
Abstract
Designing scaffolds, with optimized microstructure and function for promoting the release of neuro-related factors, is significant in peripheral nerve regeneration. Herein, a series of hydroxypropyl chitosan/soy protein isolate composite sponges (HCSS) were fabricated by a freeze-drying technique. The physicochemical properties of the resultant HCSS were examined by a Fourier infrared spectrometer, X-ray diffractometer, scanning electron microscope, water absorption assay, water retention assay, and compressive strength assay. The results indicated that HCSS exhibited an interconnected porous microstructure and a high water retention ratio with the increase in soy protein isolate (SPI) content. The biological characterization found that the HCSS-50 containing 50% SPI content profoundly promoted the proliferation of RSC96 cells and the secretion of neuro-related factors without excessive reactive oxygen species production. In addition, HCSS-50 could significantly promote the expression of neuro-related factors; for example, the expression of TGF-β was three times higher than that of the control group. Finally, an optimized HCSS-based conduit was fabricated from HCSS-50 to repair sciatic nerve injury in rats with the combination of bone marrow mesenchymal stem cells (BMSCs) or BMSC-derived Schwann cells (SCs). The results suggested that the constructed HCSS-based conduit accompanying BMSC-derived SCs could effectively promote axonal regeneration and upregulate the expression of neuro-related factors such as Krox20, Zeb2, and GAP43. Collectively, a newly engineered nerve conduit system was developed by incorporating HCSS-50 and BMSC-derived SCs, which could be an alternative candidate for peripheral nerve regeneration.
Impact statement
Peripheral nerve repair is of paramount significance in the clinical. This work describes a hydroxypropyl chitosan/soy protein isolate conduit, which could effectively promote axonal regeneration and upregulate the expression of neuro-related factors. Thus, we provide a potential candidate for peripheral nerve regeneration.
Introduction
Peripheral nerve injuries (PNIs) may be caused by accidental tissue damage and surgical interventions, affecting millions of individuals every year.1,2 These injuries reduce the quality of life and aggravate economic burdens to society. Autograft is the current gold standard method for PNI repair, but it is limited by finite donor tissue supplies, donor site morbidity, and painful neuromas.3,4 To address these limitations, the use of biomaterial-based nerve guidance conduits (NGCs) has become the primary alternative nerve grafting technique.5,6
Various polymers, such as chitosan, gelatin, cellulose, and polylactic acid, are used to construct NGCs.7,8 Hydroxypropyl chitosan (HPCS), as a functional derivative of chitosan, is obtained by introducing hydroxypropyl into the chitosan molecular chain unit glucosamine C6-O. Compared with chitosan, HPCS not only maintains good biocompatibility, biodegradability, and antimicrobial activity but also improves the water solubility, thus broadening the application of chitosan.9–13 Soy protein isolate (SPI) is rich in amino acids, and several studies have shown that SPI-based material can promote cell proliferation and be applied in nerve tissue regeneration.14–16 For example, the SPI-based conduits could efficiently promote sciatic nerve repair. 16 Additionally, previously experiments with HPCS/SPI film have indicated good cytocompatibility with L929 cells as well as good hemocompatibility and skin regeneration efficiency. 17 Therefore, in this work, a series of hydroxypropyl chitosan/soy protein isolate composite sponges (HCSS) with a porous structure were fabricated by a freeze-drying technique. The obtained products were further applied as NGCs. According to previous reports,17,18 the application potential of NGCs is sensitive to their chemical composition. Thus, we tried to identify the optimal ratio of HPCS and SPI to construct HPCS/SPI sponge-based nerve conduits.
In addition to the NGCs that provide structural support and allow access to the PNI location, the incorporation of seed cells, such as Schwann cells (SCs) and stem cells, is a vital factor in the process of peripheral nerve regeneration.19,20 The SCs can be reprogrammed to proliferate and to secrete numerous healing factors. 21 The growth rate after nerve suture was 3.45 mm/day in the rabbit model and 2.5 mm/day in the dog model. The repair rate of nerves is related to the length of the nerve gap. For instance, 22 the times of recoveries vary only from 23 to 26 days for distances of 8–10 mm. Nevertheless, the supply of autologous SCs is derived from invasive nerve biopsies and is limited by low proliferative capacity and high donor site morbidity. 23 Bone marrow mesenchymal stem cells (BMSCs) are accessible seed cell resources that have attracted considerable interest as candidates for cell transplantation because of their inherent high plasticity and low immunogenicity.24,25 Experimental studies using rat BMSCs have indicated that they can differentiate into BMSC-derived SCs in vitro.26,27 Thus, BMSC-derived SCs may be a more attractive source to replace primary SCs for peripheral nerve regeneration.
Among the peripheral nerve repair strategies, the combination of NGCs and stem cell technologies represents a powerful approach for clinical applications. The prepared NGCs provide a channel for axonal growth, and the porous wall structure facilitates the exchange of nutrients and metabolic wastes. Meanwhile, the incoming BMSC-derived SCs can continuously secrete neuro-related factors, creating an appropriate microenvironment for cell proliferation and growth. As shown in Figure 1, this study aims to prepare an optimal HCSS through a series of assays. The BMSCs or BMSC-derived SCs were then incorporated into the conduit to construct a series of cell-loaded NGCs. Finally, the potential of the cell-loaded NGCs for peripheral nerve regeneration was investigated by in vivo experiments.
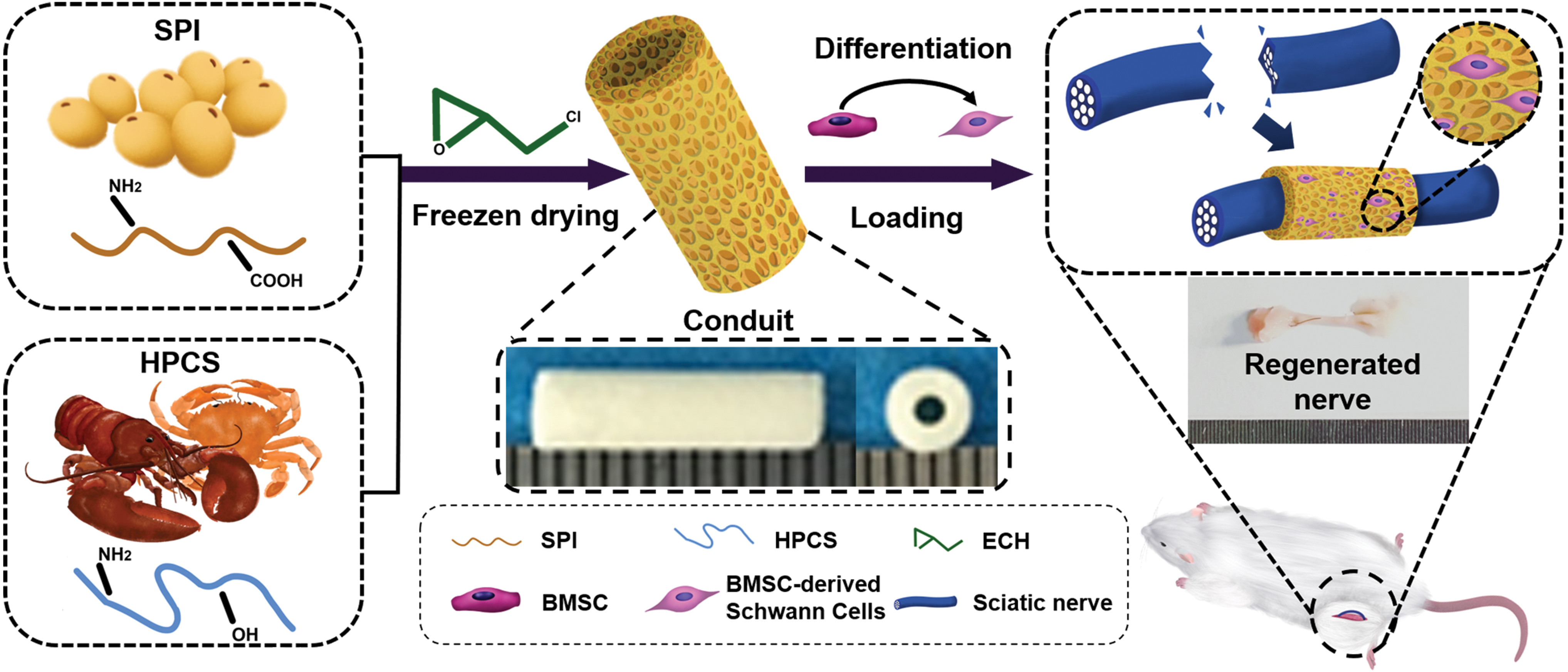
A BMSC-derived Schwann cell-loaded hydroxypropyl chitosan/soy protein isolate composite sponge conduit was constructed to promote peripheral nerve regeneration. BMSC, bone marrow mesenchymal stem cell. Color images are available online.
Materials and Methods
Materials
HPCS (1595642578, 80% degree of substitution) was purchased from Nantong Lvshen Bioengineering Co., Ltd. SPI (Molecular weight = 2.05 × 105) was obtained from DuPont-Yunmeng Protein Technology Co., Ltd. Rat Schwann cells (RSC96) were obtained from Shanghai Institutes for Biological Science (SIBS), Chinese Academy of Sciences. Eagle's alpha minimum essential medium (α-MEM; 51492C), fetal bovine serum (FBS; F8318), penicillin–streptomycin solution (TMS-AB2), trypsin–ethylenediaminetetraacetic acid (EDTA; 59417C), forskolin (F3917), β-mercaptoethanol (M3148), and trans retinoic acid (R2625) were purchased from Sigma–Aldrich Co., Ltd. Antibodies CD11b (ab133357), CD29 (EPR16895), CD45 (ab10558), CD90 (ab3105), GFAP (ab7260), P75 (ab38334), and recombinant human heregulin-beta1 (ab198676) were purchased from Abcam Inc., United Kingdom. Antibody NF200 (AB-C1833B) was purchased from Abmart Shanghai Co., Ltd. The platelet-derived growth factor (PDGF-AA; 500-P46) and basic fibroblast growth factor (bFGF; 500-P18) were purchased from PeproTech Inc. Epichlorohydrin (ECH; 106-89-8), acetic acid (64-19-7), chloral hydrate (302-17-0), paraformaldehyde (30525-89-4), glutaraldehyde (111-30-8), and sodium hydroxide (1310-73-2) were acquired from the Sinopharm Chemical Reagent Co., Ltd. A Live&Dead Viability Assay Kit (KGA263) was purchased from KeyGen Biotech Co., Ltd. A carboxyfluorescein succinimidyl ester (CFSE) cell proliferation kit (70-MTT000) was obtained from Multi Sciences (Lianke) Biotech, Co., Ltd. Transcription Kits (KMM-101) and Reverse Transcription Kits (SCQ-401) were purchased from TOYOBO, Japan. HPCS and SPI were vacuum-dried at 60°C for 24 h before use. Other chemicals were used without further purification. This article does not deal with human trials.
Preparation of HCSS
The HPCS solution and SPI solution were prepared as previously described 17 and then mixed in proportions. ECH was added into the mixtures with constant stirring for 30 min. The content of ECH was 50% of the total weight of HPCS and SPI for the preparation of every sample. After centrifugation at 3000 rpm for 8 min, the mixtures were poured into the plates for freeze-drying treatment. The obtained products were soaked in 5% acetic acid solution for 30 min and washed using distilled water for 48 h. Finally, the HCSS were obtained by a second freeze-drying. These sponges were coded as HCSS-n (n = 30, 50, 70, and 100, respectively). The codes, compositions, and processing technique of HCSS-n are shown in Supplementary Table S1.
Preparation of HPCS/SPI composite conduits
The hollow plastic tube (5 cm in length and 2 mm in inner diameter) was applied as the mold to prepare the HPCS/SPI composite conduits. Briefly, HCSS-50 solution was injected into the mold, followed by inserting the stainless steel needle (1.5 mm in diameter) and sealing both ends of the mold with plugs. The molds were freeze-dried to obtain the HPCS/SPI composite conduits.
Physical and chemical characterization
The infrared spectrum of the HCSS-n powder, HPCS, and SPI raw materials was recorded using a Fourier transform infrared spectrometer (TNZ1–5700, Nicolet, USA) within a scanning range of 4000–500 cm−1. The X-ray diffraction spectrum of the above samples was detected by an X-ray diffractometer (D-Advance, Bruker, USA) with a scanning range of 4–55° and scanning rate of 2°/min. 28 The surface and section structure of the HCSS-n and HCSSC were observed under a scanning electron microscope (SEM; VEGA3, TESCAN, Czech Republic) with a voltage of 20 kV. HCSS-n were cut into square with a size of 3 × 3 × 3 cm and placed in a vacuum drying box at 45°C for drying. Next, the dry mass was recorded as M1. The samples were then placed into a beaker containing deionized water. At the corresponding time points, the weight of the wet sample was measured and recorded as M2. The water uptake ratio (WUR) was calculated by formula 1: WUR (%) = [(M2 − M1)/M1] × 100. The weight of samples with water absorption balance was recorded as W1, and then, each sample was placed in a dryer. The mass was recorded as W2 at the corresponding time points. The water retention ratio (WRR) was calculated by formula 2: WRR (%) = W2/W1 × 100. Finally, HCSS-n were conducted with compression testing. The compression speed was 1.0 mm/min and the applied pressure range was 0–200 N.
Proliferation evaluation
The RSC96 cells were seeded in tissue culture plates supplied with α-MEM, FBS, and penicillin–streptomycin solution and then incubated in a 37°C incubator with 5% carbon dioxide for further use. Then, the CFSE (cell tracking and cell proliferation studies 29 ) was used to evaluate the proliferation of RSC96 in HCSS-n (n = 30, 50, 70, and 100, respectively). First, the CFSE solution (5 μmol/L) was added to an equal volume of RSC96 cell suspension and incubated at 37°C for 10 min. Then, phosphate-buffered saline (PBS) containing 10% FBS was added to the cell suspension to terminate staining. The stained cells were centrifuged at 1000 rpm for 3–5 min. After centrifugation, the cells were suspended and inoculated to six-well plates. The extracts of HCSS-n samples and the medium were added to the six-well plate in a ratio of 1:3 according to GB/T16886.5–2017/ISO. After culturing for 48 h, the cells were digested for flow cytometry analysis (FACS Aria III, BD, USA).
Quantitative real-time polymerase chain reaction
After co-culture of RSC96 and HCSS-n extracts, the cells were washed with PBS twice. Next, total RNA was isolated using RNeasy™ kits (Qiagen), and cDNA was synthesized using High-Capacity cDNA Reverse Transcription Kits. 30 The samples were then loaded into a C1000 Thermal Cycler with a CFX96 Real-Time System (Bio-Rad), which was used with the following parameters: 2 min at 50°C, 15 min at 95°C, a total of 40 cycles (15 s at 94°C, 30 s at 50°C, and 30 s at 72°C). The sequences of Krox20, Zeb2, and TGF-β are shown in Supplementary Table S2.
Transdifferentiating BMSCs toward BMSC-derived SCs
Bone marrow aspirates were harvested from adult rats, and the BMSCs were isolated as follows. Briefly, bone marrow aspirates were seeded in α-MEM supplemented with 10% FBS and 1% penicillin–streptomycin. Cells were allowed to adhere for 72 h followed by removal of non-adherent cells. Medium and non-adherent cells were removed every 3 days thereafter. After a week of cultivation, the adherent cells were detached with 0.25% trypsin-EDTA and then were moved to a new culture flask at a density of 1.0 × 104. To analyze the phenotype cells, the third passage of BMSCs was collected and rinsed with PBS three times and incubated with FITC-CD90, PE-CD73, PE-CD45, FITC-CD34, or their isotypes for 30 min at 4°C. The results were detected by flow cytometry (FACS AriaIII).
To differentiate the BMSCs into a SC-like phenotype (dBMSCs), medium was removed from subconfluent cultures at P2 and replaced with a complete medium containing 1 mM beta-mercaptoethanol (Sigma). After 24 h, the original medium was removed and replaced with a complete culture medium containing all trans retinoic acid (ATRA, 35 ng/mL; Sigma). A further 72 h later, the culture medium was discarded and replaced with differentiation medium containing 10% FBS, 14 μmol/L forskolin (Sigma), 5 ng/ml PDGF-AA (Pepro Tech), 10 ng/mL bFGF (Pepro Tech), and 20 ng/mL recombinant human heregulin-beta1 (H-1β; PeproTech). The induced differentiation medium was changed every 3 days for 2–3 weeks. S100, GFAP, and P75NTR immunofluorescence (IF) staining were used to identify the BMSC-derived SCs. The expression of Ngf, Nt-3, and C-fos of BMSCs and BMSC-derived SCs was validated both by realtime fluorescence quantitative PCR (RTFQ PCR) and by Western blot.
Cell seeding
The HCSS-50 conduits were placed in a 24-well tissue culture plate. After 24 h of incubation, the soaked samples were drained, and 0.5 mL BMSCs or BMSC-derived SCs cell suspension (final density 1 × 105 cells per mL) was injected directly into the conduit lumen. After 72 h of incubation, the BMSCs or BMSC-derived SC-loaded conduits (named HC-1 and HC-2, respectively) were used for in vivo animal experiments.
Animal experiments
This study was performed in compliance with the “Guiding Opinions on the Treatment of Animals” published by the Ministry of Science and Technology of the People's Republic of China and the Guidelines of the Animal Care & Welfare Committee of the Wuhan University School of Medicine. 31 Male Sprague-Dawley (SD) rats were purchased from Wuhan University Laboratory Animal Center (Wuhan, China) and fed a specific pathogen free environment for 7 days to minimize further stress.
Forty rats were randomly divided into Control (Ctrl), HC, HC-1, and HC-2 groups, with 10 rats in each group. After being anesthetized, a longitudinal linear incision was made along the hip and knee joint line to expose the sciatic nerve. A nerve defect model 10 mm in length was resected in each rat. In the Ctrl group, the autogenous nerve segment was reversed 180° and reattached with the two broken nerve ends. In the HC group, the proximal and distal ends of the nerve were inserted into the internal channel of an HCSS-50 conduit. In the HC-1 and HC-2 groups, engineered neural tissue conduits of HC-1 and HC-2 were used to bridge the two broken ends.
To evaluate motor function recovery, the rats' behavioral indicators were tested. Red pigment was applied to the paws of SD rats. The animals were then placed in a passage covered with white papers, allowing them to move freely and recording their footprints. The nerve sciatic functional index (SFI) was calculated by formula 3:
where EPL and NPL represent the distance from the heel to the tip of the third toe. ETS and NTS represent the distance from the first toe to the fifth toe. EIT and NIT represent the distance from the second toe to the fourth toe. The E represents the experimental side, and the N represents the nonoperative side.
The regenerated nerves were harvested and then stored in liquid nitrogen and 4% paraformaldehyde, respectively. S100 and NF200 IF staining was also carried out. The IF images were captured using an inverted fluorescent microscope (IX 73 DP80; Olympus, Japan). Quantitative analysis of positively stained cells was performed with Image-Pro Plus 6.0 software. The detailed steps are as follows. The original fluorescence image was inverted as a contrast, and then by discarding all color information, it was transformed into a grayscale image. The total area of the positive NF200 staining or positive S100 staining regions and the total area of the image were measured with Image-Pro Plus software. Finally, the percentages of positive NF200 staining and positive S100 staining were calculated by dividing the positive area by the total area of the image. Toluidine blue (TB) staining was also performed to test the myelinated nerve fibers. Images of TB were captured using an inverted fluorescent microscope (IX 73 DP80; Olympus, Japan). The myelinated nerve fibers densities (number/mm2) of different groups were calculated by Image-Pro Plus 6.0 software based on the TB images. The IL-β, Krox20, Zeb2, and GAP43 gene expression of the nerve tissue was detected by RTFQ PCR. The primer sequences are shown in Supplementary Table S2.
The gastrocnemius muscles were harvested from the operative side and the normal side. The gastrocnemius muscles on both sides were weighed, and the recovery rate (Mr) was calculated according to formula 4:
where MR represents the weight of gastrocnemius muscle on the operative side, and ML represents the normal side weight.
The operative gastrocnemius muscle was fixed in 2.5% glutaraldehyde and then analyzed by Masson trichrome staining assay. The cross-sectional area of muscle fibers and the average percentage of collagen fibers were calculated.
Statistical analysis
All quantitative data are expressed as mean ± standard deviation. Statistical comparisons were performed using one-way analysis of variance with the SPSS software package (version 23.0). Statistical significance was considered at p < 0.05.
Results
Physicochemical properties of HCSS-n
The Fourier transform infrared spectroscopy spectra of SPI powder, HPCS powder, and HCSS-n are shown in Supplementary Figure S1a, b. HPCS powder exhibited two characteristic peaks at 3431 and 2970 cm−1, which were assigned to the stretching vibrations of -OH and the bending vibration of -NH2, respectively. The SPI powder showed characteristic peaks at 3298 cm−1 (-OH stretch), 1658 cm−1 (hydrogen-bonded C = O stretch), and 1533 cm−1 (-OH bending). The characteristic peaks of -OH at 3431 cm−1 of HPCS powder and 3298 cm−1 of SPI powder shifted to 3299–3440 cm−1 in HCSS-n (n = 30, 50, 70, and 100, respectively). This phenomenon suggested that the -OH of HPCS and SPI were cross-linked by ECH. Compared with the raw materials, the amide I (1630–1678 cm−1) and amide II (1505–1561 cm−1) of HCSS-n had displacement, and the peak intensity changed, indicating that the -NH2 of HPCS and SPI were also cross-linked by ECH.13,32–34
The X-ray diffraction patterns of HCSS-n and raw materials are shown in Supplementary Figure S1c, d. The SPI powder exhibited two obvious crystal peaks at 8.7° and 19.5°, and the HPCS powder exhibited two characteristic peaks at 7.6° and 21.3°. The characteristic peaks of HCSS-n became weakened or even disappeared, indicating a chemical cross-linking reaction between SPI and HPCS. The surface and cross-sectional photographs of HCSS-n (n = 30, 50, 70, and 100) are shown in Figure 2a, b. All samples exhibited porous and interconnected microstructures. With the increase of SPI content, the pore size of HCSS-n decreased gradually.
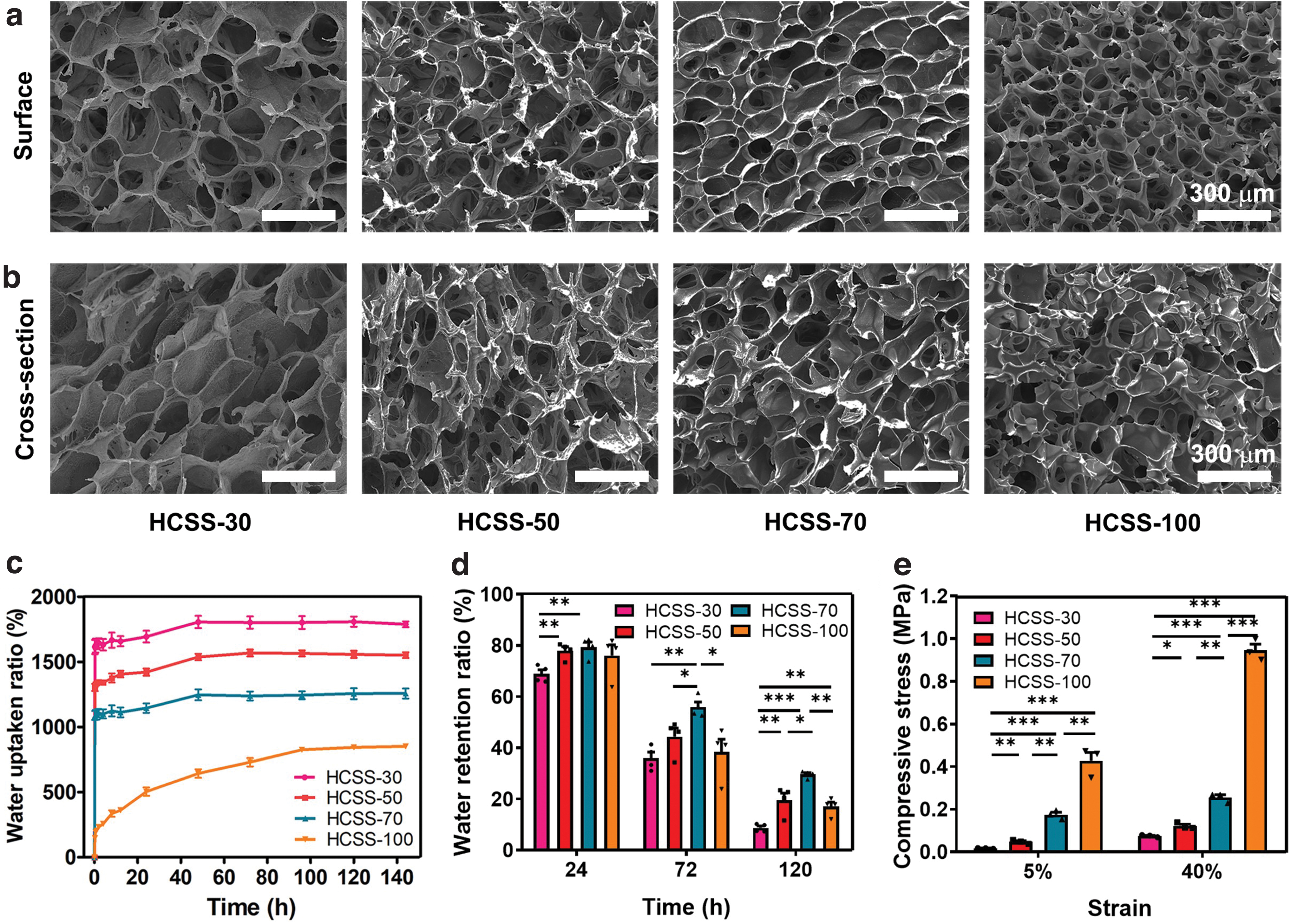
SEM images of the surface and cross-sectional structures in HCSS-n (n = 30, 50, 70, and 100, respectively)
The WUR and WRR of HCSS-n (n = 30, 50, 70, and 100, respectively) are demonstrated in Figure 2c, d. Each group's WUR rose rapidly before 2 h and then entered a slow growth process until reaching a balance around 144 h. Among them, the WURs of HCSS-30, HCSS-50, HCSS-70, and HCSS-100 reached 1788 ± 45%, 1551 ± 43%, 1257 ± 76%, and 834 ± 64%, respectively. HCSS has a strong water absorption capacity, which helps absorb medium, blood, or liquids containing neurotrophic factors. The WRRs of HCSS-30, HCSS-50, HCSS-70, and HCSS-100 reached 8 ± 1.6%, 19 ± 5.5%, 29 ± 1.4%, and 17 ± 3.5%, respectively, after 5 days of placing them in the dryer. With the increase of SPI content, the sponges' water retention ability increased gradually and reached a maximum value in HCSS-70. The WRR for HCSS-100 is lower than that for HCSS-70 at 72 and 120 h, which may be the structure of HCSS-100 gradually deteriorated due to the brittleness of SPI. Thus, HCSS-50 and HCSS-70 have good water retention ability, which is more conducive to retaining liquid, such as the medium containing seed cells, thus providing effective support for the subsequent construction of engineered neural tissue conduits (HC-1 and HC-2).
Proper mechanical properties are essential for NGCs that both support axon growth and avoid mechanical injury. To investigate the mechanical properties of HCSS-n (n = 30, 50, 70, and 100, respectively), we tested compressive stress–strain curves. As shown in Figure 2e, the compression strengths of HCSS-30, HCSS-50, HCSS-70, and HCSS-100 were 0.07 ± 0.01 MPa, 0.12 ± 0.01 MPa, 0.32 ± 0.02 MPa, and 0.94 ± 0.05 MPa at 40% strain, respectively, indicating that HCSS-n can withstand large strain and has sufficient compressibility for peripheral nerve repair surgery in SD rats. 35
Biocompatibility evaluation of HCSS
Figure 3a shows the SEM of HCSS-n (n = 30, 50, 70, and 100, respectively) cultured with RSC96 cells. The RSC96 cells could adhere to the sponges' surface and enter the interior for proliferation and growth. The RSC96 cells were mostly spherical on HCSS-100 and fusiform on HCSS-30, HCSS-50, and HCSS-70, which may be because the introduction of HPCS was conducive to the adhesion growth of RSC96 cells. The RSC96 cells showed the best growth status on HCSS-50. This result may be due to the synergistic effect of the right proportion of HPCS and SPI, which promotes cell growth. 36 In Figure 3c, the cell viability of RSC96 in HCSS-n increased with low SPI content but decreased with high SPI content. At 48 h, the cell viability of HCSS-50 was significantly higher than that of blank control. The results indicated that an appropriate SPI content could enhance cell viability.

SEM images of RSC96 cells cultured on the cell culture plate and on the surfaces of HCSS-n (n = 30, 50, 70, and 100, respectively)
The HCSS-n (n = 30, 50, 70, and 100, respectively) were cultured with rabbit whole blood for 1 h to evaluate blood compatibility. As shown in Figure 3b, the erythrocytes are round and biconcave in the Ctrl group. The erythrocytes cultured in the HCSS-n (n = 30, 50, 70, and 100, respectively) showed similar morphology to the Ctrl group. These results suggested that HCSS does not cause erythrocyte fragmentation. Figure 3d shows the hemolysis rate of HCSS-n was lower than 5%. Plasma recalcification time (PRT) is a marker of biomaterial-induced coagulation activation. 37 The PRT values of HCSS-n are shown in Figure 3e. The PRT values of HCSS-30, HCSS-70, and HCSS-100 were similar to that of the control, whereas the PRT value of HCSS-50 was significantly lower than that of the control. In conclusion, HCSS did not cause erythrocyte rupture and showed good hemocompatibility.
Effects of HCSS-n extracts on the proliferation and growth of RSC96 cells
CFSE is an indicator to detect cell division and proliferation. When CFSE enters cells, it fluoresces, and the fluorescence intensity of CFSE dye is transmitted during cell division, showing a decreasing trend in daughter cells.38,39 Figure 4a is the division diagram of RSC96 cells cultured with HCSS-n extracts. RSC96 cell division was higher in the HCSS-70 group than in the Ctrl group. Figure 4c shows the statistical analysis of the proliferation index, which indicates that HCSS-30, HCSS-50, and HCSS-70 promote RSC96 proliferation to different degrees.
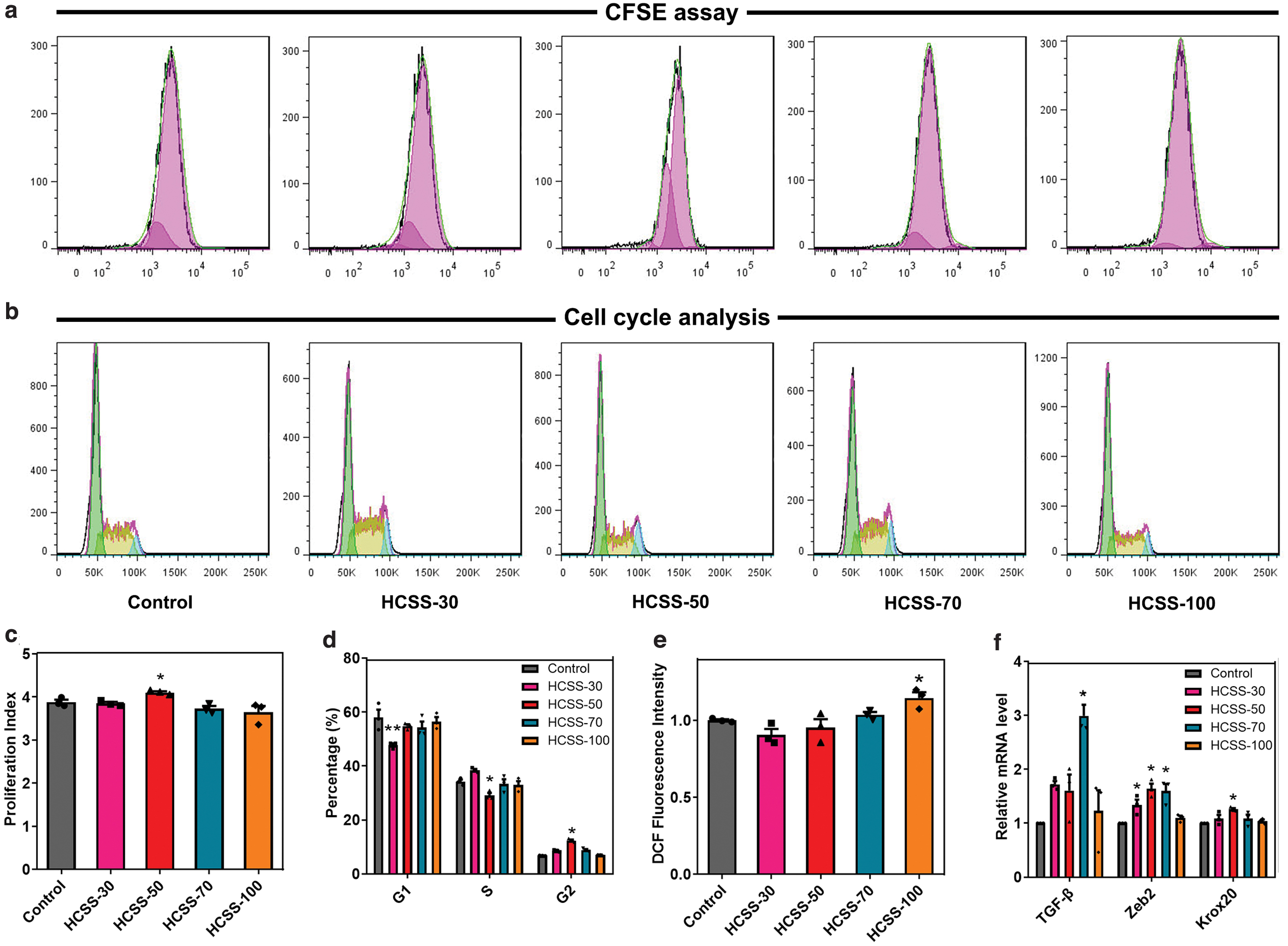
Effects of HCSS-n (n = 30, 50, 70, and 100, respectively) extracts on the proliferation and growth of RSC96 cells. RSC96 cell proliferation was evaluated by measuring CFSE fluorescence
Figure 4b, d shows the results of the cell cycle by flow cytometry. Compared with the Ctrl group, the proportion of HCSS-30, HCSS-50, and HCSS-70 groups in the G1 phase was lower than that in the Ctrl group. In contrast, the G1 proportion of the HCSS-100 group was higher than that of the Ctrl group. This pattern can be attributed to the RSC96 cells treated with HCSS-100 being mainly in the RNA and protein synthesis phase. The proliferation of RSC96 cells was speculated to be inhibited under the high SPI concentration. In the G2 phase, the DNA content of the HCSS-100 group was similar to that of the Ctrl group, whereas the DNA content of the HCSS-30, HCSS-50, and HCSS-70 groups was higher than that of the Ctrl group. These results indicated that HCSS-100 had little effect on the G2 phase of RSC96 cells.
Excessive production of reactive oxygen species (ROS) will damage cell structure and overall function and affect their viability, proliferation, and redox status. 40 As shown in Figure 4e, our study indicated that after the stimulation of HCSS-100 extracts on RSC96 cells, the intracellular ROS level was higher than in the Ctrl group, indicating that HCSS-100 had a damaging effect on RSC96 cells. Compared with the Ctrl group, ROS levels in the HCSS-30, HCSS-50, and HCSS-70 groups showed a decreasing trend. This pattern suggested that intracellular ROS production gradually decreased with decreasing SPI content, providing a more favorable microenvironment for cell growth.
Finally, we examined the RNA expression levels of TGF-β, Zeb2, and Krox20 in RSC96 cells by qPCR. TGF-β is involved in regulating cell growth, cell differentiation, apoptosis, cellular homeostasis, and other cellular functions. Zeb2 is a neuro-related transcription factor. 41 Krox20 is an important transcription factor associated with myelin formation. 42 In Figure 4f, the RNA expression of HCSS-100 was the lowest, which may be related to the SPIs high concentration. Compared with the Ctrl group, the RNA expression of TGF-β, Zeb2, and Krox20 in the HCSS-50 group was mostly higher than that in the other groups. This result indicated that the appropriate proportion of HCSS-n promoted the expression of neuro-related factors. Thus, after the above series of experiments, and according to the properties of the excellent NGCs,43–45 the composite sponge containing 50% SPI was selected to construct NGCs.
Loading BMSCs and BMSC-derived SCs in the conduits
The morphological and phenotypic identification results of rat BMSCs and BMSC-derived SCs are shown in Figure 5. According to Supplementary Figure S2a, the positive expression rates of CD90 and CD29 molecules in BMSCs reached 97.0 ± 1.34% and 97.2 ± 2.56%, and the negative expression rates of CD45 and CD11b were 9.37 ± 2.12% and 7.40 ± 1.89%, respectively. This result indicated that BMSCs collected by this work were of high purity.46,47 Supplementary Figure S2b, c shows representative morphologies of Calcein Acetoxymethyl Ester staining for BMSCs and BMSC-derived SCs, where green represents living cells. BMSCs exhibited spiral growth with a flat cell body and spindle shape. In contrast, BMSC-derived SCs exhibited the typical morphology of SCs (long spindle shape, smaller cell body, and elongated pseudopods) after 3 weeks of induction. 48 IF showed that BMSC-derived SCs expressed S100, GFAP, and P75NTR, indicating that BMSC-derived SCs had the typical structural characteristics of SCs (Fig. 5a). 49 Furthermore, as shown in Figure 5b and c, compared with the BMSCs group, the mRNA expression of c-fos, Nt-3, and Ngf, as well as the protein expression levels of S100, GFAP, and P75NTR in the BMSC-derived SCs group, were significantly increased, suggesting that BMSC-derived SCs cultured in this work can function as SCs in vitro.

S100, GFAP, and P75NTR expression in BMSC-derived Schwann cells was determined by immunocytochemistry
We also examined the loading capacity of conduits with BMSCs and BMSC-derived SCs. Figure 5d shows that a large number of BMSCs were attached in the lumen of HCSSC. BMSCs can attach to the surface and enter the sponge pores for proliferation and growth. Figure 5e shows an SEM image of cultured BMSC-derived SCs and conduit. Similarly, BMSC-derived SCs adhered tightly to the conduit's lumen and penetrated the sponge pores, clearly indicating that the sponge's structure can create a supportive microenvironment to improve cell growth. Thus, BMSCs and BMSC-derived SCs were uniformly distributed in the lumen of conduits after inoculation.
Evaluation of nerve repair effect
To detect motor function recovery, the SD rats were placed on a clean bench to walk freely, and their behaviors were videotaped. The representative images of SD rats standing are shown in Figure 6a. Rats in the HC-2 group showed a paw posture when standing, which was not significantly different from that in the Ctrl group. Figure 6b shows representative footprints of walking rats. The footprints show that the Ctrl group had the largest paw extension and the best functional recovery. Nerve sciatic functional index (SFI) is an evaluation method to measure the degree of motor function recovery in SD rats 50 ; it ranges from 0 to −100, where 0 represents complete functional recovery and −100 represents a complete functional loss. In Figure 6d, the SFI value of the Ctrl group was −51.81 ± 1.06, and those of the HC, HC-1, and HC-2 groups were −76.94 ± 1.74, −65.58 ± 2.69, and −53.2 ± 0.87, respectively. Figure 6c–e shows the compound muscle action potential (CMAP) waveform and the statistical analyses of CMAP amplitude for the operation side, respectively. The CMAP amplitude of HC was significantly lower than that of the HC-2 and HC-1 group, which suggested that the conduits loaded with BMSCs and BMSC-derived SCs could better connect the damaged sciatic nerve and conduct electrical signals after 3 months. The results of nerve conduction velocity (Fig. 6f) and latency (Fig. 6g) analyses were similar to the results of CMAP. Consequently, the repair efficiency of HC-2 was close to that of the Ctrl group, indicating that BMSC-derived SCs continuously secreted nerve-related factors to promote nerve regeneration. 26
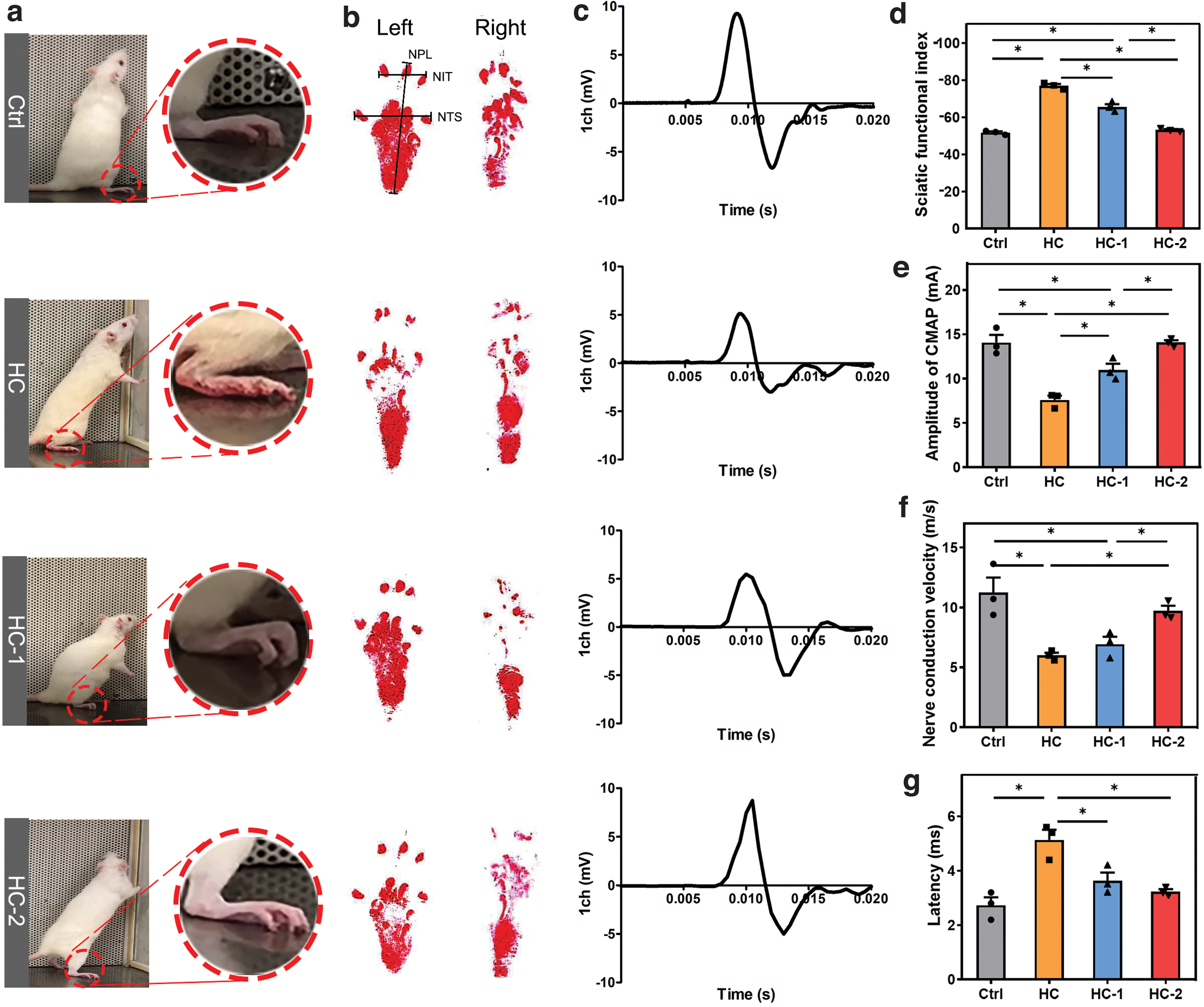
A general view of the rat standing to show the paw posture on the operative side
After the detection of SFI, the regenerated nerves were examined with hematoxylin and eosin staining and IF staining. Figure 7a shows representative IF staining images of regenerative nerves in each group. NF200 was a specific marker for axons, and S100 was a specific marker for SCs. 27 NF200 and S100 were positively expressed to different degrees in each group. Analyses of the positive expression rates of NF200 and S100 are shown in Figure 7b, c. The positive expression rates of NF200 in the HC, HC-1, and HC-2 groups were lower than that in the Ctrl group. However, the positive expression rate of S100 in the HC-2 group was similar to that in the Ctrl group. Furthermore, TB staining was performed to evaluate the myelinated nerve fibers. As shown in Supplementary Figure S4, all the groups presented the myelinated nerve fibers. The average density of myelinated nerve fibers in the Ctrl, HC, HC-1, and HC-2 groups were 1348.02 ± 179.16, 800.73 ± 112.96, 872.36 ± 79.14, and 1327.82 ± 239.72, respectively. The myelinated nerve fiber density in the HC-2 group was similar to that in the Ctrl group, indicating BMSC-derived SCs could efficiently promote the myelination of regenerated nerve fibers. Notably, the myelinated nerve fiber density of the HC-2 group was significantly higher than that of the HC and HC-1 groups, suggesting that BMSC-derived SCs could promote the myelination of regenerated nerve fibers.
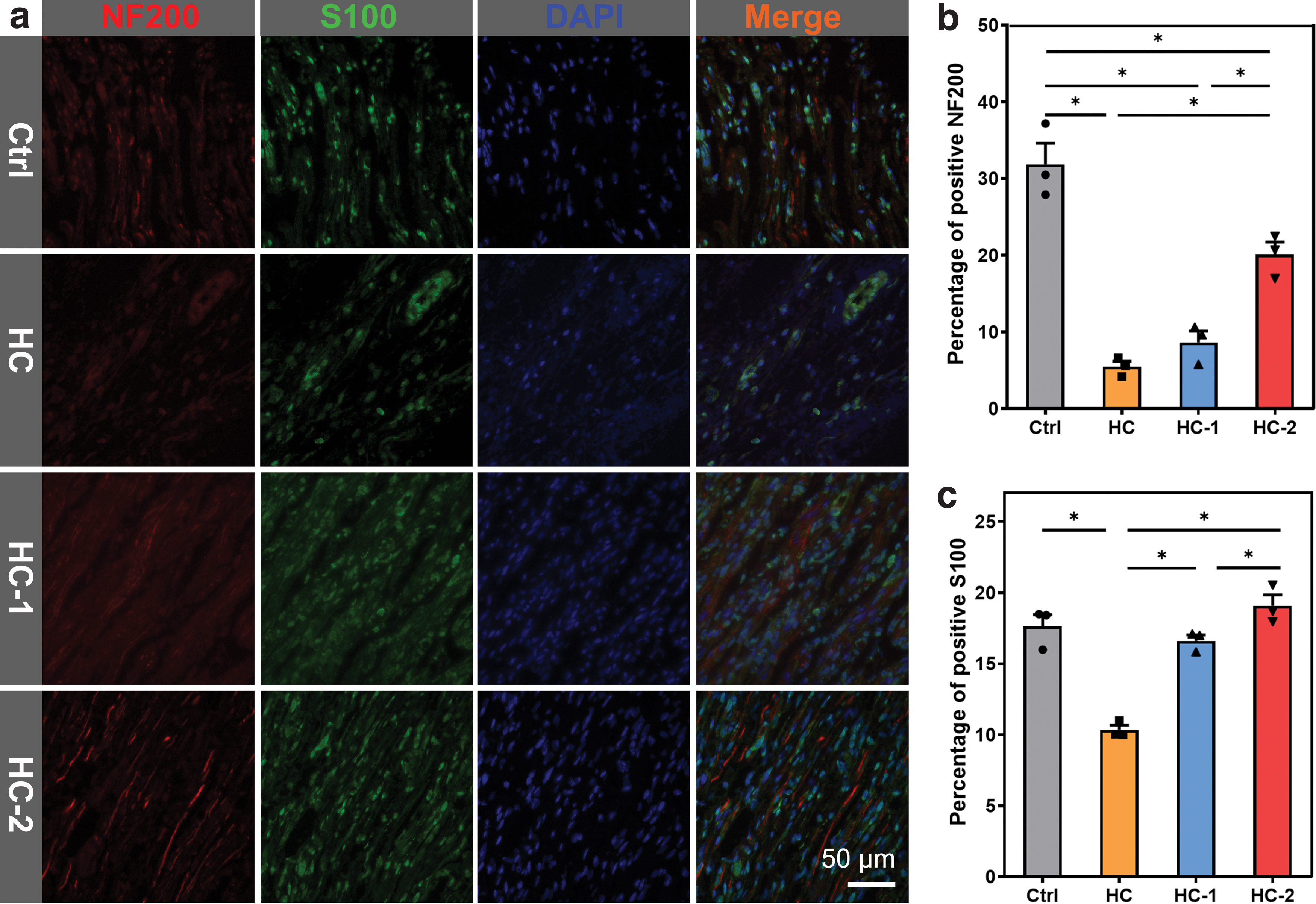
Longitudinal sections of regenerated nerve immunostained with the axonal marker NF-200 (red) and the Schwann cells marker S100 (green)
Figure 8 shows the RNA levels of IL-1β, Krox20, Zeb2, and GAP43 in the regenerated nerve. IL-1β is a pluripotent cytokine involved in many immune and proinflammatory responses. Inflammation is a unified process of injury and anti-injury. As shown in Figure 8a, the expression of IL-1β in the Ctrl group was significantly higher than that in the HC, HC-1, and HC-2 groups, indicating that the expression of 1L-β can promote nerve regeneration at an appropriate level of inflammation. 51 Krox20 is an important transcription factor associated with myelin formation. 42 Zeb2 is a neuro-related transcription factor. 41 GAP43 is a growth protein, which promotes the development of neuronal growth cones. 52 The expression level of Krox20 in the HC-2 group was higher than that in the HC and HC-1 groups and similar to that in the Ctrl group (Fig. 8b). The expression of Zeb2 (Fig. 8c) in the HC-2 group was significantly higher than that in the Ctrl group. The expression of GAP43 (Fig. 8d) in the HC and HC-1 groups was significantly lower than that in the Ctrl group, whereas the HC-2 group was similar to the Ctrl group. Consequently, the HC-2 group had a better functional return, which was significantly higher than both the HC and HC-1 groups and close to that of the Ctrl group, indicating that BMSC-derived SCs were continuously secreting neuro-related factors to recover the defective nerve.

The relative mRNA level of IL-1β (n = 3)
When the sciatic nerve is severely injured, the gastrocnemius muscle slowly atrophies until necrosis. After the defect nerve begins to recover, the atrophy of the muscle will gradually recover with the sciatic nerve's reinnervation. 53 Supplementary Figure S3a shows representative images of gastrocnemius muscles in the normal and operative sides. The gastrocnemius muscles on both sides were weighed to compare the recovery rate of each group. Supplementary Figure S3c indicates that the recovery ratios of the HC and HC-1 groups were lower than that of the Ctrl group, whereas the HC-2 group was similar to the Ctrl group. Supplementary Figure S3b shows Masson staining images of the gastrocnemius muscles on the operative side of each group. The red area represents muscle fibers, and the blue area represents collagen fibers. In the Ctrl group, the muscle fibers were tightly arranged with minimal collagen staining. The muscle fibers in the HC-1 and HC-2 groups were closely arranged, with a small amount of blue collagen fibers in the muscle space. However, the muscle fibers were loose and disorderly in the HC group, and a large amount of collagen was present. Supplementary Figure S3d, e shows that the average percentage of collagen fiber area in the HC-2 group was higher than that in the Ctrl group, but the cross-sectional area of muscle fibers was similar to that in the Ctrl group. In summary, the repair effect of the HC-1 group was intermediate to the HC group and HC-2 group. These results indicated the critical role of seed cells and that the BMSC-derived SCs were more favorable for peripheral nerve regeneration than BMSCs.
Discussion
In this study, the tissue-engineered conduits applied for peripheral nerve repair were constructed by combining HPCS/SPI sponge conduits and BMSCs or BMSC-derived SCs. Among them, the HPCS/SPI conduits could provide channels for peripheral nerve regeneration, and the seed cells BMSC-derived SCs could continuously secrete neuro-related factors to promote peripheral nerve regeneration. The strategy of combining HPCS-based nerve conduit with BMSC-derived SCs has the dual effect of promoting peripheral nerve regeneration.
Chitosan is a cationic polysaccharide derived from chitin and has been applied in the fields of wound dressing, peripheral nerve regeneration, and drug delivery carriers due to its antimicrobial activity, biocompatibility, and biodegradability. 54 Previous research has proven that chitosan can facilitate SCs proliferation and nerve axons growth. 55 Because of these merits, chitosan has been used for constructing nerve conduits by a variety of preparation methods and modification means. Rao et al. reported that the aligned chitosan nanofiber hydrogel grafted with RGI/KLT peptide nerve conduit prepared by electrospinning and mechanical stretching methods could repair sciatic nerve defects. 56 Not only chitosan-based conduits can promote nerve regeneration, but their degradation products, chitooligosaccharides (COS), can also promote nerve regeneration. For example, Zhao et al. showed that COS can promote the migration of macrophages and improve the microenvironment, thereby promoting nerve regeneration. 57 However, chitosan can only dissolve in acidic systems, which greatly limits the possibility of preparing more versatile nerve conduits. HPCS acts as the functional derivative of chitosan, not only maintains good biocompatibility, biodegradability, and antimicrobial activity but also improve the water solubility, thus broadens the application of chitosan. Previous researches revealed that HPCS-based materials have good performance for the application in tissue engineering.17,18 In this study, the resultant HPCS/SPI sponge also could support the attachment and proliferation of RSC96 cells and promoted the secretion of neuro-related factors, which is consistent with previous researches.
The nerve conduits itself only provide a substrate for the adhesion and migration of seed cells. When the number of seed cells is insufficient, it is difficult for nerve axons to regenerate. However, the application of SCs is limited by low proliferative capacity and high donor site morbidity. 23 Among the large number of seed cells, BMSCs are an accessible source that permit in vitro expansion and differentiation into BMSC-derived SCs. 24 Cai et al. reported that BMSCs could be induced to differentiate into BMSC-derived SCs, and the results of in vitro myelination provided evidence that the BMSC-derived SCs were functionally mature. 58 Even with withdrawal of inducing medium and neuronal cues, markers are associated with myelination remained. 59 As expected, in this experiment, the obtained BMSC-derived SCs have the ability to perform the basic functions of SCs, such as secreting the c-fos, Nt-3, and Ngf. These characteristics confirm that BMSC-derived SCs are more effective than BMSCs for promoting nerve regeneration.
Furthermore, the BMSC-derived SCs were inoculated as seed cells into HPCS/SPI conduits to evaluate their effectiveness in nerve repair. In the present study, animals in the Ctrl group showed largest paw extension after implantation, whereas the HC groups showed toe loss. Therefore, we preliminary found that the addition of seed cells could improve the effect of nerve regeneration. Subsequently, the IF staining and TB were performed to explore the remyelination in the Ctrl, HC, HC-1, and HC-2 groups. The results indicated that the repair effect of HC-2 was inferior to the Ctrl group. SCs are the cell type responsible for the formation of myelin. Therefore, we speculate that BMSC-derived SCs may not be able to play all functions of SCs. However, compared with the HC-1 group, the HC-2 group greatly improved the expression of S100 and NF200, as well as myelinated nerve fiber density. This result is consistent with the report of Hou et al. that an nerve conduit combined with BMSC-derived SCs could promote remyelination to a certain extent and was superior to conduits combined with BMSC. 60 Moreover, experiments including RNA levels of IL-1β, Krox20, and Zeb2, as well as gastrocnemius muscle evaluation showed the same results. Based on the above analysis, we postulate that BMSC-derived SCs may play a role in the nerve regeneration, but cannot completely replace SCs. In the future, whether the HPCS/SPI conduits can promote the differentiation of BMSCs into BMSC-derived SCs needs to be further explored. Otherwise, we also could combine the BMSC-derived SCs with neuro-related factors (such as NT-3, NT-4, BDNF, CTNF, and NGF) to improve the repair effect. 43
Conclusions
The goals of this work were to screen out the optimal HCSS and to combine it with BMSC-derived SCs for peripheral nerve tissue engineering. Based on the results of physicochemical and biological characterization, HCSS-50 exhibited the best performance in terms of mechanical properties, water retention, and biocompatibility. Then, BMSCs and BMSC-derived SCs were inoculated as seed cells into the lumen of HCSS-50 sponges to prepare cell-loaded conduits. The nerve repair effects of the obtained cell-loaded conduits were evaluated by behavioral, electrophysiological, and histological tests. The BMSC-derived SC-loaded conduits achieved better nerve tissue regeneration and functional recovery than neat HCSS-50 conduits and BMSCs-loaded conduits. Thus, this work provides a new candidate NGC for supporting peripheral nerve regeneration.
Footnotes
Acknowledgment
The authors thank the Experimental Teaching Center of Basic Medical Sciences, Wuhan University, for the technical support.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. NSFC 81871493), the Medical Science Advancement Program (Clinical Medicine) of Wuhan University (Grant No. TFLC2018003), and the Horizontal Research Program of Zhengzhou University (Grant No. 24110005).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.