
Abstract
Mammalian platelets participate in the immediate tissue injury response by initiating coagulation and further promoting tissue injury mitigation and repair. The latter properties are deployed following platelet release of presynthetized morphogens, cytokines, and growth and chemotactic factors, which launch a tissue regenerative, angiogenic, and anti-inflammatory program. Several blood-derived biologic products, like platelet-rich plasma (PRP) and platelet lysate (PL), are currently on the market to allow proper healing and tissue regeneration. However, not all growth factors are released from the platelets and the final products contain plasma proteins such as albumin, fibrinogen, complement, and immunoglobulins, increasing the risks of serum sickness or allergic reaction. To address this problem, we developed a new platelet extract where equine blood platelets are concentrated, washed, and thereafter lysed by detergent Triton X-114. Distinct from PRP, this extract is devoid of albumin, fibrinogen, and immunoglobulins and is 266-fold enriched in platelet-derived growth factor content relative to PRP. Washed equine platelet extract (WEPLEX) is amenable to lyophilization without loss of biological activity. In vitro, WEPLEX significantly inhibits human and equine T cell proliferative response to phytohemagglutinin and also polarizes murine CD45+/CD11b+ peritoneal macrophages to an IL-10+ M2-like phenotype. In vivo, WEPLEX substantially improves clinical outcome of murine experimental dextran sulfate sodium colitis. We propose that equine-sourced, zoonosis-free WEPLEX may serve as an anti-inflammatory biological therapy across mammalian species.
Impact statement
This research described the manufacture and pharmaceutical activity of a novel biological product WEPLEX, Washed Equine Platelet Extract. This extract is a washed allogeneic/xenogeneic platelet extract amenable to lyophilization and devoid of potentially immunogenic plasma-derived proteins with biological anti-inflammatory activity across mammalian species. WEPLEX use as a pharmaceutical may have broad and significant impact, both in veterinary medicine and human medical field, on acute injury of tissue, regenerative medicine, and in other medical fields where platelet derived products have been used such as dermatology, cardiology, and sports medicine.
Introduction
Experimental and clinical data established platelets to be critical in physiopathological processes such as inflammation, tissue regeneration, and revascularization. 1 In vivo, platelets are activated at the site of injury and release cytokines and growth factors are stored in the dense membrane of the α-granules. 2 These molecules, when bound to the proper receptors, promote expression of genes involved in cellular recruitment, growth, and morphogenesis. 3 Other molecules present in the α-granules (e.g., adhesive proteins, clotting factors, and fibrinolytic factors) are involved in the inflammation processes. 3 In general, activation of platelets allows the release of a large number of soluble mediators, which are essential in wound healing, clotting, and angiogenesis,1,2,4 hence the interest in platelet-rich plasma (PRP) derivatives for regenerative medicine and tissue engineering applications. 4
In the past 40 years, the pharmaceutical use of PRP has been extensively investigated.3,5 PRP was first defined as plasma with a platelet count above that of peripheral blood. 6 Ten years later, autologous PRP was used in maxillofacial surgery for its high content in fibrinogen and anti-inflammatory characteristics able to induce cell proliferation. 7 Later on, autologous PRP has been used predominantly in the musculoskeletal field in sport injuries. 6 Other fields of PRP use include cardiac surgery, pediatric surgery, gynecology, and more recently, dermatology.6,8–11
A later formulation aimed to improve the potency of PRP was the so-called platelet lysate (PL) obtained by a simple step of freezing and thawing PRP with the intent of disrupting platelets contained therein and further centrifugation to remove any residual platelet fragment.5,6 More recently, mechanical fibrinogen depletion 12 and controlled clotting procedure with CaCl2 13 have been validated to remove fibrinogen from the lysate without compromising the final growth factors and cytokine composition.
Allogeneic and autologous PRP have been tested in human medicine for tendinopathies like rotator cuff repair,14,15 ophthalmology, 16 nonhealing ulcers, 17 and persistent fracture nonunions. 18 On-going clinical trials are investigating its use in adhesive capsulitis of the shoulder (NCT01458691) and endometrial regeneration (NCT02825849). PL has been tested for epicondylitis 19 and ophthalmology. 20 Similarly, PRP has been used in veterinary medicine for the treatment of sport-related injuries in horses such as tendon and ligament injuries,21–23 and wound healing. 24 Despite the promise of PRP and PL, properly designed clinical trials with PRP and PL have had mixed outcomes for either human medicine or veterinary medicine.21,25–27
Both PRP and PL contain typical plasma proteins such as albumin, complement, and immunoglobulins that remain present in the final formulation dilute in serum. One issue found in the use of PRP and PL is the large volume of serum and the possible immunogenicity of allograft plasma protein material. 28 In some cases, the volume of serum that needs to be injected into an area is too large for the anatomical region, causing a less than ideal amount of serum injected to treat the injury. Furthermore, immune reactions can be caused using this method due to immunogenicity of allogeneic plasma proteins [akin to transfusion reactions to allogenic serum proteins], as well as transfer of diseases.28,29
We here seek to address these limitations of autogeneic/allogeneic PRP and PL in an entirely novel manner by generating a lyophilized extract of washed equine platelets. Equine platelets have been chosen over human and other mammal platelet products derived from bovine, ovine, and porcine sources. Sourcing of random donor volunteer human platelets amenable for manufacture at scale can be challenging, combined with the burden of rigorous pathogen testing required to assure safety from known and arising blood-borne diseases. Distinct from other hoofed mammals, the Equine genus is bereft of endogenous or endemic zoonosis known to afflict human. Furthermore, the ultrastructure of equine platelets is similar to that of the human and their size and shape are more uniform in respect to other mammals like human, bovine, and porcine. 30 Compared to other mammals, equine whole blood platelet counts are some of the lowest reported, yet share similar metabolic pathways resulting in qualitative composition similar between different species. 30
The virtual totality of beneficial morphogens and anti-inflammatory factors contained in PRP is derived from the platelets contained therein. However, platelets constitute a small percentage (<0.5%) of the volume of PRP, which is mostly (>99%) plasma on a volume basis. 6 Furthermore, plasma contains complement, immunoglobulins, albumin, and fibrinogen, all of which render allogeneic PRP prone to immune/allergic response. To address these issues, we propose that washed platelets from which growth factors are extracted would allow for the production of a biological product more concentrated in platelet extract (relative to standard PRP or PL) and also devoid of any plasma protein that may cause an immune reaction. This washed equine platelet extract (WEPLEX) can be further lyophilized allowing for volume reduction without affecting its biological properties. Our data support the idea that xenogeneic equine platelet-derived extract may serve as an anti-inflammatory and regenerative pharmaceutical.
Methods
Ethics statement
All procedures performed in studies involving human samples were in accordance with the ethical standards of the Institutional Review Board at UW – Madison (IRB 2019-0865). Blood was collected from healthy adults, 18–55 years old, who did not donate more than one unit of blood (450 mL) in the previous 2 months. All animals were handled and treated according to the guidelines and recommendations of the Research Animal Resources Center and the Animal Care and Use Committee at UW–Madison. Horses from which whole blood was collected were donated to the Veterinary Medical Teaching hospital, at UW–Madison, and housed at the Morrie Waud Large Animal Hospital. C57BL/6 and Vert-X mice were purchased from The Jackson Laboratory and housed in microisolator cages at the BSL2 vivarium facility of the Wisconsin Institute of Medical Research. Mice, both female and male, were used between 6 and 20 weeks of age. Mice were euthanized by asphyxia.
Collection of equine blood and isolation of platelets
Blood was collected by phlebotomy from the jugular vein from 14 horses (range, 1–33 years; mean 21 years; 6 mares and 8 geldings; 8 Quarter Horses, 2 Peruvian Horses, 1 Paint, 1 Belgian Warmblood, 1 Haflinger, and 1 Appaloosa) housed at the University of Wisconsin Morrie Waud Large Animal Hospital, without performing a sample size calculation related to breed or gender. Blood collection bags containing anticoagulant Citrate Phosphate Dextrose Adenine Solution, USP (CPDA-1), were used to collect 2 liters of blood from each horse. Each collection bag (525 mL) consists of a PL 146 Plastic primary container with 63 mL of CPDA-1 solution containing 2 g dextrose (monohydrate) USP, 1.66 g sodium citrate (dihydrate) USP, 188 mg citric acid (anhydrous) USP, 140 mg monobasic sodium phosphate (monohydrate) USP, and 17.3 mg adenine USP, to prevent coagulation. After obtaining whole blood in CPDA-1 bags, the unit was transferred in 250 mL conical tubes, under sterile conditions. The first step included the preparation of PRP by centrifugation of the anticoagulated blood at 800 g for 5 min, room temperature, with no brake applied. After the transfer of the PRP supernatant in new tubes, a second centrifugation at 800 g for 20 min at room temperature allowed the pelleting of the platelets from the plasma. The supernatant was discarded, and the pelleted platelets were washed twice with Dulbecco's phosphate-buffered saline (DPBS) without calcium or magnesium. Washed platelet concentrate devoid of equine plasma was subsequently processed for detergent lysis.
Lysis buffer determination to manufacture WEPLEX
To prepare PL, different detergent-containing solutions were tested. Lysis buffer composition was assessed by testing different % v/v of detergent in DPBS (Triton X-100, Triton X-114, NP-40, TWEEN-20, and CHAPS at concentration up to 2% v/v). To maximize the concentration density of the platelet extract, the smallest possible volume of lysis solution to allow full solubilization of the platelets was added to the washed platelet concentrate for 30–60 min at room temperature. After lysis, the insoluble material was removed by centrifugation at 1000 g, at 4°C. Removal of detergent was achieved on spin columns, DetergentOUT™ (G-Biosciences, St. Louis, MO), following manufacturer's specifications. The biochemical attributes of the different lysed platelet extracts were tested for total protein content and concentration of platelet-derived growth factor (PDGF) (R&D system, Minneapolis, MN) and the presence of plasma proteins such as total equine IgG (USBiological, Salem, MA) by ELISA, fibrinogen by mechanical clot detection/Stago Compact Max (Marshfield Labs, Middleton, WI), and albumin was measured at University of Wisconsin Veterinary Care, Diagnostic Pathology Services. The presence of endotoxin was evaluated by LAL Chromogenic Endotoxin Quantitation Kit (Pierce Biotechnology, Rockford, IL) in every final lysate before in vitro cell culture experiments.
A speed-vac from Eppendorf was used to lyophilize the sample. The sample was placed in 1.5 mL Eppendorf tubes with open cap and frozen on dry ice before using the speed-vac to prevent loss of material from bumping. Under low-pressure vacuum, the vapor-liquid equilibrium of the solvent is shifted toward the gas phase, while the proteins and molecules of WEPLEX remain in the solid phase. After all the solvent was removed by the process, a solid dry sample was left (plus salts that were present in the solvent buffer). The sample can then be resuspended in the desired amount of any buffer/water. Usually, 1 mL aliquot required 5 h to be fully lyophilized. To assess the biochemical characteristics of the lyophilized/reconstituted WEPLEX sample in respect to starting materials, total protein concentration and PDGF content were measured by BCA assay kit and ELISA, respectively, as described above.
Isolation of peripheral blood mononuclear cells and activation of T cells
Equine and human peripheral blood mononuclear cells (PBMCs) were isolated by the modified protocol described by Lan et al. 31 Briefly, anticoagulant-treated blood, diluted twofold on DPBS, was carefully layered on top of one volume of Ficoll-Paque PLUS density gradient media (GE Health Care) in a tube. The tube was then centrifuged 400 g for 30 min at 25°C. This first centrifugation step generated three layers: the upper layer was plasma, the intermediate layer containing mononuclear cells, and the bottom layer, which consisted mostly of erythrocytes and polymorphonuclear granulocytes. The plasma/Ficoll medium interface contains both PBMCs and platelets. To isolate PBMCs, this interface was carefully removed, washed with a salt-buffered solution, and then centrifuged. The supernatant, containing platelets, Ficoll, and plasma, was removed, leaving a pellet of purified PBMCs.
PBMCs were cultured in 12 or 24 tissue plates (Corning®) at a density of 1 million cells/mL in complete R10: RPMI-1640 with L-Glu (Corning), 10% fetal bovine serum -FBS (Sigma, St. Louis, MO), 2 mM L-Glutamine (Lonza), 10 mM HEPES (Lonza), 0.1 mM beta-mercaptoethanol (Gibco), and the addition of 100 U/mL penicillin/streptomycin.
T cell activation was achieved by stimulating T cells with phytohemagglutinin-L (PHA-L), Invitrogen™. 32 Cells were cultured for 3 days with PHA-L solution (500 × ) at 2 μL per mL of culture medium.
T cell proliferation assay
Equine PBMCs were labeled with 5 μM carboxyfluorescein succinimidyl ester (CFSE; Molecular Probes, Eugene, OR) in 1 mL of PBS for 15 min at 37°C. The labeling was halted by adding an excess of fetal calf serum (FCS), and the samples were washed twice with RPMI-1640 (Corning) supplemented with 10% FBS (Sigma). The CFSE-labeled cells were cultured, with a range of WEPLEX (1–10% v/v), fresh or freshly reconstituted lyophilized. The T cell CFSE fluorescence intensity was analyzed by flow cytometry after 3–4 days of co-culture and proliferation. The analysis of CD8 T cells was performed with the use of anti-equine CD8 (Invitrogen) flow cytometry antibodies. Phytohemagglutinin (PHA)-activated CD8 T cell proliferation was compared to the proliferation of WEPLEX-treated PHA-activated CD8 T cells. Unstimulated (no PHA) CD8+ T cells served as a control for basal proliferation (less than 1%).
For human T cells, Ki67 from BD biosciences was used as a means to assess proliferation following PHA activation. PHA-activated T cells were treated with a range of WEPLEX (1–10% v/v), fresh or freshly reconstituted, for 72–96 h and subjected to intracellular Ki67 staining according to the manufacturer's instructions (BD Biosciences), and identified by flow cytometry. Anti-human CD3 antibody (Invitrogen) allowed identification of T cells. PHA-activated PBMCs served as positive control for maximum T cell proliferation and unstimulated (no PHA) PBMCs to define basal T cell proliferation (1%).
Murine peritoneal macrophage isolation and analysis
Mice (C57BL/6 or Vert-X, either female or male 8–12 weeks old) were anesthetized with isoflurane and peritoneal cells were immediately isolated by washing the peritoneal cavity with sterile DPBS containing 10% FBS. To obtain monolayers of peritoneal macrophages, the cell concentration was adjusted to 5–10 million cells/mL in complete R10 media. The cells were allowed to adhere to the plate by culturing them for 16 h at 37°C. Nonadherent cells were then removed by gently washing a few times with warm PBS. Complete R10 was added to the tissue plate with the addition of a range of WEPLEX (0.01–0.1% v/v) for 24 h.
Expression of IL-10 secreted in the culture media was measured by ELISA (Invitrogen). Expression of IL-10 by flow cytometry was carried using peritoneal macrophages from GFP/IL-10 reporter mice (Vert-X mice) and by staining for CD45 and CD11b antibodies (Invitrogen).
Dextran sulfate sodium-induced colitis in mice
Colon inflammation was induced by the administration of dextran sulfate sodium (DSS) in the drinking water. 33 To induce colitis, DSS (molecular weight 5000; MP Biomedicals, Inc., Eschwege, Germany) was dissolved in ultrapure water at a concentration of 4%. C57BL/6 (either female or male 8–12 weeks old) mice were exposed to the DSS solutions ad libitum for 7 days. After 7 days, DSS water was changed with fresh water. Mice were randomly allocated into four groups (n = 5 each). Fresh WEPLEX was prepared and lyophilized WEPLEX was reconstituted in the same volume from which it was generated. Mice received 1 mL intraperitoneal (IP) injection of WEPLEX (fresh or lyophilized/reconstituted) or PBS on day 2 and 4. The body weight was measured daily for each mouse and is depicted as the percentage of the initial value measured before the onset of the experiment. Experiments were blinded and repeated thrice.
Statistical analyses
All statistical analyses were performed using Prism 8 Software (GraphPad Software, San Diego, CA). Unless otherwise indicated, tests of statistical significance were conducted using a two-tailed Student's t test. Data display normal variance. p-Values <0.05 were considered to be statistically significant.
Results
Optimization and characterization of extract derived from washed equine platelet concentrate
Two liters of equine whole blood was collected from individual horses by phlebotomy at the School of Veterinary Medicine, University of UW–Madison, in CPDA-1 bags. Undiluted whole blood was transferred to a 250 mL conical tube under sterile conditions and centrifuged at 800 g for 5 min at room temperature. This low-speed centrifugation allows the separation of erythrocytes and leukocytes from the PRP. Two distinct layers could be observed: bottom layer consisting of red blood cells and granulocytes (accounting for 50–80% of the total volume), and a top layer with straw-colored PRP. After centrifugation, the PRP above the red blood cell layer was transferred into new tubes. A second centrifugation at 800 g for 20 min at room temperature allows the separation of the platelets (as a pellet) from the plasma. The supernatant was discarded, and the pelleted platelets were washed twice with DPBS without calcium or magnesium to remove any residual plasma. Pelleted platelets are stored at −80°C for later processing.
The next step involved platelet disruption with a detergent that would allow for maximal yield of protein content (PDGF served as a surrogate) and could serve as a contemporaneous viral inactivation step (for enveloped viruses). To identify an optimal lysis buffer, the protein extraction efficiency from the platelets was tested with an array of detergents amenable to latter good manufacturing process processing (Table 1). We tested Triton X-114 (0.5–2%), CHAPS (1–0.1%), NP-40 (0.1–2%), and TWEEN 20 (0.1–2%) in DPBS (Table 1). The detergent conditions were defined for maximal release of proteins mg/mL (e.g., CHAPS 1%, TWEEN 20 1%, NP40 1%, and Triton X-114 2%) for further biochemical characterization (Table 1). Table 2 shows the biochemical characterization of PL for every detergent tested. Pelleted platelets derived from 100 to 200 mL of PRP were lysed for 1–2 h at 4°C and insoluble components were removed by centrifugation 4000 g at 4°C. Removal detergents from extracts were carried out with DetergentOUT spin column following manufacturer's instructions.
Protein Released Following Platelet Lysis in Dilutions (v/v %) of Tested Detergents
Data represent mean ± SD of three different preparations.
Protein concentration detected with BCA Protein Assay.
SD, standard deviation.
Detergents and Platelet Lysis Protein Yields
Data represent mean ± SD of three different preparations.
Protein concentration detected with BCA Protein Assay.
Total equine IgGs.
PDGF concentration detected by ELISA.
Measured at Marshfield Labs.
University of Wisconsin Veterinary Care, Diagnostic Pathology Services.
N.D., not detected; PDGF, platelet-derived growth factor.
Following detergent removal, total protein content was measured by BCA assay, PDGF and IgG by ELISA, and fibrinogen and albumin (Table 2). PDGF yield efficiency was calculated as amount of PDGF per total protein in respect to the one obtained from PRP (Table 2). Among tested detergents, Triton X-114 2% was most efficient in yielding a protein-rich platelet extract 266-fold higher than PRP (Table 1). Equine IgG was only detected in the PRP (150 ± 5 mg/mL) and no trace of equine IgG could be detected in the extract derived from washed platelets, independent of the lysis detergent used. Similarly, no albumin or fibrinogen could be detected in the equine platelet extract generated with 2% Triton X-114 (Table 2). No endotoxin was detected in the final preparations (detection limit of the kit was 0.06 EU/mL). This final formulation is hereafter referred to as WEPLEX.
Lyophilization of WEPLEX
Lyophilization of WEPLEX was carried out by freezing and drying with Eppendorf's speed-vac. Throughout the lyophilization process, the WEPLEX solution remained clear and without any precipitate. The final rendered powder was stored at room temperature and reconstituted in 1 mL of deionized water before testing. Microporous filtration with a membrane with 0.2 μm pore size was used to maintain sterility and prevent contamination postlyophilization of the reconstituted WEPLEX solution.
Total protein and PDGF contents were measured. No significant difference in protein concentrations (3.5 vs. 3.1 mg/mL for the fresh and lyophilized/reconstituted WEPLEX, respectively) and PDGF (87 vs. 78 ng/mL for the fresh and lyophilized/reconstituted WEPLEX respectively) was noted between the two formulations. The overall yield intended as amount of PDGF relative to total proteins was identical (25 ng PDGF/mg protein) (Table 3).
Comparison of Fresh and Lyophilized and Reconstituted Washed Equine Platelet Extract
Freshly prepared WEPLEX was divided in to two different aliquots. One aliquot was lyophilized and the other kept at 4°C. The day after the lyophilized sample was reconstituted in deionized water and filter sterilized, protein concentrations and PDGF content were measured. Data represent mean ± SD of three different preparations.
Protein concentration measured with BCA Protein Assay Kit.
PDGF concentration detected by ELISA.
WEPLEX, washed equine platelet extract.
T cell suppression assay with WEPLEX
To measure the ability of WEPLEX to suppress inflammation, a T cell suppression assay was carried out. PHA-activated PBMCs, incubated with or without WEPLEX, were collected and analyzed by flow cytometry (Fig. 1A–C). The proliferation of equine PBMCs was tested by labeling the cells with CFSE before stimulation and thereafter assessing dye dilution as a surrogate of replication (Fig. 1A, C). Concentration as low as 1% v/v of WEPLEX was able to significantly decrease the proliferation of equine CD8+ T cells (41.8% CD8+CFSE+ proliferating T cells with respect to 34.9% CD8+CFSE+ with 1% v/v WEPLEX, Fig. 1A). Full suppression of proliferation was achieved with 10% v/v of WEPLEX added to the culture media (2.4% CD8+CFSE+, Fig. 1A). Similar results were observed with the lyophilized/reconstituted WEPLEX formulation (Fig. 1A, C).

T cell suppression assay.
For human T cell proliferation, PBMCs were labeled post-PHA activation with Ki-67 (Fig. 1B, C). Concentration of 1% v/v of WEPLEX, both with the fresh or lyophilized/reconstituted WEPLEX formulation, decreased the proliferation of human CD3+ T cells more than threefold with respect to the PHA-stimulated control (12.5% CD3+Ki67+ proliferating T cells with respect to 4.1% CD3+Ki67+ with 1% v/v of WEPLEX, Fig. 1B, C). Full inhibition of human T cell proliferation was achieved with concentration of 4% v/v of WEPLEX (0.7 CD3+Ki67+, Fig. 1B, C).
WEPLEX and M2 polarization of mouse peritoneal macrophages
We tested the ability of WEPLEX to convert resting peritoneal macrophages to an IL-10+ M2-immunosuppressive like phenotype. Peritoneal cells from Vert-X mice were cocultured with 0.1–0.01% v/v of WEPLEX. After overnight culture, the overlying media were tested for IL-10 secretion and cells were stained for CD45 and CD11b and screened for the co-expression of GFP (IL-10) by flow cytometry (Fig. 2). As a positive control for macrophage activation, macrophages were treated with lipopolysaccharides (LPS) for 24 h. Macrophages treated with LPS secreted, in the culture media, the highest amount of IL-10 (1.1 ng/mL), while macrophages treated with 0.01% or 0.1% v/v WEPLEX secreted 0.3 ng/mL of IL-10. These cells also displayed an increase in frequency of GFP+/IL10+ macrophages (21.2% CD11b+CD45+GFP+ for 0.01% v/v of WEPLEX, Fig. 2). IL-10 could not be detected in the macrophages cultured in complete media without WEPLEX.
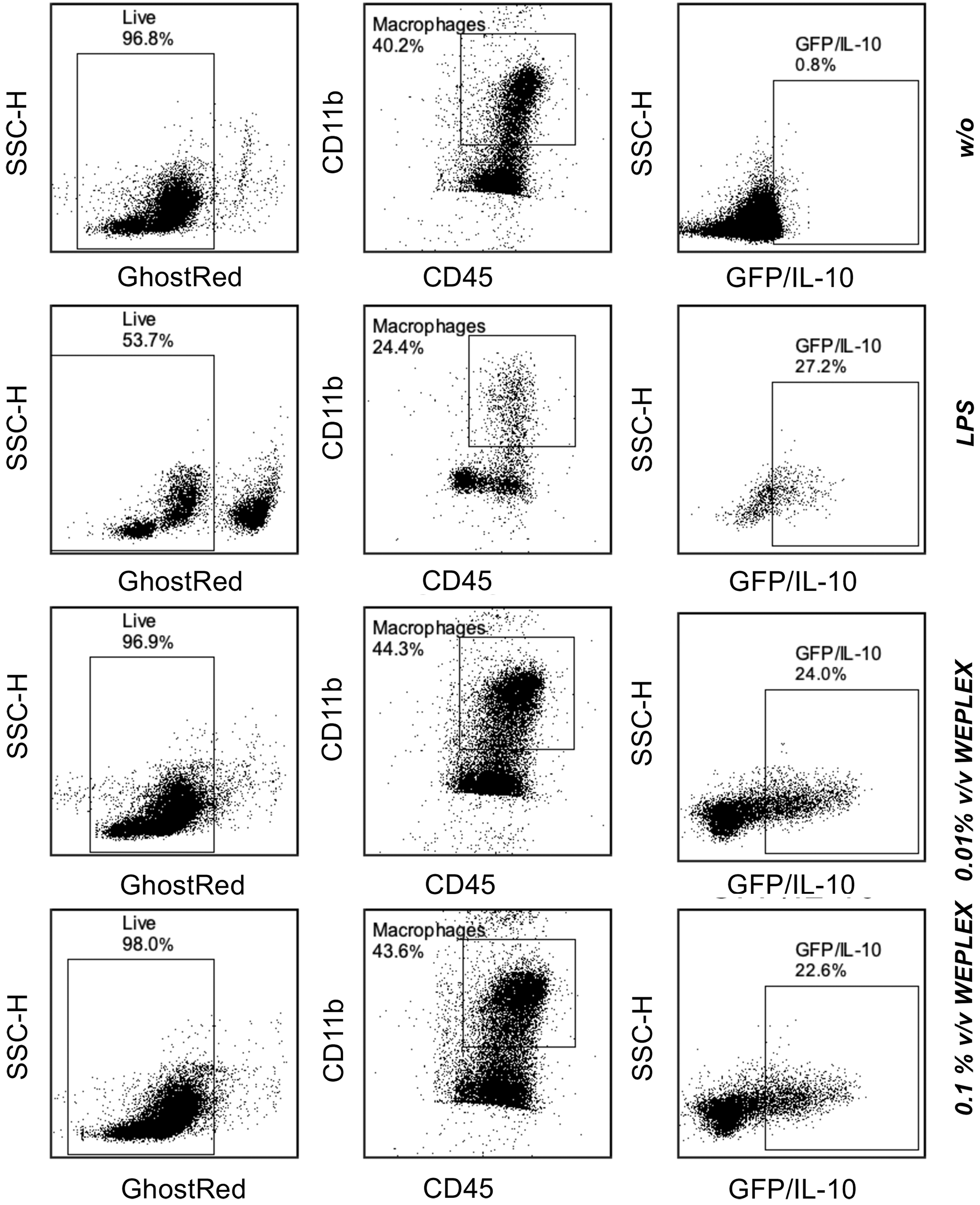
GFP/IL-10 expression in macrophage cells isolated from Vert-X mice. Live cells were selected as Ghost-reddim. Macrophages were selected as double positive CD45+CD11b+. Gate of GFP+/IL10+ cells was established in C57BL/6 mice.
Treatment of DSS-induced colitis in mice with WEPLEX
We used murine DSS-induced colitis as an acute tissue injury model to test the therapeutic pharmaceutical properties of WEPLEX administered parenterally. Mice were allocated in four groups and the relative change in body weight and survival for the different groups is shown (Fig. 3). The first two groups received either fresh WEPLEX or PBS (Fig. 3A), and the other two groups lyophilized/reconstituted WEPLEX or PBS (Fig. 3C). In the first 3–4 days of the experiment, no significant difference was detected between WEPLEX-treated (either fresh or lyophilized/reconstituted) or PBS-treated mice. From day 9 on, both WEPLEX-treated groups showed a significant increase in body weight compared to the PBS-treated groups (Fig. 3A, C). An improved survival in both WEPLEX-treated groups compared to PBS-treated mice was observed (Fig. 3B, D).
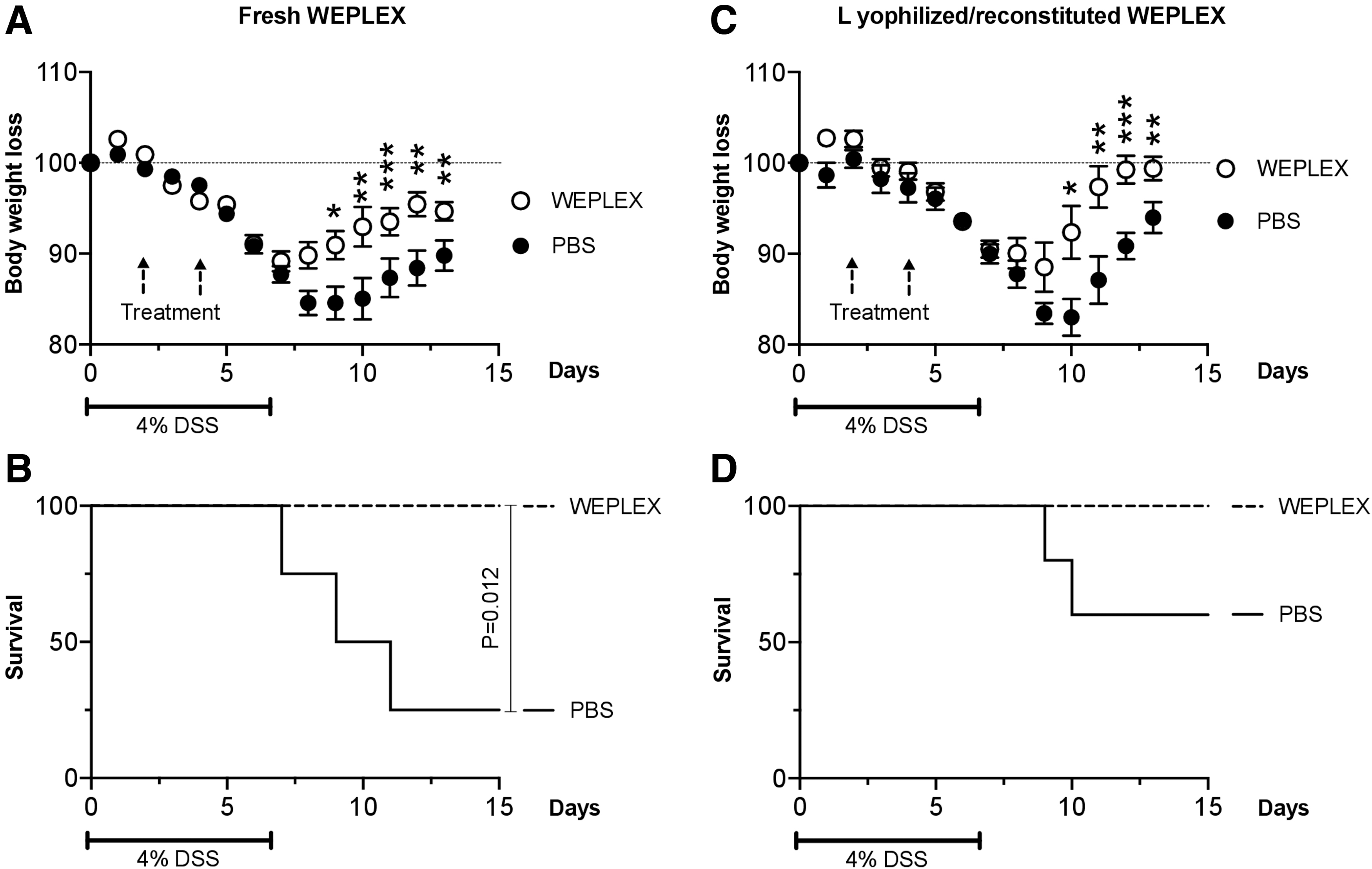
Effects of IP injection of WEPLEX or PBS in the DSS-induced colitis model. C57BL/6 mice were treated with 4% DSS in the drinking water for 7 days and then received injection IP of WEPLEX (fresh or lyophilized/reconstituted) or PBS on day 2 and 4. The body weight was measured daily for each mouse and is depicted as the percentage of the initial value measured before the onset of the experiment (n = 5 in each group).
Discussion
Platelets and blood plasma derivatives such as PRP and PL have been extensively tested in regenerative medicine and stem cell-based tissue engineering.5,34 There are numerous studies, both in vitro and animal models, demonstrating the efficacy of these blood plasma derivatives.21,35–39 Unfortunately, the use of these products in clinical settings, mostly with autologous PRP or PL, has shown mixed clinical results in dermatology, dentistry, ophthalmology, orthopedics, and others.11,40,41 A major theoretical constraint of all the blood plasma derivatives is the limited availability of platelet-derived growth factors and cytokines administered at the injection site. 42 Furthermore, composition variation is correlated with age, sex, and patient conditions, 43 an issue addressed by generating PRP and PL from a pool of multiple donors. 44
In this study, we addressed these limitations of autogeneic/allogeneic PRP and PL in an entirely novel manner. A novel biological product was manufactured with our formulation of WEPLEX. The platelet content of PRP is low on a v/v basis (<0.5%), while our novel biological product concentrated by centrifugation increased v/v yield to >99%. In addition, concurrent multiple washes with saline solution during the creation of our product, remove immunogenic allogeneic plasma proteins such as albumin, immunoglobulins, fibrinogen, and complement and decrease the risk of immune reaction or serum sickness following administration. The concentrated platelet pellet is solubilized and lysed by the addition of a buffer-containing detergent. The detergent emulsifies the platelet membranes and also allows the removal of endotoxins and potential encapsulated viruses. Moreover, we showed the possibility to lyophilize WEPLEX and store the powder for later use, a process that is not feasible with albumin-rich and serum dilute PRP or PL. This novel formulation would permit off-the-shelf use of batch-produced platelet extract for human and veterinary medicine. The final product is a platelet extract 266-fold more concentrated in PDGF compared to all the platelet products currently available on the market, including PL. Lyophilization also allowed for attaining high concentration of platelet extract after reconstitution by using a smaller volume of diluent. The new formulation would allow injection of active substances in small volumes, not otherwise achievable in restricted spaces (e.g., ligaments, tendons, and joints).
No known endemic or endogenous zoonosis can be transmitted from horse to human making equine-sourced platelets more desirable than other mammal platelet sources where endemic or endogenous zoonosis risks exist (e.g., porcine, ovine, and bovine). However, there are reported acquired equine viral diseases in certain geographical areas, like Hendra virus and West Nile fever, which theoretically can be transmitted to humans. 45 The transmission of these viruses to humans is very limited in number and the possibility to easily screen a priori for the presence of these viruses in donor horses45,46 or to vaccinate donor horses for West Nile virus46,47 makes an equine-derived platelet product still a good choice.
After testing multiple clinical-grade detergents and considering lysis efficiency as the amount of PDGF over total proteins released compared to PRP, 2% v/v Triton X-114 in PBS was identified as optimal. Indeed, 2% Triton X-114 allowed for the highest protein release in the extract as well as the highest amount of PDGF content that served as a surrogate of platelet extraction efficiency. 48 The washing of platelets prelysis allowed no detectable IgG, fibrinogen, or albumin as residual plasma protein contamination.49,50 Moreover, Triton X-114 has a relatively low clouding point at 22°C and it can be efficiently removed from the final preparation by phase separation. Indeed, centrifugation at 15,000 g at 27°C allowed the separation of the detergent phase (lower phase) with the aqueous/protein phase (upper phase). The detergent phase allowed the removal of endotoxin from the final protein extract.51,52
Denaturation of proteins or loss of biological activity can occur by heating, freezing, or agitation in aqueous solutions. Furthermore, long-term storage of liquid solutions of proteins can limit the shelf life of a product. Lyophilization can protect against some of these effects and can extend the shelf life of protein preparations. 53 Under vacuum (very low pressure), the vapor-liquid equilibrium of the solvent is shifted toward the gas phase, while our protein extract remains primarily in the solid phase. Using vacuum, we can readily remove solvent with no effect on the quality attributes of WEPLEX, leaving a dry, solid sample. The dry powder was easily reconstituted in deionized water and no significant difference could be detected biochemically based on total protein or PDGF content.
PRP and related PL derivatives have been shown to be potent suppressors of T cell functionality, including TCR-mediated mitogenic response. 54 To determine the T cell immunosuppressive potential of WEPLEX, we analyzed its ability to suppress the replicative response of equine or human PHA-activated PBMCs. In vitro suppression assay showed that lyophilized/reconstituted WEPLEX was substantially equivalent to the one freshly generated. Indeed, with equine or human PHA-activated T cells, both lyophilized/reconstituted and freshly generated WEPLEX were able to suppress the proliferation of T cells in vitro. We further tested the immunosuppressive properties of WEPLEX on isolated peritoneal macrophages. Monocytes/macrophages are innate immune effectors, which can be polarized to an M2 immune suppressor state functionally characterized by their production of IL-10. 55 Depending on the microenvironment, macrophages can polarize to M1 (inflammatory) or M2 (anti-inflammatory) phenotypes. We established the conversion to an M2-like phenotype by detecting the IL-10 secreted in the media and by establishing the amount of IL-10+ cells by flow cytometry. We used peritoneal macrophages derived from Vert-X IL-10 reporter mice (REFERENCE). Macrophages derived from Vert-X mice that express IL-10 will co-express GFP and can be easily detected by flow cytometry. 56 WEPLEX was able to polarize peritoneal macrophages to a M2-immunosuppressive phenotype.
Finally, we demonstrated the protective properties of WEPLEX in an acute tissue injury model of DSS-induced colitis in mice. WEPLEX was able to protect the mice from any major weight loss induced by DSS and to have significantly improved survival compared to the mice treated with PBS only. These results indicate that WEPLEX has a protective effect in an acute tissue injury model and it can also be used in a species-mismatched, xenogeneic setting.
Our findings show that WEPLEX is likely to have a broad and significant impact on acute injury of tissue in both the veterinary and human medical field akin to what was expected from PRP.57–59 This new formulation of platelet extract could potentially be used not only in acute tissue injury syndromes and regenerative medicine but also in other medical fields where platelet-derived products have been already used with variable results such as dermatology, cardiology, and sports medicine.
Footnotes
Disclosure Statement
A.P., S.B., and J.G. are listed on a provisional patent application covering findings reported in this article.
Funding Information
The research has been supported by the University of Wisconsin Foundation, Department of Medicine.