
Abstract
Stem cell–replacement therapies have been proposed as a potential tool to treat sensorineural hearing loss by aiding the regeneration of spiral ganglion neurons (SGNs) in the inner ear. However, transplantation procedures have yet to be explored thoroughly to ensure proper cell differentiation and optimal transplant procedures. We hypothesized that the aggregation of human embryonic stem cell (hESC)–derived otic neuronal progenitor (ONP) cells into a multicellular form would improve their function and their survival in vivo post-transplantation. We generated hESC–derived ONP spheroids—an aggregate form conducive to differentiation, transplantation, and prolonged cell survival—to optimize conditions for their transplantation. Our findings indicate that these cell spheroids maintain the molecular and functional characteristics similar to those of ONP cells, which are upstream in the SGN lineage. Moreover, our phenotypical, electrophysiological, and mechanical data suggest an optimal spheroid transplantation point after 7 days of in vitro three-dimensional (3D) culture. We have also developed a feasible transplantation protocol for these spheroids using a micropipette aided by a digital microinjection system. In summary, the present work demonstrates that the transplantation of ONP cells in spheroid form into the inner ear through micropipette 7 days after seeding for 3D spheroid culture is an expedient and viable method for stem cell replacement therapies in the inner ear.
Impact statement
Sensorineural hearing loss affects millions of people worldwide. Although inner ear stem cell replacement therapies offer a promising method to mitigate this hearing loss by aiding in the regeneration of spiral ganglion neurons, few in vivo studies have been performed in this area. By providing detailed cell spheroid characterization, determining an optimal development stage for transplantation, and establishing a reliable and reproducible transplantation protocol, we aimed to provide the necessary details to bridge the gap between in vitro experiments and in vivo studies of stem cell replacement therapies in the inner ear.
Introduction
Stem cell replacement therapies in the inner ear may aid regeneration of spiral ganglion neurons (SGNs) and mitigate the effects of sensorineural hearing loss (SNHL), which typically results from destruction of hair cells followed by loss of SGNs.1–3 Recently, human embryonic stem cells (hESCs) have become invaluable tools to study tissue regeneration because they can differentiate toward an otic neuronal lineage, the precursor to SGNs.4–7 Despite promising initial results, however, low engraftment and immature otic neuronal differentiation still pose significant challenges. These challenges are at least partially due to the inner ear's inhospitable environment, which consists mainly of bone and the nutrient-poor perilymph.8–10 As a result, transplanted hESC-derived cells cannot interact with native surrounding cells to regulate and promote proliferation, differentiation, and survival. Recent tissue engineering advancements have enabled the generation of more hospitable artificial microenvironments (i.e., stem cell niches) with the necessary cellular components, extracellular matrices (ECMs), and secreted factors to support transplanted cells.11–13
The use of three-dimensional (3D) cell culture techniques to replicate these niches have gained traction: for example, the formation of cellular aggregates (e.g., spheroids) enhances cellular functional properties.14,15 It has been reported that the formation of spheroids can upregulate the secretion of neurotrophic factors. 16 Furthermore, the formation of hESC spheroids demonstrably enhances the secretion of ECM components, including fibronectin and laminin, which can promote hESCs' neuronal differentiation, prolong cell survival, and provide better protection from neurotrophic factor deprivation.17–19 In addition, neural cells in particular have been grown successfully in 3D culture, which provides such benefits over flat culture as increased neurite outgrowth. 20 Lastly, it is known that neural cells can self-assemble into spheroid structures without the requirement for a provided matrix.21,22
However, the precise impact of 3D spheroidal cellular organization and structural environment on hESC-derived otic neuronal progenitors (ONPs) has not been investigated. We hypothesized that the formation of hESC-derived ONP spheroids facilitates otic neuronal differentiation and improves cell viability after transplantation in the inner ear, as an alternative to single-cell transplantation. Since in vivo animal studies are expensive, low-throughput, and unpredictable, 3D spheroids must be extensively analyzed before transplantation to ensure maintenance of proper cell characteristics and to determine the ideal time for transplantation. In particular, hESC-derived ONP spheroids must be examined to ensure that they maintain the ONP characteristics necessary for regeneration of SGNs. Moreover, a protocol must be developed to establish that these spheroids can be effectively transplanted at the optimum point for maximum cell survival.
Therefore, we first constructed spheroids from pluripotent hESCs. These spheroids were characterized to evaluate cellular, molecular, mechanical, and functional properties to ensure that the cells displayed neuronal progenitor characteristics and to determine the best transplantation point. We also investigated specific parameters for a spheroid transfer procedure to efficiently transplant spheroids into the inner ear with minimum stress on the cells. Results indicate that hESC-derived ONP spheroids possess attractive characteristics for stem cell replacement therapies in the inner ear and that culturing spheroids for 7 days in vitro allows sufficient development of ONPs before transplantation.
Materials and Methods
Generation of hESC-derived ONP spheroids
hESC lines H1, H7, and H9 (WiCell, Madison, WI) were differentiated toward an otic neuronal lineage according to our previously described protocol (Fig. 1). 5 Upon reaching the late ONP stage (Fig. 1) on day 25 (D25), cells were passaged to a 96-well Clear Round Bottom Ultra-Low Attachment Microplate® (U-bottom, #7007; Corning Life Science, Tewksbury, MA) and seeded at either 25,000; 50,000; or 75,000 cells per well. The plate was then centrifuged at 201 relative centrifugal force for 10 min and left incubating at 37°C for 7–28 days. For our mechanical assessment, additional late-stage ONP spheroids were generated in a different culture system by seeding 1.8–2.5 million cells in an EZSPHERE™ culture plate (Cat#:4000-900SP, Nacalai USA, San Diego, CA, manufactured by AGC Techno Glass, Shizuoka, Japan), which has 2300 microwells designed for spheroid formation on its surface. Spheroid day in vitro 1 (DIV 1) was defined as the first day when late-stage ONPs were transferred into a spheroid-forming plate. Spheroids generated by U-bottom plate were imaged regularly after formation with a Nikon Eclipse TS100 microscope (Nikon, Tokyo, Japan). Each week, spheroids seeded at 50,000 cells per well were fixed for 20–30 min with 4% paraformaldehyde (Cat#: 158127; Millipore Sigma, Burlington, MA) and rinsed twice with Dulbecco's phosphate-buffered saline (DPBS) (Corning, Life Sciences). Fixed spheroids were stored in DPBS at 4°C before use. Spheroids seeded at 50,000 cells per well were used for all experiments, except for the analyses of diameter, circularity, and roundness.
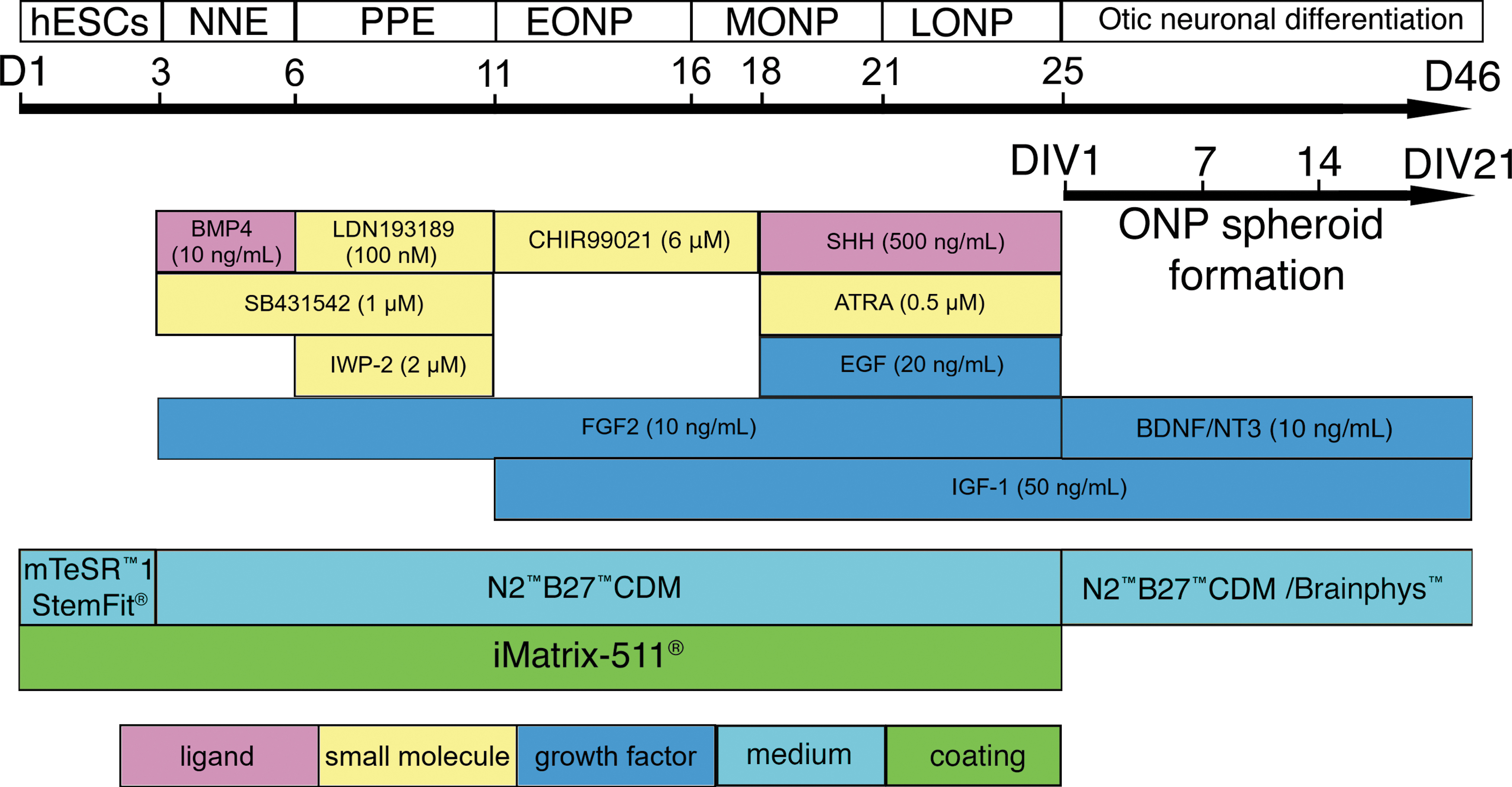
Schematic summary of the protocol and timeline for generating late-stage ONPs from undifferentiated hESCs. Developmental stage is shown at the top, with key treatments shown below. D, day of cell culture; DIV, day in vitro after three-dimensional seeding; hESCs, human embryonic stem cells; NNE, non-neuronal ectoderm; PPE, preplacodal ectoderm; EONP, early otic neuronal progenitor; MONP, mid-otic neuronal progenitor; LONP, late otic neuronal progenitor; BMP4, bone morphogenetic protein 4; SHH, Sonic hedgehog; ATRA, all-trans retinoic acid; EGF, epidermal growth factor; IGF-1, insulin-like growth factor 1; FGF2, fibroblast growth factor 2; BDNF, brain-derived neurotrophic factor; N2B27-CDM, chemically defined medium containing N2 and B27 supplements. Color images are available online.
Image acquisition and cell quantification
Samples were imaged with a Leica TCS SP5 confocal microscope (Leica, Wetzlar, Germany). For quantitative analyses on hESC-derived ONP spheroids, the average diameter of each spheroid was measured. Here, the geometric mean diameter of a spheroid (Dg) is defined as follows:
where a and b are the major and minor axes of the diameter, respectively. Using this equation, we measured the spheroids' diameters using Microscope Imaging Software (Leica Microsystem, Inc., Buffalo Grove, IL).
Sphericity and roundness are metrics to measure the compactness of a shape.
23
Sphericity describes the similarity to pure spheroidal geometry for each spheroid. Given that we measured the diameter of a spheroid in single two-dimensional (2D) images, we used the circularity index (C), which is the 2D equivalent of the true 3D spheroid index and ranges from 1 (perfect sphere or circle) to 0 (elongated shape).
24
For the circularity index,
where S is the spheroid surface area as measured on a 2D spheroid image with the largest diameter, selected from the Z-stack images of each spheroid. P is the spheroid perimeter. We also used a roundness measurement to describe the similarity of the spheroid images to a perfect circle (the shape's gross feature). Roundness was defined as follows
24
:
where S is the surface area and a is the major axis of the diameter.
Terminal deoxynucleotidyl transferase dUTP nick-end labeling
DNA fragmentation was detected by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining to evaluate cell viability in spheroids. At weekly time points, spheroids seeded at 50,000 cells per well were fixed, frozen, and sliced into 10 μm sections on a Leica CM3050 S cryostat (Leica). Center sections were treated with the In Situ Cell Death Detection Kit (Roche, Basel, Switzerland) according to manufacturer's instructions to detect the DNA fragmentation indicative of cell death. Samples were then imaged with a Leica TCS SP5 confocal microscope. Cells with TUNEL staining restricted to the nuclei as confirmed by DAPI double staining were considered apoptotic (TUNEL-positive); diffusely TUNEL-stained cells with no apoptotic morphology were excluded based on previously used criteria. 25 TUNEL-positive cells were counted and expressed as a ratio of total cells in sections counterstained with DAPI for each region. Data are expressed as percent DAPI-stained cells.
Three-dimensional immunocytochemistry
Fixed spheroids were blocked overnight at room temperature in 10% bovine serum albumin (Sigma-Aldrich, St. Louis, MO) with 0.1% Triton X-100 (Sigma-Aldrich) in DPBS before incubation with primary antibodies (Supplementary Table S1). After 2 days, spheroids were washed three times with DPBS with 0.1% Triton X-100 and then incubated in secondary antibody solution (Supplementary Table S2) for 3 days. Spheroids were counterstained with TOTO-3 iodide (1:5000–1:10,000 in DPBS, T3604; Thermo Fisher Scientific, Waltham, MA) for 45 min followed by a final wash. Immunocytochemistry controls were performed as previously described.26,27 Primary antibody controls were obtained with hESC-derived ONPs with a second primary antibody to the same antigen. 26 Secondary antibody controls were also obtained each time multiple primary antibodies were used. 26 Labeling controls (detection controls) were performed for a sample from each batch of hESC culture (Supplementary Fig. S1). 26 Samples were imaged with a Leica TCS SP5 confocal microscope (Leica). The quantification of immune-positive cells has been previously described in detail (Supplementary Data).5,6
Neurite arborization assay
The pan-neuronal marker β-III tubulin was used to calculate neurite arborization area. Individual confocal optical Z-stack sections were examined through the extent of each spheroid at the observation time point using ImageJ ver. 2.0.0-rc-69/1.52p (National Institute of Health, Bethesda, MD). 28 The watershed algorithm was extensively used to identify individual nuclei. 29 Each Z-stack photomicrograph was transformed into an 8-bit grayscale.30,31 Quantification was assessed after defining a threshold for background correction using the “MaxEntropy” method of the AUTO THRESHOLD plugin.32,33 Arbor surface area (μm2) was calculated using an established protocol and was normalized with each Z-stack spheroid surface area, as spheroid sizes varied depending on the culture length. 32
Whole-cell patch-clamp recordings
To test the efficacy of certain culture conditions for neuronal differentiation, ONP spheroids were cultured in early neuronal stage differentiation medium as previously described. 5 ONP spheroids were cultured with a polyhedrin delivery system (PODS®, #PPH1-50; Cell Guidance Systems, Cambridge, United Kingdom) carrying brain-derived neurotrophic factor (BDNF), designed to release BDNF at a regular rate into medium. Spheroids were also cultured with GrowDex-T® (#200103010, UPM-Kymmene Corporation, Helsinki, Finland), a nanofibrillar cellulose hydrogel used to simulate an ECM. PODS were supplied at 800,000 per spheroid. At given time points, these spheroids were examined by patch clamp. Standard voltage- and current-clamp recordings were performed using an Axopatch™ 200B amplifier (Axon Instruments, Inc., Union City, CA) following established protocol (Supplementary Data). 34
Mechanical characterization of ONP spheroids
To examine the mechanical properties of hESC-derived ONP spheroids, spheroids formed using both a 96-well Clear Round Bottom Ultra-Low Attachment Microplate (U-bottom) and an EZSPHERE culture plate were grown until DIV 4, at which point spheroids were immobilized and tested with a single indentation protocol using a Piuma nanoindenter (Optics11, Amsterdam, Netherlands). Subsequent nanoindenter testing was performed on DIV 8 and DIV 11. Control data were obtained using mouse brain samples (Supplementary Data). MATLAB (version 9.5.0 1067069 (R2018b) Update 4, The MathWorks, Inc., Natick, MA) was used to analyze the load–displacement data. The Hertz method was used for analysis because the use of the loading portion of the load–displacement curve is far more suitable for soft biological samples.
35
The Hertz model assumes a linear elastic material response. The elastic Young's modulus was calculated as follows:
where E is the elastic Young's modulus, h is the cantilever deflection (nm), R is the radius of the spherical indenter tip (μm), and v is Poisson's ratio (Supplementary Data).
ONP spheroid mock transplantation using a micropipette
For mock transplantation of hESC-derived ONP spheroids, we used a cell-holding system originally developed for in vitro fertilization, designed to capture cell aggregates at the tip of a micropipette with negative pressure. Spheroid transfer pipettes were pulled from borosilicate glass with a Sutter P-97 Flaming/Brown Micropipette Puller (Sutter Instrument, Novato, CA). Micropipettes were then broken to size using the score and break technique, with a Sutter Instruments Ceramic Tile for Scoring Glass (NC9569052; Sutter Instrument) used to score the glass and allow for a clean break. Pipettes were not fire polished. Please also see the Supplementary Data for details.
The transplantation procedure was simulated using a Xenoworks Digital Injector (Sutter Instrument) system with micropipettes in a range of sizes. For transfer, the pipette tip was placed at the surface of a spheroid and the negative pressure manipulated to hold the spheroid in place. Upon moving the pipette tip with the captured spheroid out of the initial culture medium to the desired area of transfer, positive pressure released the spheroid from the micropipette. Spheroids were placed in a 96-well culture dish filled with culture medium to simulate placement into the fluid-filled scala tympani. A range of micropipette sizes was used to determine the optimal conditions for transferring spheroids with a low amount of stress. Pressures ranged from −124 to +55 hPa.
Live/dead assay of transferred spheroids
After transfer, a subpopulation of spheroids was cultured in medium with the Invitrogen LIVE/DEAD™ Viability/Cytotoxicity Kit (Invitrogen, Carlsbad, CA) for 2–3 h according to manufacturer's protocol. Positive and negative controls are shown in Supplementary Figure S2. Images of the surface of each transferred spheroid were taken using a Leica TCS SP5 confocal microscope. Using ImageJ, a threshold was set on each image to distinguish live and dead cells. Images were made binary and the total area of cells was measured for each category, live and dead. Cell survival was quantified as the ratio of live cell area to combined live and dead cell area.
Nonlinear curve-fitting analysis
Data were transferred to JupyterLab (version 1.0.2) (Project Jupyter, Worldwide) as a comma-separated value file for a nonlinear curve-fitting analysis using Python (version 3.8.0) (Python Software Foundation, Wilmington, DE). The following Python libraries were used for the analysis: Scipy, Numpy, Matplotlib, and Seaborn.36–39
A monoexponential equation was used to fit data using the “scipy.optimize.curve_fit” and “scipy.optimize.least_squares” modules in Scipy. These two modules use the Levenberg–Marquardt algorithm based on the Gauss–Newton method.40,41 The algorithm returns a vector of estimated coefficients by the nonlinear curve-fit using the model specified by the following equation:
where y is an estimated spheroid diameter, t is time,
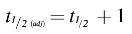
Statistical analyses
When appropriate, and as indicated in each figure, statistical analysis was performed. The value of “n” represents biological replicates. For instance, in our measurement of elastic Young's modulus, it was necessary to confirm that each measurement was reliably performed and as such, we used three to five technical replicates for the measurement on each spheroid to determine one biological replicate (i.e., n).42,43 The sample size (n) is stated in each figure caption. Experimental values are typically expressed as mean ± standard error (SE). Median, standard deviation (SD), and first and third quartiles were used in some cases. All statistical analyses were performed with R (version 4.0.0. 2020-04-24, Vienna, Austria). 44 Normal distributions were assumed unless mentioned otherwise. Analysis of variance (ANOVA) was performed with a Tukey's Honest Significant Different (HSD) post hoc test. Shapiro–Wilk normalization tests and F-tests were also performed. p values smaller than 0.05 were considered statistically significant.
Results
Characterization of hESC-derived ONP spheroids
Human ESC-derived ONPs were seeded at three different densities—25,000; 50,000; and 75,000 cells per well—and allowed to grow for 3 weeks. All spheroids consistently decreased in size (Fig. 2A), and the rate of shrinkage slowed over time. Images of prespheroid cultures taken on spheroid day in vitro (DIV) 1 an hour after seeding show a primarily monolayer culture, with a 3D structure beginning to form around the culture perimeter of each well (white arrow). Figure 2B shows the diameter of hESC-derived ONP spheroids at six different time points. Nonlinear monoexponential curve fit was performed on each cell density (Fig. 2C). On each analysis, the decay constant (τ) and the half-life time (t½) of a fitted curve were calculated. The R2 and RSE values were also calculated for evaluating goodness of fit (Table 1). The decay constant was greater at the seeding cell density of 75,000 cells/well than 25,000 cells/well, suggesting that greater cell density leads to a faster decay. Half-life times (t½) of fitted curves for all cell densities showed an average of 2.07 ± 0.374 days (mean ± SD), whereas t½ (adj) had an average of 3.07 ± 0.374 days. Finally, the R2 and RSE values, along with the visual examination of fitted curves, demonstrated reasonable curve fit of the three cell densities. Figure 2D and E show circularity and roundness, respectively, of spheroids at six different time points. The mean circularity index of spheroids at the cell density of 25,000 cells/well was significantly lower than the other two conditions (p < 0.05) (Fig. 2D). Circularity indices and roundness slowly declined after DIV 7.
Development of hESC-derived ONP spheroids over a 3-week culture period.
The Decay Constant (τ), Half-life Time (t½), Adjusted Half-Life Time (t½(adj)), Correlating Coefficient (R 2 ), and Residual Standard Error for the Nonlinear Monoexponential Fitting Curve for the Diameter Change of hESC-derived ONP Spheroids Over Time with Three Different Cell-Seeding Densities (25,000 (25K); 50,000 (50K); and 75,000 (75K) Cells/Well)
M, mean; SD, standard deviation.
TUNEL assay
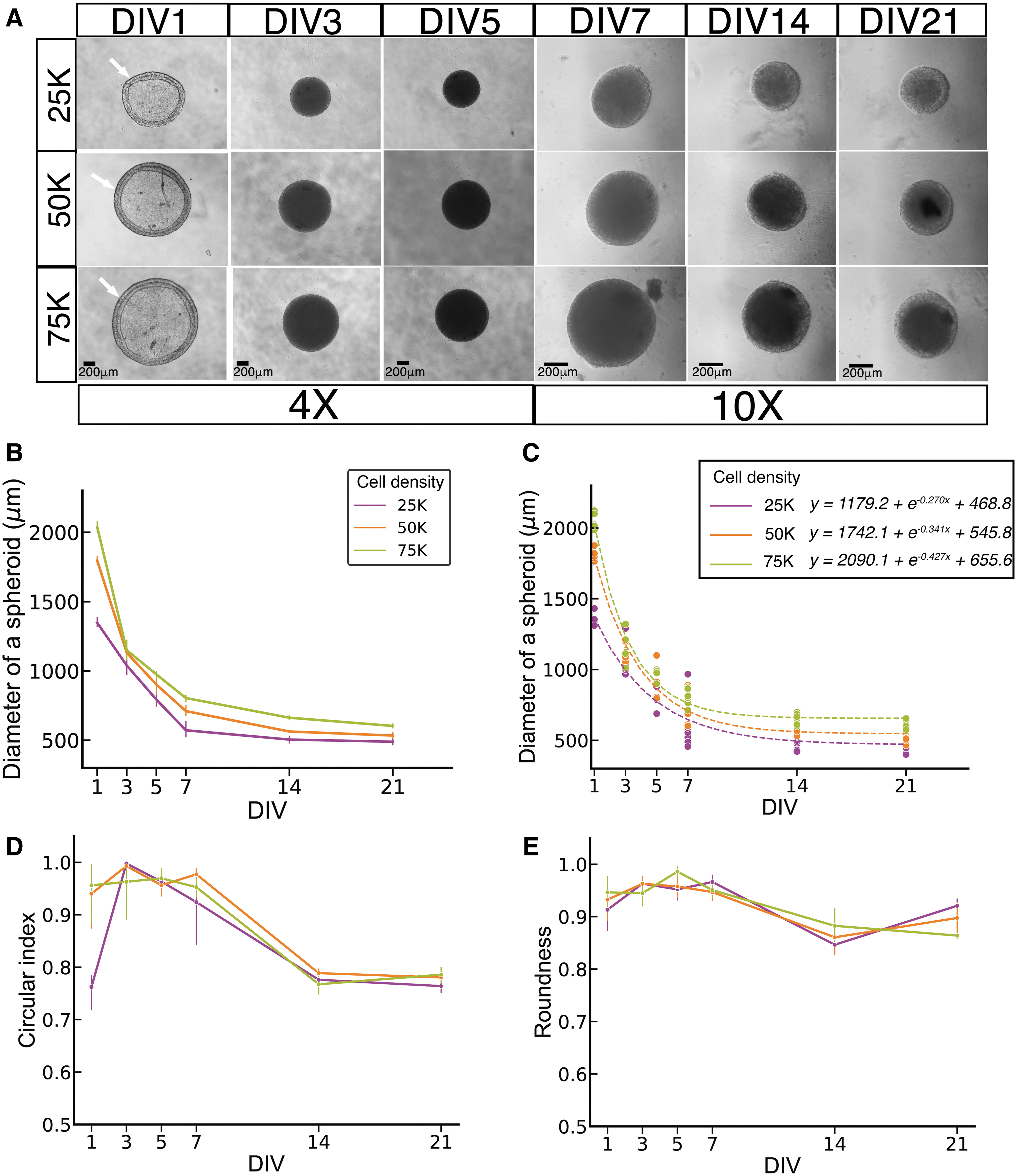
TUNEL-positive cells displaying apoptotic morphology were present in spheroids at all time points (Fig. 3A). However, quantitative analysis (Fig. 3B) indicated that the ratio of such cells increases significantly after DIV 14, with nonconcentric configuration.
Terminal deoxynucleotidyl transferase dUTP nick-end labeling.
Immunocytochemistry
Immunocytochemistry was performed on spheroids to assess ONP characteristics (Fig. 4A). All spheroids highly expressed GATA3, an otic lineage marker; PAX8, another otic lineage marker that is positive for the otic placode and vesicle; and pan-neuronal marker β-III tubulin at DIVs 3, 7, 14, and 21.45–50 Spheroids also expressed the neuronal progenitor marker nestin at DIVs 3 and 7, but nestin expression was significantly reduced after DIV 14 (Fig. 4B). 45 Glutamatergic receptor 2 (GLUR2), a neuronal marker found in SGNs, and vesicular glutamate transporter 2 (VGLUT2), a marker specific to glutamatergic neurons, showed significantly low expression at DIV 3, but high expression after DIV 7.51,52 Finally, a neurite arborization assay showed significantly less axonal branching at DIV 3 compared with DIVs 7, 14, and 21 (Fig. 4C).
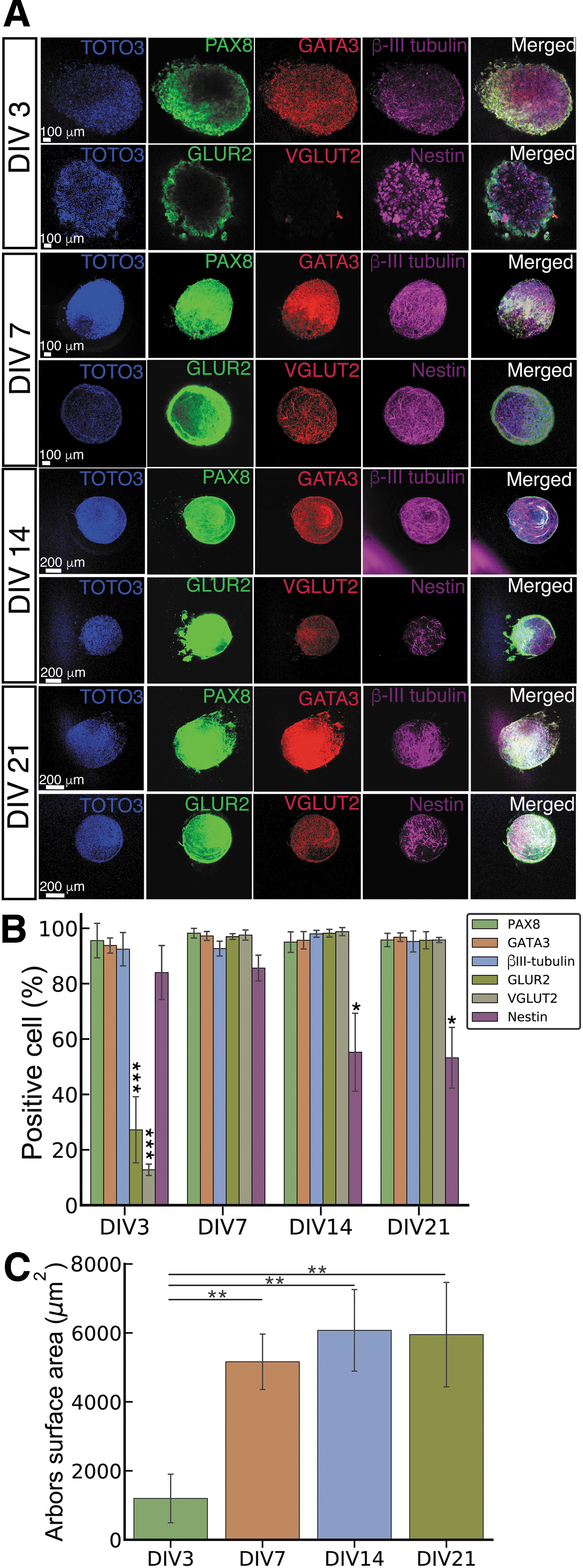
Immunocytochemical analysis of hESC-derived ONP spheroids.
Whole-cell patch-clamp recordings
Whole-cell patch-clamp recordings were performed to investigate the spheroids' electrophysiological properties. Average resting membrane potentials in DIV 5 and 7 ONP spheroids (mean ± SD) were −19.0 ± 7.9 mV (n = 7) and −28.5 ± 7.8 (n = 8), respectively. While there were virtually no current activities in whole-cell voltage-clamp recordings on DIV 5 (Fig. 5Aa), DIV 7 spheroids showed voltage-activated current efflux above −20 mV (Fig. 5Bb), indicative of a large voltage-gated potassium (Kv) channel activity. Consistently, whole-cell current-clamp recording in DIV 7 spheroids showed clearly detectable transmembrane potential (Vm) repolarization after current-induced cell depolarization (Fig. 5Bc), an essential electrophysiological characteristic found in all types of neurons. In contrast, DIV 5 spheroids indicated no signs of Vm repolarization (Fig. 5Ac).

Electrophysiological responses and properties of hESC-derived ONP spheroids.
Mechanical properties of hESC-derived ONP spheroids
Figure 6A shows a representative force curve obtained from an hESC-derived ONP spheroid using the Piuma nanoindenter. Note the smoothing effect of the Hertzian curve fit to the raw data. Note also that the indentation depth used in this study was kept below 10 μm to avoid detecting the substrate on which the spheroids were resting.21,35 As shown in Figure 6B, elastic Young's modulus increased over time, but did not increase at equal rates for the two culture plates. The Young's modulus of spheroids from both plates at DIV 4 is not significantly different from that of the mouse brain control tissue. By DIV 8, the spheroids formed in U-bottom plates had a Young's modulus significantly greater compared with the mouse brain tissue and were stiff enough to hold up to the stress of mock transplant. In contrast, the spheroids formed in an EZSPHERE plate took until DIV 11 to reach this point; at DIV 8, the elastic Young's modulus of U-bottom spheroids is significantly greater compared with the EZSPHERE spheroids.
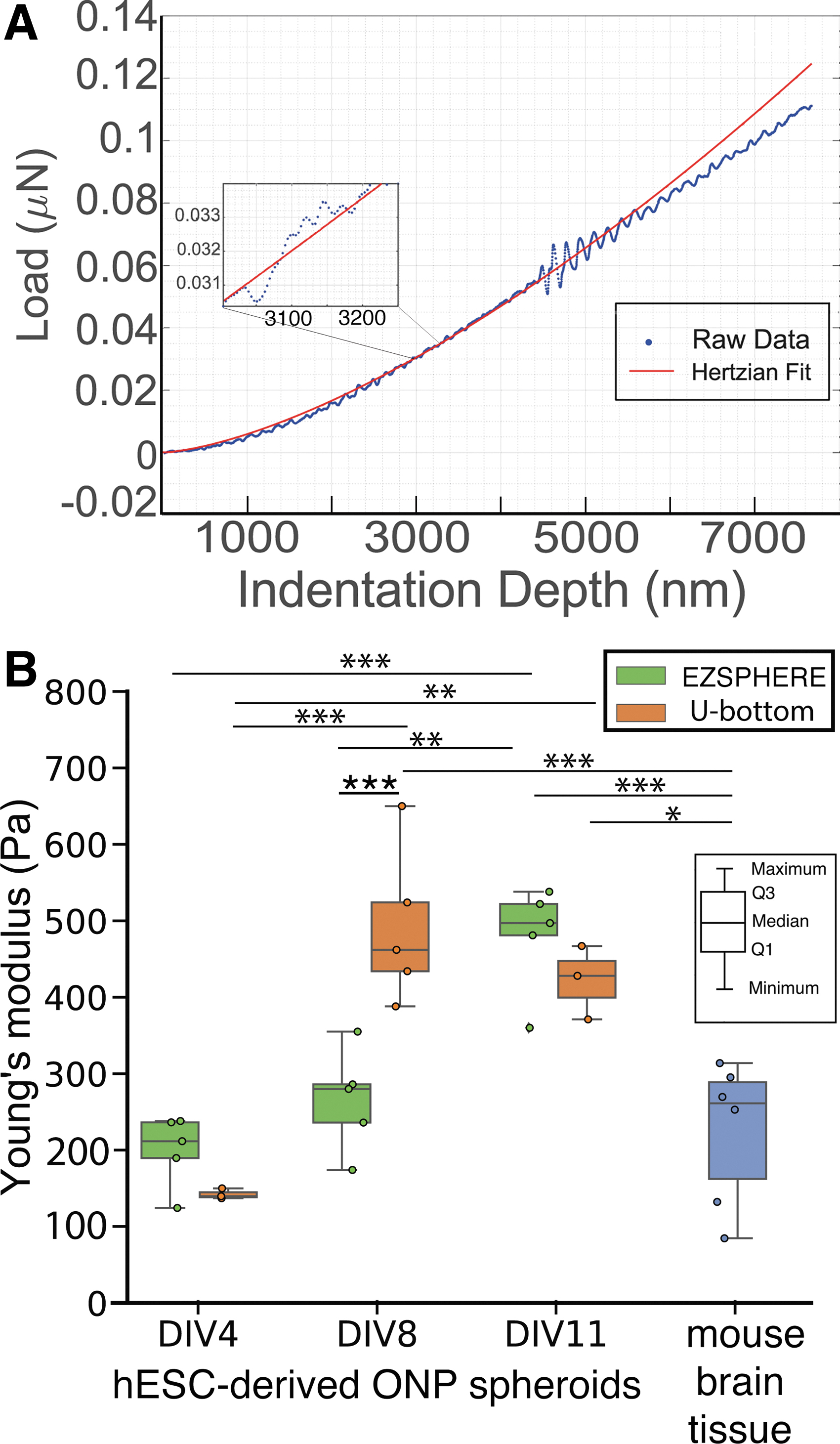
Mechanical properties of hESC-derived ONP spheroids.
Simulated transplantation of ONP spheroids using a pressure-based delivery system
Our micropipette transfer system is depicted in Figure 7, which shows spheroids held on or within a micropipette by means of negative pressure (ΔP = 5 hPa). Using this system, mock transplantation was performed with a number of spheroids and a range of micropipette diameters. Micropipettes were chosen with inner diameters (ID) covering a range from 10% to 85% of the diameters of the spheroids (based on values used for in vitro fertilization).53,54 (Supplementary Figs. S3 and S4 show the photomicrographical images used to determine this range.) Figure 8A shows the pickup and release pressures necessary to capture and transfer spheroids; the pressure changes required decrease as micropipette ID increases (from 175 to 385 μm) (Supplementary Fig. S5 for a pilot study supporting this trend). The pressure data in Figure 8B indicate that the closer the size of the micropipette to the size of the spheroid, the less negative pressure required to capture the spheroid and the less positive pressure required for release. A nonlinear monoexponential curve fit was performed on the pressure data (Fig. 8B, green line). The decay constant (

Micropipette apparatus for simulated transplant of spheroids. Negative pressure holds hESC-derived ONP spheroids at the tip of the glass micropipette (see white arrows for the spheroid location).

Simulated transplant procedure for hESC-derived ONP spheroids and analysis of cell survival.
With this trend in mind, cell survival was analyzed for spheroids in each of three “zones”: Zone I spheroids were transferred with micropipette of IDs 10–30% of the spheroid diameters, Zone II with micropipettes of ID 30–50% of spheroid diameters, and Zone III with micropipettes of ID 50–85% of spheroid diameters. All spheroids showed some proportion of dead cells (Fig. 8C), but for Zone I and II spheroids, dead cell populations were clustered particularly around the point of contact of the pipette (Fig. 8C, white dotted lines: the third column in Zone I and II). Cell survival was assessed as the ratio of live cell area compared with total cell area (Fig. 8D). In Figure 8D, the SD of live cell area in Zone I is greater compared with Zone II and III. To verify this difference statistically, a Shapiro–Wilk normality test was performed first to confirm the normal distribution of these data (p = 0.987 [Zone I], 0.845 [Zone II], and 0.876 [Zone III], respectively). After confirming the normal distribution, we performed F-tests between Zone I and Zone II, and Zone I and Zone III. The p-value of the F-test indicated p < 0.05 (0.015 for Zone I and II, 0.012 for Zone I and III), demonstrating that the SD in Zone I was statistically larger compared with Zone II or Zone III. Finally, one-way ANOVA indicated no statistical difference in the mean among Zones I–III (p > 0.05).
Discussion
Our data demonstrate that spheroid culture of hESC-derived ONPs is an advantageous system to create cell aggregates suitable for transplant by micropipette. Spheroids retained 3D form over the culture period. However, cell viability patterns altered over time: younger spheroids contained some apoptotic cells around the edges, and apoptosis increased with time, with apoptotic cells distributed uniformly (in a nonconcentric fashion) in older spheroids. Previous studies that performed TUNEL staining on 3D spheroids mostly demonstrated a concentric configuration of apoptotic cells, with center cell death presumed to occur due to hypoxia and nutrient deprivation in the spheroid core.55,56 It is conceivable that the duration of our culture period may not be long enough to show this concentric apoptosis pattern. In addition, the high viability of the spheroids could be due to high biocompatibility of our spheroid-generating platforms. Although cell death was not extensive enough to destroy the spheroids' 3D configuration, future studies should explore methods to increase cell viability further throughout the spheroids for extended culture periods.
Immunocytochemistry on the 3D spheroid culture showed that component cells expressed markers specific to the ONP stage (the precursor to SGNs) by DIV 7: β-III tubulin, GLUR2, and VGLUT2. Spheroids also showed continuous high expression of the otic markers GATA3 and PAX8. Our previous study demonstrated that hESC-derived ONPs did not express S-100 (a Schwann cell marker), PAX6 (a marker for the central nervous system), E-cadherin, or myosin VIIA (hair cell markers), indicating guidance toward the otic lineage. 5 Nestin, the only observed marker to drop significantly in expression over the culture period, is a neural marker found most often in progenitor cells.45,57 Its decrease in expression indicates that 3D culture promotes continued ONP differentiation from the progenitor stage. Additionally, our neurite arborization assay shows maximized neurite growth by DIV 7, indicating that spheroids should be cultured until at least DIV 7 before transplant to maximize expression of these specific neuronal progenitor markers.
The whole-cell voltage-/current-clamp recordings showed early signs of functional development in our ONP spheroids. Although spheroids at DIV 5 showed little ion channel activity, by DIV 7 a large voltage-dependent outward current was clearly apparent, indicating functional Kv channel expression. Additionally, the transmembrane potential repolarization following current-induced cell depolarization seen in DIV 7 spheroids may be an early sign of neuronal maturation, although this can be also seen in a variety of cells in animals. 58 The repolarization was not present in spheroids at DIV 5, demonstrating the need to allow additional time for neuronal development. We acknowledge that the ONP spheroids generated in this study were functionally not fully matured; improved resting membrane potentials (−50 to −60 mV) were found in some ONP cells at DIV 10 or later, but none showed significant inward currents upon cell depolarization. Previous studies that investigated electrophysiological properties of hESC-derived SGNs have shown that cells at least later than DIV 10 showed voltage-activated inward currents and/or action potentials.7,47,59,60 We, therefore, speculate that these activities can be promoted by extended culture, which should be explored in future study.
Spheroids seeded in both culture platforms, U-bottom and EZSPHERE, increased in stiffness over time, but not at the same rate. As early as DIV 4, all spheroids had Young's moduli similar to that of mouse brain and were too fragile to hold up to the stress of mock transplantation. By DIV 8, U-bottom spheroids were stiff enough for transplantation, and EZSPHERE spheroids by DIV 11. We conclude that both systems are sufficient to form spheroids robust enough to hold up to transplantation, but U-bottom spheroids would allow us to perform a transplantation procedure more quickly. If, however, large-scale production of spheroids is required, the EZSPHERE platform is preferable, since it does not require time-consuming seeding of individual spheroids.
The increase in elastic modulus over time may be correlated with changing cell morphology within the 3D structure. For instance, cell stiffness and motility are regulated by cytoskeletal structures such as F-actin filaments; it is therefore possible that F-actin filaments on the spheroid surface contribute to the increasing stiffness.21,61 Future study can explore this further by cytoskeletal F-actin imaging. An improved understanding of the mechanisms responsible for mechanical changes over time may provide new insight into approaches for chemically targeting cytoskeletal structures that regulate cell stiffness and motility. 61
Our observations of elastic Young's moduli of hESC-derived ONP spheroids were consistent with previously published data using microtweezers or atomic force microscopy.21,62 It is worthwhile mentioning that a different Poisson's ratio (e.g., 0.3) would change the estimation of Young's modulus (e.g., increasing by 21%); however, our purpose in this study was to demonstrate relative differences in the Young's moduli rather than absolute values. We used a Poisson's ratio of 0.5 based on a previous study.62–65 Additionally, previous studies indicate that measurements of the elasticity of biological samples by different methods—for example, optical tweezers or rheometers—have quite a wide range of variance.62,66,67 Although the variance may arise from differences in cell culture conditions, the particular geometry of the probe used and the amount of mechanical stress put on the cells in each experimental method play a significant role. 66 Thus, it should be noted that absolute values between experiments are rarely directly comparable.66–68
Our mock transplantation procedure assessed the damage done to ONP spheroids during transfer. All spheroids exhibited a proportion of dead surface cells after micropipette manipulation. Smaller micropipettes showed larger variations in proportions of living cells, however, indicating lower reliability for transplantation. Additionally, smaller micropipettes required a vast pressure differential to capture and release spheroids, creating significantly more stress and leading to a greater degree of spheroid deformity. It can therefore be concluded that micropipette tips with sizes similar to the spheroids themselves are most suitable for use. The Sutter Xenowork system we used in this study has been widely used for somatic cell nuclear transfer and intracytoplasmic sperm injection, but to our knowledge, this is the first study that used glass micropipettes in conjunction with a microinjector for stem cell spheroid transfer. 69 This work thus constitutes an intermediate step toward the development of in vivo transplantation procedures for hESC-derived 3D aggregates.
We found that DIV 7 was the optimal time point for our in vivo spheroid transplantation. Neurite arborization reached a maximum at this point and did not statistically differ thereafter. In addition, the elastic properties of our spheroids indicated that U-bottom spheroids were stiff enough to support transplantation after DIV 7 (and EZSPHERE spheroids by DIV 11). Furthermore, using our curve fit equation (Fig. 2C), it took 7 days for our hESC-derived ONP spheroids to approach a diameter of 500–700 μm—the diameter of the round window in the mouse. 70 Based on these results, we conclude that it is most suitable to transplant our hESC-derived ONP spheroids to the inner ear at or after DIV 7. Our curve-fitted spheroid diameter model (Fig. 2C), in addition to our mock transplant data (Fig. 8), can be used to identify suitable seeding density, culture period, and micropipette size for optimal transplant of hESC-derived ONP spheroids to the inner ear in future studies. This information can be utilized by the auditory research community for further development of improved stem cell replacement therapies.
Several findings of this study warrant further discussion. First, this study solely focused on hESC-derived ONP spheroids. Stem cells can also be obtained from autologous (i.e., mesenchymal stem cells or inner ear stem cells) or allogeneic sources (i.e., mouse embryonic stem cells). The results we reported in this study are only applicable for hPSCs, and further investigations will be needed for other types of cells. Second, while our TUNEL viability assay demonstrated that apoptosis occurred in a nonconcentric fashion over time, further investigations are needed to evaluate the status of the core of the spheroids and determine whether cell death was due to necrosis or programmed apoptosis. Third, the data on mechanical properties should be viewed with discretion, as testing was performed solely with a nanoindenter; future experiments will use a second method to verify our results.
Taken together, our data describe a comprehensive system for developing hESC-derived ONP spheroids and optimizing their transplant. Our cells self-aggregate into spheroids easily and show the characteristics of neuronal progenitors; these characteristics are most apparent after at least 7 days of spheroid culture. Moreover, our transplantation protocol is successful in transferring these cells as aggregates with minimal destruction of the spheroids. To the best of our knowledge, this is the first study to investigate the transplantation of 3D cell aggregates—spheroids or organoids—using a micropipette aided by a digital microinjector. Although preliminary, our findings indicate that these hESC-derived ONP spheroids are a favorable candidate for inner ear transplantation therapy to aid in the regeneration of SGNs and mitigate the effect of SNHL.
Footnotes
Acknowledgments
The authors thank Peter Dluhy, Constadina Arvanitis, PhD, David Kirchenbuechler, PhD, and Wensheng Liu, MD at the Northwestern University Center for Advanced Microscopy for assistance with confocal microscopy. Chemical and biological analysis was performed in the Analytical bioNanoTechnology Core Facility (ANTEC) of the Simpson Querrey Institute (SQI) at Northwestern University. ANTEC is currently supported by the Soft and Hybrid Nanotechnology Experimental (SHyNE) Resource (NSFECCS-1542205). They also would like to thank Alexandra Kolot, Charles Bressan (ANTEC/SQI, Northwestern University), and Jason Luo, PhD (Optics11) for assistance in their rheological and viscoelastic experiment. Finally, the authors would like to also acknowledge Chris Pernstich, PhD (Cell Guidance System), Tony Kiuru, MS, Markus Nuopponen, PhD, Lauri Paasonen, PhD (UPM Biofore), Machi Muto, PhD (Nacalai, USA), Tatsuaki Miwa, PhD, and Hiroko Tsukamoto, PhD (AGC Techno Glass) for insightful comments and suggestions on this article.
Disclosure Statement
This article has not been considered to be published in another journal. The authors have no competing and financial interest.
Funding Information
This work was supported by the American Otological Society Clinician Scientist Award (A.J.M.), the Triological Society/American College of Surgeons Clinician Scientist Award (A.J.M.), the Department of Otolaryngology, Feinberg School of Medicine, Northwestern University (A.J.M.), NIH K08 Clinician Scientist Award K08DC13829–02 (A.J.M.), and the Office of the Assistant Secretary of Defense of Health Affairs through the Hearing Restoration Research Program (Award #: RH170013:WU81XWUH-18-0712).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.