
Abstract
The manipulation of human chondrocyte sheets in target areas frequently results in their tearing because they are thin and fragile. In this study, human cancellous bones were used as a supporting material to create chondrocyte sheet-cancellous bone tissues, and their properties were evaluated. Using cell sheet technology, human chondrocytes were constructed into triple-layered chondrocyte sheets that displayed chondrogenic properties. After transferring the chondrocyte sheets onto cancellous bones, the top area of the chondrocyte sheet-cancellous bone tissues exhibited a smooth surface topography without cell sheet floating within 7 days of culture. The immunofluorescence staining of collagen type II (COL2A1) and fibronectin (FN1) was also performed and examined. Using the shotgun proteomic analysis, the proteins associated with cell adhesion, extracellular matrix (ECM) organization, cell–substrate junction assembly, and cell adhesion mediated by integrin were observed in the chondrocyte sheets, cancellous bones, and chondrocyte sheet-cancellous bone tissues. Three integrin members, including integrin β4 (ITGB4), ITGB6, and ITGB8, were found in the chondrocyte sheets. Only ITGB8 was found in the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues. During 48 h, the mean velocity of the individual cell migration was low, which did not affect the structure and chondrogenic properties of the chondrocyte sheets. Staining of the filamentous actin (F-actin) cytoskeleton in the migratory cells also provided a better understanding of the dynamic communication between the cell cytoskeleton and adhesion molecules through ITGB8, which may play a key role in the attachment of the chondrocyte sheets and the synthesis of the cartilage ECM. Therefore, we suggest that cancellous bone could be used as a supporting material to construct chondrocyte sheet-cancellous bone tissues for potential treatment of osteochondral lesions.
Impact Statement
We proposed a method to construct an osteochondral-like tissue by placing human chondrocyte sheets onto cancellous bone. The stationary chondrocyte sheets and the low mean velocity of the individual cell migration on the cancellous bone with the expression of COL2A1 indicated that the cancellous bone served as an appropriate supporting material. Moreover, the cellular mechanism for the adhesion of the chondrocyte sheets on the cancellous bone based on ITGB8-mediated adhesion through the rearrangement of filamentous actin provided a better understanding to improve the construction of osteochondral-like tissues, and to predict the repair mechanism in osteoarthritis therapy.
Introduction
Tissue engineering and regenerative medicine are promising alternative approaches to repairing articular cartilage defects, which can ultimately progress to osteoarthritis. 1 Articular cartilage damages can be categorized into chondral (partial and full thickness lesions) and osteochondral defects. Chondral defects resemble clefts found only in the articular cartilage layer, which has a low capacity for self-repair because of the limited vascularity, whereas in osteochondral defects, the clefts pass through the zone of the calcified cartilage and disturb the subchondral bone. 2
Currently, available treatments by microfracture mostly result in the formation of fibrocartilage, while the replacement of damaged cartilage by the mosaicplasty technique limits the supply of healthy cartilage donors. 3 To fulfill these unmet medical needs, tissue engineering strategies have now been developed to reconstruct cartilage tissues by means of scaffold-based tissue engineering combined with cells and bioactive molecules. 4 Various biodegradable scaffolds have been reported as having the potential for use in clinical trials, such as collagen, chitosan, and polyglycolic acid. 5 However, the degradation time of these scaffolds and release of chemical by-products require optimization in each treatment. 5 Moreover, the handling of cells to promote cell attachment and their functions on the scaffolds are limited because only a small percentage of cells are located in the target areas. 4
Among the possible tissue engineering approaches, cell sheet engineering has recently gained increasing attention, as it can overcome the problems of single-cell seeding and the use of biodegradable scaffolds. 6 Based on the techniques of cell sheet engineering, cells seeded on temperature-responsive polymers can grow as monolayer cells at 37°C. These cells create their own extracellular matrix (ECM) and adhesion factors, such as fibronectin (FN1) and integrins with tight cell-to-cell junctions. By incubating the temperature-responsive culture surface at below 32°C, the cell layer is simply harvested as an intact sheet without using enzymatic treatment. This harvested cell sheet with its own ECM can then be layered either on another cell sheet to create a multilayered sheet or on a substrate to generate a cell-to-substrate connection. Presently, cell sheet-based engineering has been combined with available surgery to improve the clinical outcomes. 7 However, the manipulation of the cell sheet and the relationship between the cell sheet and supporting material during manipulation have been rarely reported.
In our previous work, cell sheet engineering was applied to create a triple-layered human chondrocyte sheet. Chondrocytes were the only cell type found in the articular cartilage and comprised only about 5% of the total volume of the cartilage, mainly containing water, collagen type II (COL2A1), and proteoglycans. 8 We demonstrated that chondrocyte sheet manipulation both with and without the use of a gelatin manipulator generated changes in the stress fiber formation and the expression of COL2A1. The chondrocyte sheets harvested without the gelatin manipulator exhibited suppressed stress fiber formation with higher levels of COL2A1 compared with cell sheets harvested using the gelatin manipulator. Although cell shrinkage occurred when a gelatin manipulator was not applied, the chondrocyte sheets exhibited cartilage-specific genes and proteins, which had chondrogenic potential to regenerate the cartilage defects.9,10
Therefore, to the best of our knowledge, a chondrocyte sheet was very fragile and required the use of a gelatin manipulator as a supporting material with more time required to remove the gelatin from the cell sheet. Moreover, adequate incubation time of the cell sheet on a new substrate is strongly required. 10 A polyvinylidene fluoride (PVDF) membrane, as a commercially available material used for transferring cell sheets, could be used to support the cell sheets, but the chondrocyte sheets were prone to sudden shrinkage and tear when the PVDF membrane was removed.
We, therefore, proposed the use of natural materials that could preserve the cell-to-cell junctions and introduce cell-to-substrate connections without removing the material and supporting the chondrocyte sheet manipulation. One of the natural materials typically used in orthopedic surgery is cancellous bone, which has a large trabecular surface area. 5 Cancellous bone has been routinely used to infill bone defects in patients to provide fibrovascular ingrowth and integration at the recipient site. 11 The degradation behavior of cancellous bone is related to natural bone remodeling, and the release of natural by-products can be eliminated naturally from the body. Generally, cancellous bone can be prepared as fresh, deep-frozen, and freeze-dried cancellous bones. 12 Unlike fresh and deep-frozen cancellous bone allografts, the freeze-dried cancellous bone allograft has no cellular content and it has lowest immunogenicity with a low risk of disease transmission. 13 Therefore, the use of freeze-dried cancellous bone allograft has been recommended for use in many orthopedic procedures. 14
In this study, we aimed to evaluate the use of human chondrocyte sheets combined with freeze-dried human cancellous bone allograft to construct tissue-engineered cartilage by observing the cell attachment, cell growth, and expression of the chondrogenic markers. The molecular mechanisms involved in the chondrocyte sheet adhesion to cancellous bone by means of dynamic communication of filamentous actin (F-actin) cytoskeleton, FN1, and integrin were also discussed.
Materials and Methods
Human osteoarthritic cartilage and chondrocytes
Human osteoarthritic cartilage was obtained from patients aged between 70 and 80 years old, who underwent total knee replacement in Siriraj Hospital, Bangkok, Thailand. All procedures using human cartilage were approved by the Siriraj Institutional Review Board (COA No. Si429/2019). The cartilage fragments were washed in Hank's Balanced Salt Solution (HBSS) containing 1% antibiotic-antimycotics (Gibco, Grand Island, NY), and they were chopped into small pieces. Following the enzymatic treatment, the cartilages were incubated in a digestion medium: low glucose Dulbecco's modified Eagle's medium (DMEM) containing 1.5 mg/mL of collagenase II (Gibco) and 1% antibiotic-antimycotics (1 mL per 100 mg of cartilage) on a magnetic stirrer at 80 rpm. After being incubated overnight, the digested chondrocyte solution was collected by centrifugation at 1,500 rpm for 5 min, and the cells were repeatedly washed with HBSS containing 1% antibiotic-antimycotics. Afterward, the chondrocytes were cultured in a growth medium: DMEM supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% antibiotic-antimycotics at 37°C in a humidified CO2 incubator. The freshly isolated chondrocytes were cultured in a T225 flask (Corning, Inc., NY), and the growth medium was changed every 3–4 days.
Human cancellous bone
Human cancellous bones were provided by the Bangkok Biomaterial Center under the Patronage of H.R.H. Princess Galyani Vadhana Krom Luang Naradhiwas Rajanagarindra, Faculty of Medicine, Siriraj Hospital. After collecting the cancellous bone from the donors, the bones were kept in a quarantine chamber at −70°C until used. Following the advanced tissue banking guideline, the cancellous bones were ultrasonicated in double biofiltered water to remove the bone marrow and fat. After that, the cancellous bones were immersed in hydrogen peroxide to clean the bone and remove any red blood cells. To inactivate the human immunodeficiency virus and hepatitis B virus, the decellularized cancellous bones were submerged in 0.5% sodium hypochlorite solution for 2 h. Then, the cancellous bones were subjected to a freeze-drying process at −50°C for 72 h, before being sterilized by gamma radiation.
Mercury intrusion porosimetry
The porosity and pore size distribution of the freeze-dried cancellous bones were measured by means of mercury intrusion porosimetry (MIP; AutoPore V, Micromeritics, Norcross, GA). The cancellous bones were prepared as a cylindrical shape (diameter and length ∼5 mm; n = 3), and they were incubated in a hot air oven at 40°C for 15 h. The MIP measurement was carried out with the intrusion pressure ranging from 0.10 to 1,000 psia.
Preparation of the chondrocyte sheets and chondrocyte sheets attached to freeze-dried cancellous bone
The human chondrocytes were expanded on the T225 flasks, and passage 2 (P2) chondrocytes were used to construct the cell sheets, as previously described. 10 In brief, the chondrocytes were seeded as cells per area at a density of 6 × 105 cells/cm2 on the FBS-coated 24-well temperature-responsive plates (CellSeed, Tokyo, Japan). The cells were cultured in a growth medium at 37°C in a humidified CO2 incubator for 24 h to form monolayer cells. To create the chondrocyte sheets, the chondrocytes on the temperature-responsive surface were incubated with the serum-free medium at 20°C for 30 min. Next, a monolayer cell sheet was spontaneously detached by slight agitation of the serum-free medium with a plastic tip. Afterward, the detached chondrocyte sheet was transferred onto a new tissue culture polystyrene (TCPS) dish, and it was incubated at 37°C for 30 min to allow the attachment of the cell sheet on the new culture's surface. By repeating these processes, the triple-layered chondrocyte sheets were obtained. Finally, the triple-layered chondrocyte sheets were incubated at 37°C for another 1 h to further promote the attachment between each layered chondrocyte sheet.
In this study, the human cancellous bones were cut into a cylindrical shape (diameter = 5 mm; height = 3 mm) using a surgical trepan bur and saw with a micromotor (MicroNX 300D, Daegu, Korea). The cancellous bones were washed with HBSS to remove any bone dust, and they were then incubated overnight in a growth medium to remove any air bubbles. To create chondrocyte sheet-cancellous bone tissue, the triple-layered chondrocyte sheets were raised from the TCPS dish by aspirating the edge of the triple-layered sheets with the serum-free medium. After that, the floated cell sheet was carefully placed on the cancellous bone using tweezers. Next, the chondrocyte sheet-cancellous bone tissue was subjected to 2 h of preincubation at 37°C by adding an ∼300 μL of the growth medium every 30 min to prevent the cell sheets from drying and floating up. Then, the chondrocyte sheet-cancellous bone tissue was transferred into a cell culture insert set in a 24-well plate (Falcon). The plate was then incubated at 37°C in a humidified CO2 incubator for 7 days, and the growth medium was refreshed every 3–4 days.
Low-vacuum scanning electron microscope
To observe the morphology of the chondrocyte sheets adhered on the cancellous bones, the cancellous bones with and without triple-layered chondrocyte sheets at day 7 were imaged using a low-vacuum scanning electron microscope (TM4000Plus; Hitachi High-Technologies, Tokyo, Japan). The chondrocyte sheet-cancellous bone tissues were fixed with 4% paraformaldehyde for 15 min and washed with phosphate buffered saline (PBS) before the cell morphology was observed. The element composition of the freeze-dried cancellous bone was determined using TM series energy dispersive X-ray spectrometer (EDS; Bruker Nano GmbH, Germany).
Immunofluorescence analysis of the frozen tissue sections
Triple-layered chondrocyte sheets on either the TCPS or cancellous bones were subjected to a frozen section process after 7 days of culture. After fixation in 4% paraformaldehyde for 10 min, the chondrocyte sheet-cancellous bone tissues underwent decalcification in Osteosoft® (Merck Millipore, Billerica, MA) at room temperature for 3 weeks. Immunofluorescence staining of the cryosections was performed to observe the expression of collagen type I (COL1A1), COL2A1, FN1, and integrin β1 (ITGB1). The sections were fixed with 4% paraformaldehyde and incubated with 0.5% polyoxyethylene (10) octylphenyl ether in PBS for permeabilization. Nonspecific binding proteins were reduced in a Block Ace solution (Dainippon Sumitomo Pharma Co., Ltd., Osaka, Japan) for 90 min. The sections were incubated with the primary antibody for COL1A1, COL2A1, FN1, and ITGB1 (Santa Cruz Biotechnology, CA) at 4°C overnight. After washing the sample slides with tris-buffered saline (TBS), a solution of the secondary antibody (Alexa Fluor 488 donkey anti-goat IgG, Molecular Probes, OR or Mouse IgG kappa binding protein conjugated to CruzFluor™ 488, Santa Cruz Biotechnology) was added, and they were then incubated for 1 h. Another incubation was conducted for 40 min with 4′,6-diamidino-2-phenylindole (DAPI) nuclear staining dye and fluorescein-rhodamine phalloidin (Molecular Probes) to enable the visualization of the nuclei and F-actin, respectively. The fluorescence images were obtained using a Cytell™ System (GE Healthcare Life Sciences, Pittsburgh, PA). The images and the thickness of the triple-layered chondrocyte sheets were analyzed on ImageJ software.
Proteomic analysis
To observe the adhesion proteins involved in the attachment of the chondrocyte sheets onto the cancellous bones, the cultures of the cancellous bones with and without the chondrocyte sheets at day 7 were subjected to label-free shotgun proteomics as previously described. 15 In brief, the cells were lysed and the total proteins were then solubilized in 0.1% sodium dodecyl sulfate solution. After determining the protein concentration, the samples were digested with sequencing grade porcine trypsin (1:20 ratio) for 16 h at 37°C. The tryptic peptides were dried using a speed vacuum concentrator and resuspended in 0.1% formic acid for nano liquid chromatography tandem mass spectrometry (nanoLC-MS/MS) analysis. The samples were injected in triplicate into a Hybrid Quadrupole Q-TOF Impact II LC-MS system (Bruker Daltonics Ltd., Hamburg, Germany), which was coupled with a nanoLC system: UltiMate 3000 LC System (Thermo Fisher Scientific, Madison, WI). Raw LC-MS data and a human protein database from the Universal Protein Resource (UniProt) were analyzed by MaxQuant 1.6.5.0. Following gene ontology (GO) term in biological process primarily involved in the adhesion process, the proteins in terms of cell adhesion (GO:0007155), ECM organization (GO:0030198), cell–substrate junction assembly (GO:0007044), and cell adhesion mediated by integrin (GO:0033617) were observed. The STITCH database (version 5.0) was used to analyze the common and predicted functional interaction networks between the selected proteins in this study and the molecules related to the cartilage functions.
Time-lapse fluorescence observations
Triple-layered chondrocyte sheets were stained with NucBlue™ Live ReadyProbes Reagent (Molecular Probes, Life Technology) for 20 min according to the manufacturer's protocol. After that, the chondrocyte sheets were transferred to either a cancellous bone or a TCPS dish. Then, the chondrocyte sheet-cancellous bone tissue was subsequently subjected to 30 min of preincubation in a CO2 incubator with a small amount of the growth medium. After confirming the attachment of the chondrocyte sheets to the cancellous bone, the fresh growth medium was carefully added. Time-lapse images were obtained every 10 min at several positions for 48 h using a BioStation IM-Q microscope equipped with a 10 × magnification objective (numeric aperture 0.5) and a chamber set at 37°C with 5% CO2 (Nikon, Inc., Melville, NY). The measurement of the cell trajectory and the mean nonoverlapped velocity of the chondrocytes, defined as the mean velocity of all tracked objects in a nonoverlapped state, were analyzed using CL-Quant software version 3.30 (Nikon).
Statistical analysis
The thickness of the triple-layered chondrocyte sheets and the mean velocity of the chondrocytes were expressed as the mean ± standard deviation. Differences between the cells' velocities at the central and peripheral areas were analyzed using an unpaired t-test, whereas those at different time points were analyzed using one-way analysis of variance. Significant differences were determined by Fisher's Least Significant Difference post hoc test using SPSS software ver. 17.0. Differences were significant when the p-value was <0.05.
Results
Properties of the triple-layered human chondrocyte sheets
Using cell sheet technology, human osteoarthritic chondrocytes could be constructed as triple-layered chondrocyte sheets. After the detachment process, the chondrocyte sheets were simply transferred to a PVDF supported membrane (Fig. 1A–C). The average thickness of the triple-layered chondrocyte sheets was 119.22 ± 10.53 μm. After culturing for 7 days, the expression of COL1A1 (dedifferentiated marker) and COL2A1 (chondrogenic marker) in the triple-layered chondrocyte sheets was observed. The expression of COL2A1 was found in the entire chondrocyte sheets, while the expression of COL1A1 was partially detected (Fig. 1E, H). The sectioned chondrocyte sheets were also incubated with DAPI nuclear staining dye and fluorescein-rhodamine phalloidin to enable the visualization of the nuclei and F-actin, respectively. The nuclei of the chondrocytes displayed round shape in the entire triple-layered cell sheets. The staining of F-actin was partially observed at the cell membrane of some chondrocytes in the triple-layered chondrocyte sheets, but it was strongly visualized at both the basal and upper sides of the triple-layered chondrocyte sheets (Fig. 1D, G, J, M).
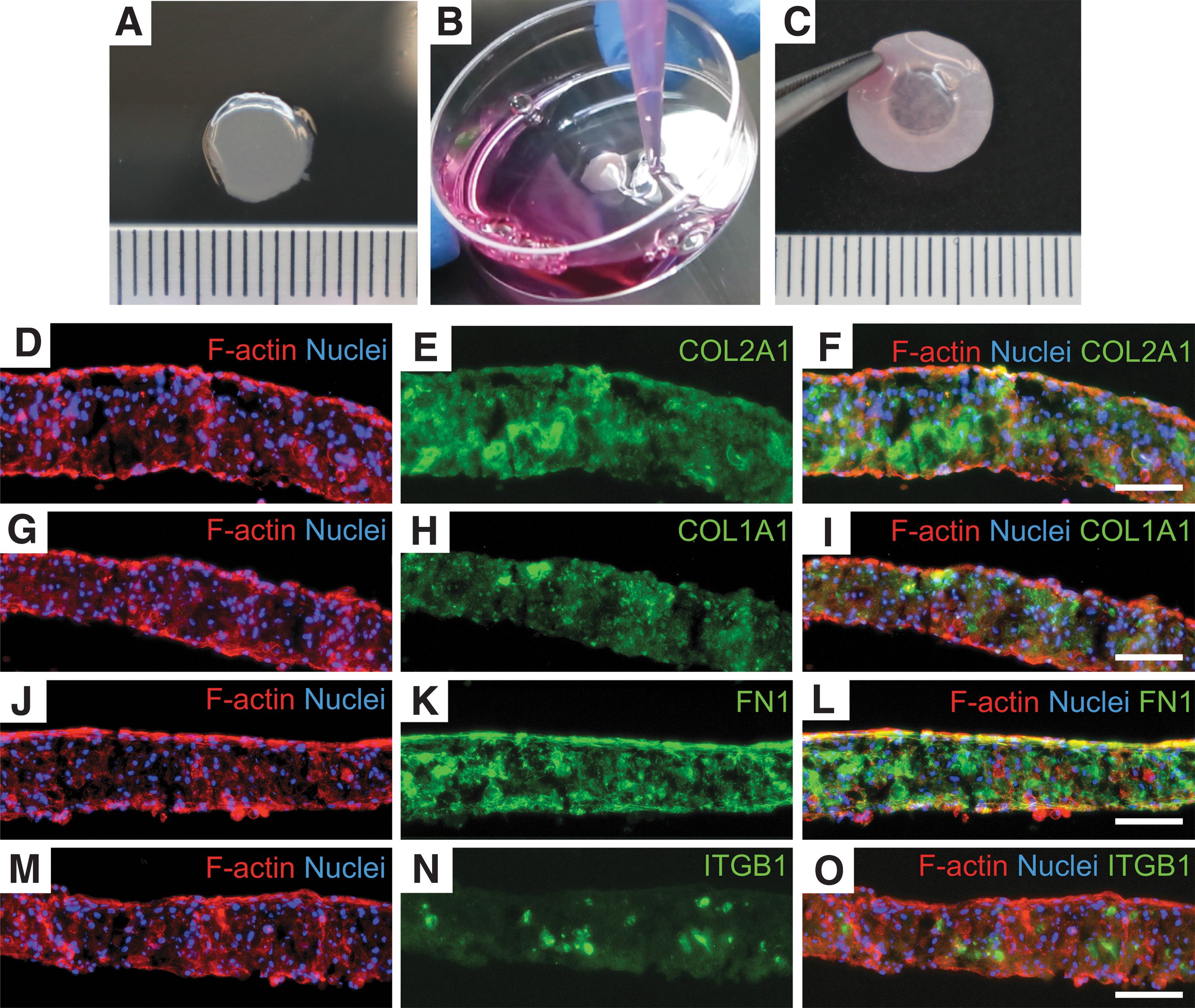
Images and immunofluorescence analysis of the human chondrocyte sheet. The triple-layered human chondrocyte sheet
To observe the potential attachment of the triple-layered chondrocyte sheets on other surfaces, the expression of FN1 was chosen because FN1 adhered to the basal side of the cell sheet after the cell detachment from the temperature-responsive surface. Hence, it functioned as a glue to attach the cell sheet to other surfaces. 16 As shown in Figure 1K, the expression of FN1 was detected in the whole chondrocyte sheet, especially at both the basal and apical sides of the cell sheets. In this study, the expression of ITGB1 was also investigated because it was considered to be the most common transmembrane receptor for COL2A1. 17 In the triple-layered chondrocyte sheets, however, the expression of ITGB1 was rarely detected similar to that of ITGB1 in the native osteoarthritic articular cartilage (Fig. 1N and Supplementary Fig. S1).
Characteristics of freeze-dried human cancellous bone
The surface topography and element composition of freeze-dried human cancellous bone characterized by the scanning electron microscope (SEM) and EDS analyses are shown in Figure 2 and Table 1. The cancellous bone showed a number of pores and interconnected porous structures (Fig. 2A, B). By randomly selecting the area in the SEM images of the cancellous bone, the EDS spectrum exhibited the peaks of calcium (Ca) and phosphorous (P) with a Ca/P ratio of 2.47 ± 0.17 by weight and a Ca/P atomic ratio of 1.91 ± 0.13, which were related to hydroxyapatite in natural spongy bone.
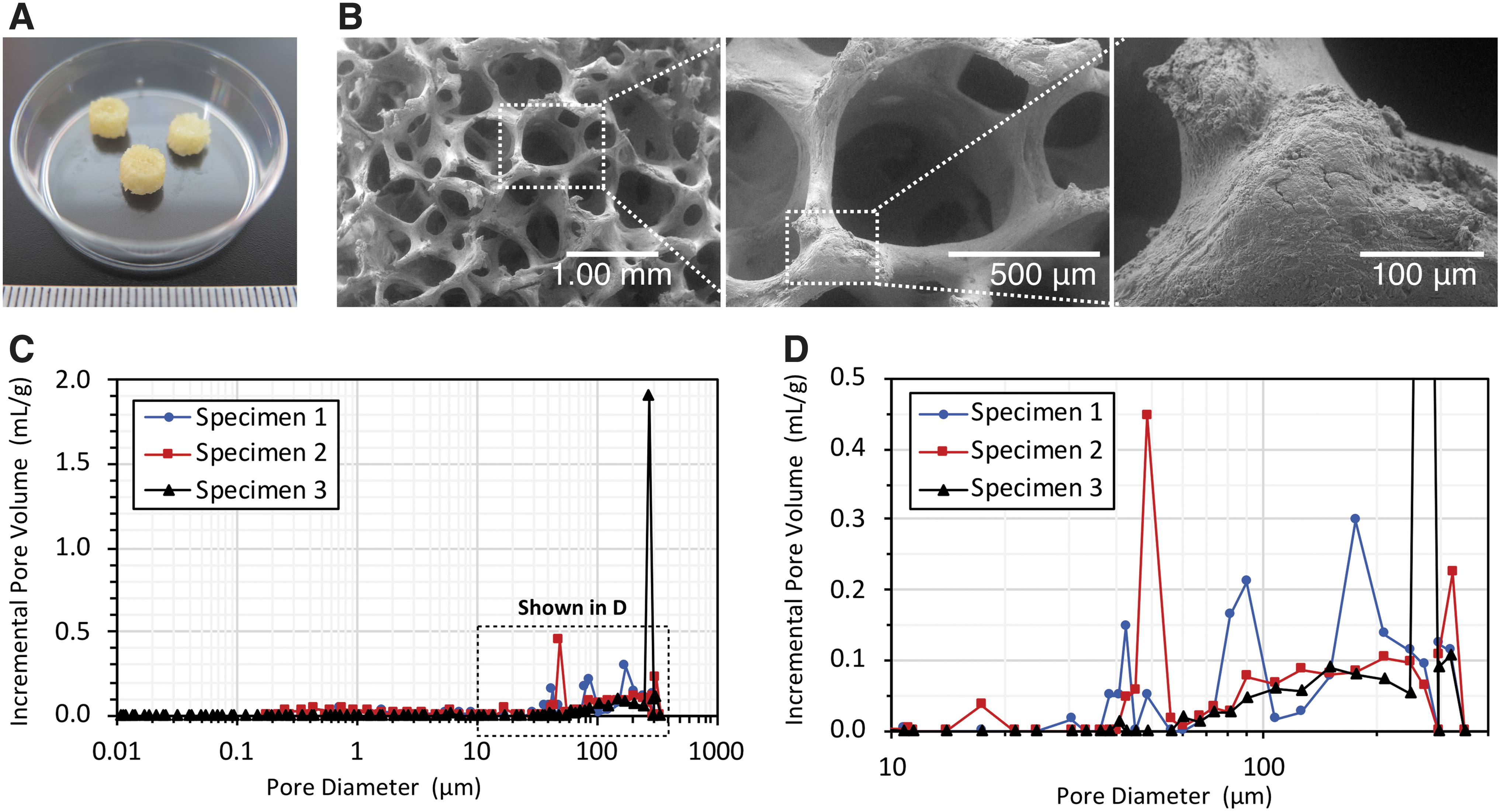
Characteristics of freeze-dried human cancellous bone. The cancellous bones were in a cylindrical shape (5 mm in diameter; 3 mm in height)
Elements of Human Cancellous Bone and Ratio of Calcium (Ca) and Phosphorous (P) by Energy Dispersive X-ray Spectrometer Analysis
The pore size distribution and porosity of the cancellous bone analyzed by MIP revealed that the cancellous bone comprised both micro (<20 μm) and macro (>50 μm) pores. As shown in Figure 2C and D, the three samples of cancellous bones designated as Specimens 1, 2, and 3 showed the pore size diameter in a range of 0.01–350 μm. The median pore diameter (volume) of the specimens was 180.60, 54.24, and 279.10 μm. The cancellous bones had a total pore area of 0.004, 0.013, and 0.003 m2/g. The open porosity values were 13.75, 24.07, and 22.84% (Table 2), respectively.
Pore Structure Characteristics of Human Cancellous Bone by Mercury Intrusion Porosimetry Measurement
Attachment of the triple-layered chondrocyte sheet on cancellous bone
The surface topography of the chondrocyte sheets after being attached to the cancellous bone was examined at day 7 of the culture using macroscopic evaluation and SEM analysis. As shown in Figure 3, the chondrocyte sheets had successfully adhered to the cancellous bone without cell sheet floating (Fig. 3A, B). The incubation time required for the strong attachment of the chondrocyte sheets on the cancellous bones was suggested to be 2 h. After 7 days of the culture, the chondrocyte sheets covered all the pores at the top area of the cancellous bone with the smooth surface topography (Fig. 3E, F). Although some chondrocytes distributed into the pores, most of the chondrocytes remained on the top layer of the cancellous bone (Supplementary Fig. S2). Compared with the top side of the chondrocyte sheet attached on the cancellous bone, the basal side of the chondrocyte sheet showed a compacted cell arrangement with a rough surface (Fig. 3C, D). These results revealed that the manipulation of the chondrocyte sheets to the osteochondral defects using cancellous bone was feasible because the cancellous bone could support the attachment of the chondrocyte sheets. Moreover, the cancellous bone could retain the shape of the chondrocyte sheets without removing the bone.

Human chondrocyte sheet-cancellous bone tissue
Adhesion protein mediated the attachment of the chondrocyte sheets on the cancellous bones
From the attachment of the chondrocyte sheets onto the cancellous bones, the proteins involved in the adhesion process of the chondrocyte sheet-cancellous bone tissues were investigated. Using the shotgun proteomic analysis, a total of 1,689 proteins were identified; 272 proteins were commonly found in both the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues, while 426 and 614 proteins were unique to the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues, respectively (Fig. 4A). With regard to GO term in cell adhesion (GO:0007155), Venn diagram shows that 10 and 19 proteins were unique to the chondrocyte sheets and the chondrocyte sheet-cancellous bone tissues, while 5 proteins were unique to the cancellous bones, respectively (Fig. 4B). Among the proteins in terms of cell adhesion in the chondrocyte sheets and the chondrocyte sheet-cancellous bone tissues, seven proteins (integrin beta-8 [ITGB8], FN1, integrin beta-like protein-1 [ITGBL1], laminin subunit alpha-2 [LAMC2], thrombospondin-1 [THBS1], nidogen-2 [NID2], tenascin [TNC]) were also identified in terms of ECM organization (GO:0030198), cell–substrate junction assembly (GO:0007044), or cell adhesion mediated by integrin (GO:0033627) (Table 3). The results also show that FN1 was identified in the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues with the term of cell–substrate junction assembly (GO:0007044).
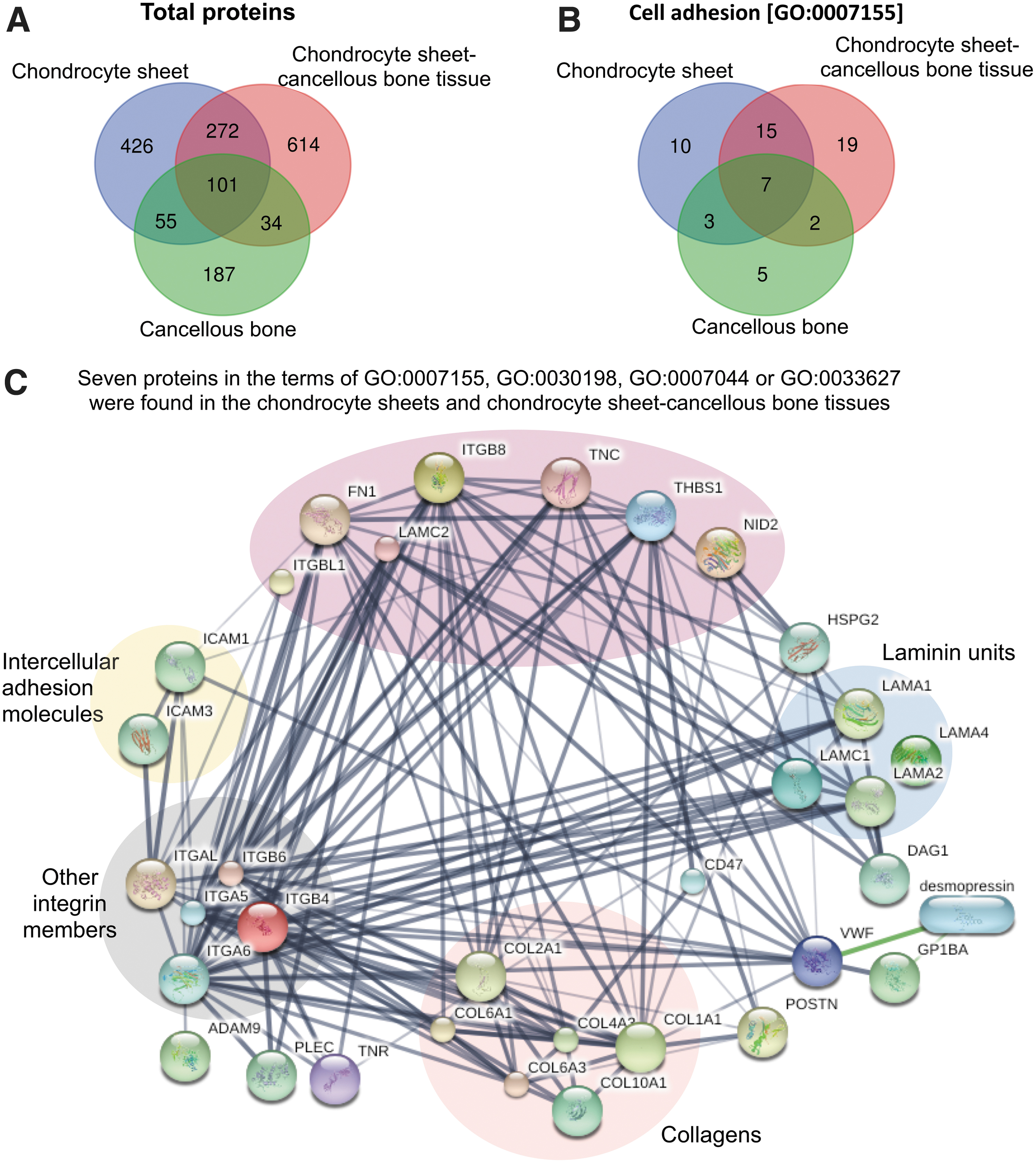
A Venn diagram showing the content overlap and differences of the total proteins
List of Proteins in Chondrocyte Sheets, Chondrocyte Sheet-Cancellous Bone Tissues, and Cancellous Bone in Terms of Cell Adhesion [GO:0007155] Also Exhibited the Biological Process in Terms of Extracellular Matrix Organization [GO:0030198], Cell–Substrate Junction Assembly [GO:0007044], and Cell Adhesion Mediated by Integrin [GO:0033627]
GO, gene ontology.
Since seven proteins showed the properties that support the attachment of chondrocyte sheets on to the cancellous bones, the possible functional interactions and associations between these proteins and other proteins available in the published article database were predicted using STITCH. As shown in Figure 4C, seven proteins were mainly shown to interact with other integrin members, collagens, and laminin units. Three proteins (FN1, TNC, and THBS1) were shown to directly interact with ITGB8, other integrin members, and COL2A1. Both FN1 and THBS1 were also shown to be connected to intercellular adhesion molecules (ICAM1 and ICAM3).
Expression of COL2A1 and FN1 in the chondrocyte sheet-cancellous bone tissues
From the interaction between the integrin members with COL2A1 and FN1, the expression of COL2A1 and FN1 in the chondrocyte sheet-cancellous bone tissues was performed by immunofluorescence staining after 7 days of the culture. As shown in Figure 5, the sectioned chondrocyte sheet-cancellous bone tissues clearly exhibited the layers of the chondrocyte sheet, adjacent zone, and cancellous bone areas. The expression of COL2A1 was detected throughout the chondrocyte sheet-cancellous bone tissues (Fig. 5B). The expression of FN1 was also observed to identify potential adhesion proteins involved in the adhesion of the chondrocyte sheets to the cancellous bone. As shown in Figure 5E, the expression of FN1 was distributed in the whole cell sheet, especially in the distinct adjacent zones between the basal side of the cell sheet and the upper side of the cancellous bone.
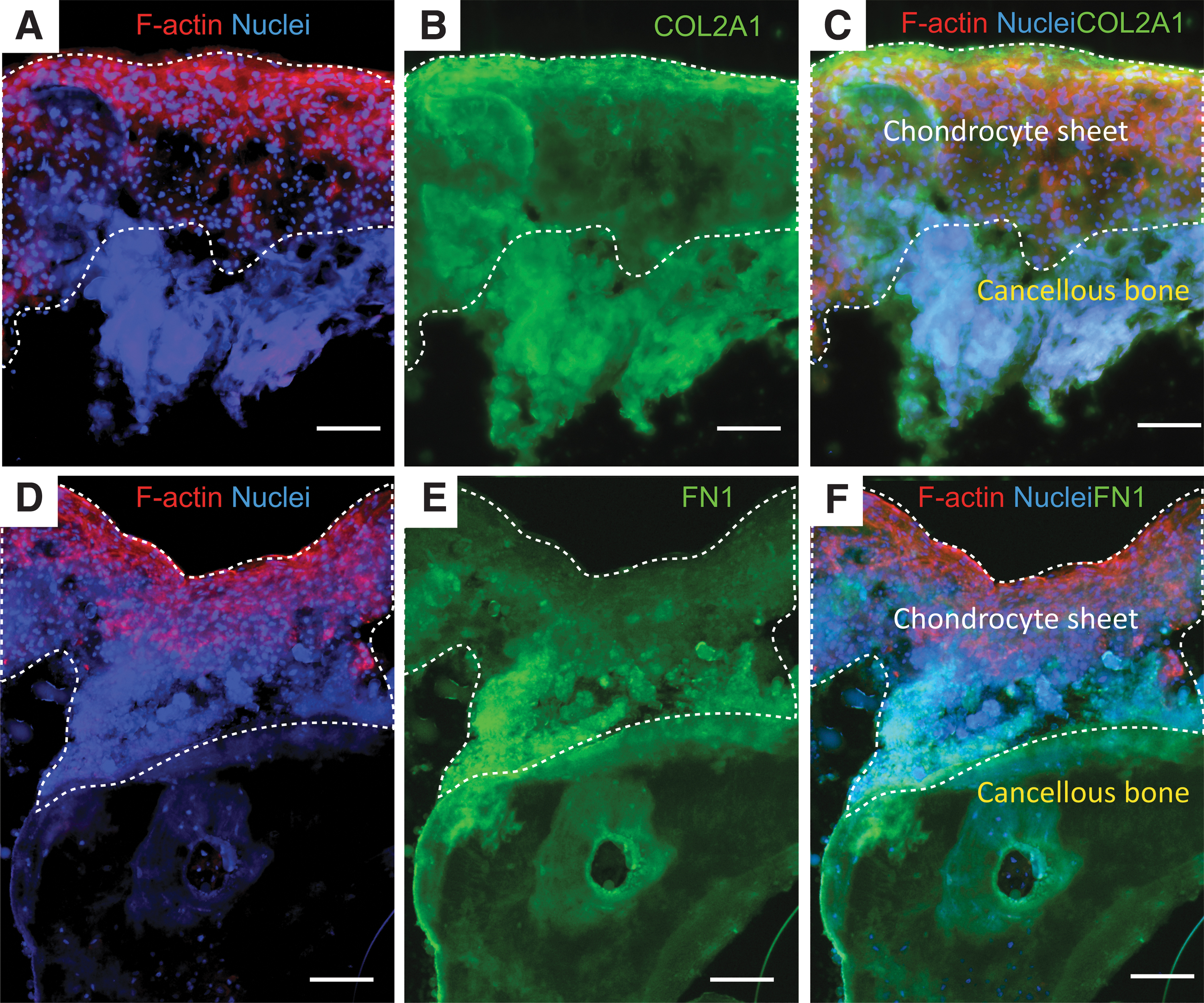
Immunofluorescence staining for COL2A1
In the chondrocyte sheet-cancellous bone tissues, the distribution of F-actin was detected throughout the layer of the chondrocyte sheets, especially in the uppermost area (Fig. 5A, D). The findings of the chondrocyte sheets adhered on the cancellous bones and the staining of F-actin with different shapes of nuclei raised the question of whether the chondrocytes were able to migrate on the chondrocyte sheet-cancellous bone tissues.
Behavior of the triple-layered chondrocyte sheets and individual chondrocyte
To elucidate the behavior of the chondrocyte sheets and the migration of individual cells on the cancellous bone, the time-lapse observation of the representative cell sheets on day 0 after placing the triple-layered chondrocyte sheets on the cancellous bone was conducted. During the 48 h of cultured chondrocyte sheets on the cancellous bones, representative videos revealed that there was no distinct migration of the whole chondrocyte sheet on the cancellous bone. In other words, the chondrocyte sheet remained stationary on the cancellous bone. However, the migration of individual chondrocyte was observed throughout almost the entire of cancellous bone both at the focal and pore areas (Fig. 6B, Supplementary Movie S1).

The behavior of the triple-layered chondrocyte sheets and individual chondrocyte on the TCPS dish and cancellous bone. Individual chondrocyte exhibited cell motility on both the TCPS dish
At the focal areas of the cancellous bone, the morphology of the chondrocytes showed a combination of round and spread cells with the round nuclei, respectively. The majority of the chondrocytes on the cancellous bone exhibited a back and forth movement by stretching along and parallel to the trabecular surface of the cancellous bone. They were connected to each other as cell clusters without cell floating in the growth medium. Moreover, the time-lapse images clearly displayed that the chondrocytes at the periphery of the cell sheet exhibited cell membrane protrusion at the focal areas of the cancellous bone (Fig. 6B, red arrows). This indicated that the chondrocytes at the focal areas of the cancellous bone did not only maintain cell–cell contact but also preserved cell–substrate adhesion. At the pore areas of the cancellous bone, most of the chondrocytes exhibited a spindle-like shape. They displayed random mobility with collective cell migration and maintained cell–cell contact. However, the chondrocytes were moving back and forth between the focal and pore areas leading to the loss of cell–cell contact in some regions of the cell sheets at the bottom of the pore zone. During the 48 h of the culture, the cell trajectory was not detected in the cancellous bone without the chondrocyte sheets, which indicated that any cell migration came from the chondrocytes (Fig. 6C).
Unlike the chondrocyte sheets on the cancellous bone, the majority of the chondrocytes at the center of the cell sheet on the TCPS possessed a spherical shape and moved randomly in all directions. Most of the chondrocytes on the top layer of the cell sheets did not grow on top of each other, as shown by the contact inhibition within 48 h (Fig. 6A, Supplementary Movie S2). At the periphery of the chondrocyte sheet, the cells displayed a directional migration toward the free areas of the polystyrene surface with the protrusion of the cell membrane (Fig. 6A, red arrows).
To observe the migratory direction of the chondrocytes on the different surface topographies, the trajectories in X and Y directions of the chondrocytes adhered on the cancellous bone and TCPS were compared. As shown in Figure 6D and E, the trajectories of the chondrocytes adhered on the cancellous bone (Fig. 6E) were similar to those of the chondrocytes adhered on the TCPS (Fig. 6D). The chondrocytes at the central area moved in all directions, whereas the chondrocytes at the peripheral area moved toward the free areas. However, the migratory distance of the chondrocytes on the cancellous bone and TCPS was slightly different. At the central area, further movement of the chondrocytes was found on the cancellous bone. The range of cell migration distance in X direction on the cancellous bone was from −46 μm to 46 μm and the range of distance in Y direction was from −63 μm to 21 μm, whereas the range of cell migration distance in X direction on TCPS was from −16 μm to 16 μm and the range of distance in Y direction was from −27 μm to 9 μm. At the peripheral area, however, the migratory distance of the chondrocytes adhered on the cancellous bone was less than those of the chondrocytes adhered on the TCPS (Cancellous bone: X direction; −13 μm to 39 μm and Y direction; −32 μm to 32 μm vs. TCPS: X direction; 0 μm to 148 μm and Y direction; −82 μm to 82 μm).
Since the chondrocytes were slightly motile with different patterns of the cell trajectory between the central and peripheral areas on both the cancellous bone and TCPS, we then observed the velocity of the chondrocyte migration, which could have been affected by the roughness of the cell substrate. During the 48 h, although the chondrocytes were adhered on the different surface roughness, there were no significant differences between the average cell migration velocity of the chondrocytes on the rough cancellous bone and on the smooth TCPS (velocity <0.30 μm/min) (Fig. 6F, G). However, the migration velocity of the chondrocytes on the rough cancellous bone and the smooth TCPS was reduced at the central area compared to those at the peripheral area. Interestingly, the chondrocyte migration velocity was significantly reduced on the TCPS at the central area compared to the peripheral area at 12 h (0.07 ± 0.01 μm/min vs. 0.13 ± 0.02 μm/min; p < 0.05), 24 h (0.07 ± 0.02 μm/min vs. 0.12 ± 0.02 μm/min; p < 0.05), 36 h (0.05 ± 0.01 μm/min vs. 0.11 ± 0.02 μm/min; p < 0.05), and 48 h (0.05 ± 0.02 μm/min vs. 0.11 ± 0.01 μm/min; p < 0.05) (Fig. 6F). For the migration velocity of the chondrocytes on the rough cancellous bone, there was no significant difference between the central and the peripheral areas, as well as among the time points at 12, 24, 36, and 48 h (Fig. 6G).
Discussion
This study presented new cartilage bone tissue constructed by the attachment of human chondrocyte sheets on freeze-dried human cancellous bone to treat osteochondral defects. We demonstrated that the biological properties of the human chondrocyte sheets and the physical characteristics of the human cancellous bone mainly supported the attachment and growth of the individual cells in the layered chondrocyte sheets. The findings also provided insights into understanding the dynamic communication of the F-actin cytoskeleton and adhesion proteins in chondrocyte sheet-cancellous bone tissue.
Based on cell sheet technology, human chondrocyte sheets harvested without enzymes mainly preserved cell–cell connection and ECM, which played a key role in the adhesiveness of each layered cell sheet and the attachment to other surfaces. 18 However, without the use of any supporting materials after the cell detachment process, the manipulation of human chondrocyte sheets onto the new surface resulted in the tearing and the shrinkage of the cell sheet. 10
In this study, we showed that the harvested chondrocyte sheets adhered well on the cancellous bone as a supporting material facilitating the maintenance of chondrocyte sheet architecture during an in vitro culture, and it could potentially be used in the tissue manipulation during an in vivo study. From the results of the immunofluorescence staining, the chondrocyte sheet-cancellous bone tissues showed the expression of COL2A1 and FN1 similar to that of the chondrocyte sheets only (Figs. 1 and 5). This indicated that after adhering the chondrocyte sheets onto the cancellous bones, the expression level of cartilage-specific protein with the adhesive property was maintained in the chondrocyte sheet-cancellous bone tissues. Regarding the structure of the cell sheet, the thin and rough surface at the basal side of chondrocyte sheets, which had been attached to the TCPS (Fig. 3), suggested that the surface of the chondrocyte sheets contained adhesive properties and allowed the cell attachment to other surfaces. 16 Moreover, the identification of FN1 and ITGB8 in the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues from the proteomic result led us to propose that the predominant adhesion proteins mostly impacting the initial attachment of the chondrocyte sheets were FN1 and integrin in the layered chondrocyte sheets.
In addition to the biological properties of the chondrocyte sheets, we were concerned with the physical characteristics of the cancellous bone scaffolds that supported the attachment and growth of the layered chondrocyte sheets. The physical characteristics of cancellous bone were shown to primarily affect the cellular attachment and growth involved in the pore size and porosity. 19 In general, the scaffolds with a very small pore size and low porosity limited cell migration, as well as nutrient diffusion and waste removal, but the cell–cell contact was increased. Conversely, the scaffolds with an extensively large pore size resulted in a reduction of the cell–cell contact because the cells recognized their surfaces as a two-dimensional (2D) substrate instead of a 3D structure.19,20 Based on cartilage tissue engineering, scaffolds with pore sizes in the range of 250–350 μm supported the proliferation of the chondrocytes and their ECM production. 21 In this study, the porosity of freeze-dried human cancellous bone was ∼13–24%, and the pore sizes were in the range of 0.1–350 μm (Fig. 2, Table 2), which were slightly different ranges from other studies.22,23 The porosity and density of the cancellous bone typically displayed a wide variability from one region to another in the same long bone due to its structural heterogeneity. 24 Therefore, we suggested that the low porosity of cancellous bones might be because the cancellous bones were collected from the locations next to the cortical bone, and the sample preparation differed from other studies.
Unlike the use of a scaffold for cell seeding throughout the structure,25,26 the cancellous bone scaffold in this study acted as a supporting material to generate cell–substrate adhesion mainly on the top surface of the bone and to maintain cell–cell contact in the layered chondrocyte sheets. Using cell sheet technology, the chondrocytes remained in a compact shape as a 3D structure in the layered chondrocyte sheets. Although some chondrocytes moved into the pores of the cancellous bone, most cells inside the layered chondrocyte sheets were located in the uppermost area of the cancellous bone and exhibited the expression of COL2A1. This demonstrated that the pore size and porosity of the cancellous bones of this study did not impede the ECM production of the chondrocytes, while still providing the adherence of the chondrocytes on the cancellous bone during the 7 days of the culture. Moreover, the freeze-dried human cancellous bones provided a similar property of natural bone containing hydroxyapatite, which typically allowed the attachment of the bone cells. Therefore, the physical characteristics of human cancellous bone facilitated the further attachment of the layered chondrocyte sheets and might support the ingrowth of new tissues, which would be beneficial for tissue engineering applications.
Since the chondrocytes are naturally embedded in a dense ECM, they could not migrate to the damaged lesions and start healing the cartilage resulting in the low synthesis of a new ECM and poor cartilage repair. 27 At the osteochondral lesions, therefore, one of the crucial roles for healing damaged cartilage was the migration and proliferation potential of chondrogenic precursor cells.28,29 Generally, cell migration velocity was used as a key biophysical parameter to observe the cell ability to migrate from healthy to damaged areas. 30 It has been shown that cell migration velocity is mostly affected by the surface topography and the stimulation by growth factors, which subsequently impacted the cell fuctions.30,31 Compared with native chondrocytes, the migratory cells harvested from damaged cartilage expressed higher levels of proliferative and migrating genes, but lower level of cartilage matrix genes such as COL2A1 and aggrecan.32,33 The previous study also reported that chondrocytes attached on the fibronectin-coated porous membrane exhibited low speed movement and retained the ability to synthesize COL2A1. 34
According to the result of the migration of individual chondrocyte into the pores of the cancellous bone in this study, we observed whether the individual chondrocyte migration affected the structure of the chondrocyte sheets and the synthesis of cartilage-specific proteins in the chondrocyte sheet-cancellous bone tissue. From the results of the time-lapse observation, it clearly revealed the stability of the layered chondrocyte sheets on the cancellous bone and the individual chondrocyte migration during 48 h (Fig. 6). The migration velocity of the individual chondrocyte adhered on the cancellous bone and the TCPS was low; nearly the same as that of the chondrocytes on the tissue culture plate in the previous study (∼0.15 μm/min). 29 Moreover, the chondrocyte sheets maintained their smooth surface structure without the loss of their chondrogenic phenotype, as shown by the SEM result and the immunofluorescence staining of COL2A1, respectively. These indicated that the movement of individual chondrocyte did not affect the structure of the chondrocyte sheets and the synthesis of COL2A1 in the chondrocyte sheet-cancellous bone tissues. Moreover, the cancellous bones, where the surface was not continuous due to the nature of the spongy bone, not only served as appropriate supporting material but also provided a suitable microenvironment for cell growth.
In the migrating cells, the morphological behavior is mainly driven by a dynamic F-actin cytoskeleton, which is coupled to the ECM by means of a focal adhesion (FA) complex.35,36 The FA complex acts as the contact sites of the cells to the ECM on the substrates through the integrin transmembrane proteins and, subsequently, processes cellular events, such as cell survival, differentiation, proliferation, and division.36,37 In this study, the staining of the F-actin cytoskeleton was found at the periphery of the layered chondrocyte sheet and the uppermost layer of the chondrocyte sheet-cancellous bone tissues. This suggested that these areas were mostly prevailed by migrating cells. The migration of cells on the cancellous bone may occur through the dynamic communication of the cell cytoskeleton and some adhesion proteins.
From the shotgun proteomic results, FN1 was detected in both the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues, while ITGB8 was identified in these tissues instead of ITGB1, which had been mostly reported to bind collagens and fibronectin in chondrocytes. 38 Unlike other reports, the previous study by a microarray analysis showed that the dominant beta integrin subunits in human articular chondrocytes were ITGB1, ITGB5, ITGB7, and ITGB8. 39 Moreover, one of the integrin subunits consistently detected in the chondrogenesis model was ITGB8, 40 which was also found to interact with fibronectin and vitronectin. 41 In this study, the STITCH database showed the interaction of ITGB8 with FN1 and the collagens in the chondrocyte sheets and chondrocyte sheet-cancellous bone tissues. The tissues also exhibited cell attachment, cell migration, and the staining of the F-actin cytoskeleton (Fig. 4). These findings indicated a possibility that the mechanism was involved in the chondrocyte sheet attachment to the cancellous bone and initiated individual cell migration that occurred through the dynamic communication of the F-actin cytoskeleton, FN1, and ITGB8 (Fig. 7). In the chondrocyte sheet-cancellous bone tissues, the contact sites of the chondrocyte sheets could be connected to either FN1 or collagens and adhered on the cancellous bones through the ITGB8 subunit. After the cell attachment process, the dynamic rearrangement of F-actin in migratory chondrocytes might sense the communication of adhesion molecules at the cell membrane and, subsequently, lead to the synthesis of the new ECM in the chondrocyte sheet-cancellous bone tissues.
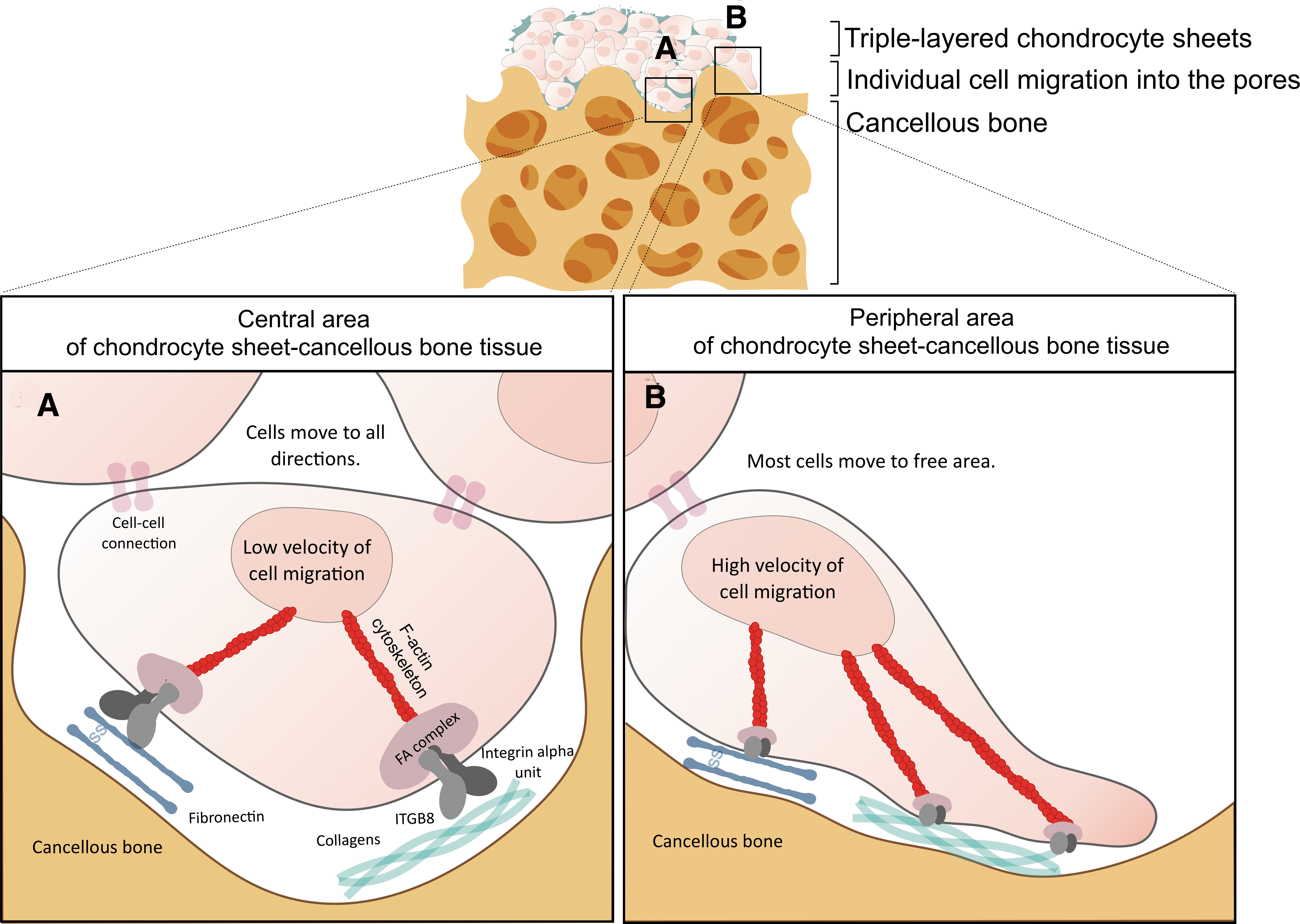
A schematic illustration of the chondrocyte sheet-cancellous bone tissue showing the behavior of the chondrocytes at the central
In osteochondral tissue engineering, the contrasting needs of the cartilage and bone tissues, as well as the integration between the two layers, should be carefully designed due to their structural and functional complexity. 42 A variety of studies reported the methods to construct osteochondral-like tissues, which provided an appropriate environment for cartilage ECM production at the top layer and supported the expansion of stem cells at the bottom layer.26,43 However, the interface between the two layers required additional steps to produce well integrated bone-cartilage interfaces such as using an iterative layering freeze-drying process, 44 suturing the tissue construct together, 45 and using gelatin or fibrin glue to stick all the layers of tissue constructs.26,45 In this study, we demonstrated an effective method to construct osteochondral-like tissues using the triple-layered chondrocyte sheets and cancellous bones without those additional steps, which would be helpful in reducing the cost and time during the tissue preparation in a preclinical study. Compared to native articular cartilage, however, the cultures of chondrocyte sheet-cancellous bone tissues in a growth medium for 7 days showed the densely packed of chondrocytes with a small portion of the ECM. Thus, further studies are needed to determine the ECM production at longer time points with additional growth factors and to assess their molecular mechanisms for making a precise structure of the complex tissue with the proper functions. Currently, the thickness of cartilage-like tissue constructed by cell sheet technology was less than 4 mm, which did not meet the mechanical integrity requirement as a native cartilage in the range of tissue thickness at 2–4 mm. 46 A thicker chondrocyte sheet, which could support the mechanical properties of the tissue-engineered constructs, would be one of the greatest challenges. Furthermore, the integration of the chondrocyte sheet-cancellous bone tissues to the osteochondral defects without tissue delamination needs further investigation to ensure the effectiveness of the osteochondral-like tissues in a preclinical study.
Conclusions
Human chondrocyte sheet-cancellous bone tissue was successfully constructed by the attachment of triple-layered chondrocyte sheets on freeze-dried cancellous bones. We emphasized that adhesion proteins, including FN1 and ITGB8 in the chondrocyte sheets, play an important role in the attachment process during the 7 days of the culture in a growth medium. Using cancellous bones as a supporting material, the structure of the triple-layered chondrocyte sheets could be retained, which further supported the cell sheet attachment, individual cell migration, and their ECM production. Moreover, a better understanding of cell behavior, which involved the communication of the F-actin cytoskeleton and adhesion molecules, may pave the way for strategies to improve the construction of osteochondral-like tissues and to predict the repair mechanism of osteochondral lesions using chondrocytes combined with scaffolds in translational research.
Footnotes
Acknowledgment
The authors acknowledge King Mongkut's University of Technology Thonburi for funding Postdoctoral Fellowship to S.W.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by the Health Systems Research Institute (HSRI), Ministry of Public Health, Thailand.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.