
Abstract
To investigate the effect of soluble factors released from human nasal chondrocytes (NCs) on cocultured human bone marrow mesenchymal stem cells (MSCs) and NC tissue-engineered constructs. Cartilage engineered from pure NCs on a three-dimensional (3D) porous collagen scaffold was cultured indirectly in a Transwell system with cartilage engineered from a direct coculture of human bone marrow-derived MSCs and NCs on a 3D porous collagen scaffold. The soluble factors were measured in the conditioned media from the different chambers of the Transwell system. Engineered cartilage from cocultures exposed to the pure NC construct exhibited reduced chondrogenic potential relative to control constructs, shown by reduced extracellular matrix deposition and increased expression of hypertrophic markers. Analysis of the soluble factors within the conditioned media showed an increase in inflammatory cytokines in the coculture chamber exposed to the pure NC construct. Principal component analysis revealed that the majority of the data variance could be explained by proinflammatory factors and hypertrophic chondrogenesis. In conclusion, our data suggest that inflammatory cytokines derived from NCs reduce the chondrogenic potential of coculture engineered cartilage through the induction of hypertrophic chondrogenesis.
Impact statement
The use of engineered cartilage from cocultured nasal chondrocytes (NCs) and mesenchymal stem cells for nasal cartilage reconstruction may be problematic. Our data suggest that the soluble factors from surrounding native NCs in the cartilage to be fixed can compromise the quality of the engineered cartilage if used in reconstructive surgery.
Introduction
Nasal chondrocytes (NCs) have been an attractive option for engineering cartilage tissue as they present an ectopic source of chondrocytes that can be obtained through minimally invasive conditions, which reduces complications associated with donor site morbidity.1–5
NCs can be easily isolated from the nasal septum and possess a robust capacity to redifferentiate and express a functional extracellular matrix (ECM) in chondrogenic culture.1,5–7 Supplementation of expansion culture medium with combined fibroblast growth factor-2 (FGF2) and transforming growth factor beta-1 (TGFβ-1) has been found to maintain the capacity of NCs to undergo chondrogenic redifferentiation and is commonly used in tissue engineering applications.4,6 In addition, compared to chondrocytes isolated from articular cartilage, NCs in culture have been shown to proliferate more rapidly and exhibit reduced donor-to-donor variability in chondrogenic capacity.5,6,8 NCs have been previously used for the repair of articular cartilage in the knee 2 and have been investigated for the repair of intervertebral discs9,10
Mesenchymal stem cells (MSC) have also emerged as a viable option for generating tissue-engineered cartilage, as they can be isolated from a variety of tissues.11–13 Bone marrow-derived MSCs (BM-MSCs) can proliferate extensively in culture and their inherent plasticity provides the potential to form cartilage under the appropriate culture conditions.14,15 However, the use of BM-MSCs in engineered cartilage is limited due to loss of chondrogenic capacity in later passages,16,17 as well as a tendency to undergo vascular calcification in ectopic transplantation in mice. 18
Coculture is a method of cell culture, whereby two distinct cell types are cultured in the same environment. In a coculture, the different cell types can interact through cell-cell contacts and/or through the release of soluble factors into the shared culture medium.19–21 Previous studies have demonstrated that the direct coculture of human NCs and BM-MSCs has a synergistic effect on chondrogenic capacity and sulfated glycosaminoglycan (GAG) production. 20 It is likely that the observed chondro-inductive effects observed were due to direct cell-to-cell contacts, as these effects were not observed when NCs and BM-MSCs were cultured in a Transwell model.20,21 In contrast, Wu et al., attributed the chondro-inductive effect observed in cocultures of articular chondrocytes and BM-MSCs to soluble trophic factors. 21 A Transwell model allows two distinct cell populations to be physically separated, but cultured in the same environment. This allows for the cells to communicate through soluble factors that either population may release into the culture media. These soluble factors can have significant effects on cellular function and particularly cell types with higher plasticity, such as MSC.22–25 Bovine chondrocytes have been shown to release signaling molecules that promote the chondrogenesis of undifferentiated cell types.26,27 MSCs isolated from goats cultured in chondrocyte-conditioned media exhibited a chondrocyte-like morphology upon expansion 26 and human embryonic stem cells cultured in a Transwell system with chondrocytes expressed type II collagen and showed increased GAG production, 28 which are hallmarks of chondrogenesis. Similarly, Ahmed et al. demonstrated that rat cartilage chips released soluble chondrogenic factors that induced chondrogenesis in rat bone marrow MSCs. 24 Thus, we hypothesized that soluble signaling factors released from pure NCs will further enhance the synergistic chondrogenesis observed in direct cocultures of NCs and MSCs, and suppress chondrocyte hypertrophy.
We employed a Transwell system to examine the effects of chondrocyte-derived soluble factors on the chondrogenic capacity of human NC-MSC coculture constructs. In contrast, to our hypothesis, our results showed that coculture constructs of NCs and MSCs after exposure to pure NC construct through the Transwell system exhibited reduced GAG matrix, aggrecan (ACAN) expression, lower Bern scores as a metric of engineered cartilage quality, and elevated expression of matrix metallopeptidase 13 (MMP13), which are collectively indicative of a reduced chondrogenic capacity with chondrocyte hypertrophy. To examine this further, we analyzed the relative levels of various cytokines within the conditioned media and found that specific soluble factors were elevated in the Transwell cultures relative to the cocultured control. Correlations in the data were probed by using a principal component analysis (PCA) to determine which soluble factor(s) may be responsible for the observed reduction in chondrogenic capacity of the NC-MSC coculture construct.
Methods
Cell harvest
Tissues were collected with the approval of and in accordance with the University of Alberta's Health Research Ethics Board. Nasal septal cartilage was collected from six donors (all male; mean age: 33.5 years; range: 23–43 years) at the Leduc Community Hospital. Five of the donors were hospitalized for a deviated nasal septum. One of the donors was hospitalized for chronic rhinosinusitis. Bone marrow aspirates were collected from the iliac crest of four different donors during routine orthopedic procedures (all male; mean age: 42 years; range: 28–58 years) at the University of Alberta Hospital. One bone marrow aspirate was collected from the right humerus during a humeral fracture repair.
Cell isolation and culture
Nasal cartilage was enzymatically digested in 0.15% (w/v) type II collagenase (300 U/mg; Worthington Biochemical Corporation) in Dulbecco's modified Eagle's medium (DMEM; Sigma) supplemented with heat-inactivated Fetal Bovine Serum (FBS; 5% v/v; Gibco), 100 U/mL penicillin and streptomycin with L-glutamine (2 mM; Thermo Fisher), and HEPES (0.1 M; Thermo Fisher). Samples were incubated at 37°C for 22 h in a shaking incubator. Tissue digests were passed through a 70-μm nylon strainer to remove undigested particulate tissue. NCs released from the digest were washed thrice in sterile phosphate-buffered saline (PBS; Sigma), resuspended in standard media, and plated in tissue culture flasks in a humidified incubator under normal atmospheric conditions (normoxic; 21% O2, 5% CO2). After 24 h, adherent cells were removed by trypsinization and the cell populations were counted using a hemocytometer and Trypan Blue. After isolation, NCs were plated at 104 cells/cm2 and expanded in DMEM complete (referring to supplementation with heat-inactivated FBS (10% v/v; Gibco), 100 U/mL penicillin and streptomycin with L-glutamine (2 mM; Thermo Fisher), and HEPES (0.1 M; Thermo Fisher), 1 ng/mL TGFβ-1, and 5 ng/mL FGF-2 at 37°C under normoxic conditions. Medium changes were performed twice a week. NCs were used for experiments at the end of passage one with an average cumulative population doubling of 3.32 (range: 2.72–4.05).
Bone marrow-mononucleated cells (BM-MNCs) were isolated from bone marrow aspirates using Histopague-1077 (Sigma) according to the manufacturer's instructions. In brief, donor bone marrow aspirate was diluted 1:1 in PBS. The diluted sample was layered over top of Histopaque-1077 at a 2:1 ratio (sample: Histopaque-1077). Samples were centrifuged at 1200 rpm for 30 min at 25°C. BM-MNCs were isolated from the interface of the plasma-Histopaque-1077 layers and diluted 1:3 in α minimum Eagle's medium (αMEM; Gibco) complete (i.e., supplementation with heat-inactivated FBS (10% v/v; Gibco), 100 U/mL penicillin and streptomycin with L-glutamine (2 mM; Thermo Fisher), and HEPES (0.1 M; Thermo Fisher). Diluted samples were centrifuged at 1500 rpm for 10 min at 25°C. The resulting supernatant was aspirated and cell pellets were resuspended in 2–5 mL of αMEM complete (volume dictated by the size of the pellet). Samples were again centrifuged at 1200 rpm for 5 min at 25°C. BM-MNC number was determined using crystal violet and a hemocytometer. BM-MNCs were then plated at 5000 cells/cm2 in αMEM complete with 5 ng/mL FGF-2 at 37°C under normal atmospheric conditions (normoxic; 21% O2 and 5% CO2). Medium changes were performed twice a week. BM-MNCs were also plated at 100 cells per 100 mm diameter petri dish in αMEM complete with 5 ng/mL FGF-2 at 37°C under normoxic conditions, for colony-forming unit- fibroblastic assay (Cfu-f) to determine the proportion of BM-MNCs at initial plating, accounting for the BM-MSC content of the bone marrow aspirates (data not included). BM-MSCs were used for experiments at the end of passage two with a mean cumulative population doubling of 14.12 (range: 12.55–16.15).
Transwell culture model
The Transwell culture system allows small molecules to freely diffuse from one culture chamber to another. After counting, NCs and BM-MSCs were resuspended in serum-free chondrogenic media (DMEM supplemented with 100 U/mL penicillin and streptomycin (Thermo Fisher), 2 mM L-glutamine (Thermo Fisher), 100 mM HEPES, insulin-transferrin-selenium (ITS +1), 0.1 μM dexamethasone, 0.1 mM ascorbic acid 2-phosphate, 0.1 mM L-proline, and 1 ng/mL TGFβ-1). Because of limited donor numbers, two of the BM-MSC donor samples were used twice, paired with a different NC donor. Six different donor pairs were set up for the coculture constructs. For each donor pair, six replicate constructs were analyzed, with two replicates for each type of analysis (GAG quantification, gene expression, and histology). Each construct was seeded with 5 × 105 cells, as per Bornes et al. 29 Cocultures consisting of 5 × 105 cells (1.25 × 105 NCs and 3.75 × 105 BM-MSCs for a ratio of 25:75 or 1:3) were seeded onto the porous side of type I and III collagen membrane scaffolds (Chondro-Gide; Geistlich Pharma) and cultured in the well chamber (WC), referred to as the WC construct. The chosen ratio reproducibly led to chondro-induction in coculture studies involving articular chondrocytes or meniscus fibrochondrocytes, or NCs with bone marrow MSCs.20,30,31 The membrane scaffolds measured 6 × 2 mm (diameter × thickness). The insert chamber (IC) contained 5 × 105 NCs seeded onto a type I collagen porous sponge scaffold (Integra Life Sciences), referred to as the IC construct. The sponge scaffolds measured 6 mm by 3.5 mm (diameter × thickness). As a control, cocultures (NCs:BM-MSCs at 1:3) were seeded on the membrane scaffolds and cultured in the well without the NC construct in the insert, referred to as the control chamber (CC) construct. Chambers consisted of six-well plates, with inserts within the WC, to form the IC. Each chamber, including the IC, contained 3 mL of chondrogenic media throughout the experiment. A schematic of the experimental setup is shown in Figure 1. Constructs were cultured in chondrogenic media for 3 weeks before analysis. The media were collected and changed twice per week.
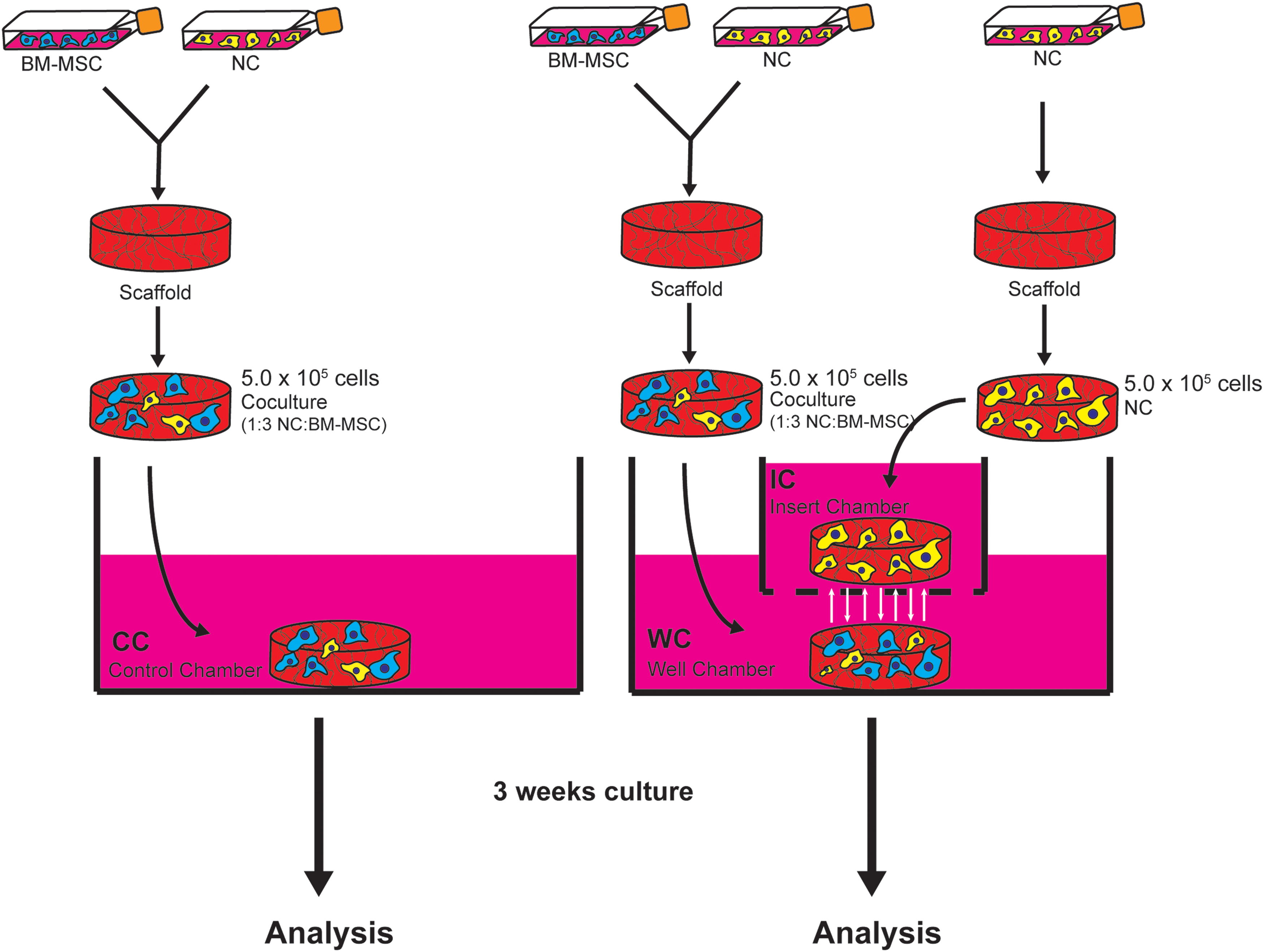
A schematic of the Transwell experimental setup. Shown is a detailed schematic of the experimental setup employed using the Transwell system. BM-MSCs were expanded in tissue culture flasks to passage two and NCs were expanded in tissue culture flasks to passage one. Cocultures of NCs:BM-MSCs (25:75) were seeded onto scaffolds and incubated in chondrogenic media in the Transwell apparatus alone (labeled “Control Chamber” or “CC”) or in the presence of an NC construct (labeled “Well Chamber” or “WC”). The NC construct contained NCs only seeded onto a scaffold and incubated in chondrogenic media in the insert, labeled “Insert Chamber” or “IC.” The constructs in the IC and the WC are unable to interact physically, but can communicate through soluble factors, which can pass through the permeable membrane between the two chambers. After 3 weeks of culture in chondrogenic media, constructs were analyzed. BM-MSC, bone marrow-derived mesenchymal stem cell; NCs, nasal chondrocytes. Color images are available online.
Sulfated glycosaminoglycan and DNA quantification
Coculture constructs from each condition were rinsed in PBS and frozen at −80°C. Constructs were thawed and digested overnight in 1 mg/mL proteinase K (Sigma) at 56°C. To quantify the GAG content, a 1,9, dimethylmethylene blue (DMMB; Sigma) assay was performed. Chondroitin sulfate (Sigma) was used as the standard. Colorimetric readings were measured at 530 nm using a V-max kinetic microplate reader (Molecular Devices). DNA content was measured from each sample using the CyQUANT Cell Proliferation Assay Kit (Thermo Fisher) according to the manufacturer's instruction. The supplied λ-bacteriophage DNA was used as a standard. Fluorescence emission was measured at 530 nm (excitation 450 nm) on a CytoFluor II fluorescence multiwell plate reader (PerSeptive Biosystems). The data were plotted in GraphPad (Prism 8) and statistical significance was determined by an unpaired Student's t-test using GraphPad (Prism 8).
Histology
Constructs from each chamber were fixed in 10% (v/v) formalin (Anachemia) overnight at 4°C. The constructs were then washed in PBS, processed, and embedded in paraffin wax. Embedded constructs were cut at 5-μm thickness using a Leica RM2125 RTS rotary microtome (Leica Biosystems). Sections were stained with 0.1% (w/v) Safranin O (Sigma) and counter-stained with 1.0% (w/v) Fast Green FCF (Sigma). Sections were mounted in Richard-Allan Scientific Mounting Medium (Thermo Fisher). The resulting stained sections were imaged on a Nikon Eclipse Ti-S microscope coupled to a DS-U3/Fi2 Color CCD camera using 10 × and 20 × objective lenses. Stained sections were also imaged using a 40 × lens and a Zeiss Primovert microscope coupled to a Canon Rebel T6i camera. Bern scores were assigned for each construct, as previously described. 32 All Bern scoring was performed blinded by five trained observers. The scores shown are an average of the scores assigned. Raw Bern scores for each construct were plotted using GraphPad (Prism 8).
Immunofluorescence
Constructs were fixed, embedded, and sectioned, as described above. Sections were deparaffinized as described above and progressively rehydrated in ethanol (100%, 95%, 70%, and 35%). Following complete rehydration in water, antigen retrieval was performed with 1 mg/mL protease XXV (Thermo Fisher) for 30 min at room temperature. Sections were washed in PBS and treated with 1 mg/mL hyaluronidase (Sigma) for 30 min at 37°C. Sections were blocked in 5% bovine serum albumin (BSA; Cell Signaling Technology) for 30 min at room temperature. Type X collagen (rabbit) (ab58632; Abcam) primary antibody (1:100 dilution in 1% BSA) was incubated with sections overnight at 4°C. Sections were washed thoroughly in PBS and incubated for 30 min at room temperature with goat anti-rabbit IgG Alexa Fluor 594 (ab150080; Thermo Fisher) secondary antibody (dilution 1:200 in 1% BSA). Sections were washed thoroughly in PBS and incubated in 4′,6-diamidino-2-phenylindole (DAPI) for 5 min at room temperature. Sections were mounted with 1:1 Glycerol:PBS and imaged on a Nikon Eclipse Ti-S microscope coupled to a DS-U3/Fi2 Color CCD camera using 10 × and 20 × objective lenses.
Quantitative real-time polymerase chain reaction
Coculture constructs from each condition were placed in tubes and immersed in TRIzol reagent (Thermo Fisher) and frozen at −80°C. Samples were later thawed to room temperature and cells were lysed by grinding scaffolds with a pestle. Total RNA was isolated by chloroform extraction. Complementary DNA (cDNA) was synthesized from 100 ng of total RNA using the GoScript Reverse Transcriptase kit (ProMega) and 1 μg of oligo(dT) primers. Quantitative PCR was performed using Takyon DNA Polymerase and SYBR Green detection (Eurogentec) on a CFX Connect Real-Time PCR Detection System (Bio-Rad Laboratories). Primer sequences for ACAN, COL1A2, COL2A1, COL10A1, MMP13, IHH, CNMD, PTHLH, SOX9, and YWHAZ were designed using Primer Express 3.0.1 (Thermo Fisher). Primer sequences for ACAN, COL2A1, COL10A1, SOX9, and YWHAZ were previously validated in Andrews et al. 5 Primer sequences for HMBS and B2M were used from and previously validated in Vandesompele, et al. 33 Primer sequences are shown in Table 1. Transcript levels for ACAN, COL1A2, COL2A1, COL10A1, SOX9, MMP13, IHH, CNMD, and PTHLH were normalized to the housekeeping genes B2M, HMBS, and YWHAZ using the delta CT method (2−ΔΔCT).34,35 Fold change (FC) and the log2FC between samples were calculated using Microsoft Excel. The data were plotted in GraphPad (Prism 8) and statistical significance for each gene's relative expression was determined by an unpaired Student's t-test using GraphPad (Prism 8).
Primer Sequences
Cytokine array
Conditioned media were collected from the control, well, and IC for each construct from five separate experiments (five different donor sets). The cytokine levels were measured from the conditioned media that were obtained from the final medium change and analyzed using the RayBio C-Series Human Cytokine Antibody Array C5 kit (RayBiotech), according to the manufacturer's instructions. In brief, array membranes were incubated overnight at 4°C in conditioned media. The membranes were then washed and incubated in a biotinylated antibody cocktail, followed by a labeled streptavidin solution. Chemiluminescence was visualized on a FluorChem Q Imaging System (Alpha Innotech Corporation) coupled to an HD2 CCD camera, using FluorChem Q software. Membranes were exposed for 45 s. Relative cytokine expression was determined by densitometry of the imaged membranes and normalized to the positive controls. The raw data were analyzed by a Microsoft Excel-based Analysis Software provided by the manufacturer (RayBiotech). FC and the log2FC between samples were calculated using Microsoft Excel. The data were plotted in GraphPad (Prism 8) and statistical significance was determined by Dunnett's test using GraphPad (Prism 8).
Data and statistical analysis
We used PCA to summarize all quantitative data. Individual measurements that fell below a Kaiser-Meyer-Olkin (KMO; measures for sampling adequacy) of 0.5 were excluded. We used SPSS version 25 to perform PCA. Prism 8 was also used for statistical analysis as indicated in other assays.
Results
Nasal chondrocyte-derived soluble factors have a negative effect on coculture matrix formation
To analyze the effect of NC-derived soluble factors on the chondrogenic potential of tissue-engineered constructs, matrix formation was determined by measuring the GAG content of each construct after 3 weeks of culture in chondrogenic media. Interestingly, total GAG content was significantly reduced (p < 0.05) in WC constructs relative to CC constructs (Fig. 2A). In addition, no significant difference in DNA levels was observed between WC constructs and CC constructs (Fig. 2B). GAG was also measured relative to the DNA content within each construct and displayed as GAG/DNA. It was also found that WC constructs possessed significantly lower (p < 0.01) GAG/DNA than CC constructs (Fig. 2C). GAG/DNA from CC constructs measured 38.61 ± 11.34 (μg/μg), whereas WC constructs measured 24.78 ± 7.66 GAG/DNA (Fig. 2C). IC constructs measured 29.49 ± 6.05 GAG/DNA (Fig. 2C). This suggests that the presence of NCs within the Transwell insert has a negative effect on GAG production and matrix formation in the coculture construct within the WC.

Coculture constructs exposed to NCs within the IC of the Transwell system exhibit reduced chondrogenic potential. Constructs from each chamber were digested in proteinase K overnight. DNA and GAG from each were measured from each construct.
Matrix formation was also analyzed by histology, using Safranin-O staining. 36 Safranin-O staining revealed decreased GAG deposition in sections from WC constructs relative to the CC constructs. This was evident from reduced Safranin-O staining intensity and uniformity throughout the entire construct (Fig. 2D). In addition, higher magnification images (20 × and 40 × ) reveal decreased spacing between cells and more spindle-shaped cells in the WC constructs compared to the CC constructs (Fig. 2D and Supplementary Fig. S1, respectively), both of which are indicative of poorer ECM formation. 32 To quantify these features, the Bern scoring method was employed. The Bern score provides a visual evaluation system for Safranin-O staining assessment of tissue-engineered cartilage constructs. 32 Each construct was scored blinded by five trained independent ob. Overall, Bern scores were reduced for WC constructs relative to CC constructs. In each respective donor set, WC constructs were consistently scored lower than those from the control. CC constructs were assigned an average Bern score of 7.56 ± 0.39, whereas WC sections scored 6.68 ± 0.91 (Fig. 2E).
NC-derived soluble factors affect the expression of ACAN and MMP13 in the well chamber construct
Gene expression analysis showed differential expression of various chondrogenic markers among the different constructs. ACAN was significantly reduced (p < 0.01) in cells from the WC construct relative to CC construct cells showing a 1.33 log fold reduction in expression (Fig. 3A). In addition, the expression of MMP13 showed a 1.38 log fold increase in expression (p < 0.01) in WC construct cells relative to the control (Fig. 3A). Similarly, the expression of Indian hedgehog (IHH) was reduced, although not significantly, in cells from the WC construct relative to the control (Fig. 3A). Type X collagen (COL10A1) was found to be downregulated 0.70 log fold in WC cells relative to the control (Fig. 3A), although not to a significant extent. However, when constructs were analyzed for type X collagen by immunofluorescence, it appeared that WC constructs exhibited stronger type X collagen immunostaining than CC constructs (Fig. 4). In contrast, no considerable differences were observed in type X collagen immunostaining between IC constructs and CC constructs. The expression of type I collagen (COL1A2) and type II collagen (COL2A1), parathyroid hormone-like hormone (PTHLH), chondromodulin (CNMD), and the transcription factor SOX9 was unaffected in WC construct cells relative to CC construct cells (Fig. 3A).
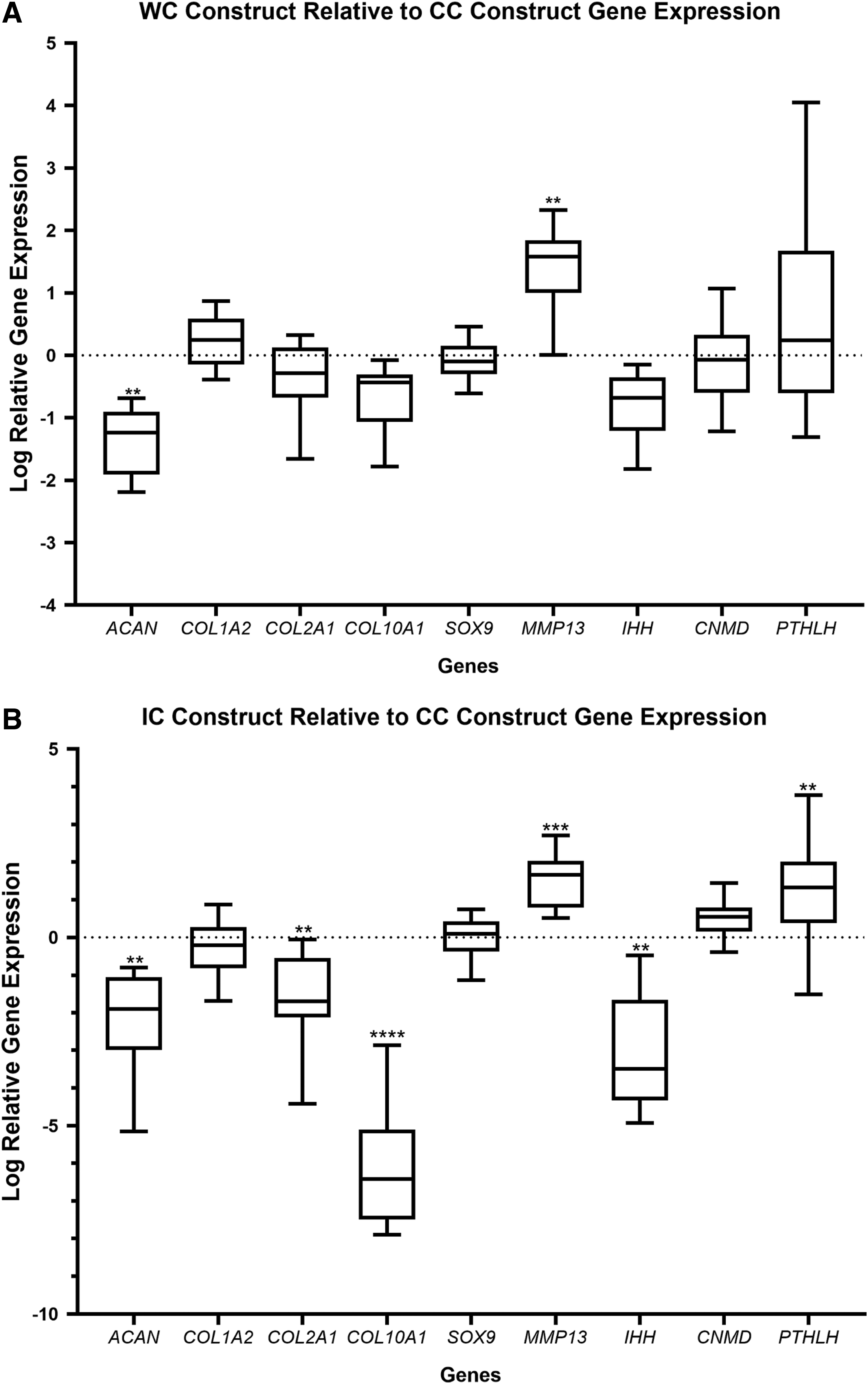
Chondrogenic markers are differentially expressed in coculture constructs exposed to NCs within the insert of the Transwell system. Total RNA was extracted from each construct and cDNA was synthesized. qPCR was employed to measure the relative transcript levels of ACAN, COL1A2, COL2A1, COL10A1, SOX9, MMP13, IHH, CNMD, and PTHLH in each construct. Genes were normalized to B2M, HMBS, and YWHAZ using the delta CT method (2−ΔΔCT).
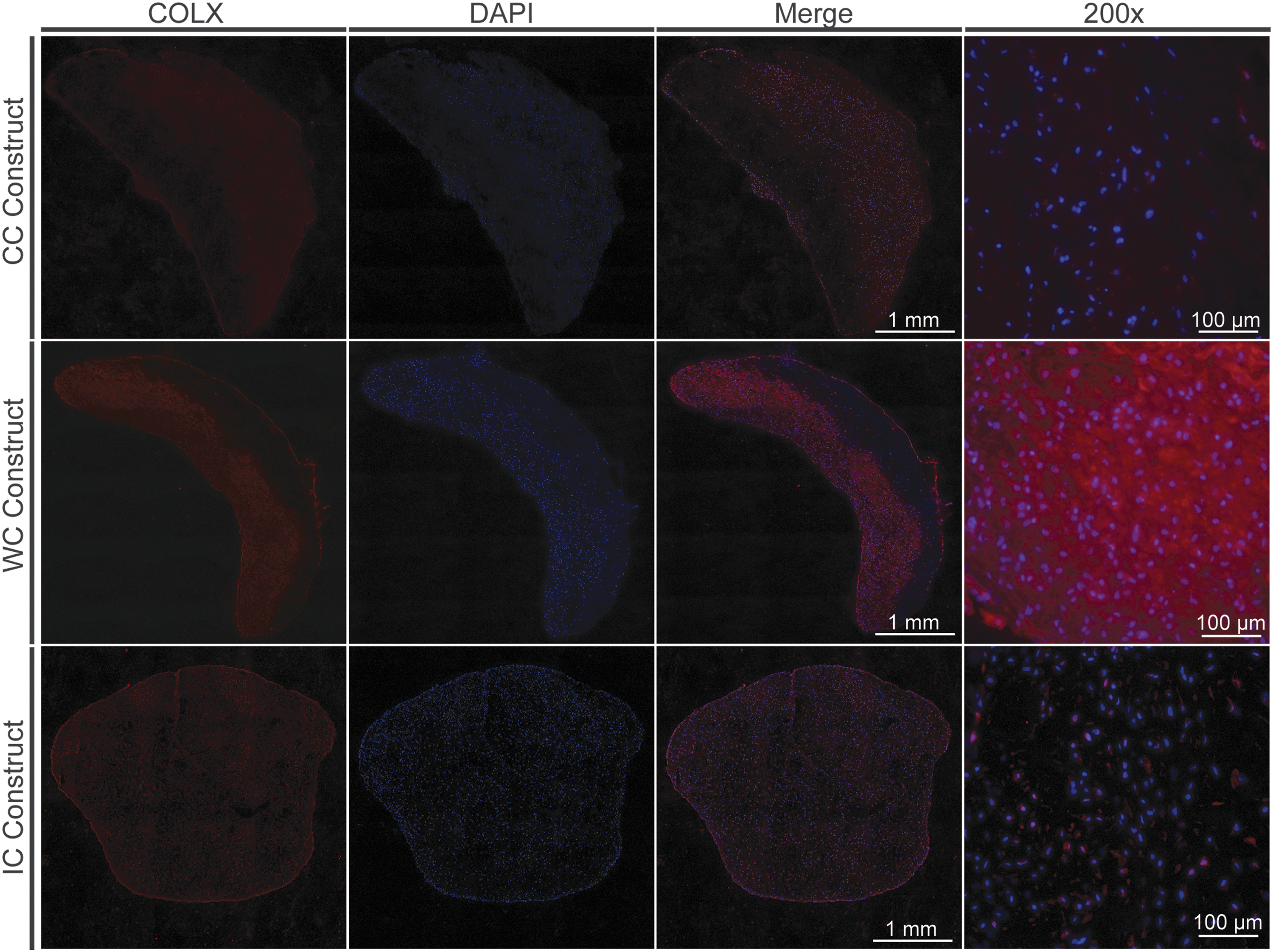
COLX immunostaining is more pronounced in WC constructs. Constructs were fixed in 10% formalin, sectioned, and probed with type X collagen (COLX) antibody and DAPI. Sections were imaged by fluorescent microscopy. The construct, CC, WC, or IC is indicated to the left of each image row. COLX is shown in red in the first column, DAPI is shown in blue in the second column, and the merged images are shown in the third column at a 100 × optical zoom. A 200 × optical zoom image of each section is shown in the far-right column. The scale bar for 100 × images represents 1 mm. The scale bar for the 200 × images represents 100 μm. Color images are available online.
Gene expression analysis of the IC construct also showed significant differential expression of several genes, relative to the control. Similar to the WC construct, ACAN mRNA expression was significantly reduced 2.08 log fold (p < 0.01) (Fig. 3B). In addition, COL2A1 and COL10A1 showed a 1.64 (p < 0.01) and 6.12 (p < 0.0001) log fold reduction in expression, respectively, in the IC constructs relative to the CC construct (Fig. 3B). MMP13 showed 1.54 log fold elevated expression (p < 0.001) and IHH showed a 3.1 log fold reduction in expression (p < 0.01) relative to the CC construct (Fig. 3B). The expression of PTHLH was elevated 1.16 log fold (p < 0.01) relative to the CC construct (Fig. 3B).
Osteoprotegerin, B lymphocyte chemoattractant, and insulin-like growth factor binding protein-1 levels were elevated in the WC-conditioned media
To determine which soluble factors may be influencing the chondrogenic capacity of the cells in the WC construct, a cytokine array was performed on conditioned media from each of the chambers. The cytokine levels were measured from the conditioned media that were obtained from the final medium change. Imaged arrays are shown in Supplementary Figure S2. Conditioned media from the WC were measured relative to the CC and each cytokine measurement was expressed as the log relative expression, displayed in Figure 5. Of the 79 cytokines measured, most were present at higher levels in the conditioned media of the WC relative to the CC. However, only three showed significantly differential expression: osteoprotegerin (OPG), B lymphocyte chemoattractant (BLC), and insulin-like growth factor binding protein-1 (IGFBP-1). OPG showed a 1.05 ± 0.53 log fold increase in expression (p < 0.01) in the WC relative to the CC (Fig. 5). BLC and IGFBP-1 showed a 1.09 ± 1.15 (p < 0.01) and a 0.85 ± 0.83 (p < 0.05) log fold increase in expression, respectively, in the WC relative to the CC (Fig. 5). OPG was also found at elevated levels in the IC-conditioned media (Supplementary Fig. S3).
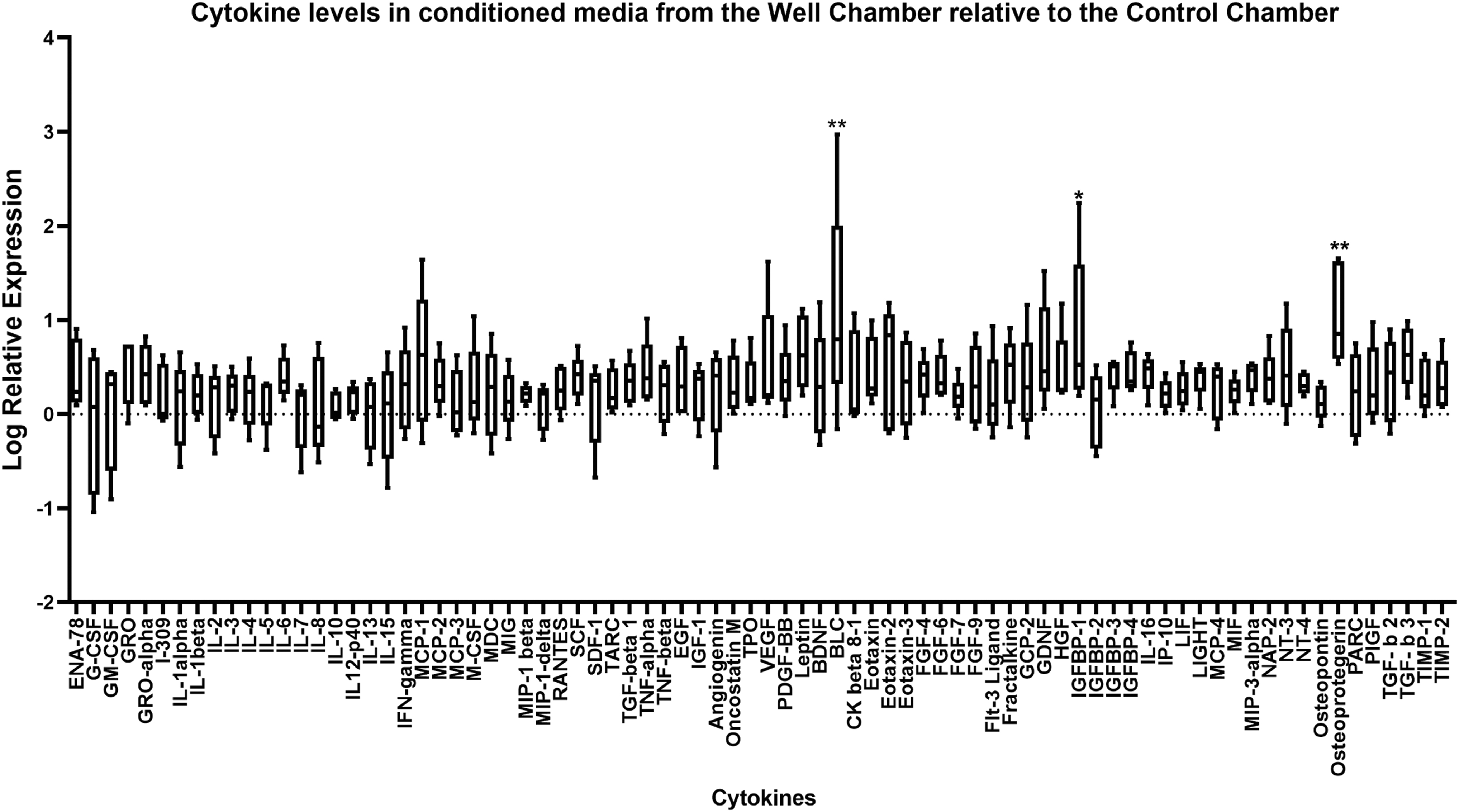
OPG, BLC, and IGFBP-1 are elevated in the conditioned media of the WC relative to the CC. The conditioned media from each chamber were analyzed using the Ray-Bio C-Series Human Cytokine Antibody Array C5 kit. Array membranes were incubated in conditioned media from each respective chamber overnight, followed by incubation in a biotinylated antibody cocktail, and labeled streptavidin solution. The membrane arrays were visualized by chemiluminescence and cytokine levels were determined by densitometry. The relative expression of cytokines in the conditioned media from the WC relative to the CC was determined and expressed as the log relative expression, shown in the graph. Each cytokine analyzed is shown on the x-axis and the log relative expression is shown on the y-axis. Statistical analysis was performed using a Dunnett's test. *p < 0.05; **p < 0.01. The whiskers represent the minimum and maximum, and the box extends from the 25th to the 75th percentile with the center line denoting the median. Conditioned media from five different donors were used in the cytokine array. OPG, osteoprotegerin; BLC, B lymphocyte chemoattractant; IGFBP-1, insulin-like growth factor binding protein-1.
Principal component analysis revealed a proinflammatory and hypertrophic chondrogenesis pathway
PCA was used to summarize pooled quantitative data and determine relationships between the measured variables. On the initial PCA, a six-component solution resulted in each component having eigenvalues exceeding 1. However, four of the six components accounted for less than 4% of the variation in the pooled data, and two (PC1 and PC2) of the six elements, collectively, accounted for ∼88% of the variation in the data. PC1 and PC2, respectively, described ∼81% and ∼7% of the total variance with respective eigenvalues of 73.4 and 6.3 in GAG/DNA, DNA, CNMD, PTHLH, COL1A2, COL2A1, ACAN, COL2A1, SOX9, COL10A1, IHH, MMP-13, and the proteins represented in the RayBio C-Series Human Cytokine Antibody Array C5 kit (Fig. 6–Varimax rotated plot of PCA). GAG content was poorly represented on both PC1 and PC2; therefore, this measured variable was removed from the two-component analysis. PC1 correlated most strongly with MMP-13 and proinflammatory cytokines (e.g., IL-1α, IL-1β, IL-2, IL-3, TNFα, and TNFβ), and PC2 connected most strongly with COL2A1, IHH, ACAN, CNMD, COL10A1, and PTHLH (Table 2–PCA correlation matrix). To that end, PC1 components were deemed to encourage a proinflammatory phenotype, and PC2 primarily described hypertrophic chondrogenesis that is perhaps orchestrated by the IHH signaling pathway. PC2 was therefore interpreted to support a hypertrophic phenotype.
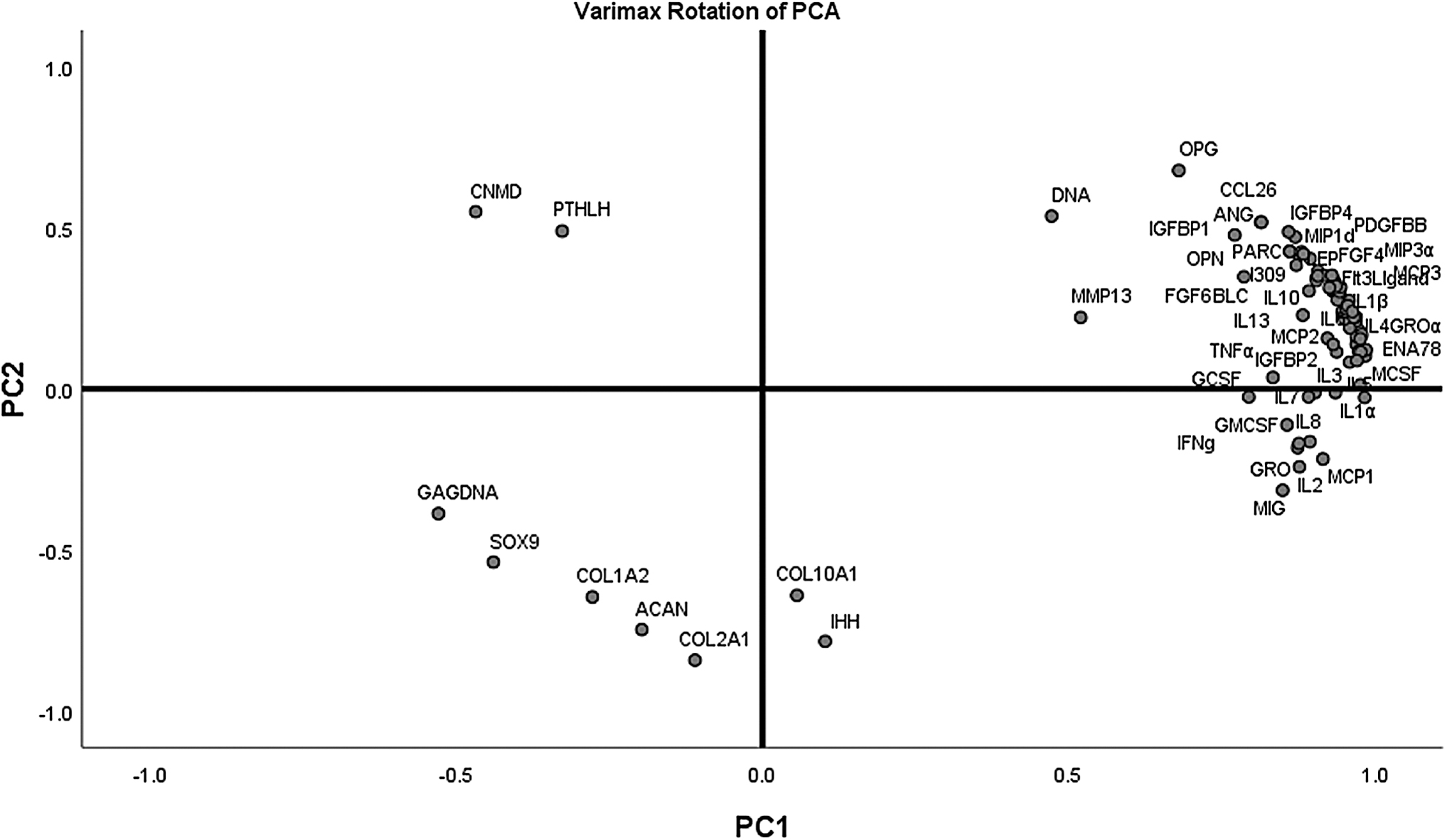
A varimax rotated plot of a two-component solution of principal component analysis of measured variables. Measured variables are as follows: GAG/DNA, DNA, CNMD, PTHLH, COL1A2, COL2A1, ACAN, COL2A1, SOX9, COL10A1, IHH, and MMP-13 and cytokines in Ray-Bio C-Series Human Cytokine Antibody Array C5 kit. Principal component 1 is on the x-axis and principal component 2 is on the y-axis. The position of each variable on the plot indicates the degree to which it loads on the two different principal components.
Principal Component Analysis Correlation Matrix
Extraction Method: PCA.
Rotation Method: Varimax with Kaiser Normalization.
PCA, principal component analysis.
Discussion
In contrast to our study's hypothesis, we observed a reduction in the quality of ECM in WC constructs relative to control constructs. A reduction in GAG/DNA in WC constructs was consistent with lower Bern scores in WC constructs, which suggests that the presence of NCs within the IC has a negative effect on chondrogenic capacity and matrix formation of the coculture within the WC. Reduced ACAN expression was also consistent with the observed reduction in ECM production in WC constructs, as ACAN is the major proteoglycan in the ECM of cartilage tissue. 37 Because of the nature of the Transwell setup, the cells within the IC could only influence cells within the WC through soluble factors. Therefore, it is likely that the soluble factors released into the media are responsible for the change in ECM quality observed. Because the media collected from the WC were analyzed relative to the CC and were found to exhibit increased levels of particular cytokines, it is likely that said cytokines are released from the NCs within the IC. It is also possible that the cytokines were released from the coculture construct within the WC in response to soluble factors from the NC construct within the IC.
One consideration to make is the conditions in which the constructs were cultured in. In this study, all constructs were cultured in normoxic conditions. Higher oxygen levels have been shown to induce hypertrophy and osteogenic differentiation in chondrocytes.38–40 It is possible that the culture of these constructs in hypoxic conditions might mitigate some of the hypertrophic effects observed. However, WC constructs were analyzed relative to the CC constructs, of which both were cultured in normoxia. Therefore, we reason that the hypertrophic effects observed were a result of the soluble factors released from the IC construct, as discussed.
Gene expression analysis revealed a significant upregulation of MMP-13 in WC constructs, a hypertrophic marker in chondrocytes. 41 Again, this is consistent with reduced chondrogenic quality, as MMP-13 is an endopeptidase that cleaves collagens and degrades ECM components. 42 Type X collagen (COLX) is also a marker of hypertrophic differentiation in chondrocytes.43,44 COLX immunostaining appeared more pronounced in WC constructs. However, there appeared to be a disconnect with the gene expression, which showed no significant change in COL10A1 expression. It is unclear as to why this is the case; however, it is possible that the COLX protein was stabilized and exhibited reduced turnover in the WC constructs. While the central dogma of biology dictates that mRNA levels are directly correlated with protein levels, which is not always the case, the discrepancy between RNA and protein levels has been previously observed. One study that analyzed RNA and protein levels of 1066 gene products in 23 different human cell lines found that only one-third of the gene products analyzed found significant correlations between RNA and protein levels. 45 In addition, this particular discrepancy has been previously observed in chondrocytes.46–48 In combination with the observed decrease in ECM quality, this suggests hypertrophic differentiation of the WC constructs. Furthermore, upon PCA, it was revealed that 7% of the data variance could be explained by chondrogenic hypertrophy (Fig. 5). Therefore, the data suggest that the release of soluble factors from the NCs within the IC induces hypertrophic differentiation of the coculture within the WC.
A general increase in inflammatory cytokines was detected in the conditioned media of the WC relative to the CC (Fig. 4) and PCA's PC1 components were identified as proinflammatory (Fig. 5), which explained ∼80% of the data variance. The elevation of OPG as recorded in our study supports the existence of a proinflammatory microenvironment in the conditioned media. OPG has been reported to be secreted by human articular chondrocytes in response to stimulation with interleukin-1β, a known proinflammatory cytokine expressed in osteoarthritic joints.49,50 However, it is interesting to note that OPG was reported to decrease in conditioned media from 2 weeks of cocultured pellets of primary human articular chondrocytes and BM-MSCs, although in the absence of soluble factors from pure primary human articular chondrocytes as in the case of pure NCs, as in this study. 20 Therefore, it is possible that the cytokines released from the NCs within the IC initiated an inflammatory response from the WC coculture, causing hypertrophic chondrogenesis. Cytokine upregulation has been previously linked to hypertrophic differentiation and the progression of OA in cartilage tissue. BLC, also known as CXCL13, is an inflammatory cytokine that has been linked to forms of arthritis.51,52 Interestingly, BLC was also reported to be upregulated in the conditioned media of pellets of cocultures of human articular chondrocytes and BM-MSCs after 2 weeks of in vitro culture. 20 In addition, IGFBP-1 has been suggested to reduce proteoglycan synthesis in cartilage because of its ability to sequester and block the activity of insulin-like growth factor-1 (IGF-1), a potent anabolic of cartilage ECM. 53 Both BLC and IGFBP-1 were found to be significantly upregulated in the WC chamber relative to the CC chamber. Increased cytokine expression results in a disruption of the balance between anabolism and catabolism in cartilage tissue, which contributes to OA progression. 54 It has been shown that antagonists to cytokine receptors for TNFα and IL-1 resulted in decreased expression of MMP-13 in chondrocytes from OA patients. 55 In addition, the treatment of infrapatellar fat pad-derived MSCs with conditioned media from OA cartilage fragments, which showed upregulated cytokine expression, resulted in increased hypertrophic differentiation in the MSC. 56 Inflammatory cytokines have been previously implicated in the degradation of ECM within cartilage tissue.57–62 BLC, also known as CXCL13, was significantly elevated in the WC-conditioned media and has also been implicated in early progression of RA. 52 In addition, IGFBP-1, which was significantly elevated in the WC-conditioned media (Fig. 4), has been linked to the progression of RA, caused by low bioavailability of IGF-1.63,64
The Transwell setup described presents a potential model for studying the effects of inflammatory cytokines and chronic inflammation on the degradation of cartilage tissue. Overall, the observed reduction in ECM quality in WC constructs can be explained by a general increase in inflammatory cytokines in the conditioned media. Cartilage erosion in RA is characterized by chronic inflammation 65 and has been linked to some of the cytokines found to be elevated in the WC-conditioned media. Furthermore, our data raise a reasonable concern in the use of engineered nasal cartilage from cocultured NCs and MSCs, to reconstruct nasal cartilage defects. It is probable that the soluble factors released by a surrounding native nasal cartilage may negatively impact the engraftment and quality of the engineered nasal cartilage from cocultures of NCs and MSCs after reconstructive implantation surgery of focal cartilage defects. These factors must be taken into consideration for improving the quality of tissue-engineered constructs for surgical implantation.
Footnotes
Acknowledgments
We thank Integra LifeSciences, Inc., for in-kind donation of type I collagen porous scaffold, Duragen (Integra LifeSciences, Inc., NJ). We thank Geistlich for in-kind donation of the hybrid type I and III collagen membrane scaffold, ChondroGide (Geistlich Pharma AG, Switzerland).
Disclosure Statement
No competing financial interests exist.
Authors' Contributions
M.A.B. and M.K. conducted the experiments and were responsible for data acquisition and analysis. M.A.B. drafted the initial article, and A.M.S. was involved in cell culture. M.O., K.A., and H.S. were involved in nasal cartilage procurement, article writing, and review. A.B.A. conceived the study, supervised the study, performed data analysis, including PCA, and was responsible for writing and final review of the article. All authors read and approved the final article.
Funding Information
Financial support was provided by Canadian Institutes for Health Research (CIHR PS 159661) to A.B.A., M.O., K.A., and H.S., Alberta Cancer Foundation-Mickelborough Interfacial Biosciences Research Program (ACF-MIBRP 27128) to A.B.A., M.O., and K.A., Canadian Foundation for Innovation (CFI 33786) to A.B.A., and University Hospital Foundation (RES0028185) to A.B.A.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.