 (Fig. 4A). Considering that prelamin is a part of the nuclear constituents, it dissolves rather easily and is not able to exert lasting effects on RAW264. Meanwhile, prelamin at higher concentrations decreased the expression of Col2 in chondrocytes, suggesting that prelamin from necrotic chondrocytes may be directly detrimental to the maturation of chondrocytes.
In this study, we have shown that maturation of tissue-engineered cartilage could worsen when less viable chondrocytes are included in the constructs. In these constructs, localization of macrophages and neutrophils was more apparent, suggesting that DAMPs from necrotic chondrocytes could activate an immunological response. Also, prelamin was identified as a significant molecule from necrotic chondrocytes. In general, cells used for regenerative medicine should meet criteria to secure clinical safety, and evaluation based on cell count and cell viability is the most fundamental and widely used method. However, there could be damage of cells, not critical enough to affect cell viability at the time-point of cell harvest, but eventually leading to cell necrosis. Therefore, it would be desirable to adopt several criteria to secure the quality of cells for regenerative medicine, and one option is determining the content of DAMPs in the culture medium. Our data suggest the possibility of lamins being useful for evaluating the condition of cultured chondrocytes.
Footnotes
Acknowledgments
We are grateful to Mr. Tomoaki Sakamoto for his laboratory assistance.
Disclosure Statement
No competing financial interest exists.
Funding Information
This work was supported by a Grant-in-Aid for Scientific Research (C) from the Ministry of Education, Culture, Sports, Science and Technology of Japan (No. 15K11236).