
Abstract
Engineered neural stem cells (NSCs) have recently emerged as a promising therapy. Acting as a tumor-homing drug-delivery system, NSCs migrate through brain tissue to seek out primary and invasive tumor foci. NSCs can deliver therapeutic agents, such as TNFα-related apoptosis-inducing ligand, directly to the tumor and suppress glioblastoma (GBM) in murine models. While the mainstays for evaluating NSC migration and efficacy have been two-dimensional chemotaxis assays and mouse models, these low-throughput and small-scale systems limit our ability to implant and track these cells for human translation. To circumvent these challenges, we developed a three-dimensional culture system using a matrix of poly-
Impact statement
Tumor-homing and tumor-killing-engineered neural stem cell (NSC) therapies have shown immense promise in both preclinical and clinical trials. However, as cell therapies continue to evolve, cost-effective and high-throughput screening assays are needed to assess the proliferation, migration, and efficacy of these cells. In this study, we developed a bioinspired brain matrix for the evaluation of engineered NSCs. Importantly, this matrix is easy to fabricate, scalable, and allows for sterile real-time, noninvasive imaging using our custom bioreactor. We then utilized the bioinspired brain matrix system to answer key questions around the tumor-homing migration and efficacy of engineered NSC therapies that are challenging to address with traditional models.
Introduction
Primary malignant brain tumors are responsible for the most years of life lost of any cancer. 1 Of these, glioblastoma (GBM) is the most common and aggressive. Surgery, followed by chemoradiotherapy remains the standard of care, but unfortunately results in a median survival of under 2 years and a 5-year survival rate of <5%. 2 A major reason GBM remains difficult to treat is the highly invasive nature of the disease, where migratory tumor cells invade healthy brain tissue, making recurrence and associated patient mortality nearly inevitable.3–5 Additionally, there is a lack of noninvasive methods of local delivery. Systemic delivery, on the other hand, is hindered by both the blood/brain barrier and vascular dysfunction in the tumor, which is exacerbated by the fact that many current drugs have short half-lives and therefore do not accumulate to therapeutic concentrations in the brain.3,6,7 Moreover, brain metastases outnumber primary brain tumors significantly in frequency and result in an even lower median survival: <1 year. 5 New treatments are desperately needed to improve outcomes in all types of brain cancers.
In efforts to develop therapies that are more effective against brain cancer, neural stem cells (NSCs) have been increasingly explored as drug delivery vehicles. Their inherent tropism to various cancer types, especially cancers of the brain, makes NSCs attractive carriers for tumoricidal agents, to which they confer increased persistence along with active targeting.7–10 Based on the strength of promising preclinical data in mice, NSC therapy recently entered human clinical trials. Despite the promising advancements, numerous questions around NSC therapy remain due to gaps in relevant models that bridge the size and scale of NSC brain therapy from murine models to human patients.
Tumor-selective homing is one of the most critical and unique aspects of NSC therapy, yet little is known about tumor homing at distances unachievable in size-constrained mouse models. Studies using large animal models are costly to perform in vivo and cannot be recapitulated at scale in commonly used, less-expensive rodent cancer models. As such, in vitro cell migration-based assays remain the mainstay of NSC homing. These include: (1) measurement of cell movement across a scratch or gap, (2) detection of cells having moved through a membrane from one chamber to another, or (3) measurement of cell movement in microfluidic devices capable of three-dimensional (3D) culture.11–14 Each strategy is limited in terms of the number of cells that can be investigated, and these techniques do not recapitulate migration at the longer distances necessary in a human brain. 15
Moreover, two-dimensional (2D) culture especially fails to incorporate the physical and chemical cues that regulate NSC migration in vivo and restricts cell–cell interactions.
13
In fact, cells grown in 2D cultures differ significantly in phenotype and genotype from cells grown in vivo or in 3D bioinspired hydrogels.
16
To overcome these limitations we developed a custom, scalable 3D hydrogel system referred to as bioinspired brain matrix (BBM). The model is composed of an agarose base and has poly-
Materials and Methods
Cell lines
U87 glioma cells were purchased from the American Type Culture Collection and cultured according to manufacturer's specifications. Human fibroblasts (NHF1) were provided by W. Kauffman (University of North Carolina School of Medicine). 231-BR cells were provided by Carey Anders (University of North Carolina Lineberger Cancer Center). C17 mouse NSCs were used in all NSC experiments and were acquired from Sigma-Aldrich. Each cell line was cultured at 37°C, 5% CO2 in Dulbecco's modified Eagle's medium (DMEM, 10% fetal bovine serum [FBS], 1% penicillin–streptomycin; Gibco). Cells were engineered to express green fluorescent protein (GFP), mCherry (mC) fluorescent protein, and/or TNFα-related apoptosis-inducing ligand (TRAIL) through a 24-h transduction with lentivirus (LV) in the presence of polybrene (8 μg/mL; Sigma). All LV constructs were packaged as LV vectors in 293T cells using a helper virus-free packaging system as described previously.18,19
BBM fabrication
The BBMs were fabricated by mixing 3D cell culture medium (DMEM [1% antibiotic–antimycotic, 1% glutamine, 2% FBS, no phenol red]; Gibco) with a 1.2% w/v agarose (Gene Mate LE Agarose; BioExpress) dissolved in phosphate-buffered saline (PBS) at a 1:1 ratio. This solution was poured into a plastic mold with 2.2 mg/mL meltblown PLA 6100 scaffold microfibers, which were obtained from the Non-Wovens Institute at North Carolina State University. Before being added to the mold, the PLA fibers were sterilized using ultraviolet (UV) light and hydrated in the 3D medium overnight. Next, the fibers were manipulated using forceps to distribute the fibers homogenously throughout the BBM solution as it cooled. The BBMs were incubated at room temperature to allow gelation before transferring from the mold to the appropriate cell culture incubator. BBMs were kept hydrated using 3D medium and incubated at 37°C/5% CO2. Medium was changed every 3–4 days.
BBM cell seeding
To inoculate the BBM with NSCs and tumor cells, the cells were suspended at the desired concentration in DMEM. To allow for injection without backflow, the pipette was inserted 10 mm into the BBM, then withdrawn ∼5 mm to create a cavity in which to inject the cells. Cells were injected at a concentration of 5.0 × 104 cells in 3 mL for the mini-BBMs or 3.0 × 107 in 100 μL for the life-size BBMs. The seeded BBM was incubated at room temperature for 30 min to allow the cells to adhere before being immersed in 3D media and incubated at 37°C, 5% CO2.
Scanning electron microscopy
BBM samples were prepared, seeded, and cultured as described above. Fourteen days postseeding, BBMs were fixed using 10% formalin then transferred to 1 × PBS. Using an 8 mm biopsy punch and scalpel, samples were trimmed to the region surrounding the injection site, where cells would be most likely to persist. The samples were then dehydrated sequentially in 50%, 75%, 90%, and 100% ethanol. Next, samples were dried using a critical point dryer (Tousimis Autosamdi-931), mounted on aluminum stubs, and sputter coated with 8 nm of gold–palladium (Cressington Sputter Coater 108auto). The samples were imaged on a FEI Helios 600 Nanolab Dual Beam System microscope using a 2 kV accelerating voltage.
BBM serial imaging
The BBMs were imaged on an in vivo imaging system (IVIS Kinetic; Caliper LifeSciences) for fluorescence. Media were aspirated from the samples before imaging. Cells expressing GFP were probed for using an excitation and emission wavelength spectrum of 465–500 nm, and the mC-expressing cells were observed using a spectrum of 570–620 nm. An exposure time of 5 s was used for all imaging sessions. Tumor signal was quantified using Living Image software. Following imaging, the BBMs were resuspended in 3D media and returned to the incubator.
Evaluating BBM optical transparency
Mini-BBMs were fabricated as previously described and kept in a six-well plate with 3D media. Absorbance was read at 426 nm using a Molecular Devices SpectraMax M2 plate reader. Percent transmittance was calculated using the following formula,
Statistics
Data were analyzed by the Student's t-test, one-way analysis of variance/Bonferroni, or log-rank test when appropriate. Data were expressed as mean ± standard deviation. Significance between groups was denoted by *p < 0.05; **p < 0.01; ***p < 0.001. Replicate number is defined by n in figures.
Results
BBM fabrication and bioreactor design
To develop cost-effective models that will allow investigation of NSC therapy on a larger scale, two versions of the 3D hydrogel model were produced. The first was a life-sized BBM that measured ∼1.5 × 103 cm3 in volume, and the second was a mini-BBM that was ∼7.5 cm3, which is comparable to small rodent in vivo models. Figure 1A depicts schematically the development and use of BBMs to study cell growth and migration. Briefly, PLA was sterilized through UV light for 30 min in a tissue culture hood and incubated overnight in 3D cell culture medium (DMEM, 1% FBS, 1% antibiotic–antimycotic). Next, 0.6% agarose (w/v) was dissolved in 1:1 PBS:3D cell culture medium and poured over PLA in the plastic mold. Gelation occurred as the agarose cooled at room temperature. Importantly, the resulting BBM was optically transparent to enable noninvasive tracking of cell populations within the matrix.

BBM and bioreactor design.
The mini-BBM (Fig. 1B) could be cultured in 6-well polystyrene plates (Fig. 1C). However, it was necessary to design a bioreactor in which to culture the human-size BBM (Fig. 1D). To this end, a 6-L round polycarbonate container was used as a central housing. An acrylic lid was laser cut to the container's dimensions. The lid was rationally designed with filter inserts to allow for proper gas exchange while remaining sterile. The bioreactor was designed to be compatible with serial noninvasive optical imaging, thus the walls and lid comprised material that was optically transparent, whereas the base and fittings were designed with black material to minimize the reflection of light during imaging. A custom impeller was 3D printed to circulate nutrients throughout the bioreactor without creating a circular flow pattern that could shear parts of the BBM contained within, a common issue with standard stir bar-based bioreactors. 20
Cell–BBM interactions
To next investigate the interaction between the PLA fibers and agarose, as well as cell distribution and attachment to the matrix, we utilized scanning electron microscopy (SEM). The SEM images showed that while the PLA fibers were primarily dispersed evenly throughout the matrix, more condensed groups of fibers were present in certain locations (Fig. 2A, B). The average PLA fiber diameter was determined to be 15.83 ± 1.99 mm through ImageJ analysis (n = 10) (Fig. 2C). With increasing magnification of the BBM agar component, the continuous morphology of the material was interspersed with cracks, which were likely an artifact of the critical point drying procedure (Fig. 2D). Following injection of tumor cells, cell clustering and attachment to the agar matrix was observed, directly adjacent to the PLA fibers (Fig. 2E, F).

BBM characterization. Interaction between PLA and agar is visualized in
Implanting GBM tumors and tracking growth within BBMs
Initial experiments sought to characterize growth rates of two different types of brain cancer in both the small and large BBM systems. The tumor cell lines, U87 and MDA-MB-231-BR (231-BR), were selected due to their sensitivity to TRAIL.21,22 To first investigate GBM, the human glioma line U87 was engineered to express the optical reporter mC, and tumor cells were injected 2 mm deep into mini-BBMs at the same cell density used in traditional animal models: 5 × 105 cells in 3 μL (Fig. 3A).3,23 Thirty minutes after implantation, BBMs were immersed in 3D cell culture medium and moved to an incubator for long-term culture. A similar method was used in the life-size BBM model, where 3.0 × 107 mC cells were implanted ∼40 mm deep into the hydrogel, which was then transferred to the bioreactor (Fig. 3B). Tumor volume changes were then serially tracked using IVIS imaging. We found that 1 week after implantation, U87 tumor volumes increased 48-fold in the life-size BBM and greater than 52-fold in the mini-BBM (Fig. 3C). These growth rates are comparable to experimentally determine in vivo growth rates when these tumors are established in nude mice. 19 Thus, remaining experiments were all conducted using the same tumor cell density as described here.
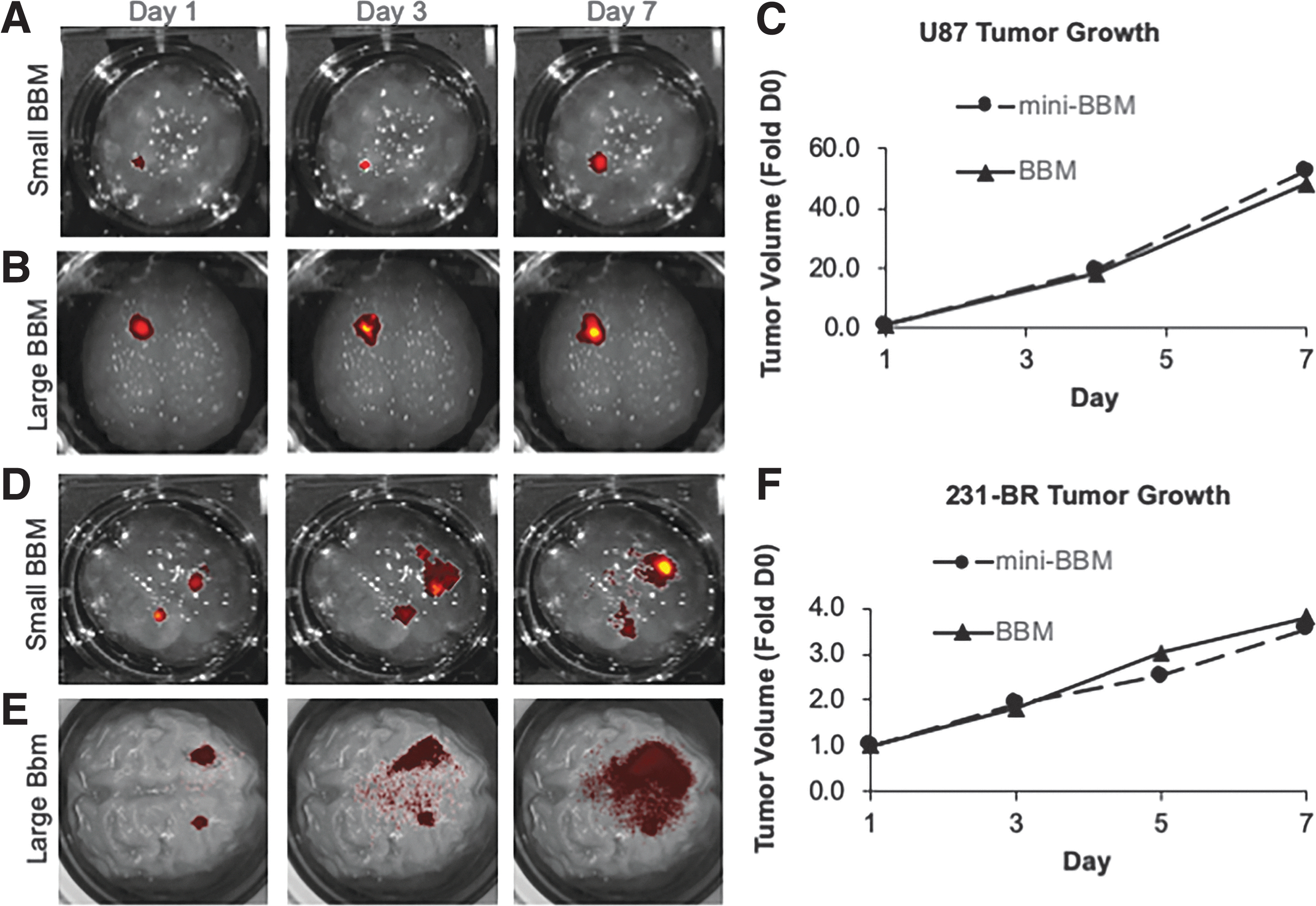
Brain tumor growth in BBMs. Representative images of GBM fluorescence over 1 week of
The testing was then expanded to explore metastatic brain cancer, specifically breast-to-brain metastases. Human 231-BR cells, a brain-seeking variant of the common metastatic breast cancer line MDA-MB-231, was tested because it preferentially metastasizes to brain tissue. 24 In the clinic, brain metastases often present as a multifocal disease. To model this growth pattern, multiple 231-BR tumor foci were established through implants of 2.5 and 5.0 × 105 cells contralateral to each other in both mini- and life-size BBMs (Fig. 3D, E). One week following implantation, serial kinetic imaging showed that tumor volume increased approximately fourfold in both the mini- and life-size BBMs after 1 week (Fig. 3F), which also closely mimics the growth pattern that has previously been reported in nude mice. 25 Altogether, these data provide evidence that tumor growth in BBMs closely mimic in vivo tumor growth kinetics, an important factor when designing redosing studies and tracking tumor invasion.
NSC homing to established tumor in BBM
The ability of NSCs to migrate through the brain and actively seek out solid and invasive tumor foci is one of the most beneficial aspects of NSC-based therapy for brain cancer. The tumor-homing capacity of NSCs has been extensively studied in mouse models of brain cancer, yet little is known about NSC migration distance capabilities in a human patient. The impact of NSC distance from the tumor and the resulting efficacy is unknown beyond the scale of a mouse. To assess NSC migration in the BBMs, mini-BBMs were first implanted with U87-mC to establish cancer foci (referred to as “tumor”) or with unlabeled fibroblasts to model nontumor foci (referred to as “nontumor”). Three days posttumor implantation, NSCs engineered through transduction with LVs to express GFP (NSC-GFP) were implanted ∼2 mm lateral to the tumor or fibroblast focus (Fig. 4A). On days 1 and 7 postimplantation, a mini-BBM was fixed, and biopsies containing both cell types were collected and sectioned into ∼1 mm slices using a rat brain slicer matrix. Fluorescent micrographs of the slices were captured using a stereomicroscope and fluorescent signal was quantified in ImageJ. Analysis showed that NSCs migrated more than 100-fold further in the presence of tumor-containing BBM compared with nontumor-containing BBM, traversing 1.3 ± 0.05 mm of the 2 mm gap between the sites of NSC and GBM implant over the 7-day period (Fig. 4B). Quantification of GFP signal in different regions near the injection site or GBM focus further confirmed this migration, as analysis of the fluorescent intensity showed 78% ± 2.0% of NSC's migrating region of the BBM when implanted near glioma cells for 1 week. When cocultured with fibroblasts instead, NSCs exhibited no directional migration, with only 25% ± 0.9% of GFP signal found in the migrating region in the nontumor BBM ImageJ (Fig. 4B).
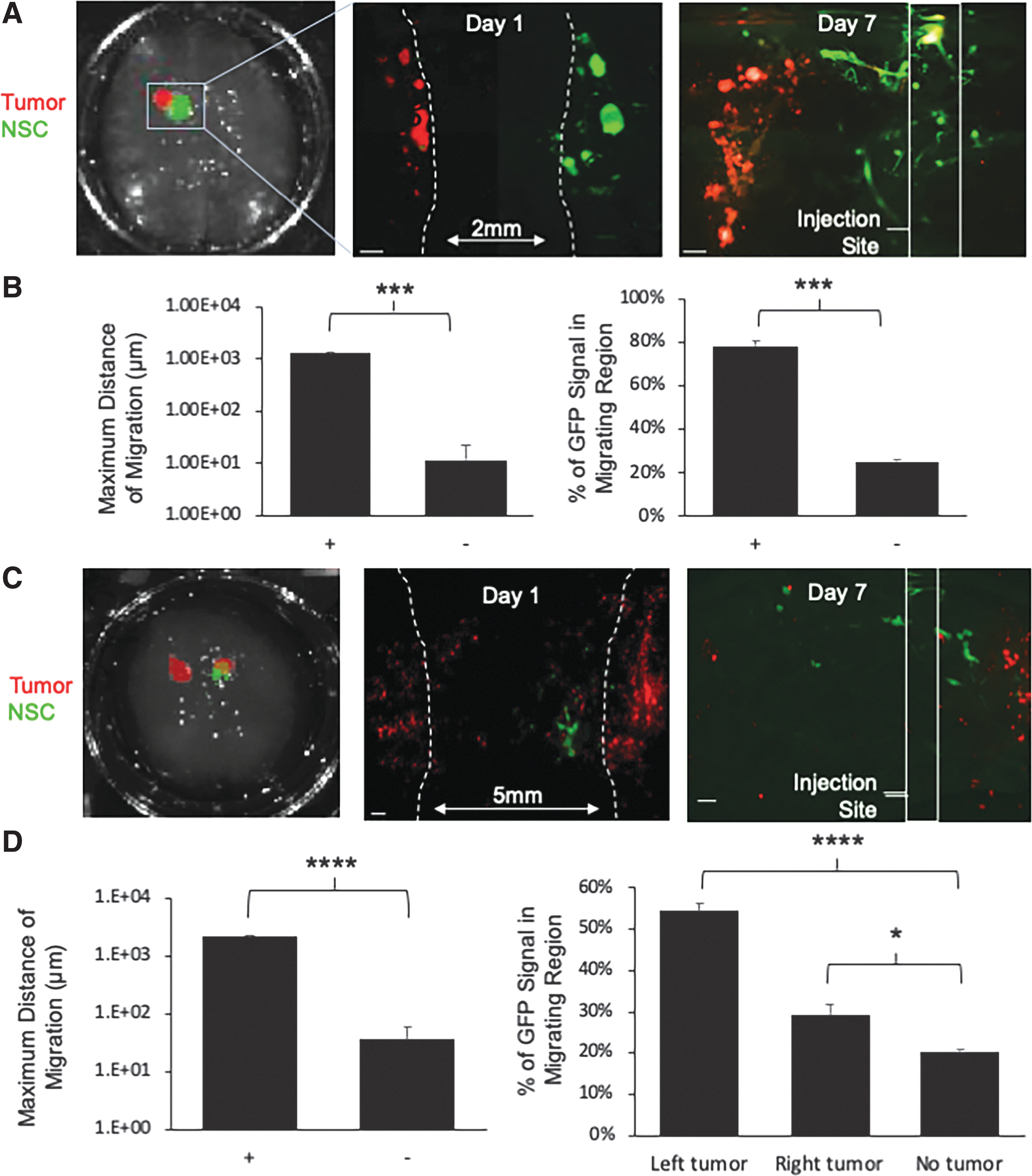
NSC tumor homing in BBMs.
In the clinic, metastatic brain cancer often presents as multifocal disease. To investigate NSC migration to multifocal tumors, we next implanted mC-labeled 231-BR cells on contralateral hemispheres of the mini-BBMs ∼5 mm apart. Three days later, NSCs were implanted ∼1 mm medially to the right tumor (RT), as shown in Figure 4C. After 1 week of coculture, mini-BBMs were processed and fluorescent micrographs were captured and analyzed in ImageJ as before. Analysis showed that NSCs migrated 61-fold further in the presence of tumor compared with nontumor BBM, traversing 2.2 ± 0.1 mm of the 4 mm gap between the NSC injection site and left tumor (LT) implant site over the 7-day period (Fig. 4D). Quantification of NSCs at the injection site or near the tumor foci further confirmed this migration, as analysis of the GFP intensity showed that 54% ± 1.8% of NSCs homed directionally toward the LT, which was, interestingly, further from the initial injection site compared with the RT. Around 29.2% ± 2.4% of NSCs homed directionally toward the RT. Both enabled significantly more NSC migration than nontumor BBMs, where only 20% ± 2.0% of GFP signal could be found outside the injection site region (Fig. 4D). These results validate the tumor-homing property of NSCs and the usefulness of mini-BBMs for interrogating this key aspect of NSC therapy.
Investigating the impact of distance on NSC-induced tumor kill
In human patients, brain cancer foci are likely to present at varying distances from the site of NSC implant or infusion. However, the impact of tumor distance on NSC therapy is not easily investigated in vivo as the small size of the murine model makes implanting multiple tumor foci challenging and limits the distances that can be investigated. To use the BBM system to explore the impact of tumor distance on NSC therapy, U87-mC were implanted in the mini-BBMs and NSCs engineered to express the antitumor agent TRAIL or control GFP were implanted at 0, 2.0, 5.0, or 10 mm laterally from the tumor (Fig. 5A, B). Tumor progression was tracked with weekly kinetic imaging of U87-mC fluorescence for 1 month. As shown in the summary graph (Fig. 5C), NSCs delivered directly into the U87 tumor foci rapidly attenuated tumor progression, reducing tumor volumes to subdetection levels by day 10 posttreatment and sustaining suppression through 28 days. NSCs implanted 2 mm from the tumor also significantly attenuate tumor growth by day 14, with decrease to subdetection levels achieved by day 21. Attenuation of tumor growth from NSCs implanted 5 and 10 mm was also found to begin at day 14, however, reduction to subdetection levels was not achieved until day 28 posttreatment (Fig. 5C). On day 14, no statistical significance was observed when comparing the sham group to the 2-, 5-, and 10-mm groups. Statistical significance was observed on day 21; compared with the sham control, the p-values for the 2-, 5-, and 10-mm groups were calculated to be 0.00084, 0.001809, and 0.004413, respectively. On day 28, the p-values for the 2-, 5-, and 10-mm groups were calculated to be 0.00059, 0.000667, and 0.000973, respectively, when compared with the sham control.
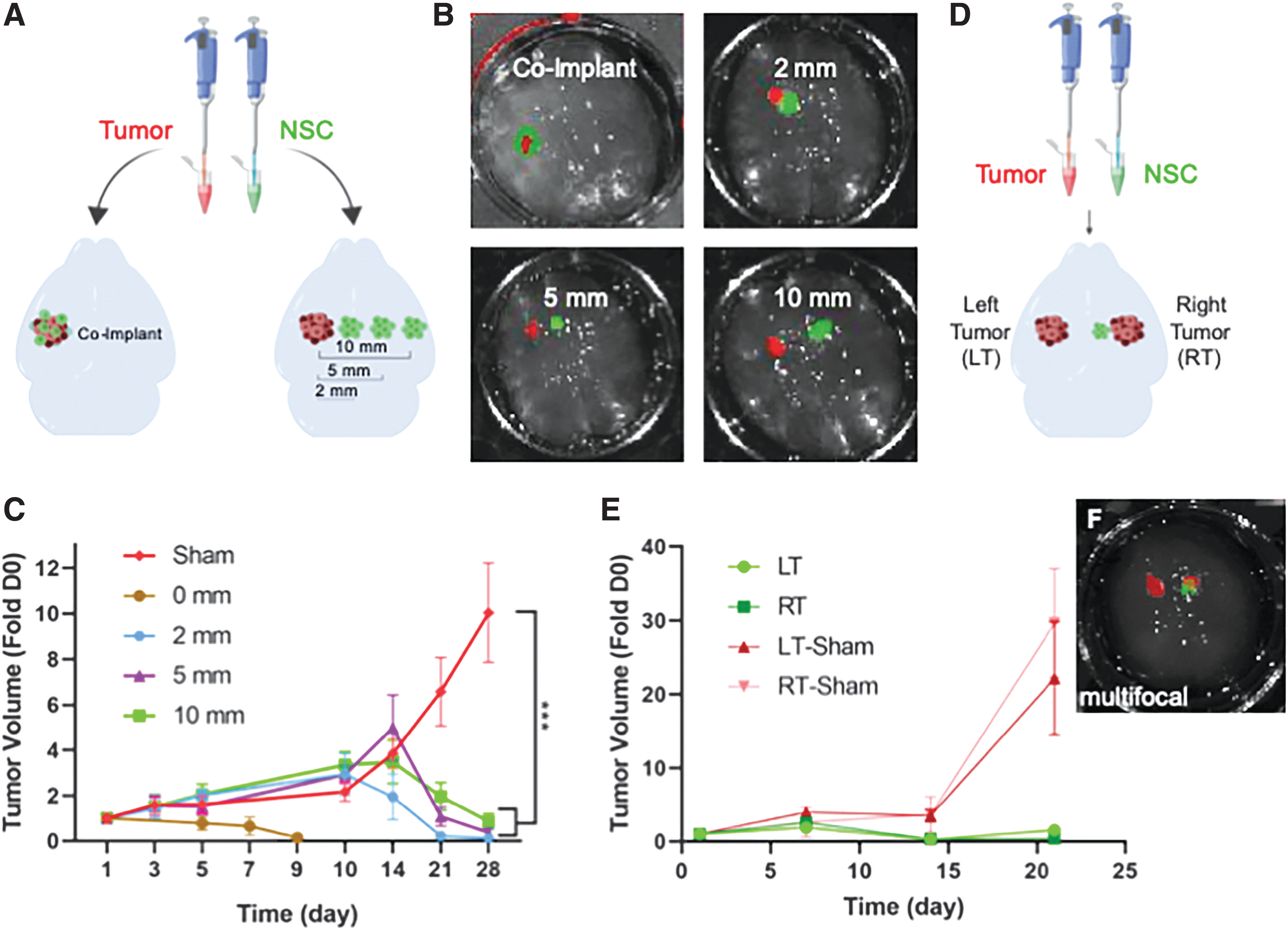
Efficacy of NSC-TRAIL as a treatment modality determined using different disease models in small BBMs.
Lastly, we explored the potential efficacy of NSC therapy for multiple tumor foci. NSC migration is dependent on chemotaxis, thus it is possible that multifocal tumors may split the dose of therapeutic NSCs across multiple foci, resulting in altered killing. To investigate this relationship, 231-BR-mC cells were implanted in each hemisphere of the mini-BBM, ∼5 mm apart. NSC-TRAIL or NSC-GFP were implanted directly adjacent to the tumor in the right hemisphere (RT), and tumor volumes were measured serially for 3 weeks (Fig. 5E). As shown in the summary data, serial kinetic imaging showed that NSC-TRAIL treatment reduced both tumor foci by greater than 70% 14 days posttreatment despite the closer proximity to the GBM in the right hemisphere (Fig. 5E). Interestingly, imaging results showed the tumor in the left hemisphere (LT) began to trend toward regrowth at day 21, and the change in signal was statistically significant compared with the GBM in the right hemisphere (Fig. 5E). In contrast, tumors in both hemispheres expanded rapidly in BBMs implanted with control NSC-GFP.
BBM fabrication and bioreactor design
Transmittance was calculated using light absorbance values of the mini-BBMs. Percent transmittance was determined to be 11.96 ± 4.43 and 23.17 ± 3.71 in mini-BBMs with and without PLA fibers, respectively (Fig. 2G).
Discussion
Engineered NSC therapy is a promising new approach to cancer therapy. Multiple aspects of NSC migration and killing are impacted by the microenvironment, making the ideal potency assay for NSC-based therapeutics for brain cancer models that recapitulate the physical properties of brain tissue. With this in mind, BBMs were made to physically resemble the brain perivascular niche through embedding of microfiber PLA.26–30 PLA was chosen due to its biocompatibility as it has been used in several cell migration assays and tissue regeneration studies.30,31 Agarose as a base also presents a number of advantages. First, low concentration agarose gels (0.2–0.6%) have been shown to mimic the poroelasticity and backflow of the brain.32,33 Second, its deformability makes it easily injectable, and collective cell migration, which is more relevant to cell behavior in vivo, can be studied.34,35 Third, it is considerably less expensive than other hydrogels designed to mimic the tissue microenvironment, such as collagen12,15,36 or Matrigel.11,37 Fourth, the Young's modulus of soft agarose closely mimics that of healthy brain tissue; this is critical, because several studies have reported that the viscoelastic properties of a substrate can impact cells' proliferation, differentiation, and migration characteristics.38–41 While agarose shows promise in this study, future optimization of BBMs will include exploration of other transparent matrices, such as alginate. Lastly, the transparent nature of the BBM allows for accessible serial imaging using in vivo fluorescent imaging systems (IVIS). IVIS imaging is efficient at locating the general presence of cells in a 2D space. To better understand the 3D spacing of cells, we performed initial experiments that explored the use of light-sheet microscopy (LSM) and confocal microscopy. For LSM, the BBM samples were prepared using the iDISCO+ protocol and stained with validated antibodies. 42 However, NSCs were not visualized; this was likely caused by the inability of the antibodies to diffuse through the dense BBM. Confocal imaging was also nonideal due to the size of the sample and the spread of the cells in the x-y plane. Future studies combining BBMs with different imaging modalities would further optimize the BBM system and also provide unique insights into cellular and molecular events that occur during both the progression of brain cancers and following response to therapies.
As a result, rationally designed BBMs were able to model both tumor growth and NSC tumor homing that occurs in vivo in nude mouse xenograft models of brain cancer.19,25,43 From there, experiments using BBMs revealed the impact of distance between NSC and tumor foci on therapeutic efficacy. Clinically, GBM tumor regrowth occurs within 2 cm of the tumor resection cavity. 44 Therefore, the distances of 2, 5, and 10 mm were selected to mimic these clinical findings. When implanting therapeutic NSCs and GBM, onset and extent of tumor suppression varied inversely with initial NSC distance from the established tumor focus. While therapeutic NSCs implanted into 231-BR models did not impact the timing of the onset of tumor suppression, the tumor contralateral to the NSC initial injection site began to recur <2 weeks later. More investigation into the mechanism for recurrence is needed, but it could be due to metastatic invasion and therefore escape from the region of effect created by diffusion of TRAIL secreted by therapeutic NSCs.5,12,32,45,46 Additionally, while SDF-1, CXCR4, VEGF, and EGF are believed to play roles in NSC migration, the exact mechanism has yet to be elucidated. 17 To maximize efficacy of NSC-based therapies, optimization of dosing strategies is needed; more research into the effects of cell density, total cell dose, frequency of dosing, and potential for cell conditioning, among others, on cell migration, and therefore potency, must be conducted.
In the case of autologous NSC-based therapies, a method for high-throughput screening of migration kinetics between cells derived from different patients is needed to determine reduction to practice. We present this multiscale 3D biomimetic model of brain cancer to fulfill this need. It is our hope that ongoing studies of efficacy using NSC-based therapies against GBM in life-size BBMs can accurately reflect what can be expected as this therapeutic approach is adapted for clinical use. Using this method to examine the effect of dosing strategy on therapeutic cells at a human-size scale should provide more reliable data than current commonly used approaches, as cells prefer to migrate through a soft matrix rather than through a membrane or across a rigid surface. 13
Footnotes
Acknowledgments
The authors would like to thank the Biomedical Research Imaging Core at the University of North Carolina at Chapel Hill. The authors also gratefully acknowledge Devin Hubbard, PhD for his assistance with the design and fabrication of the bioreactor.
Disclosure Statement
S.D.H. has ownership interest and is the CSO of Falcon Therapeutics. The remaining authors declare no competing interests.
Funding Information
This work was funded by the Eshelman Institute for Innovation at the University of North Carolina at Chapel Hill.