
Abstract
The use of induced pluripotent stem cells (iPSCs) shows potential in bone regenerative strategies. In this study, we investigated whether implantation of chondrogenically differentiated iPSC-derived mesenchymal stem cells (iMSCs) can lead to successful bone regeneration in nude mice with bone defects. Two human iPSC clones (201B7 and 454E2) were used. After generating iMSCs, chondrogenic differentiation was achieved by three-dimensional pellet culture. Thereafter, a 2-mm defect was created in the radius of nude mice, and chondrogenically differentiated iMSC pellets were transplanted in the defect. Microcomputed tomography imaging was performed 8 weeks posttransplantation to assess bone regeneration. All (100%) radii in the 201B7 cell-derived pellet transplantation group and 7 of 10 (70%) radii in the 454E2 cell-derived pellet transplantation group showed bone union. In contrast, 2 of 11 radii (18%) in the control group showed bone union. Thus, the experimental groups showed significantly higher bone union rates than the control group (p < 0.05). Histological analysis 2 weeks postimplantation in the experimental groups revealed hypertrophic chondrocytes within grafted iMSC pellets and the formation of woven bone around them. This hypertrophic chondrocyte transitioning to newly formed bone suggests that the cartilaginous template can trigger endochondral bone ossification (ECO). Four weeks postimplantation, the cartilage template was reduced in size; newly formed woven bone was predominant in the defect site. New vessels were surrounded by a matrix of woven bone, and hypertrophic chondrocytes transitioning to newly formed bone indicated the progression of ECO. Eight weeks postimplantation, the pellets were completely resorbed and replaced by bone; complete bone union was observed. Dense mature bone developed with evidence of lamellar-like bone formation. Collectively, our results suggest that using iMSC-based cartilage grafts recapitulating the morphogenetic process of ECO in the context of embryonic skeletogenesis is a promising strategy for repairing large bone defects.
Impact statement
We investigated whether implantation of chondrogenically differentiated iPSC-derived mesenchymal stem cells (iMSCs) could lead to the successful regeneration of bone defects in vivo. We implanted two different clones of human induced pluripotent stem cells into a radial bone defect model. Eleven of 11 (100%) and 7 of 10 (70%) radii in the 201B7 and 454E2 cell-derived pellet transplantation groups, respectively, showed bone union, which were significantly higher than those in the control group [only 2 of 11 radii (18%)]. Overall, our results support the use of iMSC-based cartilage grafts recapitulating the morphogenetic process of endochondral bone ossification for repairing large bone defects.
Introduction
Treatment of bone defects caused by severe trauma, nonunion, osteomyelitis, or bone tumors remains challenging in orthopedic surgery. 1 As a result, the quality of life and daily living activities of patients are disrupted. Bone grafting is the most commonly used surgical procedure for treating bone defects; autogenous bone grafting represents the clinical “gold standard.” However, the amount of available autologous bone is limited, and the harvesting procedures are associated with donor-site morbidity and various complications, such as severe pain, hemorrhage, infections, fractures, and nerve damage. In fact, in over half of reported cases, autogenous bone grafts showed unsatisfactory results. 2 To overcome these limitations, bone tissue engineering using stem cells and scaffolds has emerged as a promising strategy for bone repair. 3
Cell-based bone tissue engineering requires the large-scale production of homogeneous populations of lineage-restricted progenitor cells that can easily differentiate into specific tissues. Mesenchymal stem cells (MSCs) are considered as a promising source of progenitors for tissue engineering strategies. 4 MSCs are multipotent progenitors found in many connective tissues, including the bone marrow and fat.5–7 However, the use of MSCs is limited by their heterogeneity, availability, invasive and laborious harvest procedures, finite proliferation potential, and loss of differentiation capacity during expansion.8–10 Recently, a major breakthrough in the generation of patient-specific pluripotent stem cells was reported by Yamanaka et al.; embryonic stem cell-like pluripotent stem cells, designated as induced pluripotent stem cells (iPSCs), were generated from adult somatic cells using reprogramming techniques.11,12 iPSCs possess unlimited self-renewal capacity and the ability to differentiate into all somatic cell types. Importantly, iPSCs can be derived from the patient's own somatic cells, thereby avoiding immune rejection after transplantation as well as ethical concerns. Notably, several studies successfully achieved derivation of MSCs from iPSCs.13,14 Therefore, iPSC-derived MSCs, named as “iMSCs,” represent a highly promising and attractive source of large populations of progenitors for use in bone tissue engineering.15,16 However, the in vivo regenerative potential of iPSC-derived mesenchymal stem cell (iMSCs) in the context of long-bone repair has not been clearly demonstrated. 16
The bone repair process closely resembles normal development of the skeleton during embryogenesis, including intramembranous and endochondral ossification (IMO and endochondral bone ossification [ECO], respectively).17–19 In IMO, MSCs differentiate directly into osteoblasts that form the bone. In ECO, MSCs differentiate into chondrocytes, which form a cartilage template; next, chondrocytes within this template undergo hypertrophy, secreting cytokines such as vascular endothelial growth factors (VEGF), enzymes such as alkaline phosphatase, and proteases (e.g., matrix metalloproteinases). These factors stimulate vascularization and mineralization of the cartilage template, which is subsequently resorbed by attracted osteoclasts and replaced by bone. Importantly, ECO is the dominant mode of ossification in the natural healing of bone fractures. Conventional bone tissue engineering approaches have typically recapitulated the process of IMO through direct in vitro osteogenic priming of MSCs within scaffolds before implantation. However, this approach is associated with insufficient vascularization of the graft following implantation, thereby preventing the necessary delivery of oxygen and nutrients. 20 As a result, such strategies have not been widely used for the clinical treatment of bone defects. Recently, studies have focused on strategies that recapitulate the process of ECO. In fact, several studies successfully demonstrated the capacity of in vitro tissue-engineered cartilage to undergo ECO and generate bone tissue in vivo.17–19 Previous studies showed that chondrogenically primed constructs seeded with chondrocytes isolated from chick embryos, 21 human embryonic stem cells, 10 and human MSCs10,22 act as osteoinductive templates and transform into bone tissues following implantation in rodent animal models. However, to date, bone repair using iPSCs through the process of ECO has not been attempted.
In this study, we investigated whether implantation of chondrogenically differentiated human iMSCs could lead to the regeneration of radial bone defects in nude mice through ECO.
Methods
For more detailed procedures, please see the Supplementary Data.
Derivation of human iMSCs
The culture of human iPSCs was approved by the Institutional Review Board of Kobe University, Graduate School of Medicine (#26–41). Two clones of human iPSCs, 201B711 and 454E2, 23 were used in this study. MSCs were generated from iPSCs using a modified “direct-plating method” omitting the embryoid body (EB) formation step.24–26 We referred to the cells obtained through this direct-plating method as iMSCs. The morphology of the different cell colonies is shown in Figure 1A–C.
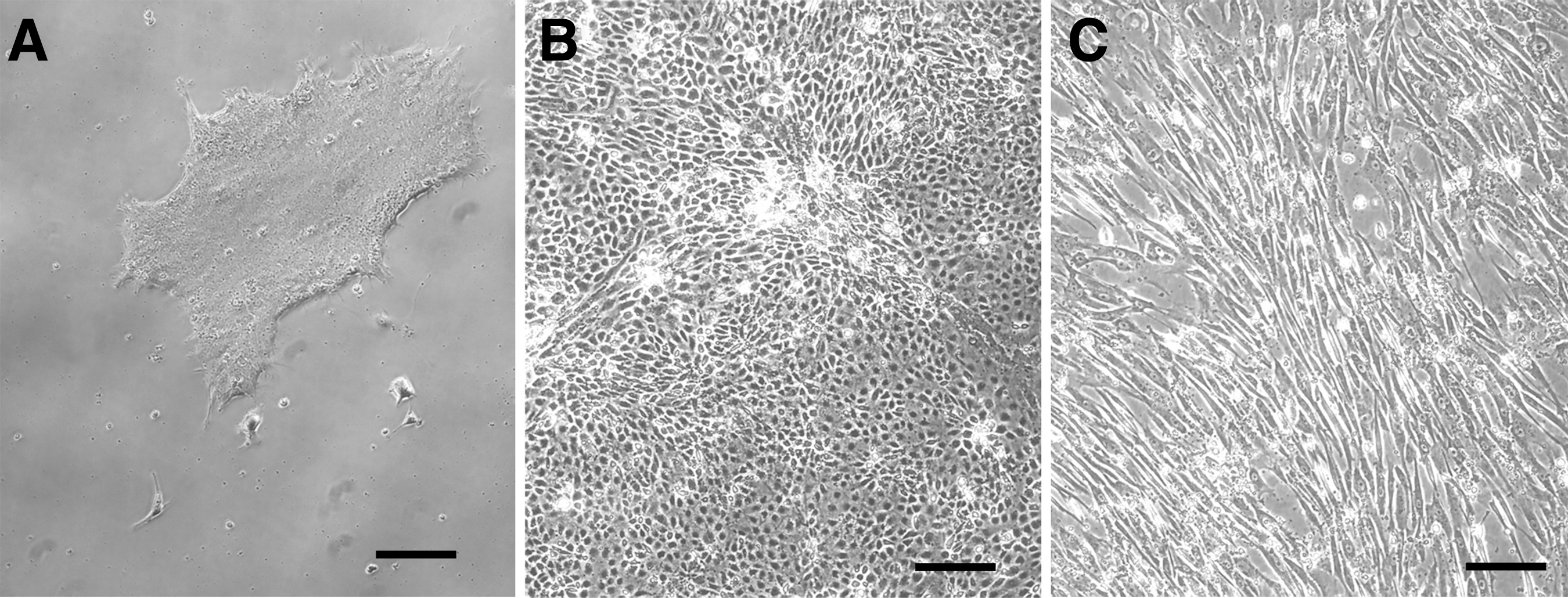
Morphology of
Surface antigens and multidifferentiation potential of iMSCs
The surface antigen profiles of iMSCs (n = 3 per iMSCs derived from 201B7 to 454E2) at passage 5 were characterized by flow cytometry. The multipotency of iMSCs was evaluated histologically. Differentiation (in vitro) into osteogenic, adipogenic, and chondrogenic lineages was investigated as previously reported.9,10
Immunohistochemistry of chondrogenically differentiated iMSC pellets
Chondrogenically differentiated iMSC pellet sections were stained with anti-human-type collagen II (Col II), type X collagen (Col X), VEGF, and stromal cell-derived factor 1 (SDF-1) antibodies.
In vivo experiments
All animal procedures were performed under the approval and guidance of the Animal Care and Use Committee of our institution. BALB/cAJcl-nu/nu mice (CLEA, Tokyo, Japan) were used. A 2-mm nonunion defect was created in the radius of each mouse, as previously described (Fig. 2A).27,28 Thereafter, chondrogenically differentiated iMSC pellets were transplanted into the defects; animals in the control group were treated with saline (Fig. 2B).

Radial defect model for orthotopic bone regeneration.
Reverse transcription–polymerase chain reaction
At week 2, the newly generated tissues around implanted pellets in the defect site were harvested. The expression of human and mouse osteocalcin (OC), type I collagen (Col I), Col II, Col X, VEGF, and SDF-1 was analyzed by reverse transcription–polymerase chain reaction (RT-PCR). Human and mouse glyceraldehyde 3-phosphate dehydrogenase was used as an internal control for normalization (Supplementary Tables S1 and S2).
Tissue staining
Histological, immunofluorescence, and immunohistochemistry analyses were carried out 2, 4, and 8 weeks postimplantation using tissue sections from the radii of the different groups of animals.
Analysis of bone regeneration through microcomputed tomography
Bone regeneration was assessed by microcomputed tomography (μ-CT) imaging at 8 weeks postimplantation by three examiners blinded to the experimental details. Bone union was defined as the absence of a bone gap or presence of a bridging callus at three of four cortices in three-dimensional (3D) μ-CT images. 29 Bone formation parameters, including the total callus volume (TV), bone surface (BS), tissue mineral density (TMD), and volumetric bone mineral density (vBMD), were also determined.
Statistical analysis
Fisher's exact test was used to compare the bone union rates between the 201B7 or 454E2 group and control group. Kruskal–Wallis test followed by Bonferroni-corrected Mann–Whitney U post hoc analysis was used to compare the capillary density and μ-CT measurements among the three groups. Statistical significance was set at p < 0.05.
Results
In vitro study
Characterization of human iMSCs
Flow cytometry analysis revealed that iMSCs were consistently positive for the MSC-related markers, CD29, CD44, CD105, and CD166, but negative for the hematopoietic stem cell markers, CD31, CD34, CD45, and CD133 (Table 1). Notably, iMSCs showed low expression of the pluripotent markers TRA-1-60 and TRA-1-81, suggesting the loss of pluripotency.
Flow Cytometry Analysis of iMSCs
Positive expression rates (%) are displayed as mean ± standard deviation.
At 21 days after osteogenic induction of iMSCs, Alizarin Red S staining revealed a mineralized matrix rich in calcium deposition (Supplementary Fig. S1A). Additionally, Oil Red O staining showed fat vacuole accumulation in iMSCs at 21 days after adipogenic induction (Supplementary Fig. S1B), whereas, at 21 days after the chondrogenic induction of 3D cultures, the iMSC pellet was spherical (1–2 mm in diameter) and glistening transparent in appearance with a uniform distribution of proteoglycan deposition as revealed by Safranin O staining (Fig. 3A). These results suggest that iMSCs can differentiate into osteogenic, adipogenic, and chondrogenic cells with multidifferentiation potential.
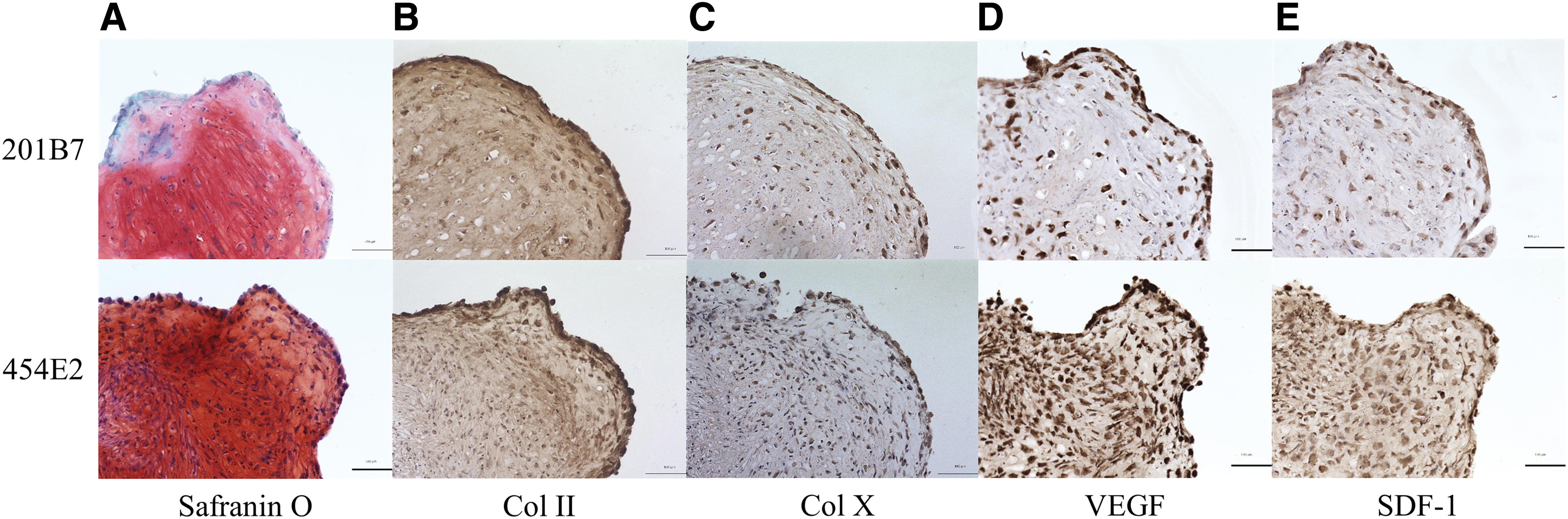
Histological evaluation of chondrogenically differentiated iMSC pellets in vitro at day 21; Safranin O staining
Immunohistochemistry of chondrogenically differentiated iMSCs
Further evidence of chondrogenesis in the context of iMSCs was provided by Col II immunohistochemistry staining (Fig. 3B). The patterns of Col II deposition corresponded to those observed by Safranin O staining (Fig. 3A). Additionally, Col X deposition within the pellets was observed as weak staining, indicating a certain degree of hypertrophy in the pellets (Fig. 3C). Notably, positive staining for VEGF and SDF-1 was observed in and around the chondrocytes (Fig. 3D, E).
In vivo study
RT-PCR analysis of bone defect sites
In the grafted groups, RT-PCR analysis revealed the positive expression of human Col II, Col X, VEGF, and SDF-1, but not of human OC and Col I in the grafted bone defect sites (Fig. 4). In contrast, in the control group, no expression of human genes was detected. In all groups, the positive expression of mouse counterpart genes was detected (Supplementary Fig. S2).
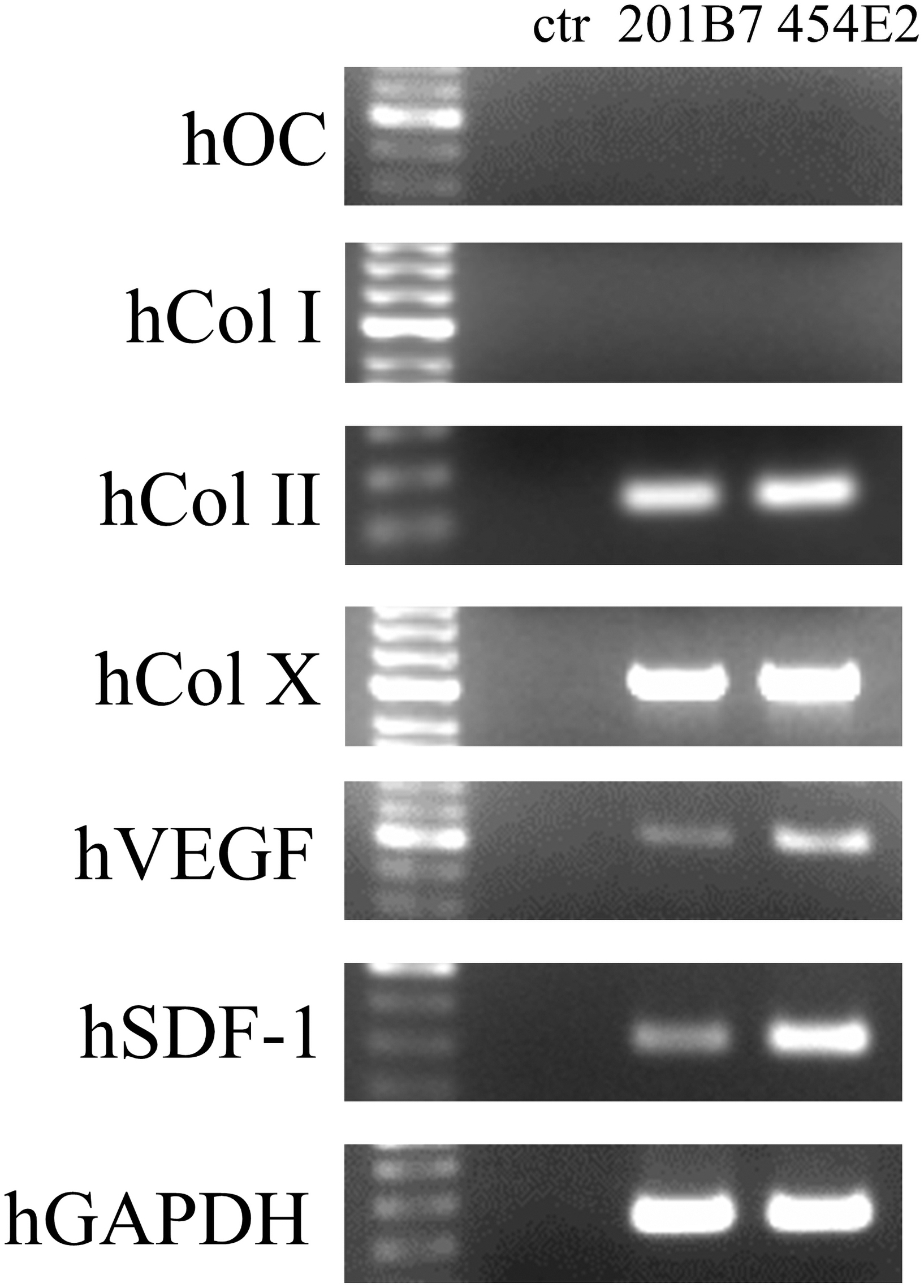
RT-PCR analysis of the expression of human OC, Col I, Col II, Col X, VEGF, and SDF-1 at the bone defect sites 2 weeks postimplantation. Col I, type I collagen; OC, osteocalcin; RT-PCR, reverse transcription–polymerase chain reaction.
Capillary density in bone defect sites
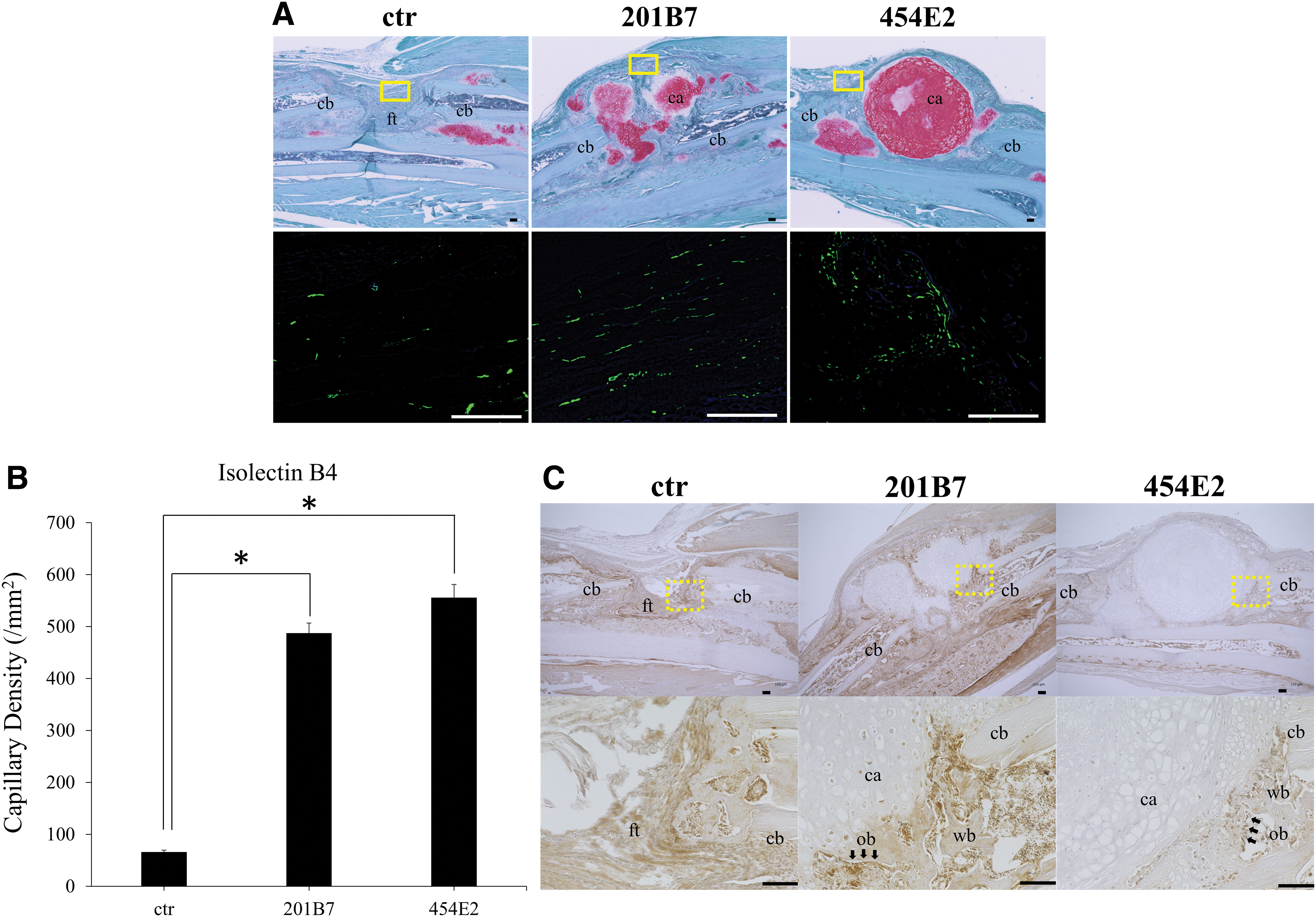
Visualization of the capillaries through immunohistochemistry staining of isolectin B4 in tissue samples collected at 2 weeks postimplantation revealed greater angiogenesis surrounding the implanted pellets in the grafted groups (Fig. 5A), with the capillary density significantly higher than that in the control group (p < 0.05; Fig. 5B). There was no significant difference between the 201B7 and 454E2 groups.
Immunolocalization of mouse OC in bone defect sites
At 2 weeks postimplantation, immunoreactivity against mouse OC (mOC) was detected around the implanted pellets (Fig. 5C). Moreover, in the control group, immunoreactivity of mOC was detected at cortical bones along the edge of bone defect sites.
Radiographic assessment of bone regeneration
Microcomputed tomography analysis performed at 8 weeks post-implantation showed striking bone formation in the defect sites in the grafted versus control groups. Regarding the rate of bone union, 11/11 (100%) radii in the 201B7 group and 7/10 (70%) radii in the 454E2 group showed bone union. In contrast, only 2/11 radii (18%) showed bone union in the control group, a significantly lower rate than in the experimental groups (p < 0.05; Fig. 6A). Quantification of bone formation revealed that the TV and BS in the defect sites in the 201B7 and 454E2 groups were higher than those in the control group (p < 0.05), whereas no significant differences were observed with respect to the TMD and vBMD among the three groups (Fig. 6B).
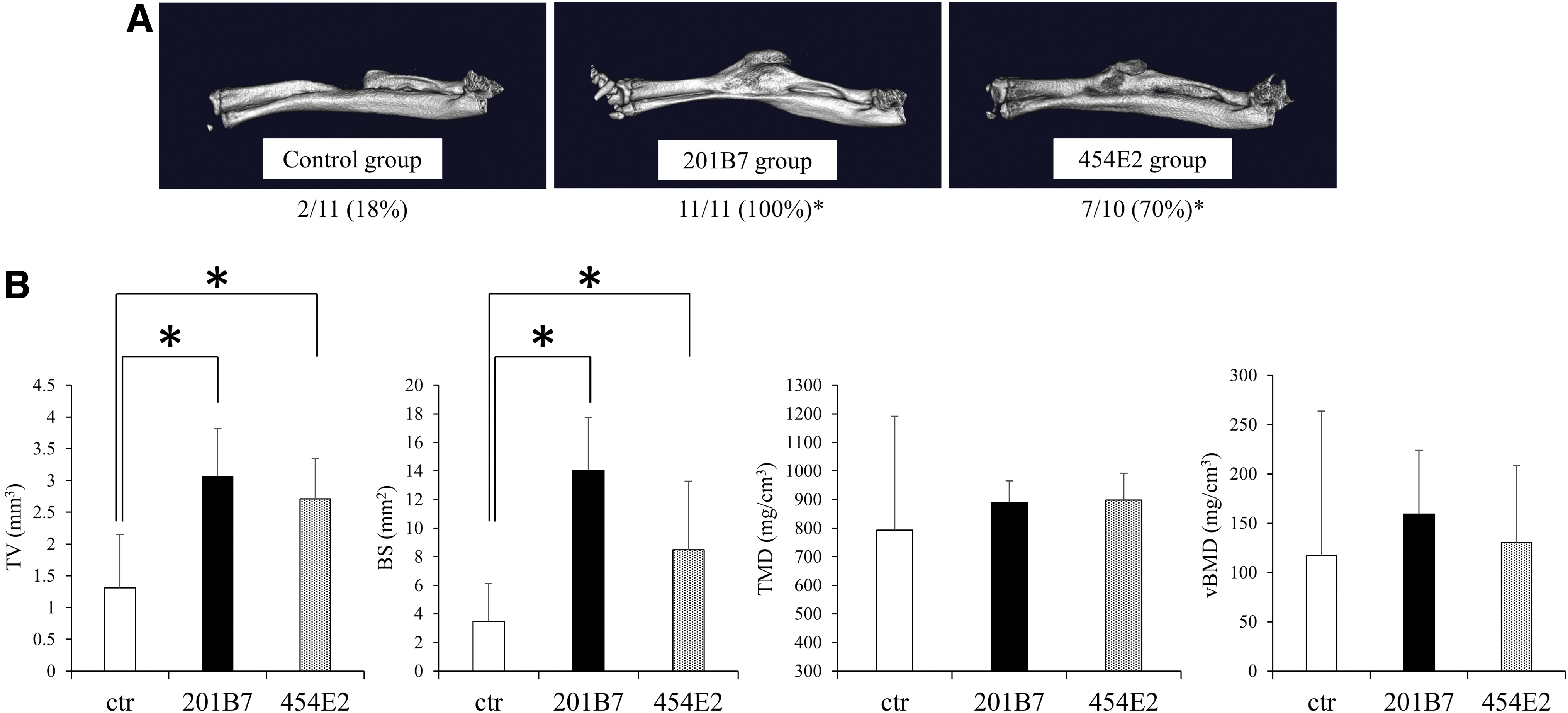
Histological assessment of bone regeneration
At 2 weeks postimplantation, the grafted pellets were still visible in the defect sites, indicating their survival (Fig. 7A); they formed a cartilage template that bridged the defect. Within the pellets, chondrocytes differentiated into hypertrophic chondrocytes, as indicated by larger lacunae with smaller uniform nuclei. At the outer rim of the pellets, the extracellular matrix was not stained with Safranin O, suggesting the beginning of chondrocyte maturation. Additionally, around the pellets, newly formed woven bone was observed. The presence of remnants of hypertrophic cartilage next to the newly formed bone suggested that bone formation occurred through an ECO process. In contrast, in the control group, the defect site was filled with fibrous tissue.
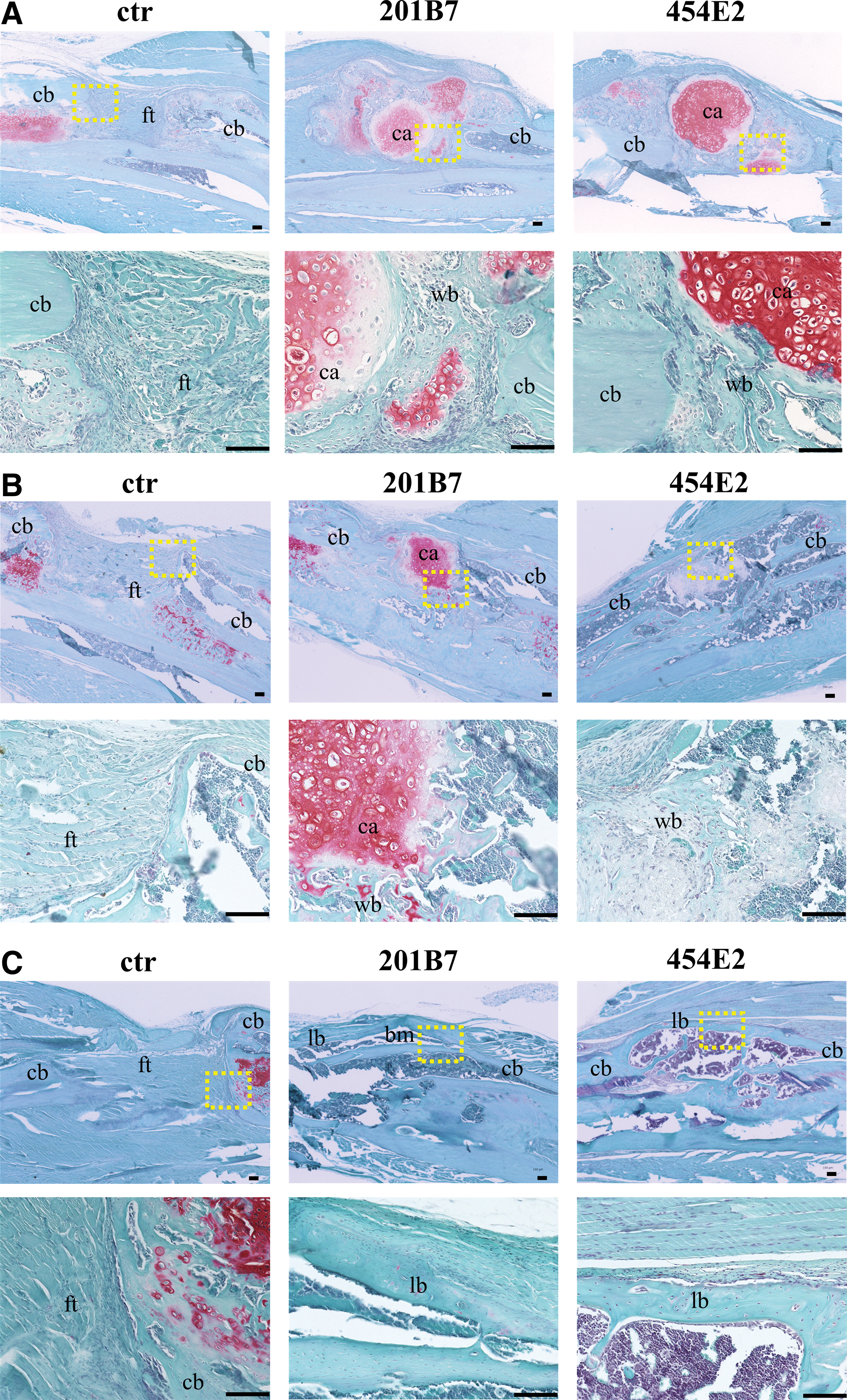
Histological evaluation. Safranin O/fast Green staining at the defect sites in the control, 201B7, and 454E2 groups
Furthermore, at 4 weeks postimplantation, the cartilage template was reduced in size in the experimental groups; newly formed woven bone was predominant in the defect site. New vessels were surrounded by a matrix of woven bone and the hypertrophic chondrocytes transitioning into newly formed bone indicated the progression of ECO. In contrast, in the control group, fibrous tissue remained in the defect site (Fig. 7B).
At 8 weeks postimplantation, the pellets were completely resorbed and replaced by bone in the experimental groups; complete bone union was observed overall, with evidence of lamellar-like bone formation. We even observed bone marrow-resembling tissues in some bone lacunae. In contrast, in the control group, the remaining gap was filled with fibrous tissue (Fig. 7C).
Overall, similar histological findings were observed in both the 201B7 and 454E2 groups. No evidence of teratoma formation was found in any mice implanted with iMSC pellets (Fig. 7A–C). The cartilage area at week 2 was significantly higher in the experimental groups than in the control group (p < 0.05; Supplementary Fig. S3).
Donor-derived bone formation: immunohistochemistry analysis
To determine the contribution of the transplanted human chondrogenically differentiated iMSCs to new bone formation, immunohistochemistry using human-specific anti-OC and antinuclear monoclonal antibodies that do not crossreact with murine antigens was performed (positive control images are shown in (Supplementary Fig. S4). At 2 weeks postimplantation, immunoreactivity against human nuclei was not detected outside of the pellets in the experimental groups (Fig. 8A). However, at 4 weeks postimplantation, immunoreactivity against human nuclei was detected in the newly formed bone around the pellets (Fig. 8B). This was also evident at 8 weeks; immunoreactivity was observed in osteocytes within the newly formed bone (Fig. 8C).
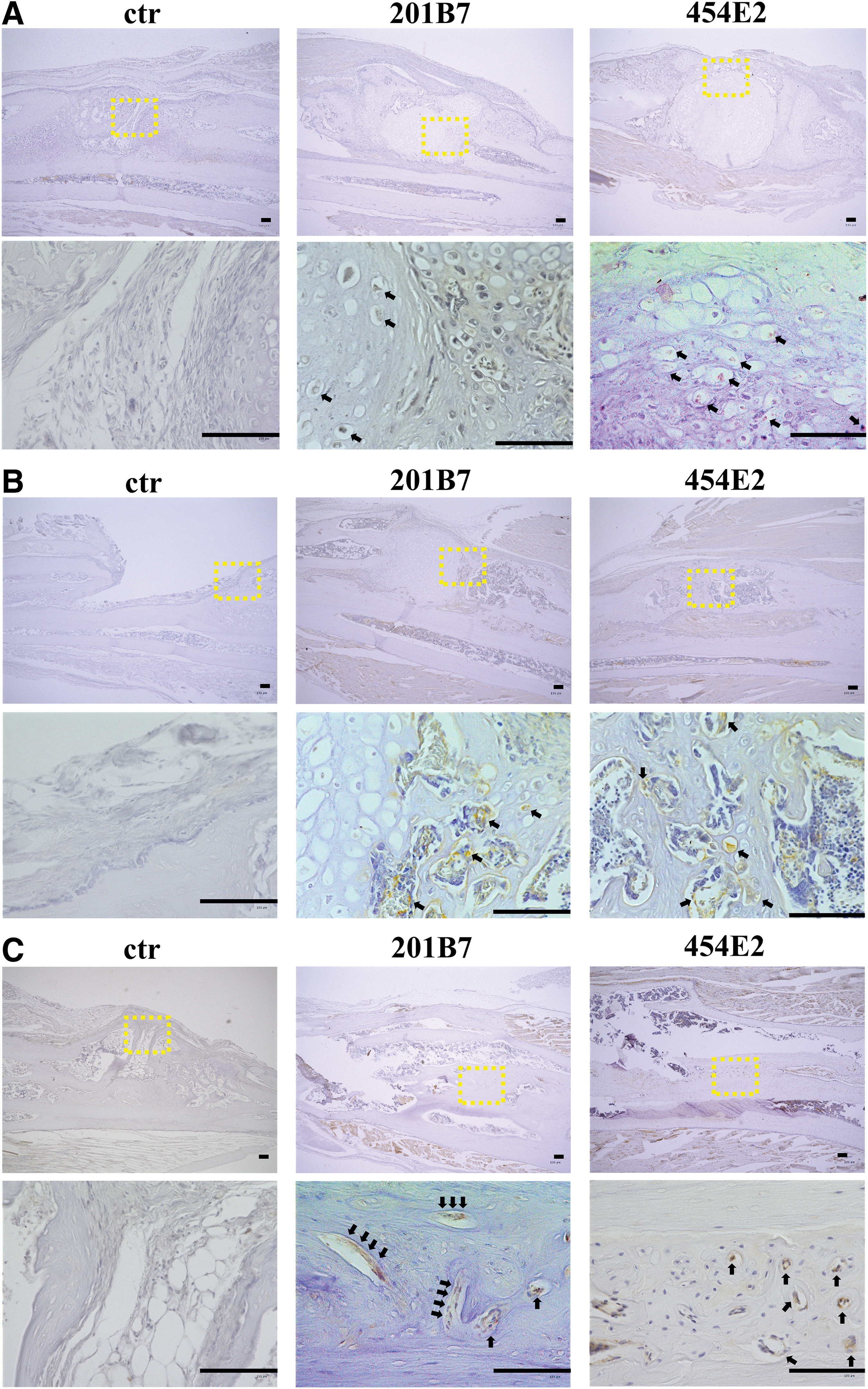
Immunohistochemistry of human nuclei at the bone defect sites in the control, 201B7, and 454E2 groups
In contrast, immunoreactivity against human OC was not detected in the experimental groups at 2 weeks postimplantation (Fig. 9A). However, at 4 weeks, osteoblast lining cells in the newly formed bone around the pellets were positive for human OC (Fig. 9B), whereas at 8 weeks, immunoreactivity against human OC was observed in osteocytes within the newly formed bone (Fig. 9C).
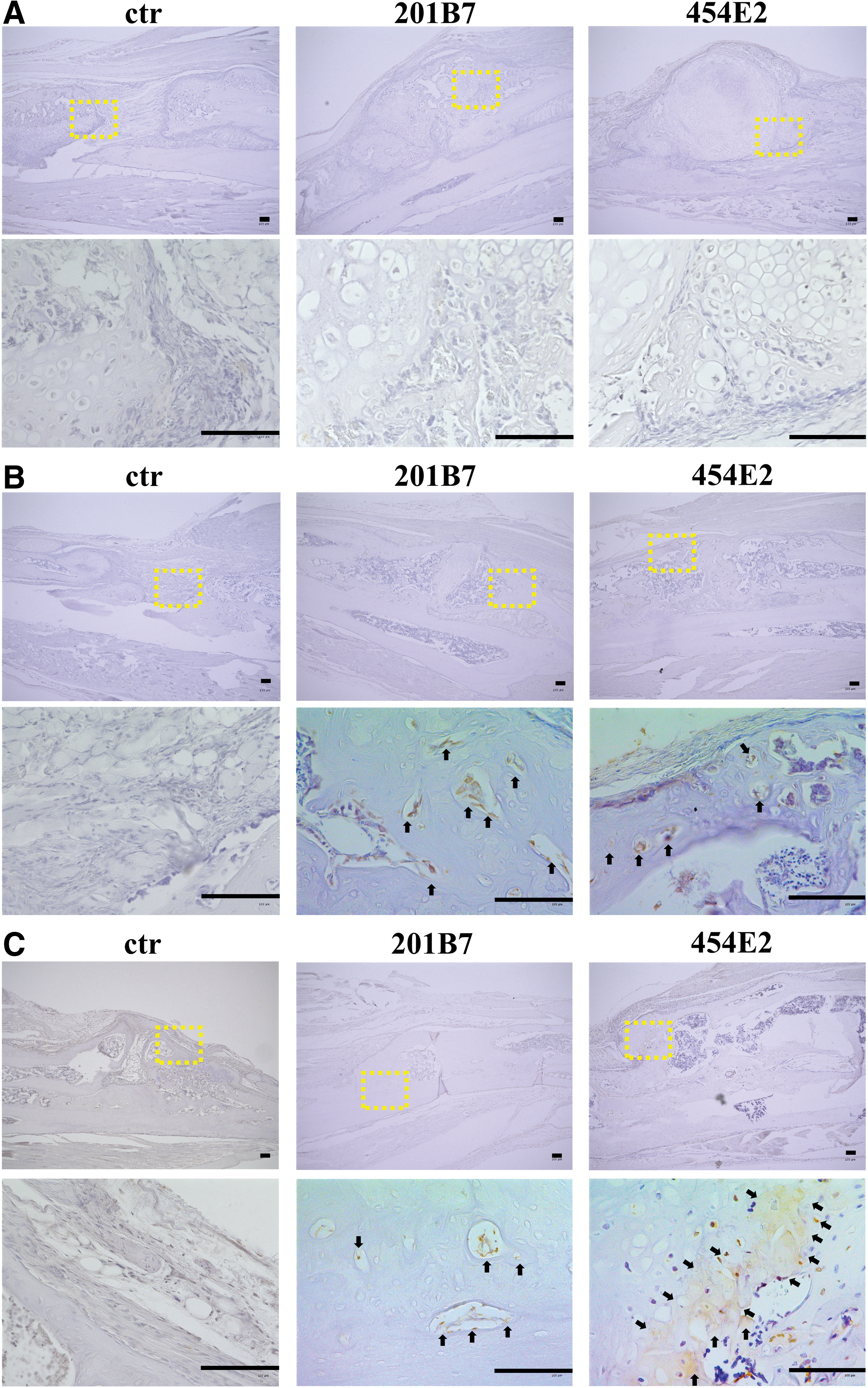
Immunohistochemistry of human OC at the bone defect sites in the control, 201B7, and 454E2 groups
Discussion
Recently, several in vitro methodologies have been developed to derive functional, multipotent MSCs from human iPSCs and embryonic stem cells, including an intermediate step using EB formation,14,28 coculture, 30 and cell sorting.30,31 In this study, using a simple direct-plating method without the need for EB formation or cell selection,24–26 we successfully generated MSCs from human iPSCs; these cells showed typical MSC characteristics.14,28 iMSCs exhibited a uniform fibroblastic morphology and expressed high levels of typical MSC cell surface markers (CD29, CD44, CD105, and CD166), but did not express hematopoietic lineage markers (CD31, CD34, CD45, and CD133). iMSCs also possessed multilineage potential, differentiating toward the osteogenic, adipogenic, and chondrogenic lineages when cultured in the presence of lineage-specific differentiation factors.8,9 These results contrast those of Xu et al., 32 who observed marked reduction of the in vitro chondrogenic differentiation potential of iMSCs (compared with that of bone marrow-derived MSCs). This discrepancy is likely related to the different methods used for iMSC derivation. Xu et al. 32 used an EB outgrowth method, as EBs are known to be composed of cells with different degrees of multipotency that may limit the differentiation potential. 33 Additionally, the iMSCs obtained in the present study did not express the pluripotency-related markers, TRA-1-60 and TRA-1-81; this fact, coupled with the observation that no teratomas were formed for up to 8 weeks postimplantation in vivo, suggests the loss of pluripotency and a reduced tumorigenic risk. As iPSCs can be expanded indefinitely, the direct-plating method has considerable potential for in vitro generation of large numbers of homogeneous MSCs for bone regeneration.
In the last decade, the “developmental engineering” paradigm: engineering “processes” should recapitulate embryonic events rather than “tissues,” was proposed in regenerative medicine. 34 Importantly, ECO during bone repair recapitulates the key process of ECO during development. Therefore, the scientific community suggested performing bone regeneration through the ECO pathway.17–19 ECO involves a complex series of highly regulated steps, in which MSCs condense, differentiate into chondrocytes, mature to the hypertrophic state, and stimulate vascular invasion and mineralization before transforming into bone. 19 A potential advantage of using cartilage grafts through the ECO (vs. IMO) route to promote bone healing is that cartilage cells are physiologically adapted to survive under avascular, hypoxic conditions. Thus, ECO-based bone formation can circumvent the issues associated with the supply of oxygen and nutrients to large, engineered constructs. Chondrogenically primed MSCs have an inherent tendency to undergo hypertrophy. 35 Indeed, our results revealed that iMSC pellets after differentiation in vitro developed a cartilage matrix expressing Col X, a hypertrophic marker (Fig. 3C). RT-PCR analysis in vivo confirmed the expression of human Col X (Fig. 4), and histological analysis revealed hypertrophic chondrocytes within grafted iMSC pellets and the formation of woven bone around them in the experimental groups. In addition, the cartilage area in the defect site at week 2 was significantly higher in the experimental groups than in the control group (Supplementary Fig. S3). These results suggest that the cartilaginous template can trigger the process of ECO following exposure to the in vivo environment. Collectively, from a bone developmental engineering perspective, iMSC-based cartilage grafts recapitulating the morphogenetic process of ECO in the context of embryonic skeletogenesis is a promising strategy for repairing large bone defects.
VEGF plays a key role in ECO, affecting angiogenesis, extracellular matrix remodeling, and ossification. 36 In the later stage of ECO during bone development and repair, chondrocytes in the cartilaginous template stop proliferating, undergo hypertrophy, and synthesize Col X. Hypertrophic chondrocytes also express and release VEGF, inducing the invasion of blood vessels into the cartilage template and facilitating bone formation. 37 In the current study, immunoreactivity against human VEGF was detected within iMSC pellets cultured for 3 weeks under chondrogenic conditions. Moreover, the expression of human VEGF was observed in vivo at 2 weeks postimplantation. We also found that the capillary density around the bone defect sites in the experimental groups was significantly higher than that in the control group (at week 2), suggesting that transplanted chondrogenically differentiated iMSC pellets enhanced angiogenesis and vasculogenesis. Overall, these data strongly support the initiation of ECO.
In fact, previous studies demonstrated that chondrogenically differentiated MSCs can form bone through ECO after ectopic subcutaneous implantation in rodents.20–22 However, ectopic implantation does not fully reflect the complex environment of bone defects (e.g., nonunion); thus, this model is not ideal for assessing constructs for bone repair. Therefore, to fully determine the bone regenerative capacity in an orthotopic environment, we used a weight-bearing bone (radius) defect model.27,28 This model has clinical relevance for treating human bone defects with large segment grafting. Several recent studies revealed endochondral bone regeneration after the transplantation of MSCs predifferentiated into chondrogenic cells in orthotopic models.38–42 For instance, Harada et al. 40 implanted poly(D,L-lactic-co-glycolic acid) scaffolds seeded with chondrogenically primed rat bone marrow-derived MSCs into critical-sized (5 mm) and massive full-thickness (15 mm) rat femoral defects; newly formed bone and full bone union were observed at 8 and 16 weeks postimplantation, respectively. Similarly, rapid healing was reported when chondrogenically primed human bone marrow-derived MSC pellets were implanted into a 6-mm rat femoral defect. 38 Additionally, Sheyn et al. 28 implanted undifferentiated BMP-6-overexpressing human iMSCs into 1.5-mm radial defects in mice and demonstrated complete bone regeneration by week 8. Jungbluth et al. 43 implanted undifferentiated human iSCs into 11-mm diameter and 25-mm depth “partial-thickness” cylindrical defects in the proximal tibia of mini pig (a model that does not mimic nonunion or refractory fracture of the long bones in patients), with only 46% of bone defect consolidation by week 6. Importantly, in the current study, we demonstrated that human iMSCs can regenerate bones in vivo through ECO without gene transfection in a preclinical orthotopic long bone defect model. We showed that in nude mice, implantation of chondrogenically differentiated iMSC pellets into 2-mm radial bone defects results in successful bone regeneration with a significantly higher bone union rate, TV, and BS in the defect sites in the grafted groups compared with those in the control group (Fig. 6).
It is unclear whether the implanted iMSCs contribute directly to bone regeneration in an autocrine manner or have an indirect effect, such as paracrine signaling-induced regeneration through the recruitment and activation of osteoprogenitor cells from host animals. In this study, paracrine-induced recruitment of host osteoprogenitor cells was suggested by the observation of mouse OC-positive cells around the implanted pellets (Fig. 5C). In agreement with this, previous studies showed that the paracrine effects of growth factors and cytokines secreted by transplanted MSC pellets can promote tissue repair and regeneration. 44 One such candidate is SDF-1, which is among the most important molecules for ECO. SDF-1 plays a pivotal role in fracture healing, promoting the migration and differentiation of progenitor cells in fracture sites, and contributing to endochondral bone repair.45,46 For instance, Kitaori et al. 45 reported that SDF-1 promotes ECO by recruiting MSCs to the injury site. Additionally, in experimental models of fracture healing, SDF-1 was expressed in prehypertrophic and hypertrophic chondrocytes within the fracture callus.46,47 Murata et al. 47 also showed that SDF-1 was expressed at the growth plate and regulated chondrocyte differentiation during endochondral bone development, indicating that SDF-1 is crucial for hypertrophic conversion and the subsequent calcification of chondrocytes. In the current study, SDF-1 immunoreactivity was detected within chondrogenically differentiated iMSC pellets in vitro (Fig. 3E). In addition, at 2 weeks after in vivo implantation, the transplanted iMSCs expressed human SDF-1 (Fig. 4). Overall, these results indicate that bone regeneration is at least partially due to the paracrine effects of implanted iMSC pellets expressing SDF-1 and consequent recruitment of host osteoprogenitor cells.
At 4 and 8 weeks postimplantation, positive immunoreactivity against human nuclei and OC was detected in the newly formed bone, indicating that the implanted iMSCs survived and directly contributed to bone regeneration (Fig. 8B–C and Fig. 9B–C). These results suggest that one of the mechanisms underlying bone regeneration is direct differentiation of iMSCs into osteoblastic cells. Such cells may have developed from the remaining iMSCs within pellets that did not undergo differentiation during chondrogenic preinduction. 48 Another possibility is that not all chondrocytes became hypertrophic and underwent apoptosis, but rather that some cells retained the capacity to transdifferentiate into osteoprogenitor cells. Recent studies showed that during the end stages of ECO, hypertrophic chondrocytes can transdifferentiate into osteoblasts and osteocytes.40,49 In contrast, at week 2, neither immunoreactivity against human OC (Fig. 8A) nor human OC expression (Fig. 4) was detected in the defect sites. Thus, at least at earlier time points, donor iMSCs are likely involved only in indirect bone formation through paracrine effects. Ultimately, the mode of action of implanted cells remains unclear. iMSC tracing experiments are required to investigate the autocrine effects of iMSCs. In the current study, we revealed the potential of implanted chondrogenically differentiated iMSCs for repairing bone defects through ECO in an autocrine/paracrine manner.
One potential limitation of our study is that although no teratoma formed during the 8-week period, iMSCs may still be tumorigenic in vivo. Further investigation (long-term observation) is necessary to support the safety of chondrogenically differentiated iMSCs in vivo. In addition, although we demonstrated successful bone regeneration using iMSCs in a murine bone defect model, further experiments in large animals are necessary before translation into human trials. Additionally, we used scaffold-free pellet cultures to form cartilage grafts from iPSCs. iMSC pellets are inherently limited in size to ∼1–2 mm in diameter and may not be sufficient to graft large bone defects in humans. Incorporation of iMSCs into appropriate scaffolds that can be scaled-up is required to repair larger, clinically relevant bone defects. In contrast, there was a discrepancy between the results of radiographic (μ-CT) and histological assessments at week 8. Although histology in the control group showed that the defect site was filled with fibrous tissue, there were no significant differences in the TMD and vBMD in μ-CT analysis among the three groups. In some control group samples, the radius at the resection stump was fused with the ulna. In such cases, the callus tissues were included in the region of interest, which may explain this discrepancy.
Conclusions
Overall, our data show that grafting chondrogenically differentiated human iMSCs into bone defects (in the radius) in nude mice leads to successful bone regeneration through the process of ECO. Therefore, endochondral bone tissue engineering using human iPSCs should be considered as a strategy to repair long bone defects.
Footnotes
Authors' Contributions
M.A.: Study conception and design, acquisition of data, analysis and interpretation of data, and drafting and revising the article. S.Y.L.: Study conception and design, analysis and interpretation of data, and drafting and revising the article. T.F.: Analysis and interpretation of data, and drafting and revising the article. K.O.: Analysis and interpretation of data, and drafting and revising the article. S.T.: Acquisition of data, and analysis and interpretation of data. T.M.: Analysis and interpretation of data, and drafting and revising the article. S.H.: Analysis and interpretation of data, and drafting and revising the article. T.M.: Analysis and interpretation of data, and drafting and revising the article. R.K.: Analysis and interpretation of data, and drafting and revising the article. T.N.: Study conception and design, analysis and interpretation of data, and drafting and revising the article.
Acknowledgments
The authors would like to acknowledge Ms. M. Yasuda, Ms. K. Tanaka, and Ms. M. Nagata (Department of Orthopedic Surgery, Kobe University Graduate School of Medicine) for their excellent technical assistance, and Prof. Takashi Aoi (Department of iPS Cell Applications, Kobe University Graduate School of Medicine, Kobe, Japan) for his generous gift of SNL cells.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a Grant-in-Aid for Scientific Research (C) from Japan Society for the Promotion of Science (17K10967) and grants from ZENKYOREN (National Mutual Insurance Federation of Agricultural Cooperatives), the Nakatomi Foundation, and Hyogo Science and Technology Association (all to SYL).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.