
Abstract
The autologous synovium is a potential tissue source for local induction of chondrogenesis by tissue engineering approaches to repair articular cartilage defects that occur in osteoarthritis. It was the aim of the present study to ascertain whether the aging of human osteoarthritic patients compromises the chondrogenic potential of their knee-joint synovium and the structural and metabolic stability of the transformed tissue.
The patients were allocated to one of the following two age categories: 54–65 years and 66–86 years (n = 7–11 donors per time point and experimental group; total number of donors: 64). Synovial biopsies were induced in vitro to undergo chondrogenesis by exposure to bone morphogenetic protein-2 (BMP-2) alone, transforming growth factor-β1 (TGF-β1) alone, or a combination of the two growth factors, for up to 6 weeks. The differentiated explants were evaluated morphologically and morphometrically for the volume fraction of metachromasia (sulfated proteoglycans), immunohistochemically for type-II collagen, and for the gene expression levels of anabolic chondrogenic markers as well as catabolic factors by a real-time polymerase chain reaction analysis.
Quantitative metachromasia revealed that chondrogenic differentiation of human synovial explants was induced to the greatest degree by either BMP-2 alone or the BMP-2/TGF-β1 combination, that is, to a comparable level with each of the two stimulation protocols and within both age categories. The BMP-2/TGF-β1combination protocol resulted in chondrocytes of a physiological size for normal human articular cartilage, unlike the BMP-2-alone stimulation that resulted in cell sizes of terminal hypertrophy. The stable gene expression levels of the anabolic chondrogenic markers confirmed the superiority of these two stimulation protocols and demonstrated the hyaline-like qualities of the generated cartilage matrix. The gene expression levels of the catabolic markers remained extremely low. The data also confirmed the usefulness of experimental in vitro studies with bovine synovial tissue as a paradigm for human synovial investigations.
Our data reveal the chondrogenic potential of the human knee-joint synovium of osteoarthritic patients to be uncompromised by aging and catabolic processes. The potential of synovium-based clinical engineering (repair) of cartilage tissue using autologous synovium may thus not be reduced by the age of the human patient.
Impact statement
Our data reveal that in younger and older age groups alike, synovial explants from osteoarthritic joints can be equally well induced to undergo chondrogenesis in vitro; that is, the chondrogenic potential of the human synovium is not compromised by aging. These findings imply that the autologous synovium represents an adequate tissue source for the repair of articular cartilage in clinical practice by tissue engineering approaches in human patients suffering from osteoarthritis, independent of the patient's age.
Introduction
Osteoarthritis (OA) is one of the diseases for which there currently exists no effective, biologically based therapeutic strategy to reconstitute the damaged layer of articular cartilage. During the early stages of OA, cartilage damage is confined to discrete structural lesions. However, owing to the intrinsically poor healing capacity of cartilaginous tissue, the lesioning process, once initiated, cannot be arrested by spontaneous repair. 1 Ultimately therefore, the entire layer is implicated and destroyed.2,3 Although several treatment strategies are currently investigated to promote the repair of articular cartilage, such as the microfracturing technique, 4 the grafting of osteochondral tissue, 5 or the implantation of autologous chondrocytes, 6 the tissue thereby formed is biologically and mechanically grossly inferior to native articular cartilage. 7
For the engineering of cartilage tissue in vitro and in vivo, multipotential stromal cells (MSCs) are often preferred to committed chondrogenic cells, since their differentiation can be induced under more physiological stimulation conditions. 8 Although MSCs are usually drawn from the bone marrow and fat tissue, the synovium is likewise a rich source of such cells exhibiting a very high chondrogenic potential.9–11 Also anatomically, the proximity of the synovium to the layer of articular cartilage would render it more a suitable source of MSCs for the in situ engineering of this tissue.
We had developed in vitro a suitable stimulation protocol for the formation of a hyaline-type of cartilage tissue from both isolated human synovial cells 12 and bovine synovial explants.13–15 Specifically, this protocol is able to arrest also the chondrocyte downstream differentiation into terminal cell hypertrophy and to prevent the associated intercellular matrix mineralization. 16
Synovial MSCs isolated from the knee joints of human OA patients of various ages have shown chondrogenic potential.10,12 On the contrary, the chondrogenic potential of synovial explants derived from OA synovium is still unknown, and in particular, the possible modulating influence of the presence of a number of other cells, such as fibroblasts, endothelial cells, and inflammatory cells, as well as the extracellular matrix (ECM) that may act also as a reservoir of cytokines.17–19 Furthermore, although many researchers have reported that the donor's age affects the differentiation potential of MSCs derived from bone marrow,20–22 but most likely not the synovium-derived ones,10,11,23 this is still unknown for synovial tissue explants.
The question thus arises whether the synovial explants derived from elderly OA patients retain their chondrogenic potential, or if this potential is deteriorating with age. It was the aim of the present study to address this question.
Materials and Methods
Explants of synovial tissue were obtained from osteoarthritic patients who were allocated to two age categories (54–65 and 66–86 years). The choice of 65 years was arbitrarily chosen, but based on the findings that the highest numbers of OA cases occur at the ages of 55–64 years, and mild OA peaks around 65 and then decreases again 24 ; however, the general prevalence of OA increases almost linearly with age.25,24
The donor exclusion criteria applied were chemotherapy, any systemic disease (such as diabetes and autoimmune diseases), septic arthritis, intra-articular use of steroids, viscosupplementation, and so on, or long-term pain therapy. After the explants were cultured for up to 6 weeks with bone morphogenetic protein-2 (BMP-2) or transforming growth factor-β1 (TGF-β1), or with a combination of BMP-2/TGF-β1(to prevent terminal differentiation to chondrocyte hypertrophy and matrix mineralization 16 ), the differentiated tissue of each category was then analyzed for its cartilage properties morphologically (chondrocyte/lacunae formation) and histochemically (volume fraction of metachromasia), as well as immunohistochemically (type-II collagen); and also the expression profiles of anabolic and catabolic marker genes were investigated.
Tissue preparation, growth factors, and culturing
Synovial tissue was obtained (surgical waste material) from the knee joints (medial and lateral femoral condyle areas) of OA patients (grade 3 or 4 of OA; aged 54–86 years) who were undergoing total knee replacement surgery. A total number of 64 patients served as donors. Informed consent and local ethics commission approval were obtained. Synovial explant differentiation was induced by BMP-2 alone, TGF-β1alone, or with a combination of BMP-2/TGF-β1. Negative control groups with an absence of growth factors were also established. In the gene expression analysis (see below), this control group is not shown separately in the Results section since used for the ratio computation of the gene activity levels.
Details of tissue preparation and culturing
Quadratic pieces of tissue with a side length of ∼2 mm (and a thickness of 1–2 mm, that is, including the intimal and subintimal tissue layers) were prepared and sandwiched between two layers of agarose for culturing as previously described.13,14 To induce their chondrogenic differentiation, the synovial explants were exposed to BMP-2 alone (2000 ng/mL; InductOs®, Pfizer/Medtronic, Minneapolis, MN), TGF-β1 (10 ng/mL; Peprotech, Rocky Hill, NJ), or a combination of the two growth factors (at the aforementioned concentrations) for 2, 4, or 6 weeks (n = 7–11 explants [donors] per experimental group); from the total of 64 donors we were able to culture on average 3–4 explants per experimental group and per donor (i.e., 12–16 explants totally per donor). Owing to the limited availability of the donor material, the explants derived from the group of younger patients were cultured for only 4 weeks. Thus, the age group comparison was restricted to the 4-week time duration of culturing.
Morphology, morphometry, histochemistry, and immunohistochemistry
At the end of each culturing period (2, 4, or 6 weeks), a portion of the specimens were processed for morphology, morphometry of cell sizes, quantification of metachromasia (sulfated proteoglycans; internal normalization being performed by referring to the staining intensity of non-OA articular cartilage areas of the same joint) after staining with Toluidine Blue O, and for the immunohistochemical demonstration of type-II collagen.
The volume fraction of metachromasia was determined from the light micrographs by the point counting technique, which was applied in accordance with stereological principles.26,27 The mean cell volumes were estimated using the point sampled intercept method 28 and systematic random sampling strategies. 26
For the immunohistochemical demonstration of type-II collagen, the sections were first exposed to hyaluronidase, and then to a type-II collagen antibody (clone CII C1; Hybridoma Bank, Iowa City, IA). Immunoreactivity was enhanced by applying first the avidin/biotin/peroxidase complex (Vector Laboratories) and then biotinyl tyramide (Perkin Elmer, Waltham, MA). Cell nuclei were counterstained with hematoxylin. The sections were evaluated and photographed in a Nikon Eclipse E1000 light microscope; for details see Refs.14,16
Isolation of RNA, reverse transcription, and real-time polymerase chain reaction analysis
At the end of each culturing period, samples were subjected to a real-time polymerase chain reaction analysis to determine the gene expression levels of key anabolic cartilaginous markers (collagen types I, II, X, and XI, aggrecan, alkaline phosphatase, cartilage oligomeric matrix protein [COMP], lubricin, matrilin-1, osteocalcin, and Sox-9) and of a panel of catabolic factors (interleukins [IL])-1β, -4, and -6, a disintegrin and metalloproteinase with thrombospondin motifs 4 [ADAMTS-4], cyclooxygenase-2 [Cox-2], inducible nitric oxide synthase [iNOS], matrix metallopeptidase-13 [MMP-13], and tumor necrosis factor-alpha [TNF-α]); for details (and abbreviations): see Tables 1–3.
Primers and Probes Used for Real-Time Polymerase Chain Reaction (Anabolic Factors)
Primers and Probes Used for Real-Time Polymerase Chain Reaction (Catabolic Factors and 18s rRNA)
Abbreviations
The total amount of RNA was isolated using the RNeasy Micro Kit (Qiagen, Basel, Switzerland) in accordance with the manufacturer's standard protocol, which included a deoxyribonuclease-catalyzed digestion step. The concentrations of mRNA were measured in a NanoDrop spectrophotometer ND-1000 (NanoDrop Technologies, Wilmington, DE). The samples of mRNA were stored at −70°C and then subjected to reverse transcription (200 ng aliquots) using an ImProm-II- Reverse Transcription (Promega, Madison, WI). For methodological details of cDNA processing see Refs.14,16; for use of primers and probes, and final concentrations: see Tables 1 and 2.
The reactivity of the primers and probes was confirmed using samples of cDNA that had been prepared from human cartilage or the stimulated human synovial explants. The levels of mRNA were quantified relative to those for 18S rRNA using the comparative cycle threshold method. 12 For these calculations, the gene expression levels in unstimulated synovial explants served as the basis for comparison. 29
Statistical analyses
All statistical analyses were conducted using Prism 8 (version 8.4.3; GraphPad Software, San Diego, CA). For volume fractions of metachromasia, comparisons between two sets of data were statistically evaluated by the unpaired t-test. Comparisons between multiple groups were evaluated by one-way analysis of variance and then by implementing Dunnett's multiple comparison test. For gene expressions, comparisons between two sets of data were statistically evaluated by the Mann–Whitney test. Comparisons between multiple groups were evaluated by the Kruskal–Wallis test and then by implementing Dunn's post-test. The correlation between the age of patients and the volume fraction of metachromasia was determined using R square values calculated by a liner regression.
Results
Donors and sampling
From the total number of 64 patients participating in this study, we were able to obtain per donor on the average 3–4 explants per experimental group for culturing and/or for morphological, histomorphometrical, histochemical, or gene expression analyses, that is, 12–16 explants totally per donor. The total duration of time over which sampling was performed was 18 months.
Morphology, histochemistry, immunohistochemistry, morphometry
Negative control groups (absence of growth factors) did not show any signs of chondrogenic differentiation (Figs. 1 and 2). Those that had been exposed to BMP-2 alone or a combination of BMP-2/TGF-β1 underwent chondrogenic differentiation, as evidenced by the intense metachromatic staining of the extracellular, and manifested the widespread appearance of pericellular lacunae—a characteristic feature of differentiated chondrocytes that have been chemically fixed in aldehyde for light microscopy 30 (Fig. 1). The intensity of metachromasia increased as a function of culturing time. At each juncture, the distribution of metachromasia over the sections was inhomogeneous. Synovial explants that had been exposed to TGF-β1 alone manifested only weak metachromasia. Immunoreactivity for type-II collagen (Fig. 2) likewise reflected the findings described above for the metachromatic staining.
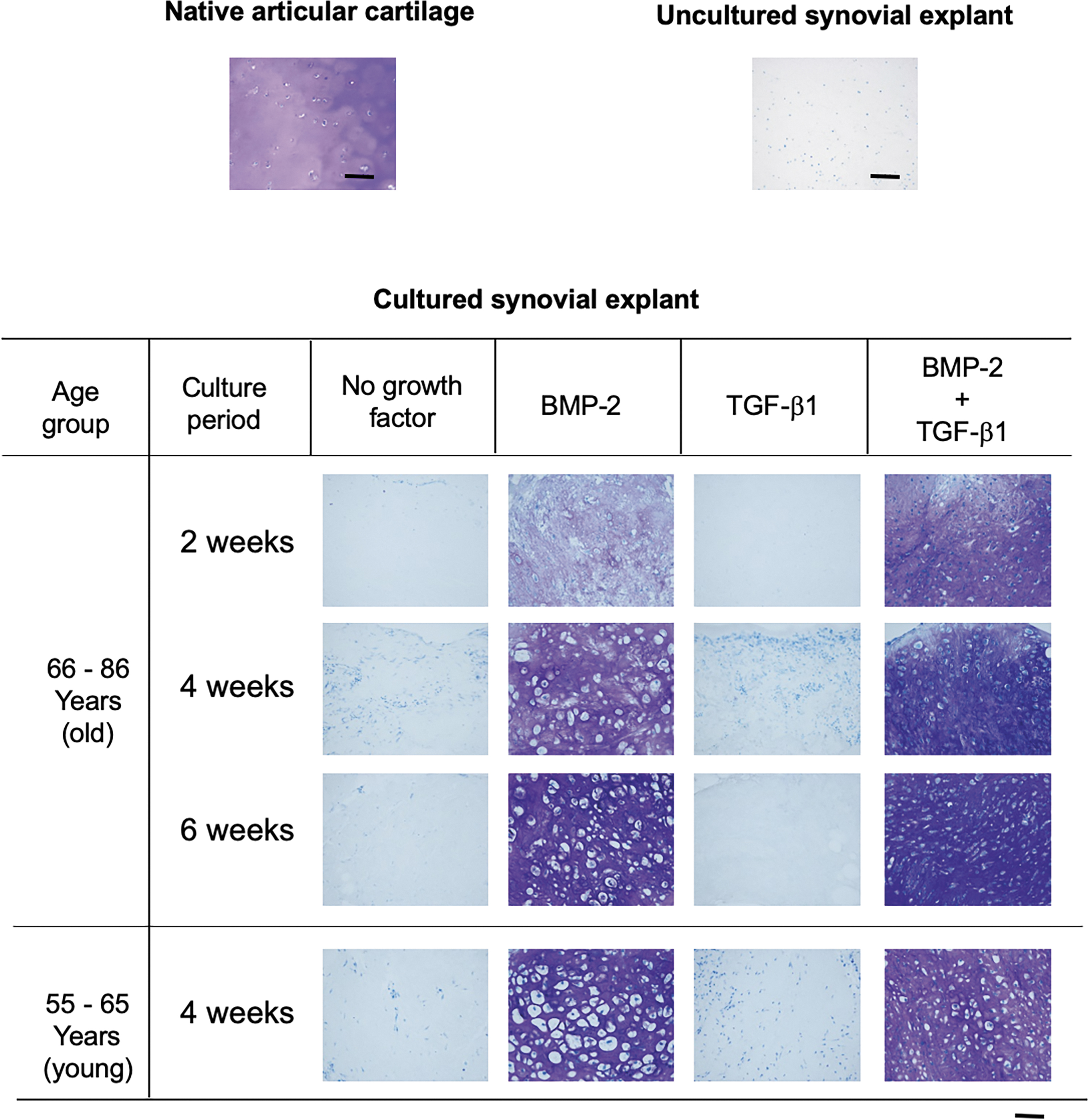
Light micrographs of sections through synovial explants in the two age categories of osteoarthritic patients, 2, 4, or 6 weeks after exposure to BMP-2 alone, TGF-β1 alone, or a combination of the two growth factors, and staining with Toluidine Blue O. Untreated synovial explants and native articular cartilage served as negative and positive controls, respectively. Synovial explants that had been stimulated with either BMP-2 alone or the BMP-2/TGF-β1 combination manifested a temporal increase in the intensity of metachromatic staining, whereas those that had been exposed to TGF-β1 alone exhibited only weak metachromasia at each juncture. The development of metachromasia was accompanied by the widespread appearance of pericellular lacunae, which are a characteristic feature of chondrocytes. Bar = 100 micrometers. BMP-2, bone morphogenetic protein-2; TGF-β1, transforming growth factor-β1.
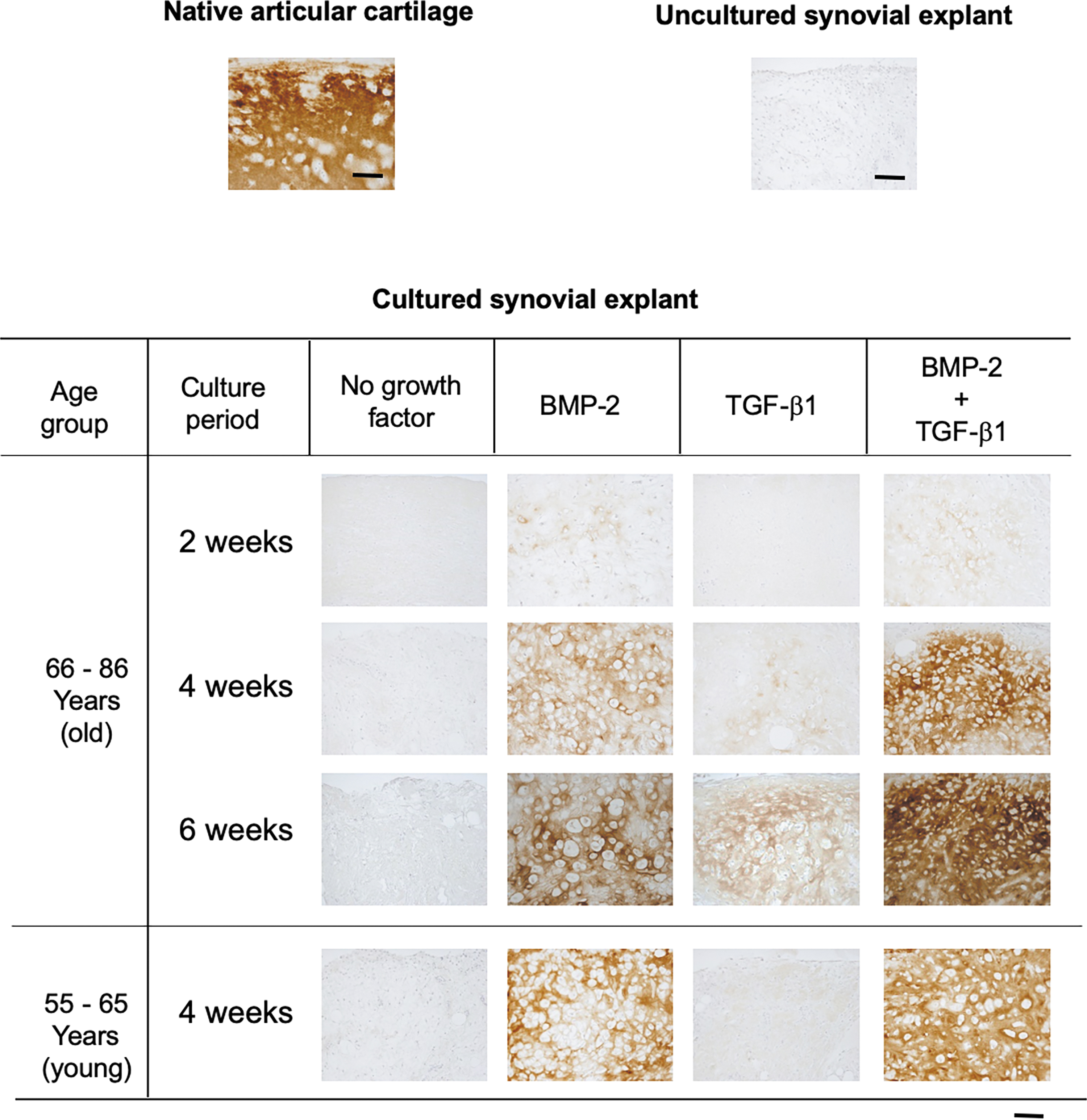
Light micrographs of sections through synovial explants in the two age categories of osteoarthritic patients, 2, 4, or 6 weeks after exposure to BMP-2 alone, TGF-β1, or a combination of the two growth factors, and immunostaining for type-II collagen. Untreated synovial explants and native articular cartilage served as negative and positive controls, respectively. Synovial explants that had been stimulated with either BMP-2 alone or the BMP-2/TGF-β1 combination manifested a temporal increase in the intensity of immunostaining for type-II collagen, whereas those that had been exposed to TGF-β1 exhibited only weak immunoreactivity at each juncture. Bar = 100 micrometers.
The histomorphometric quantification of the volume fraction of metachromasia (Fig. 3A) confirmed the time dependency of chondrogenesis induced by BMP-2 alone or by a combination of BMP-2/TGF-β1. At each time point, the volume fraction of metachromasia appeared greater after exposure to the BMP-2/TGF-β1 combinations than to BMP-2 alone. Although such a temporal increase in this parameter appears graphically apparent (Fig. 3A), a comparable level of metachromasia was attained between these two groups already after 4 weeks of exposure to the growth factors (p = 0.78 [comparison between the BMP-2 groups, 4 weeks vs. 6 weeks] and p = 0.6 [BMP-2/TGF-β groups, 4 weeks vs. 6 weeks]).
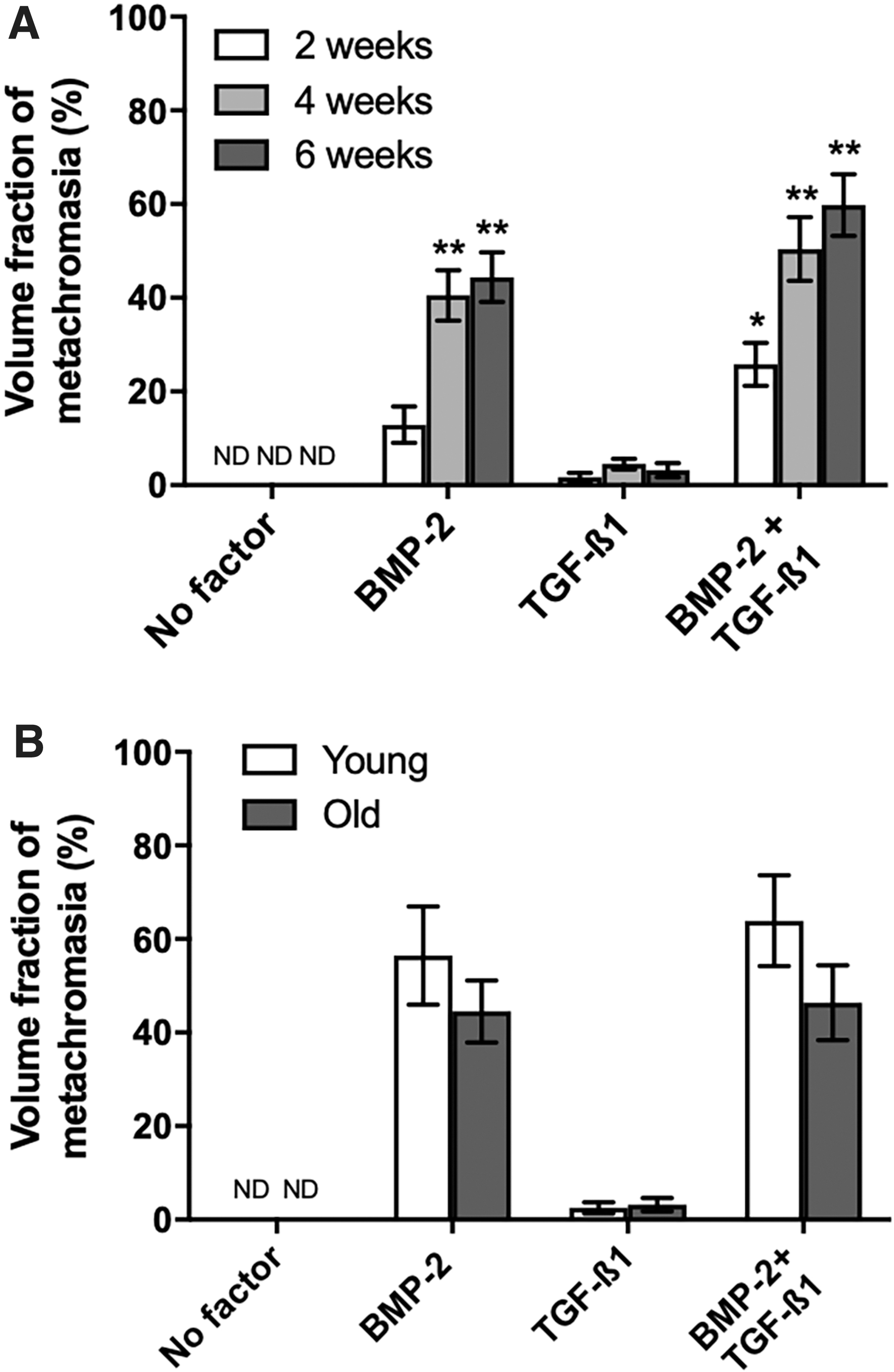
Moreover, the individual time point values show a high interindividual variability (coefficients of error (CE) ranging between 0.21 and 1.0). Figure 3B shows that the volume fractions of metachromasia were also on comparable levels when comparing the young and the old age groups with each other (p = 0.33 [BMP-2 groups, young vs. old] and p = 0.09 [BMP-2/TGF-β1groups, young vs. old]). In the TGF-β1-only group, the degree of metachromasia attained was clearly lower (e.g., in the young group: p = 0.0005 vs. BMP-2; in the old group: p = 0.002 vs. BMP-2).
The illustration of data distribution for all patients in Figure 4 shows an impressive scatter of results and confirms the age independency of the quantitative metachromasia. The correlation analyses for the BMP-2 patient materials and for the BMP-2/TGF-β1-treated groups showed a negative correlation between age and volume fraction (R 2 = 0.15 [for BMP-2 stimulation], 0.02 [for TGF-β1 stimulation], and 0.05 [for BMP-2/TGF-ß1 stimulation]) (Fig. 4). The graph also illustrates the high variability of results within each age group. The final cell volumes achieved after 6 weeks are illustrated in Figure 5. The mean final cell volume in the BMP-2 group was 15,452 μm3 (CE 1.03%, n = 754), and in the BMP-2/TGF-β1group was 1736 μm3 (CE 1.1%, n = 1064).
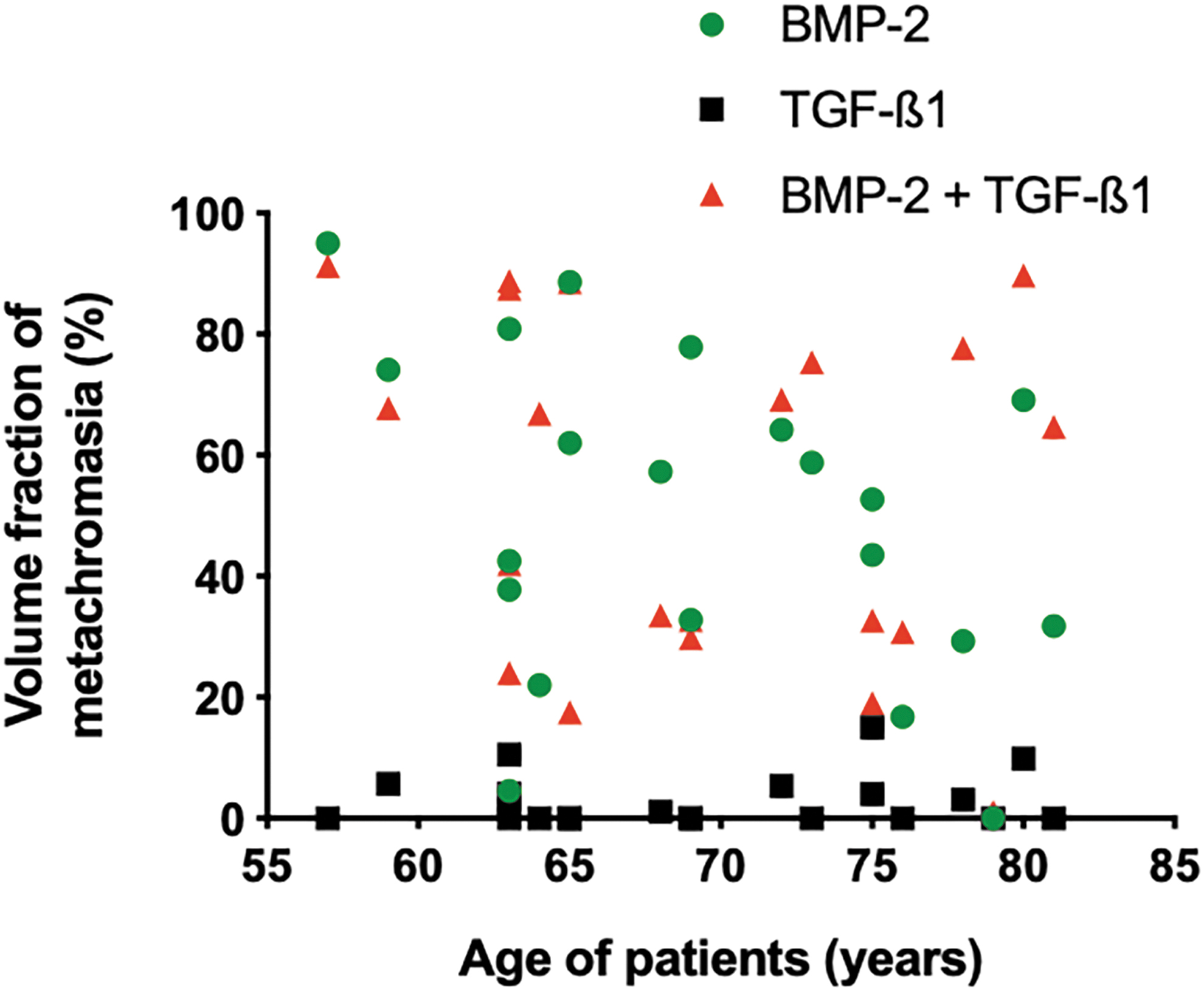
Volume fraction of metachromasia within the synovial explants of each of the osteoarthritic patients, 4 weeks after exposure to BM-2 alone, TGF-β1 alone, or a combination of the two growth factors, expressed as a function of the individual's age. A large data scatter can be identified, and no correlation with the age of the donors.

Mean chondrocyte cell volumes achieved at the end of 6-week culturing periods for the groups stimulated by BMP-2 alone or by the BMP-2/TGF-β1mixture. The final cell volumes achieved in these two groups (pooled data statistics) are significantly different from each other, ****p < 0.0001. The ones attained by the BMP-2/TGF-β1mixture correspond to the normal chondrocyte cell volume persisting throughout life in the adult human articular cartilage (radial zone), 62 whereas that of the BMP-2-generated chondrocyte volume relates in size to the terminal chondrocyte cell size achieved for the generation of maximal growth rate activities in growth plates for longitudinal bone growth. 63
Gene expression evaluation
The gene expression levels were monitored after the culturing periods of 2, 4, and 6 weeks. With the exception of matrilin-1, IL-4, and TNF-α, the mRNA levels that lay below the limits of detection, the activities of each of the investigated genes could be quantified (Fig. 6).

Temporal changes in the gene expression levels of the indicated anabolic and catabolic markers (to the left and to the right of the dividing vertical line, respectively) within the synovial explants of the group of older patients (66–86 years) after stimulation with BMP-2 alone
Among anabolic genes, collagen types II, X, and XI, aggrecan, and Sox9 were elevated after stimulation, irrespective of the nature of growth factors used (Fig. 6). The expression levels of these genes generally peaked at the 4-week juncture. COMP and lubricin were upregulated after stimulation with TGF-β1 alone or BMP-2/TGF-β1, but not with BMP-2 alone. Alkaline phosphatase was upregulated after stimulation with BMP-2 alone or BMP-2/TGF-β1, but not with TGF-β1 alone. Type-I collagen level was slightly raised only after stimulation with TGF-β1. Osteocalcin levels were not changed, irrespective of the type of growth factor applied.
The levels of catabolic genes were barely changed during culturing (Fig. 7). Only the levels of ADAMTS-4 (after stimulation with TGF-β1), COX-2 (after stimulation with BMP-2/TGF-β1), and iNOS (after stimulation with BMP-2 or BMP-2/TGF-β1) were slightly elevated (<10-fold) time dependently. On the contrary, the levels of IL-1 and IL-6 were downregulated, especially at the 2-week juncture after stimulation with BMP-2/TGF-β1.
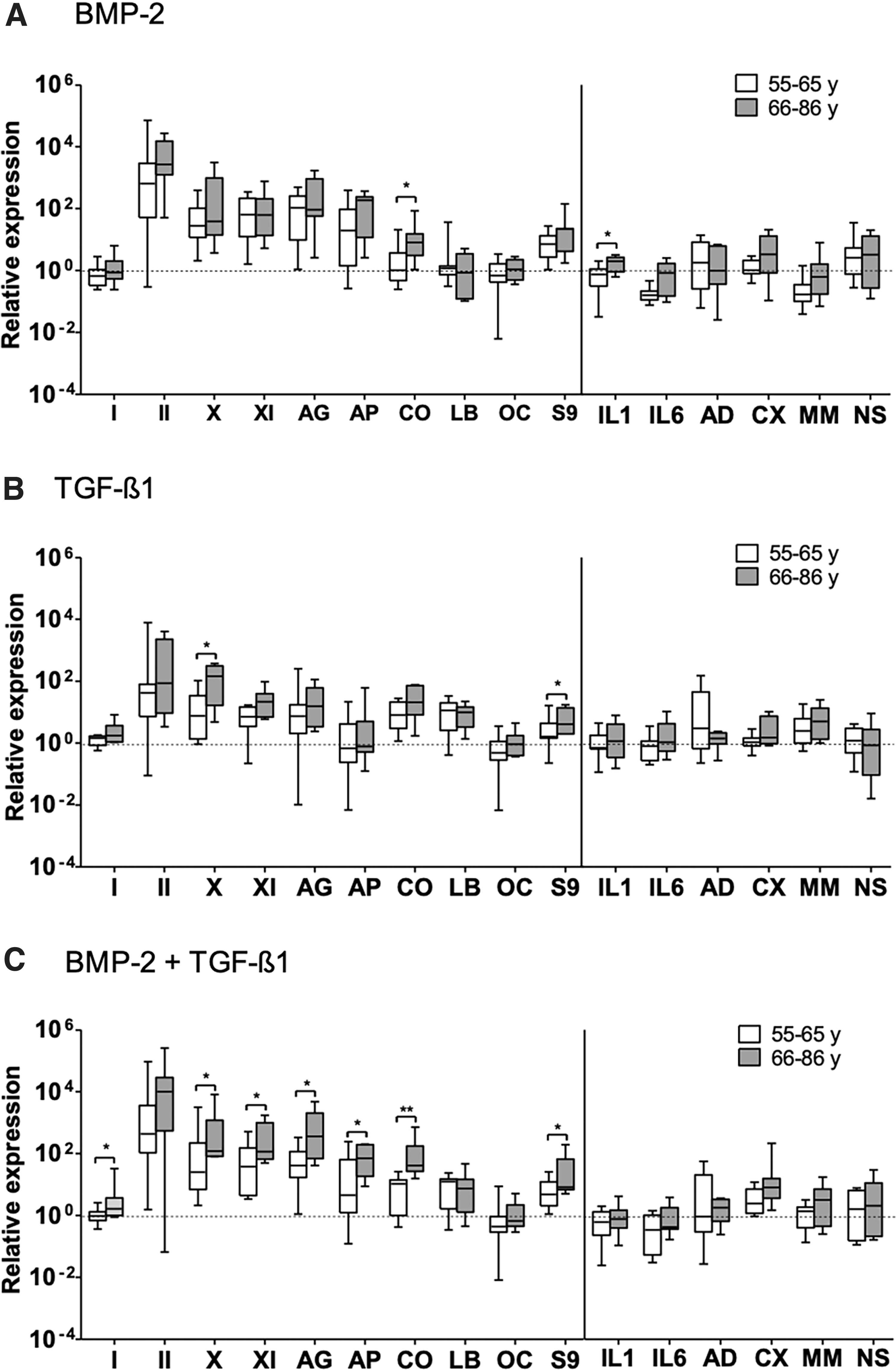
Comparison of changes in the gene expression levels of the indicated anabolic and catabolic markers (to the left and to the right of the dividing vertical line, respectively) within the synovial explants of the age groups of osteoarthritic patients, 4 weeks after stimulation with BMP-2 alone
The change of gene expression after stimulation was compared between younger and older patients at the 4-week juncture (Fig. 7); and due to limited availability of donor materials from the young age group, this comparison was not performed at the 6-week juncture. The differences between these two age groups were mainly observed among anabolic genes depending upon the stimulation condition. After stimulation with BMP-2, COMP was elevated significantly higher in older patients than in younger patients. After stimulation with TGF-β1, type-X collagen and sox9 were elevated significantly higher in older patients than in younger patients. After stimulation with BMP-2/TGF-β1, collagen types I, X, and XI, aggrecan, COMP, and sox9 were elevated significantly higher in older patients than in younger patients. Among catabolic genes, only IL-1β was significantly higher in older patients than in younger patients, but only after stimulation with BMP-2 alone.
Discussion
Compared with isolated MSCs, the most important advantage of synovial explants is that the physiological scaffold (ECM) is present around the cells and is provided by the synovial tissue itself. Such scaffold might be more conducive to chondrogenic differentiation than other nonjoint-associated scaffolds (or if ECM is absent). Indeed, the synovial tissue is known to differentiate into cartilaginous tissue under both clinicopathological 31 and experimental conditions,13,32 resulting in the formation of cartilaginous tumors and cartilage-bone-like tissues.
Furthermore, our previous studies have shown that bovine synovial explants are able to form more abundant cartilaginous matrix than isolated, alginate-cultured, and aggregate-cultured synovial MSCs.14,33,34 These findings suggest that a system using synovial explants, which can obviate the need for cell isolation and cell preculturing, could be one of the most promising strategies for the repair of articular cartilage lesions.
We had shown previously that the bovine synovial explants of healthy animals are able to differentiate into cartilage tissue after stimulation with appropriate growth factors.13,14,16 In the present study, we demonstrated that synovial explants derived from OA patients have a chondrogenic potential, and maintain this capacity irrespective of age. Given the age-independent high potential of human synovial tissue, it may thus be possible to exploit this potential for the repair of cartilage lesions in a clinical setting. 35
The activity and the differentiation potential of MSCs of various origins were found to decline with aging and with the number of passages in vitro.20,21 Consequently, the MSCs of elderly donors are considered to be unsuitable for the purposes of tissue engineering. 36 Also, the influence of donor age on the chondrogenic potential of MSCs derived from various tissue sources had been addressed by several research groups,37,38 and the data are conflicting. One possible reason for the discrepant findings might be that MSCs may not be considered a “universal” population subject to the same process of aging.
The aging of MSCs may be a tissue-specific phenomenon, or at least a process that is differentially influenced by their origin. Given the great potential value of MSCs in regenerative medicine, a clarification of this issue is important, particularly in the fields of orthopedics and rheumatology, 39 since patients who would qualify for the instigation of an autologous MSC-based cartilage repair strategy would be primarily patients with circumscribed articular cartilage defects, such as after trauma (may occur at any age), osteochondritis dissecans, a focal form of OA.
In a previous study, we demonstrated that synovial cells derived from the knee joints of osteoarthritic patients could be induced to differentiate into cartilage-producing chondrocytes (in vitro), 12 a finding that was reconfirmed by other researchers40,41 on similar grounds. However, the influence of donor age on the process of chondrogenic differentiation was not addressed, nor the issue of the generally shifted balance of cell metabolism to catabolic activities of chondrocytes in osteoarthritic joints.42,43 These phenomena most likely would affect repair cartilage formation from synovial tissues when originating from such diseased joints; to clarify these potentially adverse issues was the purpose of the present study, using synovial explants.
As evidenced by the formation of chondrocytes with lacunae and a metachromatic matrix, BMP-2 alone and the BMP-2/TGF-β1 combination induced the chondrogenic differentiation of the synovial explants to the greatest and to similar degrees. Thus, TGF-β1 in a combined use with BMP-2 exerted no synergistic/enhancing effect in the human synovial tissue (of OA joints) differentiation (unlike in synovial tissue of normal bovine joints of young adult animals 16 ). Given that the peak volume fraction of metachromasia was approximately ninefold lower after stimulation with TGF-β1 alone than after exposure to either BMP-2 alone or to the BMP-2/TGF-β1 combination, this is not a surprising finding, and it was, moreover, supported by the immunohistochemical staining profiles for type-II collagen.
Although temporal differences in the volume fraction of metachromasia were not significant, a time-dependent increase in this parameter was nonetheless graphically apparent (Fig. 3A), thereby indicating that the chondrogenic activity of the synovial MSCs could be sustained for at least 6 weeks in vitro. When the volume fractions of metachromasia were compared at the 4-week juncture, no differences were revealed. Also, when this histomorphometric parameter was displayed as a function of an individual's age, the correlation coefficients were very low. Thus, the general differentiation potential of synovial explants originating from human patients suffering from OA was not impaired as a function of donor age.
This finding confirms a previous report of de Bari et al. 10 using isolated human synovial cells. Moreover, the achieved degree of tissue transformation into cartilage-like tissue of the synovial explants was found to be of the same order of magnitude as that encountered in synovial tissue originating from healthy young adult bovine sources.13,14,16 Also, the chondrocyte sizes attained were of similar dimensions, but resulted in a more physiological size using the combination of BMP-2/TGF-β1 rather than BMP-2 alone. This finding also confirms the usefulness of the bovine tissue in an in vitro model to simulate human tissue biology (in the sense of the 3R philosophy 44 ).The osteoarthritic process thus seems not to affect the chondrogenic differentiation potential of the synovium.
The analysis of the gene expression levels of key anabolic markers of chondrogenesis permits a more discriminative evaluation of the induced differentiation process into cartilage tissue.
Type-I collagen is not a marker of chondrogenesis, but a characteristic component of fibrous tissues, and the presence of high levels of its mRNA in repair cartilage is an indication that it is deficient in hyaline-like qualities. Irrespective of the stimulation protocol, the gene expression levels of type-I collagen were extremely low and bordering on baseline values. Only after exposure to the BMP-2/TGF-β1 combination, a significant difference between the two age categories was revealed, with slightly higher levels in the group of older patients.
Type-II collagen is a characteristic component of cartilage tissues, and, irrespective of the stimulation protocol, its gene expression levels were higher than for any other marker. However, no age-related differences were revealed. This is somewhat surprising since on the basis of data in the literature relating to bone marrow-derived MSCs and their declining proliferation and differentiation potential with increasing age,20,36 as well as with perichondrial-derived cells 45 or with periosteum tissue flaps, 38 this apparently does not apply for synovial tissue, as found here, nor for isolated synovial-derived MSCs, as previously found. 10
Quantification of the immunohistochemical data was not performed since this method is intrinsically associated with such a high variance between blocks and sections that it does not provide sufficient solid ground (only intrinsic control referencing principles for each case could provide some semiquantitative indications).
Type-X collagen is a marker of terminal chondrocyte hypertrophy, and high mRNA levels indicate that the ECM is undergoing calcification, which is undesirable. Irrespective of the stimulation protocol, the gene expression levels of type-X collagen were elevated, but they were still lower than those of type-II collagen. Since TGF-β1 was found to suppress the terminal hypertrophic differentiation of chondrocytes, 16 this finding is not surprising.
Significant age-related differences in the gene expression levels of type-X collagen were revealed after stimulation with either TGF-β1 alone or the BMP-2/TGF-β1 combination, the values being higher in the group of older osteoarthritic patients than in the group of younger ones. This finding is in correlation to findings relating to normal articular chondrocytes and osteoarthritic ones from OA patients46,47 and with data from animal models 48 ; interestingly, it was also observed upon differentiation of synovial tissue from osteoarthritic patients.
Alkaline phosphatase is likewise a marker of terminal chondrocyte hypertrophy and matrix mineralization. 49 In both age categories of patients, its gene expression levels were lowest (barely above baseline values) after stimulation with TGF-β1 alone, and lower after exposure to the BMP-2/TGF-β1 combination than after treatment with BMP-2 alone.
Type-XI collagen is a marker of cartilage-specific collagen fibril formation since coexpressed with collagen type-II. 50 The gene expression levels of type-XI collagen were highest after exposure to either BMP-2 alone or the BMP-2/TGF-β1 combination. In the latter case, a significant age-related difference was revealed, the values being higher in the group of older osteoarthritic patients than in the group of younger ones.
The gene expression levels of aggrecan were higher after stimulation with either BMP-2 alone or the BMP-2/TGF-β1 combination than after exposure to TGF-β1 alone. These data are in accordance with the histomorphometric findings for metachromasia. An age-related difference was revealed only after stimulation with the BMP-2/TGF-β1 combination, the values being higher in the group of older osteoarthritic patients than in the group of younger ones.
The gene expression levels of COMP 51 and lubricin,52–54 which are markers of the articular cartilage layer as a whole and of its superficial zone and of synovial fluid, were highest after stimulation with either TGF-β1 alone or the BMP-2/TGF-β1 combination, and lowest after exposure to BMP-2 alone. Significant age-related differences in the gene expression levels of COMP were revealed after stimulation with either BMP-2 alone or the BMP-2/TGF-β1 combination, the values being higher in the group of older osteoarthritic patients than in the group of younger ones 55 ; this possibly may be related to TGF-β1effects. 56 No significant age-related differences in the gene expression levels of lubricin were observed.
The gene expression levels of matrilin-1, which is a marker of cartilage tissue organization, and that may play a role in the development of OA, 57 lay below the limits of detection in all instances. Hence, in the newly formed cartilaginous tissue, anabolic processes override catabolic activity. 58
Sox9 is a transcription factor that acts as a DNA-binding protein during chondrogenic differentiation. Irrespective of the stimulation protocol used, the gene expression levels of Sox-9 peaked—as expected—at the 4-week juncture. 59 The same temporal pattern of gene expression was observed for type-X collagen, aggrecan, and COMP, which is in accordance with the current opinion that their induction occurs via Sox9-regulated pathways. 60 Significant age-related differences in the gene expression levels of Sox9 were revealed after stimulation with either TGF-β1 or the BMP-2/TGF-β1 combination, the values being higher in the group of older osteoarthritic patients than in the group of younger ones.
The gene activities for the catabolic markers, 61 which included inflammatory factors62,63 (IL-1β, IL-4 [not detected], IL-6, TNF-α [not detected], COX-2, and iNOS) and matrix proteases (MMP-13, ADAMTS-4), were generally expressed at very low levels. As for the anabolic markers, the peak levels were generally attained at the 4-week juncture. No consistent trend in favor of anyone particular stimulation protocol was observed, with the exception of IL-1β. Also, no significant age-related differences in the gene expression levels of the catabolic markers were observed. In the case of IL-1β, the values were slightly higher in the group of older osteoarthritic patients.
The finding that the eight catabolic marker genes were expressed at very low or even nondetectable (IL-4 and TNF-α) levels indicates that the production of a cartilage matrix (attested by the stable histochemical, immunohistochemical, and morphological results) was positively balanced against degradative processes. 62 The finding that five of the six detected catabolic marker genes were expressed at similar levels in the older and the younger individuals indicates that the state of positive equilibrium between anabolic and catabolic processes was not compromised by aging.
The authors are fully aware that the gene expression activities are an indication only of possible posttranslational activities that need to be substantiated by demonstrating the presence/absence of such activities for each gene investigated. 64 Such additional investigations can now be performed on a more targeted level with this information now provided. Given the observed stability of the neoformed cartilage tissue structure, the outcome of an extensive posttranslational investigation can be expected to be most likely of a positive correlation to the observed gene activities.
Ethical Approval and Consent to Participate
Informed consent was obtained from the individual patients to donate their surgical waste material for the present study. An approval by the local ethics commission was also obtained.
Consent for Publication
All authors/coauthors agree with the text/content of this article and to submit it for publication in this Journal. The study sponsors had no influence on the study design, collection, analysis, and interpretation of data, nor on the writing of the article or the decision where to submit the article for publication.
Availability of Supportive Data
All data and supportive data of this study are available on an unrestrictive basis.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by SNF grant: 320000–118205 (to M.J.B.K. and E.V.) and the Foundation of the Clinic for Osteoporosis, Inselspital, Bern, Switzerland (to E.B.H.).