
Abstract
In knee osteoarthritis (OA), there is more pronounced cartilage damage in the medial compartment (“lesion zone”) than the lateral compartment (“remote zone”). This study fills a gap in the literature by conducting a systematic comparison of cartilage and chondrocyte characteristics from these two zones. It also investigates whether chondrocytes from the different zones respond distinctly to changes in the physical and mechanical microenvironment using three-dimensional porous scaffolds by changing stiffness and pore size. Cartilage was harvested from patients with end-stage varus knee OA. Cartilage from the lesion and remote zones were compared through histological and biomechanical assessments, and through proteomic and gene transcription analyses of chondrocytes. Gelatin scaffolds with varied pore sizes and stiffness were used to investigate in vitro microenvironmental regulation of chondrocytes from the two zones. Cartilage from the lesion and remote zones differed significantly (p < 0.05) in histological and biomechanical characteristics, as well as phenotype, protein, and gene expression of chondrocytes. Chondrocytes from both zones were sensitive to changes in the structural and mechanical properties of gelatin scaffolds. Of interest, although all chondrocytes better retained chondrocyte phenotype in stiffer scaffolds, those from the lesion and remote zones, respectively, preferred scaffolds with larger and smaller pores. Distinct variations exist in cartilage and chondrocyte characteristics in the lesion and remote zones of knee OA. Cells in these two zones respond differently to variations in the physical and mechanical microenvironment. Understanding and manipulating these differences will facilitate the development of more efficient and precise diagnostic and therapeutic approaches for knee OA.
Impact statement
This study performs a novel systematic analysis of cartilage properties in the lesion (medial) and remote (lateral) zones of knee osteoarthritis (OA), from the tissue level right down to the cell and molecular levels. It also demonstrates for the first time the effects of manipulating the physical and mechanical microenvironment on the behavior of chondrocytes from the lesion and remote zones. This study improves our understanding of the properties and responses of tissues and cells in different zones of knee OA, providing novel insights into OA pathophysiology and also demonstrating a new application of biomaterial scaffolds in studying cell–matrix interactions.
Introduction
The medial compartment of the varus joint is the most frequently affected area in knee osteoarthritis (OA). Varus knee thrust is associated with increased imbalance in joint loading in the medial knee compartment.1,2 A normal loading regime is known to be essential for the development and maintenance of articular cartilage, owing to the profound effects of mechanical loading on the composition and structure of cartilaginous extracellular matrix (ECM). 3 In varus knee OA, the mechanical loading regime causes cartilage damage in the “lesion zone” (medial compartment adjacent to osteoarthritic cartilage) to be much more severe than that in the “remote zone” (lateral compartment distant from osteoarthritic cartilage). This is evidenced by lower glycosaminoglycan (GAG), collagen type II, and chondroitin sulfate content in the medial side of femoral and tibial cartilage compared with the lateral side.4–6
The ECM is a critical and unique component of all tissues that not only facilitates cell adhesion and intercellular communication,7,8 but also imparts physical and mechanical characteristics such as pore size, stiffness, and deformability that play vital roles in directing cell survival, proliferation, and differentiation.9–11 The ECM is a dynamic structure that undergoes constant remodeling to control tissue homeostasis. 12 In healthy articular cartilage, the unique composition and complex structure of the ECM allow the tissue to withstand cyclic loading deformation throughout life. During normal cartilage loading, water in this highly hydrated tissue is gradually extruded out and reabsorbed into the ECM, leading to complex and uneven deformation fields at the cell and tissue levels.13,14
Altered cartilage loading either as a cause or result of knee OA introduces complex biomechanical changes that in turn may affect cell and tissue responses to joint load on macroscopic and microscopic levels. 15 In an interesting study that generated a numerical model of early-stage OA, microscale spatial softening of the ECM resulted in a substantial increase (30%) of chondrocyte shear strain with no apparent changes in macroscale tissue properties. 16 The need to use these numerical models to study cell behavior in healthy and osteoarthritic cartilage was mainly because of the inability to effectively decouple the influence of structural and mechanical properties in artificially synthesized scaffolds. With the latest advances in biomaterials processing technologies, we were able to fabricate gelatin scaffolds that decouple the complex interplay between the scaffold's pore size and stiffness, allowing each of these properties to be studied in isolation in terms of their effects on directing cell behavior. 11
Although there are some known differences in the macroscopic and microscopic properties of cartilage in the lesion and remote zones (or medial and lateral compartments) of knee OA, previous studies have mainly focused on the comparison of discrete aspects, such as only the macroscopic biomechanical and histological properties of cartilage tissue, or only the microscopic ECM composition or chondrocyte gene expression profile.4–6 Systematic analysis of both macroscopic and microscopic properties of cartilage in the lesion and remote zones of knee OA is still lacking. Furthermore, proteomic analysis and RNA sequencing of chondrocytes isolated from these different zones in the same patient have not been performed.
Our study addresses these identified knowledge gaps by comparing the lesion and remote zones in knee OA from the tissue level down to the cellular and molecular (protein and gene) levels. We also study, for the first time, the effects of ECM pore size and stiffness on the behavior of chondrocytes from the lesion and remote zones using an in vitro scaffold platform. Improving the understanding of these relationships, including the properties of tissues and cells in different cartilage regions, and how cell behavior in these different regions are differentially affected by structural and mechanical cues in the microenvironment, will provide novel insights into OA pathophysiology and enable the development of more effective and precise diagnostic and therapeutic tools.
Materials and Methods
All reagents were obtained from Thermo Fisher Scientific, unless otherwise stated.
Patients and samples
This research was approved by the Ethics Review Committee of Peking University People's Hospital, China (2019PHB098-01). Discarded cartilage tissues were obtained from patients undergoing total knee replacement surgery for the treatment of knee OA with varus deformity. All clinical specimens were taken after patients gave informed consent and agreed to have their data used for research purposes, including gender, age, diagnosis, OA grade, and surgical site. The clinical specimens were obtained from patients at age 68 ± 6.48 years (mean ± standard deviation [SD]) who were clinically diagnosed to be Kellgren–Lawrence grade 3 based on radiographic examination (Supplementary Table S1). Patients were excluded if they had valgus knee OA, infected arthritis, or rheumatoid arthritis. For each specimen, cartilage tissue was harvested from different regions of knee OA based on the International Cartilage Repair Society (ICRS) score (Supplementary Table S2). “Lesion zone” cartilage was harvested from the medial tibial plateau and had ICRS grade 2 or 3 (abnormal to severely abnormal), whereas “remote zone” cartilage was harvested from the lateral tibial plateau and had ICRS grade 1 (nearly normal).
Biomechanical testing
Cartilage tissue from three patients was used for biomechanical testing. The harvested cartilage tissue was stored in 0.9% sodium chloride saline solution, and biomechanical testing was conducted within 1 h from the time of harvest. Two osteochondral plugs (5 cm in diameter) were, respectively, taken from the lesion and remote zones of each specimen using an autograft transfer tool. A central region of cartilage (20 × 20 mm) from each osteochondral plug was selected for compressive modulus mapping using a ferrule-top nanoindenter together with the PIUMA controller/drive (Optics11 Life, Amsterdam, the Netherlands). The ferrule-top indenter probe was glued to a small iron part, which was hooked to a magnet anchored to a coarse z-positioner. The tissue sample was fixed on a closed-loop z-piezoelectric translator, equipped with an integrated strain gauge sensor that was controlled by a closed-loop controller. An area was sensed by screwing the z-piezoelectric translator in the xy directions, which was controlled by the software. This method allowed the measurement of reduced Young's modulus. A cantilever with a 1 μm probe was used for the indentation experiments, and the deflection of the cantilever during indentation was measured by Fabry–Pérot interferometry. The contact area of the indentation tip with the tissue sample had a diameter of <5 μm. For each measurement, 10 single indents were performed on the same spot of the sample. All indents were depth controlled with a maximum indentation of 10 μm, and the loading and unloading was set to 2 s. Before the loading and after the unloading step, the data were collected for 1 s each. The starting point (deformation = 0 nm) of the recording was determined optically using a photodiode aligned with the fourth port of the coupler. The load–displacement curves were used to determine the Young's modulus of cartilage in the lesion and remote zones. 17
Histology and immunofluorescent staining
Cartilage tissue from 18 patients was used for histological assessment. The cartilage samples were fixed in 4% paraformaldehyde for 12–24 h, followed by decalcification in 20% ethylenediaminetetraacetic acid (EDTA) solution for 2 months. The decalcified samples were embedded in paraffin and cut into 5 μm thick sections using a microtome (Leica, Cambridge, United Kingdom). A total of 10 sections were obtained for each cartilage sample, which were stained with hematoxylin and eosin (H&E) and safranin O/fast green, and imaged using a digital slide scanner (Pannoramic DESK/MIDI/SCAN/250; 3DHISTECH, Hungary). Immunofluorescent staining was performed using mouse antihuman collagen type II (ab185430; Abcam) and rabbit antihuman collagen type I (ab34710; Abcam), and imaged using confocal laser scanning microscopy (LSM 710 Meta; Zeiss, Germany). The percentage area of collagen type II and type I expression in cartilage from the lesion and remote zones were quantitatively assessed using ImageJ software (National Institutes of Health).
Safranin O-stained sections were scored using the ICRS II evaluation system, 18 and more detailed methods have been included in the Supplementary Data.
Chondrocyte isolation and culture
Cartilage samples from the lesion and remote zones of each clinical specimen were, respectively, harvested from the grossly intact areas of the medial and lateral tibial plateaus. Chondrocytes from the lesion and remote zones were isolated from the cartilage by sequential enzymatic digestion with 0.2% collagenase type II (No.1392433; Gibco) for 10 h at 37°C, as previously described. 19 Isolated chondrocytes were washed in phosphate-buffered saline (PBS), strained using 100 μm nylon meshes, and seeded at high density (1 × 105 cells/well) in six-well plates. Chondrocytes were maintained in Dulbecco's modified Eagle's medium (DMEM) high glucose supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin–streptomycin, 10 mM HEPES, and 1% minimum Eagle's medium nonessential amino acids solution, 20 until further analysis.
Analysis of chondrocyte characteristics
The characteristics of chondrocytes isolated from the lesion and remote zones of cartilage were analyzed after 3 days in culture. The deposition of collagen types I and II by chondrocytes (from three clinical specimens) was visualized and quantified through immunofluorescence staining as described in “Biomechanical testing” section. Chondrocyte diameters (50–60 cells from each of six clinical specimens) were measured using ImageJ after imaging the cells through a counting plate.
Chondrocytes (from four clinical specimens) were analyzed by flow cytometry. The cells were digested with EDTA, collected and treated as follows: 4% paraformaldehyde for 15 min; PBS containing 0.5% Triton X-100 for 10 min; PBS containing 5% bovine serum albumin for 1 h; PBS with the primary antibodies mouse monoclonal collagen type II (Abcam; 1:400) and rabbit monoclonal collagen type I (Abcam; 1:200); and PBS with the secondary antibodies Alexa Fluor-549 goat anti-mouse immunoglobulin G (IgG) and Alexa Fluor 488 goat anti-rabbit IgG. Finally, more than 1 × 104 cells were analyzed for fluorescence intensity using flow cytometry (LSRFortessa SORP; BD Biosciences). For cell size verification, chondrocytes digested with EDTA were directly identified by flow cytometry.
Proteomic analysis
Chondrocytes from three patients were used for proteomic analysis and experiments were performed in biological triplicate. In brief, chondrocytes from the lesion and remote zones of each specimen were lysed by ultrasonication. An in-solution trypsin digestion protocol was used to generate peptides for liquid chromatography–tandem mass spectrometry analysis. More detailed methods have been included in the Supplementary Data.
RNA-seq analysis
RNA-seq analysis was conducted on lesion and remote chondrocytes isolated from three patients, using 2 μg of total RNA for each sample. RNA sequencing was conducted by Anoroad Gene Technology Corporation (Beijing, China). Differential expression genes were identified by DESeq2 package, and functional enrichment for Gene Ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) were performed with GOstats package. More detailed methods have been included in the Supplementary Data.
Fabrication and characterization of gelatin scaffolds
Gelatin-based porous scaffolds were prepared as described in our previous study, 11 to give four scaffold groups differing in pore size (30 or 80 μm) and stiffness (20 or 200 kPa). In brief, 4% w/v gelatin precursor solution was prepared by dissolving gelatin powder in deionized water, mixed with two concentrations of dimethylsulfoxide (DMSO) as a cryoprotectant (1% and 5% v/v; shown to give average pore sizes of 30 and 80 μm, respectively, in our previous study 11 ). Two different concentrations of glutaraldehyde (0.1% and 0.4% w/v; shown to give average stiffness of 20 and 200 kPa, respectively, in our previous study 11 ) were then added to the premixed solutions and stirred for 20 s. For each of the four scaffold types, gelatin-based sheets (20 × 20 × 1 mm3) were made by placing 400 μL of solution in a polymethyl methacrylate mould, which was then allowed to undergo cryogelation for 16 h at −20°C. After cryogelation, scaffold sheets were treated with 1% sodium borohydride solution (pH 9.7) to remove residual glutaraldehyde, and washed with deionized water before being lyophilized for 2 h. Scaffold pieces (4 × 4 × 1 mm3) were cut out from the sheet for use in subsequent cell culture experiments.
The prepared scaffolds were coated with gold and imaged using scanning electron microscopy (SEM; FEI Quanta 200) to evaluate their morphology. The pore size distribution for each of the four different scaffold types was measured using 12 SEM images with ImageJ software. Stiffness of the scaffold types was not reevaluated in this study as this property was found to be reliably controlled by glutaraldehyde concentration in our previous study. 11
Chondrocyte culture in gelatin scaffolds
Gelatin scaffolds were sterilized under ultraviolet radiation for 2 h before cell seeding. Chondrocytes for the lesion and remote zones were pooled from six clinical specimens and seeded in the four different scaffold types (n = 8 per time point). Cells were washed, counted, centrifuged, and resuspended in DMEM supplemented with 10% FBS and 2% penicillin–streptomycin. Cells were seeded into gelatin scaffolds (4 × 4 × 1 mm3) at a final concentration of 5 × 104 cells per scaffold in 48-well plates, and were allowed to attach for 2 h at 37°C. Finally, 500 μL of growth medium was added to each well and seeded scaffolds were cultured at 37°C with 5% CO2 for up to 9 days, with medium changed every 3 days.
Viability and proliferation of chondrocytes in gelatin scaffolds
At 1, 3, 6, and 9 days, the viability (n = 4) and proliferation (n = 4) of chondrocytes in each scaffold type was tested using a Calcein-AM/PI kit (BAOLIBO, China) and Click-iT EdU Cell Proliferation Kit (Thermo Fisher Scientific), respectively, according to the manufacturers' instructions. The cell viability ratio [l/(d + l)%] was calculated from the number of red dead cells (d) and green living cells (l) counted using ImageJ from four separate regions for each sample. The proliferation rate of cells was calculated by visualizing the stained cells using a fluorescence microscope (Nikon, Japan), counting the cells using ImageJ from four separate regions for each sample, and normalizing to the area of the scaffold.
Gene expression of chondrocytes in gelatin scaffolds
Chondrocytes for the lesion and remote zones were pooled from three clinical specimens and seeded in the four different scaffold types (n = 4 per time point). Cells were seeded and cultured on gelatin scaffolds for 3, 6, and 9 days using the same procedures as described in “Biomechanical testing” section. At each time point, total RNA was isolated from the cells using the TRIzol reagent (Invitrogen) according to the manufacturer's instructions. In brief, after homogenizing the sample with TRIzol reagent, chloroform was added and RNA was precipitated from the upper aqueous layer using isopropanol. The RNA was washed and resuspended in 20 μL purified water.
cDNA was synthesized from 600 ng total RNA by reverse transcription using a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) according to the manufacturer's instructions. Expression levels of all genes were quantified using a real-time RT-PCR system (ABI 7300; Applied Biosystems) and normalized to GAPDH (primer sequences from Ruirui Boxing Kesheng Company, Beijing; Supplementary Table S3) using the comparative Ct (2−ΔΔCT) method.
Histology and immunofluorescence of chondrocytes in gelatin scaffolds
Chondrocytes for the lesion and remote zones were pooled from three clinical specimens and seeded in the four scaffold types (n = 3). Cells were seeded and cultured on gelatin scaffolds for 9 days using the same procedures as described in “Fabrication and characterization of gelatin scaffolds” section. For histology, the scaffolds were harvested and embedded in optimum cutting temperature embedding medium for frozen tissue (Sakura Finetek), and frozen at −80°C for 4 h before being fixed in 4% paraformaldehyde for 12 h. Each sample was cut using a cryostat (Leica CM1520, Germany) to generate three sections with a thickness of 8 μm. The sections were stained using Safranin O/fast green and imaged using the same process as described in “Histology and immunofluorescent staining” section. The amount of red staining was taken to represent aggrecan deposition and was quantified using ImageJ for six regions from each of the three sections for each sample. For immunofluorescence, scaffolds were fixed in 4% paraformaldehyde for 12 h, followed by immunofluorescence staining for collagen type II using the same process as described in “Histology and immunofluorescent staining” section. The amount of collagen type II deposition was quantified using ImageJ for six regions on the surface layer of each sample.
Statistical analysis
All data were expressed as mean ± SD. Statistical analysis was performed using GraphPad Prism. Significant differences between groups were checked using unpaired, two-tailed Student's t-test to compare two groups of data, or one-way analysis of variance with Tukey's multiple comparisons test to compare three or more groups of data. Values of p < 0.05 were considered statistically significant.
Results
Macroscopic, histological, and biomechanical characteristics of osteoarthritic cartilage in different zones
Macroscopic observations of cartilage specimens from patients with varus knee OA showed that cartilage had more pronounced damage in the lesion zone compared with the remote zone (Fig. 1A). Lesion zone cartilage sustained a much smaller indentation depth during biomechanical testing (Fig. 1B, left), and had significantly lower Young's modulus (150 ± 50 kPa) compared with remote zone cartilage (1637 ± 138 kPa) (p < 0.0001) (Fig. 1B, right). The lower biomechanical properties of lesion zone cartilage are suggestive of more advanced ECM degradation compared with the remote zone. Histological staining showed worse cartilage quality in the lesion zone than in the remote zone, as indicated by lower GAG content through safranin O staining, and significant surface damage and fibrillation through H&E staining (Fig. 1C, left). The ICRS II score was significantly lower (p < 0.0001) for the lesion zone (55 ± 3) compared with the remote zone (84 ± 2) (Fig. 1C, right). Qualitative and quantitative analysis of collagen expression by immunofluorescence showed that the collagen type II/type I ratio was significantly lower in the lesion zone compared with the remote zone (p < 0.0001) (Fig. 1D). Collectively, cartilage from the lesion and remote zones of varus knee OA display vastly different macroscopic, histological, and biomechanical properties suggestive of much more severe damage in the lesion zone.
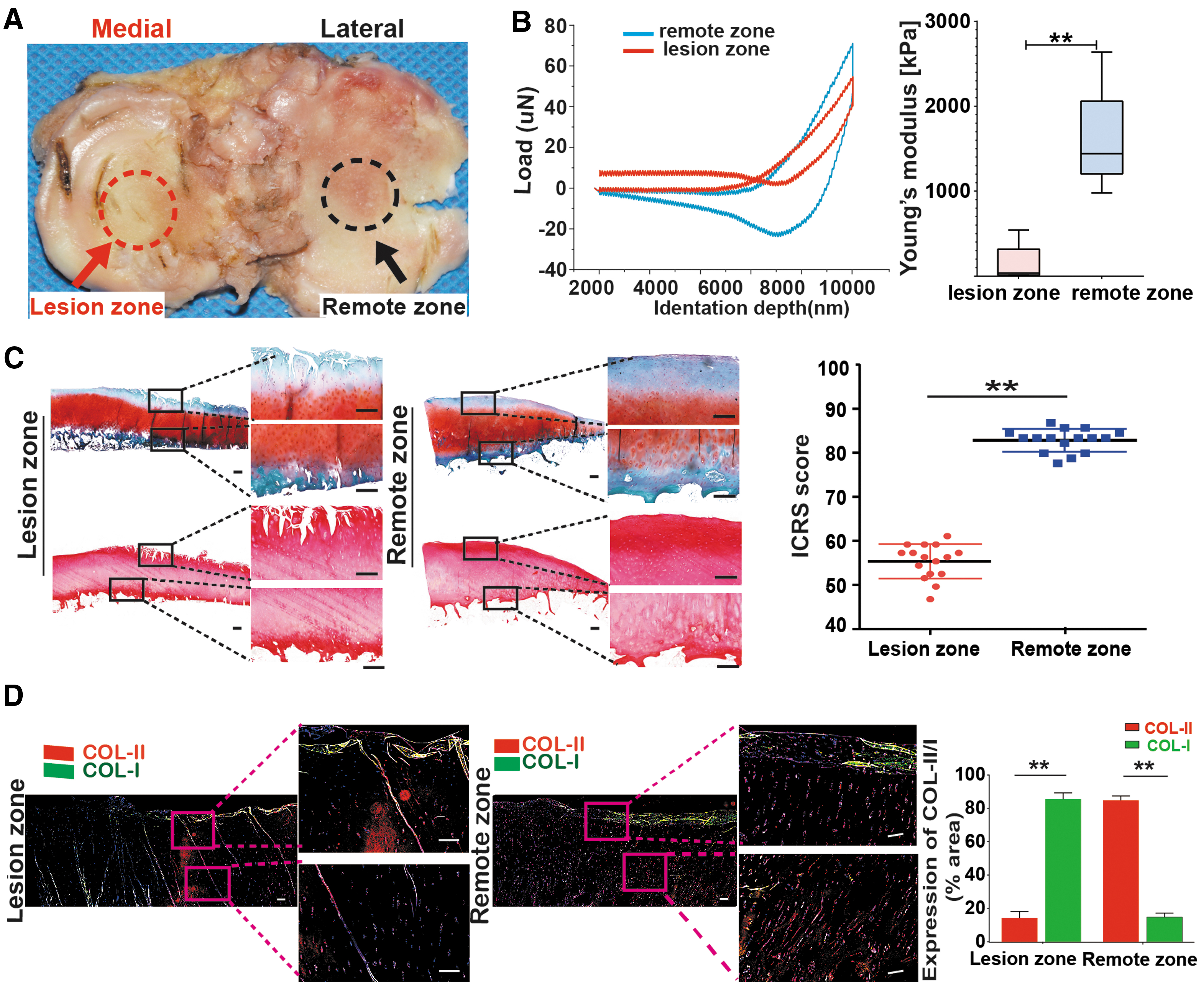
Macroscopic, histological, and biomechanical characteristics of cartilage in different zones in knee OA.
Chondrocyte characteristics of osteoarthritic cartilage in different zones
Immunofluorescence staining of chondrocytes isolated from osteoarthritic cartilage showed significantly higher collagen type I than type II expression in the lesion zone (19.3% and 78.2%, respectively), whereas the reverse trend was observed for chondrocytes in the remote zone (79.2% and 19.7%, respectively) (Fig. 2A). Flow cytometry analysis showed a ratio of ∼1:5 for collagen type II/type I expressing chondrocytes in the lesion zone, compared with a ratio of ∼4:1 in the remote zone (Fig. 2B). The average size of chondrocytes from the lesion zone (32.6 ± 8.4 μm) was larger than those in the remote zone (26.1 ± 5.9 μm), as quantified through both image analysis (Fig. 2C) and flow cytometry analysis for cell size (Fig. 2D). Collectively, chondrocytes from the lesion and remote zones of varus knee OA display significant differences that indicate greater hypertrophy and loss of the chondrocyte phenotype in the lesion zone, because increased cell size is a hallmark of chondrocyte hypertrophy.21,22
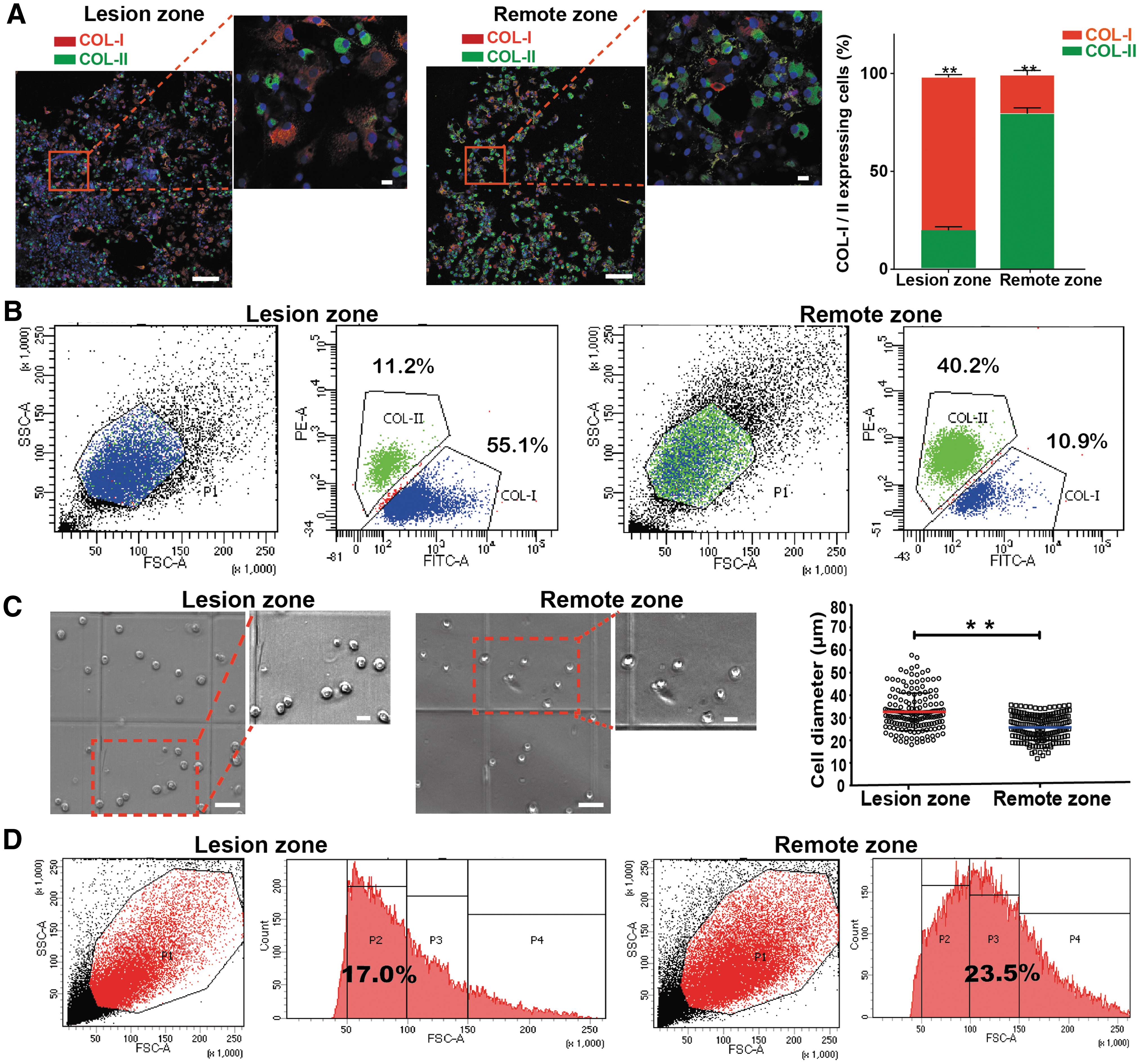
Chondrocyte characteristics in different damage zones in knee OA.
Proteomic analysis of chondrocytes from different damage zones
Differentially expressed proteins in chondrocytes from two regions in osteoarthritic cartilage (lesion zone and remote zone) were identified and quantified, with the false-positive rate (false discovery rate) set to be <1%. For the samples from three patients used for proteomic analysis, 1844 proteins were found to be differentially expressed between chondrocytes from the lesion and remote zones, of which 780 proteins were upregulated and 1064 were downregulated (Fig. 3A). Some proteins were differentially expressed between lesion and remote zone chondrocytes in two or more patients (Fig. 3B). Of these proteins, the majority were associated with the ECM and exosomes. The upregulated proteins were also associated with the endoplasmic reticulum and negative regulation of endopeptidase activity, whereas the downregulated proteins were also associated with the perinuclear region of cytoplasm and negative regulation of growth (Fig. 3C). The highly expressed proteins may contribute to callus formation, providing a new direction for further exploration of osteophyte formation in late-stage knee OA. The functional classification of differentially expressed proteins in at least one patient was analyzed by DAVID Bioinformatics Resources 6.8 (Fig. 3D), and more detailed results are included in the Supplementary Data (Supplementary Fig. S1).

Proteomic analysis of chondrocytes from different zones in knee OA (n = 3). Protein expression in lesion chondrocytes was compared with the expression in remote chondrocytes (control).
Proteomic cluster analysis of upregulated proteins in lesion zone chondrocytes from at least one patient revealed proteins associated with other disorders such as Parkinson's disease, Huntington's disease, and Alzheimer's disease (Supplementary Table S4). These proteins may provide pathways for investigating the mechanisms underlying the observations made in another study, where OA was found to be associated with the risk for Alzheimer's disease and related dementias by using cross-sectional data from a representative sample of US adults 65 years or older. 23 The complete list of differentially expressed proteins between the lesion and remote zone chondrocytes in all three patients is given in the Supplementary Data (Supplementary Table S5).
RNA-seq analysis
Differentially expressed genes in lesion and remote zone chondrocytes from osteoarthritic cartilage were found to be mostly associated with the ECM and corroborated the proteomic analysis. These results are included in the Supplementary Data (Supplementary Fig. S2). The complete list of differentially expressed RNA between the lesion and remote zone chondrocytes in at least two of three patients is given in the Supplementary Data (Supplementary Table S6).
In vitro responses of osteoarthritic chondrocytes from different zones to microenvironmental cues provided by porous gelatin scaffolds
Viability and proliferation of chondrocytes in gelatin scaffolds with different pore size and stiffness
The pore size and stiffness of porous gelatin scaffolds could be reliably controlled by adding different concentrations of DMSO and glutaraldehyde, respectively, during the fabrication process. 11 Using this method, four types of gelatin scaffolds with different pore sizes and stiffness were fabricated: 30 μm/20 kPa, 30 μm/200 kPa, 80 μm/20 kPa, and 80 μm/200 kPa. The average pore size for all scaffold groups were verified by SEM (Fig. 4A).
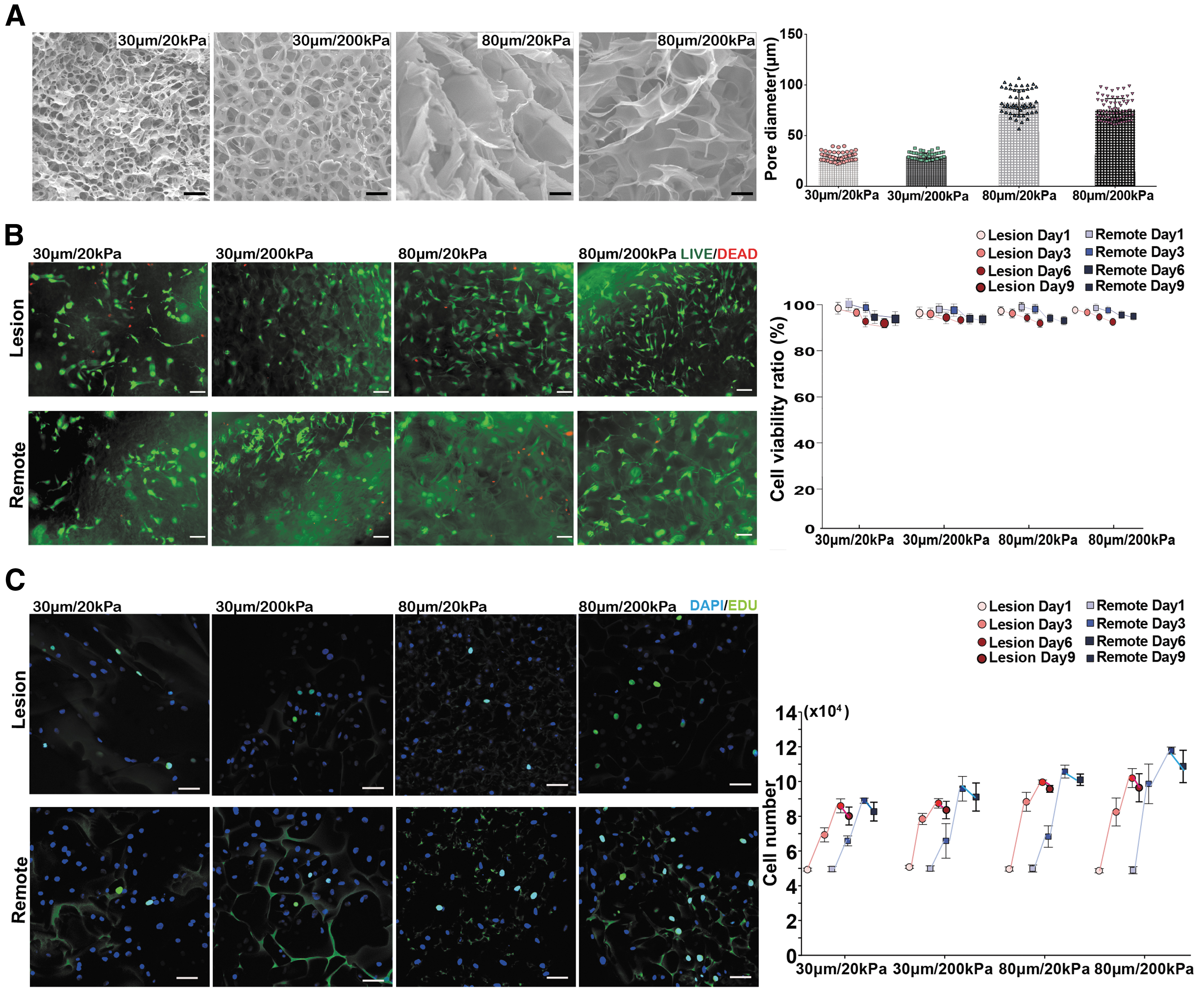
Viability and proliferation of lesion and remote zone chondrocytes in gelatin scaffolds with different pore size and stiffness.
Chondrocytes from the lesion and remote zones of osteoarthritic cartilage were cultured in the four types of gelatin scaffolds for up to 9 days. The viability of chondrocytes from the lesion and remote zones as indicated by live/dead staining was similar at all time points, and remained >92% in all scaffold types at 9 days (Fig. 4B). Chondrocyte proliferation as indicated by 5-ethynyl-2′-deoxyuridine (EdU) staining showed that cells from both lesion and remote zones increased in number in all scaffold types until day 6, and reached a plateau between 6 and 9 days (Fig. 4C). Of interest, although the remote zone chondrocytes either showed a steady proliferation rate or faster proliferation between 3 and 6 days in different scaffold types, the lesion zone chondrocytes showed faster proliferation during the first 3 days and a declined proliferation rate thereafter. At 6 and 9 days, the remote zone chondrocytes showed similar or higher cell numbers compared with the lesion zone chondrocytes in all scaffold types. Although the proliferation rate was not greatly affected by scaffold stiffness, both lesion and remote zone chondrocytes showed greater increases in cell number when grown in scaffolds with larger (80 μm) pore sizes.
Gene expression of chondrocytes in gelatin scaffolds with different pore size and stiffness
Gene expression analysis of lesion and remote zone chondrocytes cultured in the four types of gelatin scaffolds showed some interesting trends. Lesion zone chondrocytes seemed to favor gelatin scaffolds with larger pore sizes, as indicated by pronounced upregulation of markers associated with maintenance of the chondrocyte phenotype (AGG, SOX9, and COL-II) in the 80 μm scaffold groups, particularly at 6 and 9 days with the most significant increases observed at 9 days (Fig. 5A). Lesion zone chondrocytes also showed downregulation of markers associated with osteoarthritic chondrocytes (COL-I, TIMP1, and MMP13) in the 80 μm scaffold groups. Here, TIMP1 is taken to be associated with genes implicated in cartilage catabolism because of its strong coupling with MMP13 activity, being a functional regulator of MMP13-associated chondrocyte senescence and cartilage matrix aging. 24 In comparison, the remote zone chondrocytes were less sensitive to changes in scaffold pore size, with less significant differences between groups at all time points (Fig. 5B). Different from the lesion zone chondrocytes, the remote zone chondrocytes showed more upregulation of AGG, SOX9, and COL-II and downregulation of COL-I, TIMP1, and MMP13 in scaffolds with smaller pore sizes of 30 μm. Both lesion and remote zone chondrocytes were less sensitive to changes in scaffold stiffness compared with changes in pore size, although both appeared to have better maintenance of a “normal” chondrocyte phenotype in stiffer scaffolds with 200 kPa modulus. These trends were more easily observed by computing the ratio of gene expression between days 9 and 3 for all genes tested (Fig. 5C). The lesion and remote zone chondrocytes, respectively, retained their chondrocyte phenotype better in scaffolds with larger and smaller pore sizes, as shown through the relatively higher ratio of expression of AGG, SOX9, and COL-II compared with COL-I, TIMP1, and MMP13.
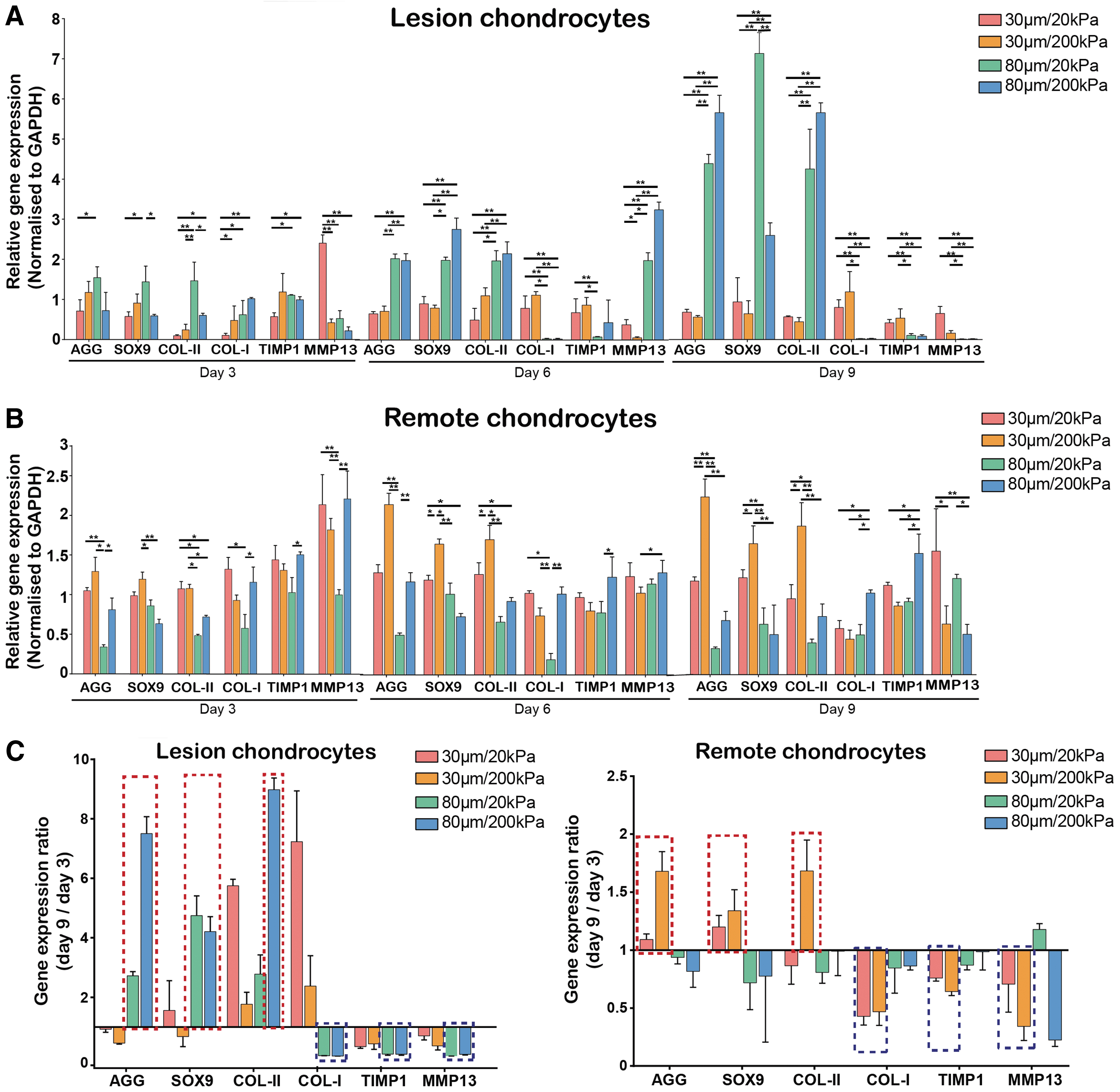
Gene expression of lesion and remote zone chondrocytes in gelatin scaffolds with different pore size and stiffness. Chondrocytes from the
Histology and immunofluorescence of chondrocytes in gelatin scaffolds with different pore size and stiffness
Safranin O staining of the scaffold sections showed that chondrocytes infiltrated and grew throughout the depth of the gelatin scaffolds. For lesion zone chondrocytes, more aggrecan deposition was seen in the 80 μm scaffold groups (Fig. 6A, left), with the highest deposition in the 80 μm/200 kPa scaffolds (Fig. 6A, right). The remote zone chondrocytes showed more aggrecan deposition in the 30 μm scaffold groups (Fig. 6A, left) and the highest deposition in the 30 μm/200 kPa scaffolds (Fig. 6A, right). Collagen type II deposition followed the same trend, with the lesion chondrocytes favoring scaffolds with larger pore sizes and remote chondrocytes favoring scaffolds with smaller pore sizes (Fig. 6B). Both lesion and remote zone chondrocytes were less sensitive to changes in scaffold stiffness, although the highest aggrecan and collagen type II deposition were observed in the stiffer (200 kPa) scaffolds. These results corroborated the trends seen through gene expression analysis.
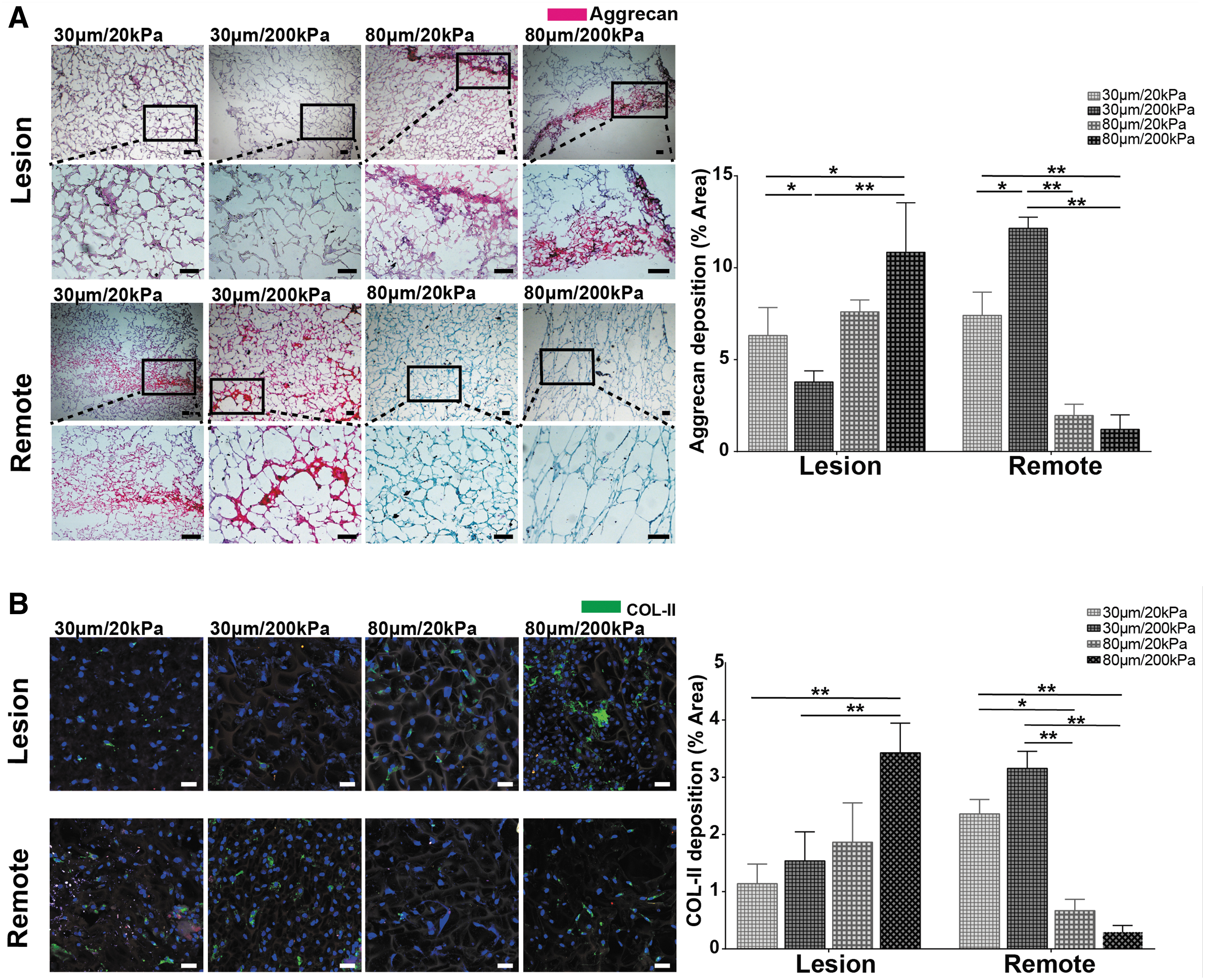
Histology and immunofluorescence of lesion and remote zone chondrocytes in gelatin scaffolds with different pore size and stiffness.
Discussion
OA is a multifactorial disease that involves a whole-joint pathological response to a range of intrinsic and extrinsic factors. Abnormal mechanical loading is currently understood to be a primary factor contributing to OA pathogenesis and progression. Altered joint biomechanics can have intertwined macroscopic and microscopic effects on articular cartilage. Disruption of the normal porosity and fluid flow within articular cartilage because of abnormal loading can cause substantial surface wear owing to inadequate lubrication, leading to degeneration of the superficial zone of articular cartilage that is a common feature of early OA. 25 In response to these degenerative changes, the cartilage matrix undergoes extensive biophysical remodeling, often resulting in matrix stiffening that further contributes to OA pathogenesis. 26 Chondrocytes can respond to these abnormal ECM changes by shifting toward catabolism and undergoing hypertrophy, which contribute to initiating a cascade of events that result in permanent cartilage damage. 27 In this study, we examined osteoarthritic cartilage and chondrocytes harvested from patients with varus knee undergoing joint replacement procedures, and found much more severe features of OA in the lesion zone (region primarily affected by abnormal mechanical stress) compared with the remote zone. Lesion zone cartilage displayed properties indicative of a loss of structural and mechanical integrity (Fig. 1), whereas the chondrocytes showed evidence of hypertrophy and a loss of chondrocyte phenotype (Fig. 2), which is consistent with the degenerative changes associated with abnormal joint loading.
Proteomic analysis of lesion zone chondrocytes revealed trends in the upregulation of proteins associated with ECM organization and interactions, and downregulation of proteins associated with mineral absorption and growth regulation compared with remote zone chondrocytes (Fig. 3). These trends match the RNA-seq data and reflect the extensive ECM remodeling seen in advanced OA. In particular, the upregulation of proteins associated with ECM–receptor interactions and focal adhesions in lesion chondrocytes suggests that the mechanical loading environment, which in turn determines the mechanical stresses experienced by the matrix, plays a major role in influencing cell-mediated matrix remodeling in osteoarthritic cartilage. The proteomic and RNA-seq analyses partly corroborated the results of other studies comparing protein28,29 and gene expression30,31 between the medial/osteoarthritic region and lateral/nonosteoarthritic region in human knees. For instance, similar to our findings, these studies showed downregulation of certain chondrogenic proteins (collagen XI and aggrecan) and genes (SOX9, COL2A1, and ACAN), and upregulation of certain nonchondrogenic proteins (collagen VI) and genes (COL1A1, COL1A2, RUNX2, and MMP3) in osteoarthritic compared with nonosteoarthritic chondrocytes. Other interesting associations arising from the proteomic analysis are the increased expression of proteins related to Parkinson's disease, Huntington's disease, Alzheimer's disease, and oxidative phosphorylation in lesion zone chondrocytes (Supplementary Table S4), suggesting that certain pathways in severe OA may concomitantly affect brain function and neuropsychiatric symptoms. These observations corroborate with studies suggesting that people with knee OA are more likely to have depression, 32 perceived memory loss, 33 and suicidal ideation. 34 The results from this study provide pointers to novel pathways that warrant investigation in future studies.
Articular cartilage is a highly specialized connective tissue with a unique ECM that fulfils essential structural and biomechanical functions to enable normal joint motion. 35 It is known that dysregulation of the composition, structure, stiffness, and abundance of cartilage ECM contributes to OA pathogenesis. However, the processes by which ECM remodeling affects disease progression and how these processes can be artificially controlled are much less well understood. 36 Part of the reason for this is the inability to effectively decouple the influences of pore size and stiffness in artificial scaffold systems, as traditional fabrication processes typically cause simultaneous changes in both parameters. For instance, stiffness adjustment of collagen scaffolds is traditionally achieved by changing the concentration of collagen. However, a higher collagen concentration results in denser and differently arranged fibers, which not only lead to higher scaffold stiffness but also smaller pores. Using these scaffolds as a model system for cell culture can make it difficult to determine whether variations in cellular responses were because of changes in scaffold stiffness or pore size. 37 The gelatin scaffolds we previously developed and subsequently used in this study address these limitations, allowing cell responses in scaffolds with different pore sizes and stiffness to be compared while only varying one parameter and not the other. 11 This novel scaffold system allowed us to demonstrate the responses of OA chondrocytes in microenvironments with independent variations in pore size and stiffness while still using the same base material for consistency, which was previously impossible using other artificial scaffolds. Of interest, although the remote zone chondrocytes proliferated at a steady rate in all scaffold types, the lesion zone chondrocytes proliferated much faster during the first few days of culture (Fig. 4). In addition, the ratio of gene expression from days 3 to 9 for lesion chondrocytes showed much greater magnitude for all genes compared with remote chondrocytes (Fig. 5C), indicating greater changes in gene expression levels over time. These results possibly suggest that the pathological phenotype of the lesion chondrocytes makes them more responsive to microenvironmental changes, and are in alignment with evidence pointing to a central role of matrix-mediated mechanotransduction in OA pathogenesis. 26
Analysis of gene expression, histology, and immunofluorescence for cells cultured in different types of gelatin scaffolds indicated that the lesion zone chondrocytes upregulated their anabolic activities and downregulated catabolic activities in scaffolds with larger (80 μm) pore sizes, whereas the remote zone chondrocytes did the same in scaffolds with smaller (30 μm) pore sizes (Figs. 5 and 6). One possible explanation is that the lesion chondrocytes were mostly in a hypertrophic state with large diameters exceeding 30 μm, which needed to be grown in a matrix with larger pores to allow better adherence and satisfaction of their metabolic requirements. On the contrary, the remote chondrocytes preferred smaller pore sizes that more closely resembled the tight spacing in chondrocyte lacunae, suggesting that their behavior was more similar to normal, quiescent chondrocytes with low metabolic activity. By this explanation, the lesion and remote chondrocytes could, respectively, achieve more complete spreading in scaffolds with larger and smaller pore sizes, which would better activate cell surface receptors associated with matrix sensing, leading to downstream intracellular signaling to maintain cell survival and activity. Of interest, although changes in the responses of lesion and remote chondrocytes to scaffolds of different pore sizes were quite obvious, their responses to variations in scaffold stiffness were more subtle, which may indicate that matrix porosity is a more important regulator of chondrocyte behavior in OA. Nevertheless, it was noted that both lesion and remote chondrocytes exhibited gene expression behavior suggesting better maintenance of the chondrocyte phenotype on stiffer (200 kPa) scaffolds. This is in contradiction with other studies in the literature suggesting that chondrogenesis and the chondrogenic phenotype are better maintained on softer (1–5 kPa) rather than harder (100–150 kPa) matrices.38,39 Given that substrates with stiffness ∼1 MPa can promote the expression of both chondrogenic and osteogenic markers, 40 it is possible that both lesion and remote zone chondrocytes were undergoing hypertrophic changes owing to OA, although to different degrees, and therefore preferred growth on stiffer substrates. Understanding and deciphering the responses of diseased chondrocytes from different zones in knee OA to changing physical and mechanical cues in the microenvironment constitute an important future research direction for OA pathophysiology.
A primary limitation of this study was that because of practical difficulties in obtaining sufficient cartilage samples and numbers of chondrocytes from individual patients, different experiments were performed using cartilage or chondrocytes harvested from a different pool of donors. It is possible that donor variability exists in the clinical specimens obtained that may have confounded the findings between experiments. However, we attempted to minimize this variability by selecting cartilage samples to constitute the lesion and remote zones based on ICRS scoring, and by randomly assigning samples for different analyses. It would also have been relevant to compare the findings in this study with the responses of healthy human articular chondrocytes, which will be explored in future studies. Another limitation was that the chondrocytes were briefly maintained in culture (<3 days) before being used for further analysis, which may have caused dedifferentiation and some loss of phenotypic characteristics. To minimize the possible influences of in vitro culture on the maintenance of chondrocyte phenotype and behavior, we conducted a relatively short-term study of 9 days using the gelatin scaffolds. However, a longer term study might allow more comprehensive analysis of chondrocyte behavior and whether phenotypic drift of chondrocytes would occur on scaffolds with different pore sizes and stiffness, which will be explored in future studies. Finally, the proteomic and RNA-seq analyses only each resembled the expression profiles of three patients, and more samples may be necessary to confirm the trends that were observed in two or less patients. Nevertheless, the number of samples used were on the same order as other studies that performed similar types of global protein 28 and gene 30 expression analyses to compare the expression profiles of osteoarthritic and nonosteoarthritic chondrocytes.
Conclusions
In this study, we systematically demonstrated that there were distinct differences in macroscopic, structural, and biomechanical properties between cartilage from the lesion and remote zones of knee OA. Chondrocytes from these different zones had different protein and RNA signatures, and were sensitive to changes in the physical and mechanical microenvironment. Lesion and remote zone chondrocytes cultured on gelatin scaffolds, respectively, showed better maintenance of phenotype on scaffolds with larger and smaller pore sizes, whereas stiffer scaffolds were preferred by chondrocytes from both zones. Understanding these cell–matrix interactions in OA may underpin the development of new diagnostic and therapeutic tools.
Footnotes
Authors' Contributions
Conception and design: Y.Z., Z.Y., Y.D., and J.L. Analysis and interpretation of the data: all authors. Drafting of the article: Y.Z., Z.Y., and J.J.L. Critical revision of the article for important intellectual content: all authors. Final approval of the article: all authors. Provision of study materials or patients: Y.D. and J.L. Statistical expertise: Y.Z. and D.X. Obtaining of funding: Y.D. and J.L. Collection and assembly of data: Y.Z., Z.Y., D.X., J.J.L., Q.Z., H.H., Z.L., S.J., Z.W., Y.Z., W.L. and L.Z.
Acknowledgments
The authors gratefully acknowledge Peng Zhao and Cheng Lyu for their assistance in preparing the gelatin scaffolds, as well as Beijing CytoNiche Biotechnology Ltd. for providing technical support. The authors also thank Dr. Jialu Ma, Huizhen Cao and Yue Sun from the Center of Biomedical Analysis at Tsinghua University for technical assistance, and all members from the Du-lab for general experimental assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (81973606, 81802204, and 81501919), Peking University People's Hospital Scientific Research Development Funds (RDY2019-11 and RDH2017-05), Beijing Science and Technology Major Projects (Z181100001818008), and the Australian National Health and Medical Research Council (APP1120249).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.