
Abstract
Irregular defects at sites of degenerative cartilage often accompany osteoarthritis (OA). The development of novel cell-/biomaterial-based cartilage tissue engineering methods to address these defects may provide a durable approach to hinder the development of OA. In this study, we fabricated a neocartilage patch by fusing cell aggregates onto a biodegradable nanofiber film for degenerative cartilage repair. Human mesenchymal stem/stromal cell (MSC) aggregates were prepared and induced for chondrogenesis in a thermosensitive hydrogel, poly (N-isopropylacrylamide-co-acrylic acid (p(NIPAAm-AA)). Cell migration mediated the formation of cell aggregates in the thermosensitive hydrogel and led to a cell-dense hollow shell structure. The chondrocytes derived from MSC aggregates in the hydrogel were evidenced by the expression of chondrogenesis-related genes and extracellular matrices. They were fused onto an electrospun film by mechanical force and spatial confinement to generate a neo-cartilage patch. The fabricated neocartilage patches may be able to integrate into the irregular defects under compressive stresses and achieve cartilage regeneration in vivo.
Impact statement
The formation of human mesenchymal stem/stromal cells aggregates in thermosensitive hydrogels was mechanistically examined. These in situ formed cell aggregates with enhanced chondrogenesis were bioengineered into a neocartilage patch for regeneration of superficial irregular cartilage defects.
Introduction
Osteoarthritis is a chronic joint disease that is always accompanied with irregular defects on the articular hyaline cartilage surface. These defects are often difficult to heal owing to the avascular nature of the tissue and the limited number of chondrogenic progenitor cells in the defect area. 1 Tissue engineering, utilizing biomaterials and/or cells, represents a potential approach to promote cartilage regeneration.2–5 However, due to the irregular structure of the damaged cartilage, it is challenging to retain the implanted biomaterials and/or cells in situ. Although injectable hydrogels can deliver cells in vivo in a minimal invasive manner to fill deep irregular defects, their poor mechanical property becomes a key constraint for regeneration of a superficial cartilage defect.6,7
Stem cell aggregates are superior to individual cells, due to their functional enhancement in secretion of endogenous growth factors and deposition of extracellular matrices (ECMs).8–10 The cell aggregates are generated through spontaneous cell aggregation-mediated fabrication 11 ; substrate-mediated fabrication 12 ; or technology-assisted fabrication. 13 While hydrogels are widely used for three-dimensional stem cell culture, few have been used to stimulate the formation of cell aggregates in situ for specific applications, and the intrinsic mechanism of cell aggregate formation within hydrogels has not been reported.14–21 We have previously reported a thermosensitive hydrogel, poly (N-isopropylacrylamide-co-acrylic acid (p(NIPAAm-AA)), which could facilitate multilineage differentiation of human mesenchymal stem/stromal cells (MSCs) in vitro and osteochondral regeneration in vivo by the formation of cell aggregates in situ.22,23 In this study, we further examined the mechanism for the formation of cell aggregates within the p(NIPAAm-AA). The differentiated cells were harvested and transferred onto an electrospun film and fused into an integrated neocartilage by spatial confinement and external mechanical compression as shown in Figure 1. The fabricated neocartilage patch could be used for cartilage regeneration by simply attaching it to the degenerated cartilage with the assistance of a medical glue24,25 or suture, 26 and the cells within the neocartilage may be able to integrate into the irregular defects by remodeling the surrounding ECM proteins.
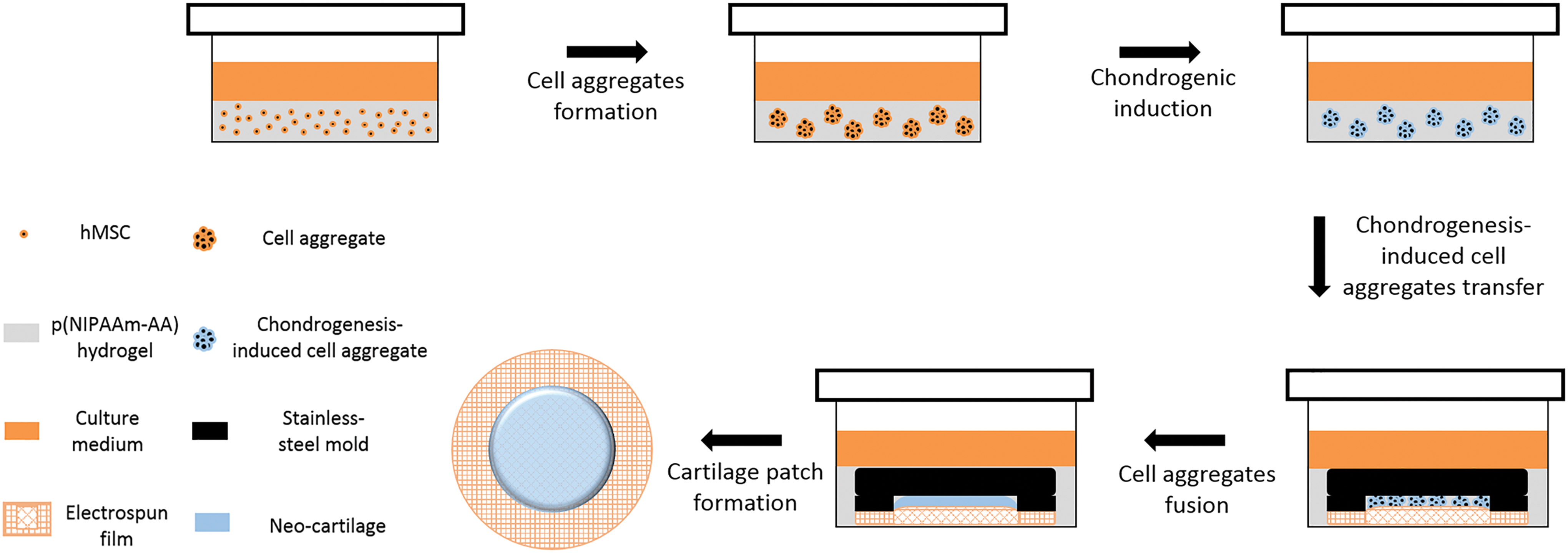
The procedure of in vitro cartilage patch fabrication. Color images are available online.
Materials and Methods
Details of the materials and methods are described in Supplementary Data.
Results
Characterization of p(NIPAAm-AA) hydrogel and electrospun film
Fourier-transform infrared spectroscopy (Supplementary Fig. S1), thermosensitive behavior (Supplementary Fig. S2) and toxicity (Supplementary Fig. S3) of p(NIPAAm-AA) hydrogels, and characterization of electrospun film (Supplementary Fig. S4) are presented and discussed in Supplementary Data.
MSC aggregates in p(NIPAAm-AA) hydrogel
When UE7T-13 cells were cultured inside the p(NIPAAm-AA) hydrogel at a MgCl2 concentration of 0.01 M on days 1 and 9, the majority of cells displayed a green color, indicating excellent cell viability and the biocompatibility of the p(NIPAAm-AA) hydrogel at a MgCl2 concentration of 0.01 M (Fig. 2). Interestingly, individual cells had a round shape on day 1, while cellular aggregates were presented in the p(NIPAAm-AA) hydrogel on day 9.
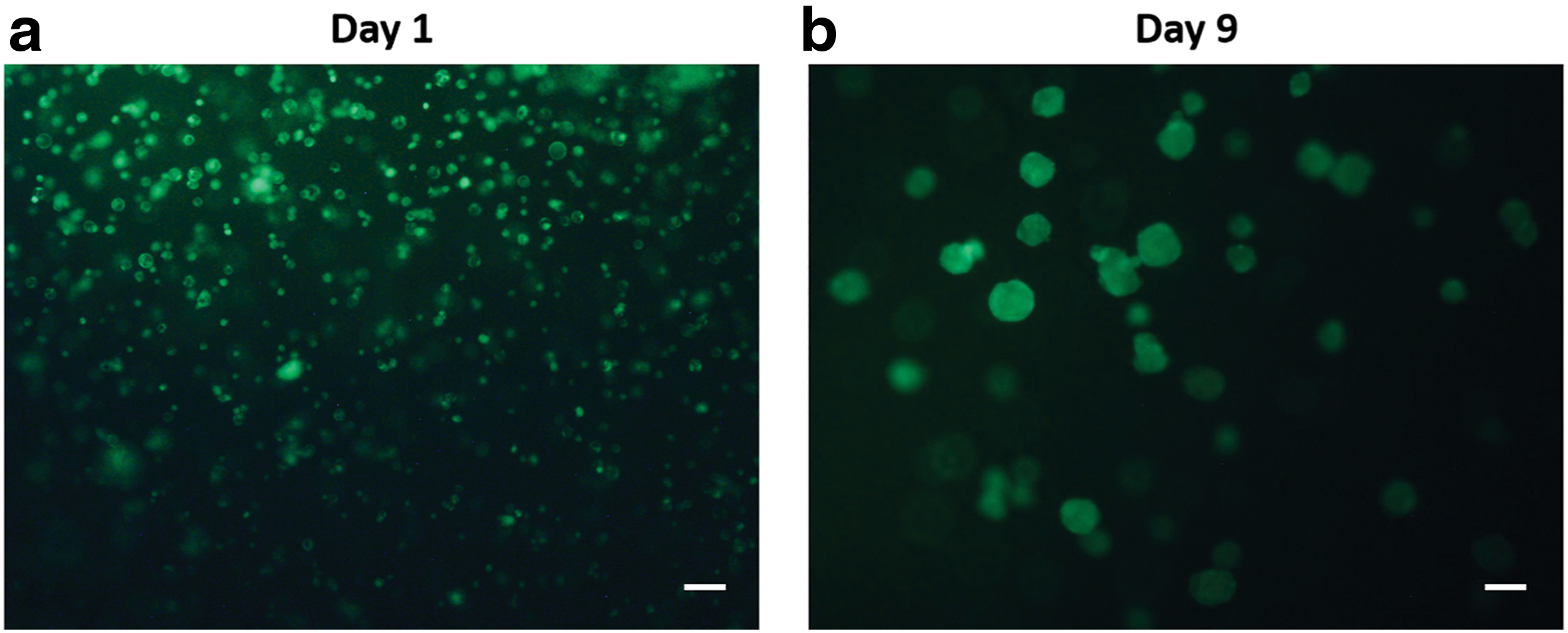
Live and dead staining of UE7T-13 cells cultured in the p(NIPAAm-AA) hydrogel on
Cell migration inside p(NIPAM-AA) hydrogel
The cell-gel hybrids were incubated on top of a porous transwell membrane in three different culture media supplemented with 10% fetal calf serum (FCS) in the upper chamber and 10% FCS in the microwell below the membrane as a control (Fig. 3a control); 10% FCS only in the microwell below the membrane (Fig. 3a downmigration); and 10% FCS only in the upper chamber above the membrane (Fig. 3a upmigration). A chemoattractant (FCS) gradient was developed in the vertical direction owing to the concentration difference of FCS for both downmigration and upmigration. When the chemoattractant concentration decreased from the chamber below the membrane to the chamber above the membrane, cells migrated inside the p(NIPAM-AA) hydrogel to the membrane of the upper chamber, resulting in approximately twofold increase in cell number on the cell membrane, compared to the control (Fig. 3b). However, a greater than 50% decrease in the cell number attached onto the membrane of the upper chamber when the chemoattractant gradient decreased from the chamber above the membrane to the microwell below the membrane. There was no significant difference at two initial concentrations of the chemoattractant (20% vs. 10%). The decrease in the cell number due to cell migration through the p(NIPAAm-AA) hydrogel was also evidenced from the fluorescent DAPI (4′,6-diamidino-2-phenylindole) staining images of the porous membrane of the upper chamber (Fig. 3c).

The cell-hydrogel hybrids were also imaged by standard error of the mean (SEM). Single individual cells were found to locate inside the p(NIPAAm-AA) hydrogel without a chemoattractant gradient on day 1. The SEM images also revealed a porous structure of the freeze-dried p(NIPAAm-AA) hydrogel (Fig. 4a). When a chemoattractant gradient was applied, a migrated cell aggregate was shown at the bottom of the hydrogel, which may migrate through the p(NIPAAm-AA) hydrogel (Fig. 4b).

UE7T-13 cells cultured inside the p(NIPAAm-AA) hydrogel behaved differently from those cultured as cell pellets in the polypropylene (PP) tubes (Supplementary Fig. S5). When cells were embedded in the p(NIPAAm-AA) hydrogel, individual cells were initially well distributed in the hydrogel, which was evident from the images obtained from a CKX41 optical microscope (Fig. 5a). Once cell aggregates were formed after 9 days, collagen, one of the major ECM proteins, was produced around the cell aggregates as shown by the Masson's Trichrome blue color staining (Fig. 5b). After culturing them in inductive media for 8 weeks, the cell aggregates displayed an atypical structure with rich collagen depositing around the cell aggregates, and connection between cell aggregates was established through rich collagen fibrils (Fig. 5c). The p(NIPAAm-AA) hydrogel in the inductive medium was contracted into a smaller size and large cell pellets were seen on the surface of the hydrogel after induction for 4 weeks (Supplementary Fig. S6a). At the late stage of chondrogenic induction, the p(NIPAAm-AA) hydrogel was further contracted and cells were found on the surface of the p(NIPAAm-AA) hydrogel with an adherent morphology surrounded with distinct blue-stained collagen (Fig. 5d). Moreover, cell pellets with a larger size were also seen on the contracted hydrogel. In contrast, large cell pellets in the PP tubes were generated after centrifugation on day 1. Cells were found to gradually detach from cell pellets from day 1 to week 4. As chondrogenic induction progressed, there were more cell aggregates with a small size in the PP tubes (Supplementary Fig. S6b, c). Moreover, these cell aggregates were found to migrate upward along the tube wall and merge with the large cell pellet into a single-cell pellet after induction at week 8.

Mechanisms of hMSCs migration within p(NIPAAm-AA) hydrogels.
Chondrogenesis
After inducing chondrogenesis for 4, 6, and 8 weeks, the expression of chondrogenesis-related genes (SOX9, ACAN, and COL 2A1) in the cells cultured in PP tubes and p(NIPAAm-AA) hydrogels was assessed using qRT-PCR with the primer pairs indicated in Table 1. The mRNA expression of SOX9, ACAN, and COL 2A1 in cells cultured inside p(NIPAAm-AA) hydrogels was significantly higher than that for cell pellets in the PP tubes at week 4 (Fig. 6a). After 6 weeks, a significant difference was still seen for the mRNA expression of SOX9 and COL 2A1, while no difference was seen for ACAN (Fig. 6b). After inducing chondrogenesis for 8 weeks, the mRNA expression of ACAN in cells cultured inside p(NIPAAm-AA) hydrogels was higher than that for cell pellets in PP tubes, while no significant difference was seen in the mRNA expression of SOX9 and COL 2A1 (Fig. 6c).
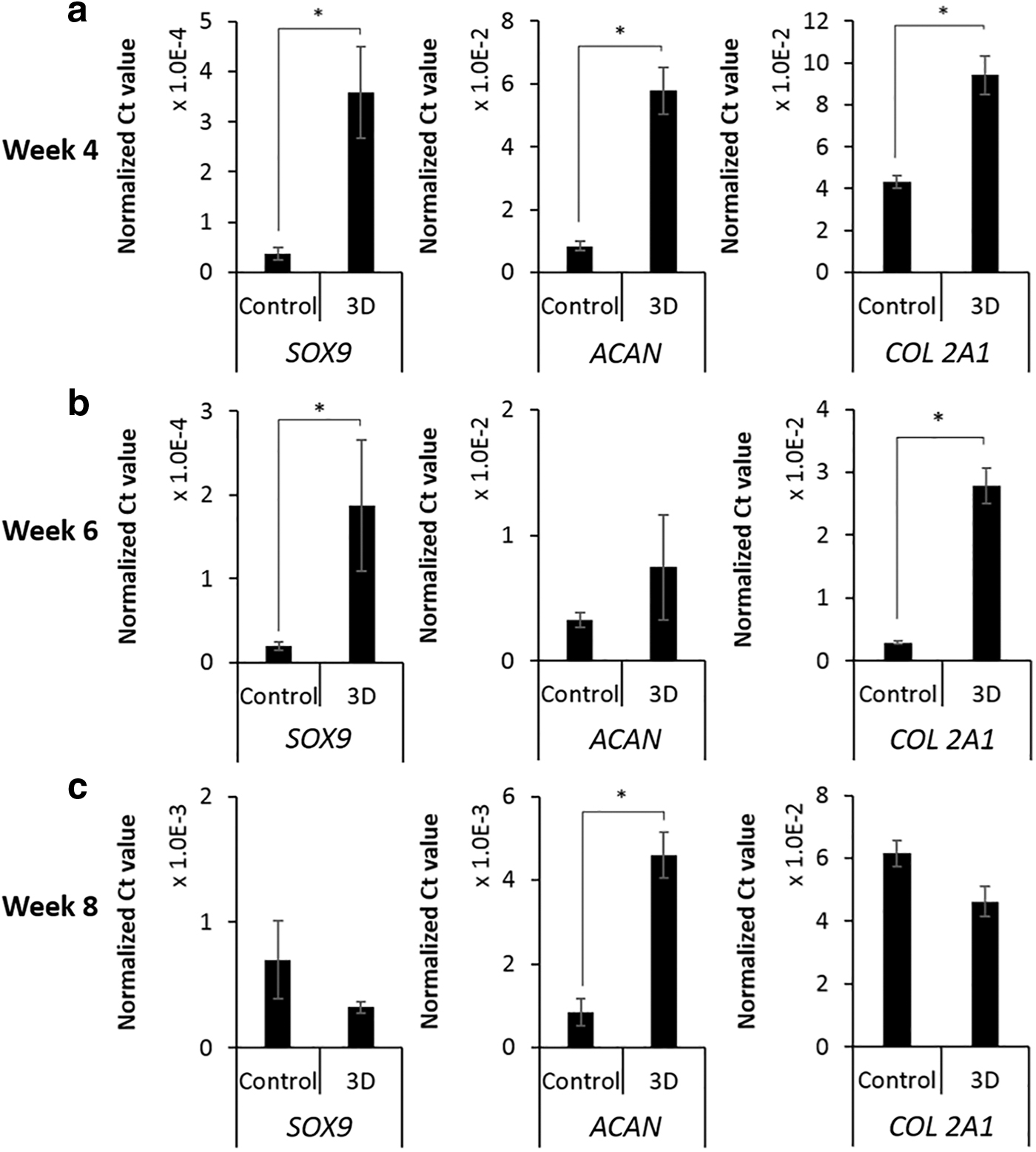
mRNA expression of chondrogenic genes (SOX9, ACAN, and COL 2A1) in UE7T-13 cells induced with chondrogenic inductive media for
Gene Sequence Of Primer Pairs
Furthermore, histological and immunofluorescent staining also revealed deposition of sulfated glycosaminoglycans (GAGs) and collagen 2A1 within the neocartilage formed in PP tubes and p(NIPAAm-AA) hydrogels (Fig. 7). The deposited sulfated GAGs were mainly distributed at the peripheral area of the neocartilages at week 4. However, sulfated GAGs produced in the neocartilage formed in the p(NIPAAm-AA) hydrogels were more distinct than those formed in the PP tubes. In addition, it was seen that the cell pellets formed in the PP tubes had a porous structure at weeks 4 and 8; however, the porous cell pellets formed in the p(NIPAAm-AA) hydrogels at week 4 possessed a more compact structure and the internal spaces between cells were filled with sulfated GAGs at week 8. In contrast, collagen 2A1 was mainly distributed in the central area of the neocartilages in both systems at week 4. A similar distribution of collagen 2A1was still seen in the neocartilages in the PP tubes after week 8, while the collagen 2A1 in the neocartilages embedded in the p(NIPAAm-AA) hydrogels was more uniformly distributed, which was in accordance with the distribution of sulfated GAGs.
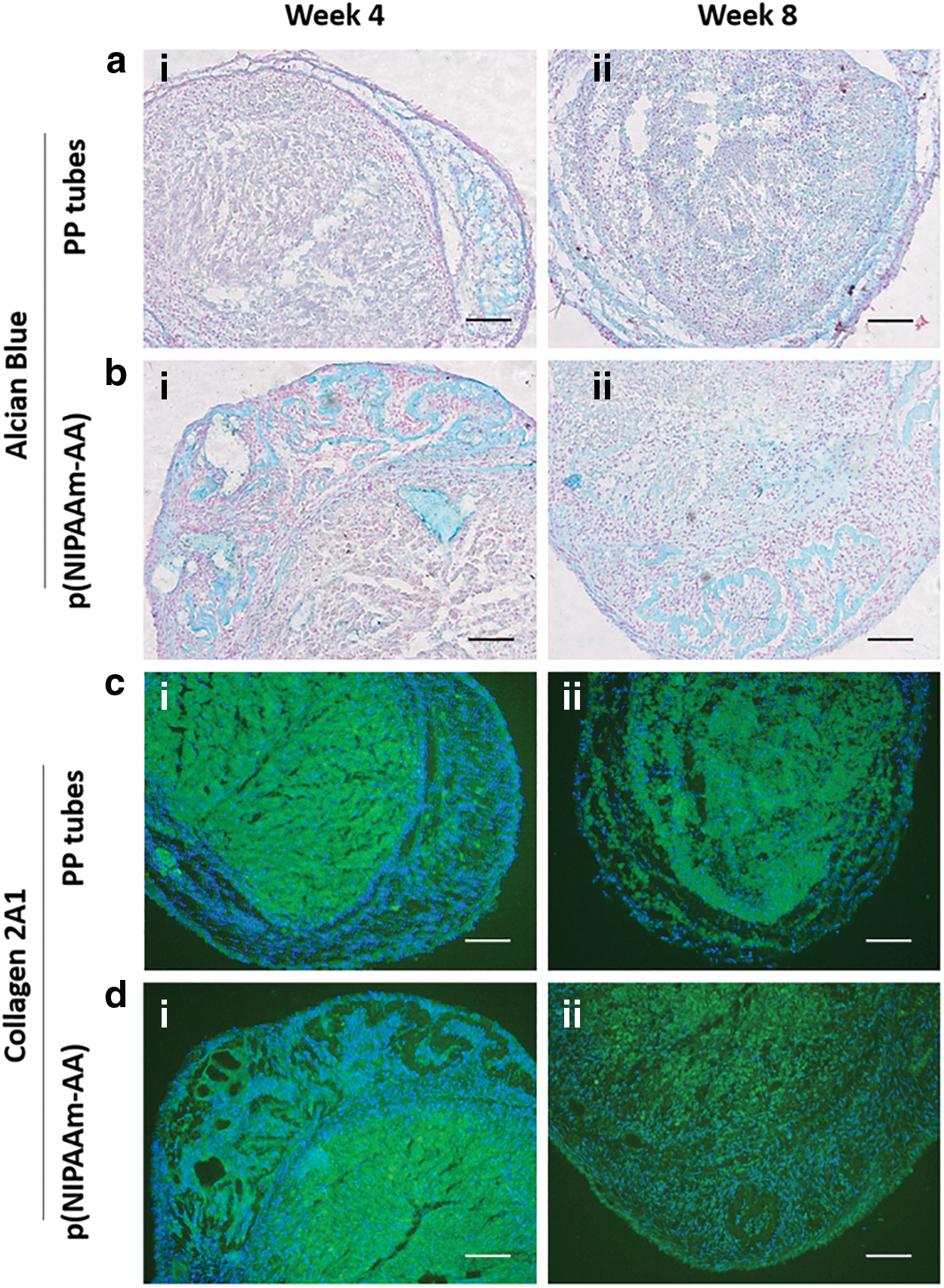
Alcian blue staining images (Red: cell nuclei and Blue: sulfated GAGs) and immunofluorescent staining images (Blue: DAPI-labeled cell nuclei and Green: FITC-labeled collagen 2A1) of UE7T-13 cells cultured
Fabrication of a neocartilage patch
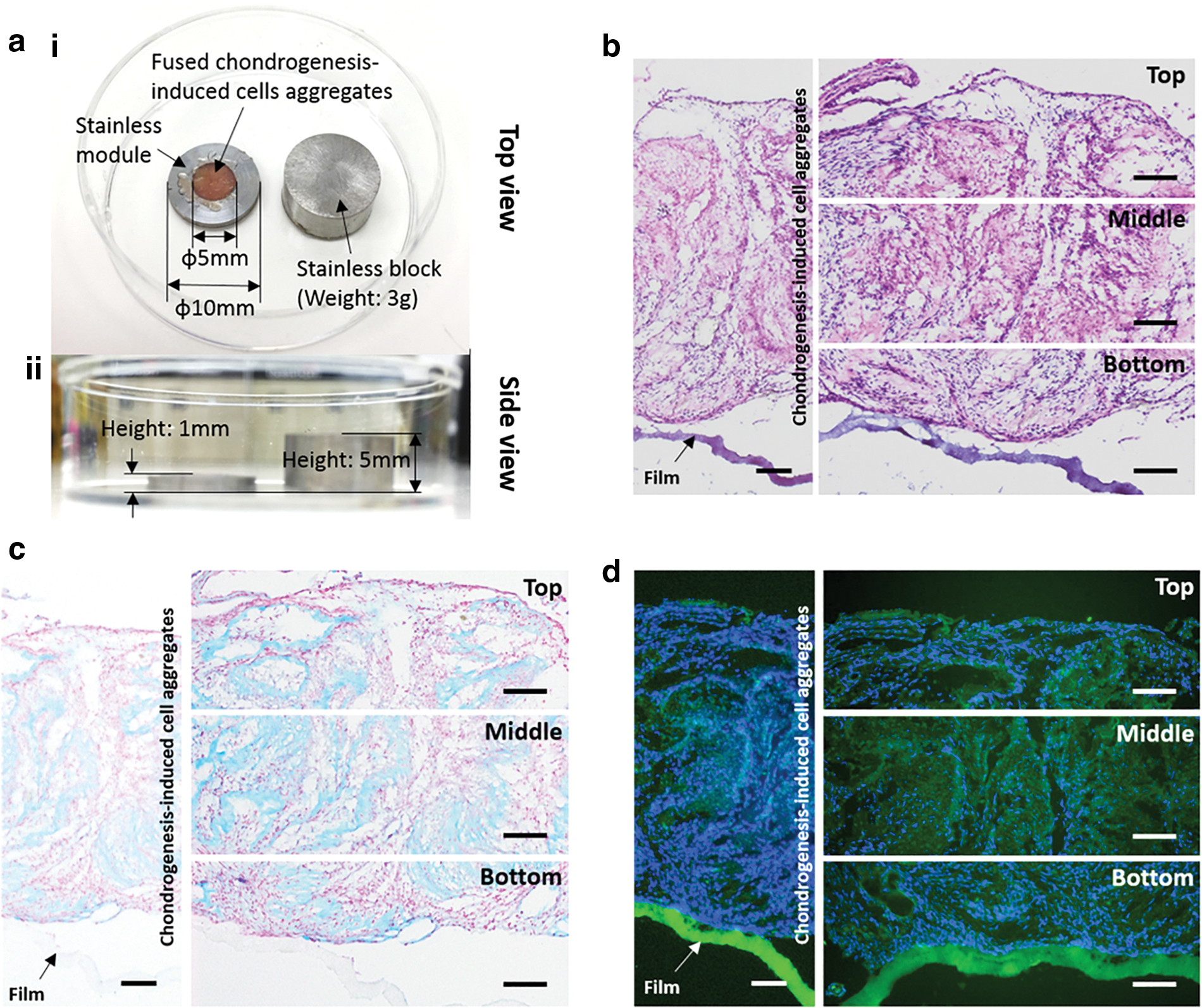
After transferring chondrogenesis-induced cell aggregates onto the electrospun film, cells displayed an adherent fibroblast-like morphology (Supplementary Fig. S7). Moreover, the cells were merged into a neocartilage with a size of 5 mm in diameter and 1 mm in height by the spatial confinement of the stainless-steel mold and block (Fig. 8a). Furthermore, the histological staining revealed that the neocartilage was attached to the electrospun film (Fig. 8b). Sulfated GAGs and collagen 2A1 within the neocartilage were stained with a blue color by the Alcian Blue staining and a green color by immunofluorescent staining, respectively (Fig. 8c, d).
Discussion
Cell aggregates were present within the p(NIPAAm-AA) hydrogel on day 9 (Fig. 2), while the mechanism of forming cell aggregates was unclear. We hypothesized that cell migration, inside the hydrogels, might contribute to formation of cellular aggregates. Mg2+-/COO−-induced ionic crosslinking and hydrophobic interaction resulted in cavities with various sizes within the p(NIPAAm-AA) hydrogel 27 (Fig. 4). When individual cells were homogeneously embedded inside the p(NIPAAm-AA) hydrogel at 37°C, due to the absence of cellular binding sites in the p(NIPAAm-AA) polymeric structure, they tend to show individual cells. However, due to the intrinsic gravity, those cells distributed within the same cavities may initiate cell-cell interactions leading to the formation of cell aggregates. This may provide an explanation for the adherent cells on the membrane of the microwell insert even without a chemoattractant gradient (Fig. 3b). When a 10% of FCS gradient was applied from the well below the membrane to the chamber above the membrane, the embedded cells moved downwards through interfiber pores due to their intrinsic gravity and a chemoattractant gradient. Amoeboid-like migration of cells might be induced due to spatial confinement and the weak interaction forces between cells and supporting hydrogels. 28 Hence, adherent cells on the membrane of the upper chamber were significantly increased compared to the control without a chemoattractant gradient (Fig. 3b) and a cell aggregate was seen to migrate through the p(NIPAAm-AA) hydrogel network to the bottom (Fig. 4b). When the chemoattractant gradient was generated by decreasing 10% FCS from the chamber above the membrane to the well below the membrane, upward amoeboid-like migration of the embedded cells might be realized through the polymeric network. Cell migration was not affected by the initial concentration of the chemoattractant, which might be due to the saturation of the FCS receptors on the cell membrane.29,30 A small number of adherent cells were still seen on the membrane even in the presence of a upward chemoattractant gradient, because these cells might retain in the bottom of the p(NIPAAm-AA) hydrogel, where the confinement for single cells was not sufficient to initiate amoeboid-like migration and overcome the cell gravity (Fig. 3c). The embedded cells might also be able to temporarily liquefy the surrounding p(NIPAAm-AA) hydrogel by chemoattractant-induced stiffening on the cell membrane and disrupting the ionic crosslinking to induce stress relaxation of the hydrogel 31 because of the weak mechanical structure of the p(NIPAAm-AA) hydrogel. 22
It was widely reported that cell aggregates were capable of enhancing secretion of endogenous growth factors and deposition of ECM proteins, 8 both of which might promote chondrogenesis inside the p(NIPAAm-AA) hydrogel compared to cell pellets, as evidencing in the higher expression of chondrogenesis-related genes (Fig. 6) and production of two major ECM components in cartilage (Fig. 7). 32 The difference in cellular behavior in two culture systems might be due to developmental properties of cell aggregates. Because the p(NIPAAm-AA) hydrogel facilitated the formation of small cell aggregates at the early stage, the surface-to-volume ratio of the cells cultured inside the p(NIPAAm-AA) hydrogel was substantially larger compared with cells cultured as cell pellets, resulting in more efficient transportation of both endogenous growth factors and exogenous inductive components present in the media for small cell aggregates. 33 The production of ECM proteins (Fig. 5b) was further enhanced around these small cell aggregates after incubating these aggregates in the chondrogenic medium. The accumulated ECM could interact with the p(NIPAAm-AA) hydrogel to build “bridges” within the p(NIPAAm-AA) hydrogel for the embedded cell aggregates (Fig. 5c). Cell aggregates could, therefore, migrate inside the p(NIPAAm-AA) hydrogel by binding to the deposited ECM proteins, 34 which led to the contraction of the p(NIPAAm-AA) hydrogel and establishment of contacting communication between cell aggregates. In contrast, it was found that even though cells were forced into large cell pellets by centrifugation, some cells gradually detached from the cell pellets, leading to a decrease in the size of cell pellets when culturing in the complete culture medium without inductive biomolecules (Supplementary Fig. S5). Those detached cells would re-form cell aggregates within the PP tubes and merge into a larger cell aggregate when cells were incubated in the inductive medium, which was quite similar to the development of cell aggregates inside the p(NIPAAm-AA) hydrogel (Supplementary Fig. S5). Meanwhile, cell migration was also observed for cells cultured in PP tubes due to the enhanced deposition of ECM by the cell aggregates on the tube wall. The mobility of small cell aggregates was better compared with large cell pellets, resulting in a longer migrated distance (Supplementary Fig. S6b), which was more distinct for cells cultured in the chondrogenic induction medium in the PP tubes (Supplementary Fig. S6c). However, the fusion of cell aggregates in the PP tubes was not as efficient as that in the p(NIPAAm-AA) hydrogels since endogenous growth factors secreted from cell aggregates would rapidly diffuse into the medium, but could not form a gradient to initiate migration of cell aggregates. In contrast, due to the obstacle of the p(NIPAAm-AA) polymeric network, endogenous growth factors could be transiently retained around the cell aggregate, generating a chemoattractant gradient to initiate the migration of cell aggregates and fusion of cell aggregates. 29 Therefore, a relatively large neocartilage was formed within the p(NIPAAm-AA) hydrogel (Fig. 5d and Supplementary Fig. S6a). Although no significant difference was found in the mRNA expression of chondrogenesis-related genes (ACAN (week 6), SOX9 and COL 2A1 (week 8) (Fig. 6b, c) and the deposition of extracellular matrices (sulfated GAGs and collagen 2A1 [week 8]) (Fig. 7ii) for cells cultured in the PP tubes and the p(NIPAAm-AA) hydrogel, more efficient transportation of biomolecules within small cell aggregates and the developmental difference of cell aggregates reinforced the p(NIPAAm-AA) hydrogel as a better platform for short-term chondrogenesis of mesenchymal stem/stromal cells.
Because of reversibility of the p(NIPAAm-AA) hydrogel (Supplementary Fig. S2), these chondrogenesis-induced cell aggregates were harvested from the hydrogel by simply diluting the hydrogels at room temperature and transferred onto the electrospun films (Supplementary Fig. S7). The mechanical loading of a stainless-steel block could, on the one hand, induce a compressive stress on the cell aggregates to enhance the chondrogenesis of cell aggregates 35 and accelerate the attachment of cell aggregates onto the electrospun film. 36 On the other hand, the stainless-steel block together with the stainless-steel ring also created a spatial confinement for the cell aggregates, which forced cell fusion into an integrated neocartilage. A neo-cartilage patch with a diameter of 5 mm and a height of 1 mm was successfully fabricated (Fig. 8). The neo-cartilage patch could be potentially used for treatment of an irregular degenerated cartilage by simply attaching it to the defect sites and initiating cartilage integration when bearing a compressive stress, even though an in vivo further study still needs to be performed.
Conclusion
We have demonstrated a neocartilage patch by forcing MSC aggregates formed in thermosensitive p(NIPAAm-AA) hydrogel onto an electrospun film, which might be potentially used for the treatment of irregular degenerated cartilage by simply attaching to the defect sites and initiating cartilage integration when bearing a compressive stress.
Footnotes
Acknowledgments
J.B.Z. thanks the IPE-UoA Scholarship (BES).
Disclosure Statement
No competing financial interests exist.
Funding Information
H.Z. acknowledges financial support from the Australian Research Council Discovery Project (Grant no. DP160104632) and The Medical Advancement Without Animal (MAWA) Trust.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.