
Abstract
In recent years, the mathematical and computational sciences have developed novel methodologies and insights that can aid in designing advanced bioreactors, microfluidic setups or organ-on-chip devices, in optimizing culture conditions, or predicting long-term behavior of engineered tissues in vivo. In this review, we introduce the concept of computational models and how they can be integrated in an interdisciplinary workflow for Tissue Engineering and Regenerative Medicine (TERM). We specifically aim this review of general concepts and examples at experimental scientists with little or no computational modeling experience. We also describe the contribution of computational models in understanding TERM processes and in advancing the TERM field by providing novel insights.
Impact Statement
Although in recent years the use of mathematical and computational sciences has increased in the Tissue Engineering and Regenerative Medicine (TERM) field, we believe that a further integration of experimental and computational approaches has a huge potential for advancing the field due to the ability of models to explain and predict experimental results and efficiently optimize TERM product and process designs. By providing an overview of existing computational models, how they have contributed to the field, as well as a future perspective, this review represents an important step to help realize TERM's ultimate goal: a cure instead of care.
Introduction
In recent years, the mathematical and computational sciences have developed novel methodologies and insights that can, or at least so they sometimes claim, revolutionize the field of tissue engineering. These approaches potentially help streamline culture conditions, can give useful insight into the collective behavior of cells or how to efficiently control gene expression using growth factors, and they help to predict long-term behavior of engineered tissues in vivo. However, to tissue engineers themselves the use of these approaches may not always be clear. In this article, we give our answers to questions on the role of mathematical and computational modeling that you may have as a tissue engineer. In doing so, we keep in mind the specific modeling challenges we face when modeling biological cells and tissues: living tissues continuously adapt through mechanisms and actions that have not yet been elucidated in quantitative detail, in contrast to simulation techniques in the physical or the chemical sciences.
Despite this lack of quantitative precision in models of biological tissues, starting early with a simple model and adding more details as the computational analysis proceeds and new experimental knowledge becomes available, often proves very fruitful. In the following sections we do not aim to provide an exhaustive literature overview. Instead, we put forward some illustrative examples to provide an easy-to-grasp overview for noncomputational experts. Where applicable, we highlight excellent reviews that can aid in further reading. In this way, we aim to give an overview of the steps to be taken when integrating computational modeling in the workflow for Tissue Engineering and Regenerative Medicine (TERM). We cover the general concepts, available modeling techniques, and discuss the decisions that need to be taken when designing a computational model, ideally in discussion with a computational collaborator.
What Are the Current Challenges in TERM That Computational Approaches May Help Solve?
There is a great clinical need for tissue and organs for transplantation and research purposes. TERM aim to target this organ and tissue shortage by engineering different components that can be combined to generate much-desired organs or neotissues (Fig. 1). In the past decades, advancements have been made in many aspects of tissue engineering, including in material sciences, cell and developmental biology, nanotechnology, culturing of the necessary cell types from induced pluripotent stem cells, and systems biology.1–5 However, challenges in the field of TERM remain and include, among others, the recreation of organs with complex higher order structures and correct physiological function, recreating adequate vascularization (in particular for large tissues and organs), understanding and controlling the immune response after neotissue transplantation or during neotissue development in case of in situ tissue engineering, the reproducibility of neotissues, and so forth.2,6,7
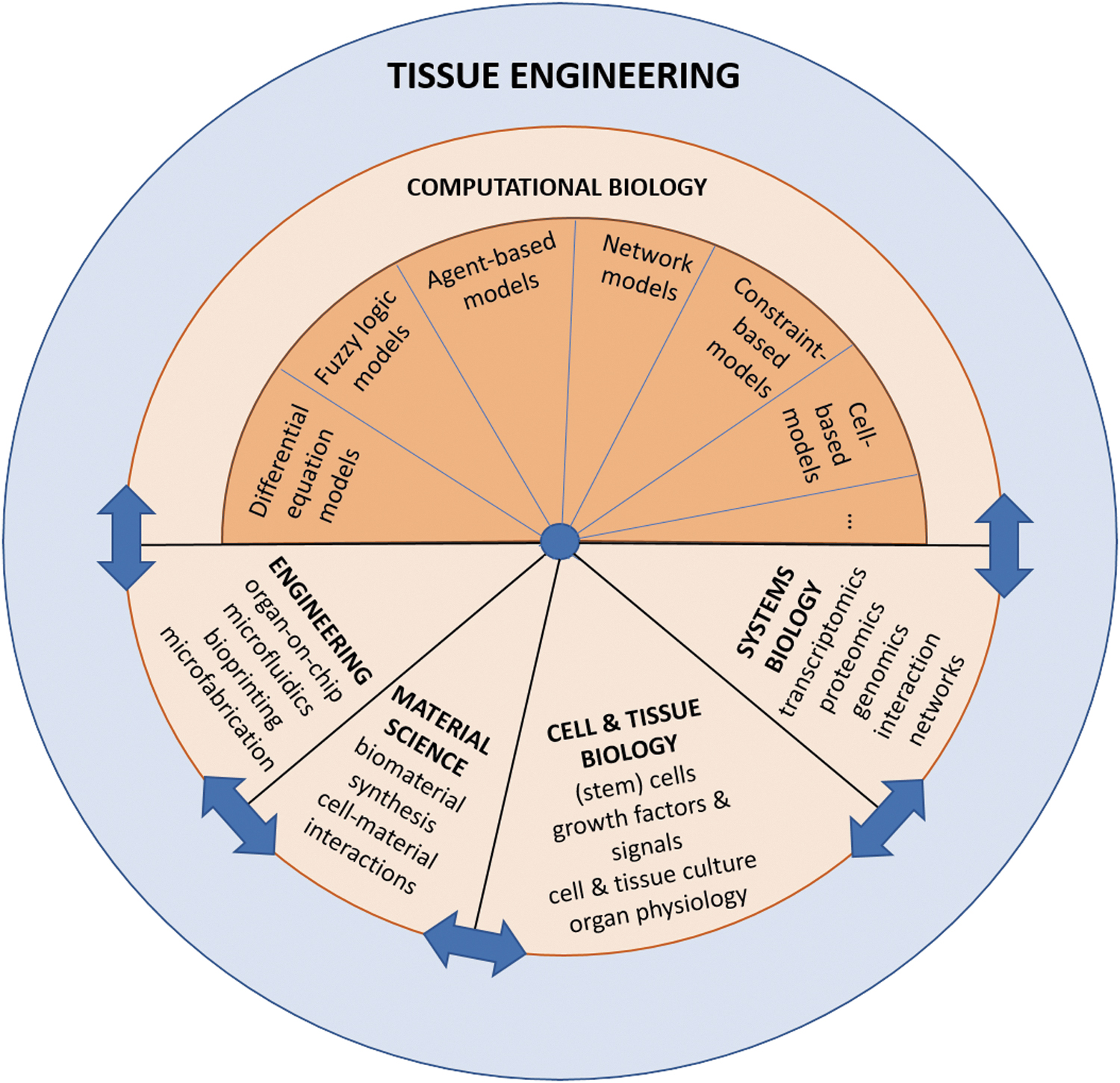
Tissue engineering is a multidisciplinary research field that aims to make complex tissues from a variety of cell sources to find personalized treatments. The increase in omics and high-throughput measurements has led to a steep increase in data that can be analyzed by systems biology. In addition, these data can be integrated and understood by the generation of executable computational models, which in turn help to prioritize data and predict novel mechanisms, thereby aiding the tissue engineer. Please note that the sizes of the pie do not reflect the contribution of the discipline to the field. Since this article aims to discuss computational biology, there is more detail about that in the figure.
Another important challenge in TERM is the huge amount of data available from literature. Systems biology aims to integrate these data into knowledge of the complexities of tissue and organ response. 8 To do so, systems biology makes use of omics approaches to collect data at the molecular and cellular levels. These approaches connect the individual components that make up the whole system. Modeling approaches attempt to put together the individual components back into functioning mechanisms, to obtain knowledge of cells and tissues as a whole. Thus, rather than focusing on single components, systems biology focuses on the integration of the components into a functioning whole. In particular, mathematical models can aid in understanding these systems and their underlying mechanisms by integrating the data into dynamic models that describe the processes.
In this review, we describe the contribution of mathematical models in understanding TERM processes and in advancing the TERM field by providing novel insights. We omit discussion of statistical models as used in bioinformatics-based systems biology, which are extensively discussed in Waters et al. 5 In the remainder of this article, we will use the term “mathematical model” to underline the mathematical aspect, but we acknowledge that very often these models are solved and simulated using computational tools.
What Is a Mathematical Model?
In general, models are simplified representations of complex processes or structures to help representing, analyzing, and understanding them. For example, signaling pathways are often represented as block-and-arrow diagrams: a conceptual visualization of a mechanism, where blocks indicate the proteins of interest, and arrows indicate the known or putative interactions. Blueprints are another example of models that aid in describing the design of a device (e.g., a bioreactor) in terms of dimensions and 3D relation of its components. In vitro and in vivo systems are also models. They have been designed to represent, simplify, and (partially) control the biology of the patient. Similarly, mathematical models represent a complex biological process in terms of equations or rules and a description of the domain (e.g., time and/or space) on which they are valid. 9
Conceptual versus mathematical models
To illustrate the use of mathematical models, imagine a cell proliferation experiment in which cells are seeded with an initial seeding density and grow until a particular end time (Fig. 2). As a research question, we aim to analyze the controlling factors of the proliferation rate, perhaps with the eventual aim of optimizing culture conditions. Without any direct observation of the process (Fig. 2a), except at the start and end time, only a very simple model can be defined, that is, a description in words: “the cell density increases due to cell proliferation.” However, when observing the system (Fig. 2b) or taking measurements at a sufficient temporal resolution (Fig. 2c), a more accurate model can be developed, that is, a verbally described hypothesis on the mechanisms underlying the observation. For example, we could state that “the cells proliferate until they have exhausted the growth medium.” Such a conceptual model can be rephrased in terms of a testable mathematical model, which predicts the dynamics of the system over time.
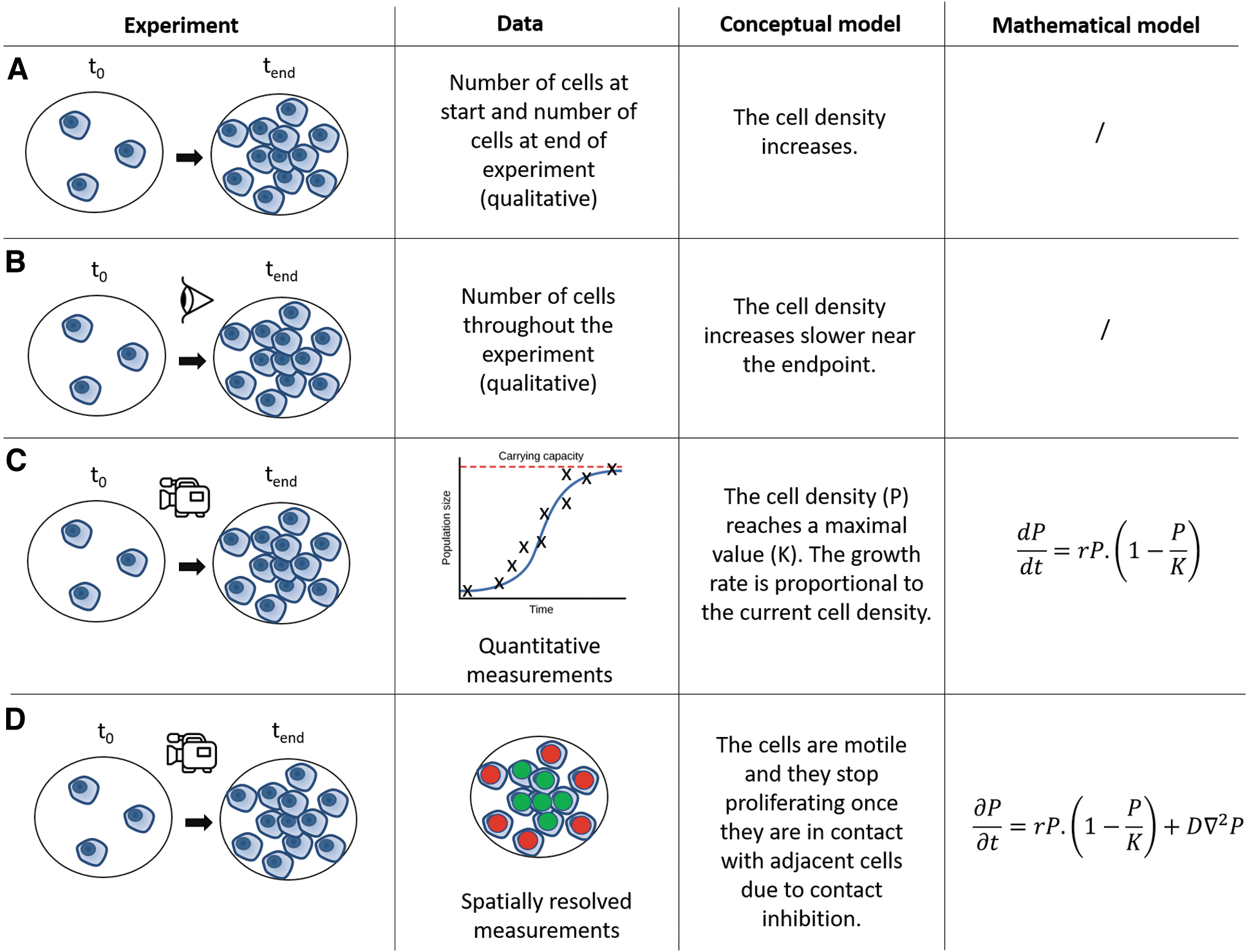
Schematic overview of the relation between experiments, data, interpretation, and models.
More specifically, in this example the mathematical formulation becomes an ordinary differential equation describing how the variable (the quantity of interest of the model, here, the cell density P) changes as a function of time has two contributions: (1) the cell density increases with a growth rate r proportional to the current cell density P, and (2) the growth rate decreases when the cell density reaches the maximal cell density K (1-P/K). This mathematical formulation is also known as the logistic growth model and defines how the cell density variable evolves over time; r, the growth rate, and K, the maximal cell density are the so-called parameters of the logistic model. Parameters are those quantities in the model that we assume do not change during an individual experiment. The (fixed) numerical values of the parameters can be measured in the experiment, and in this way, they connect the model to reality. Parameter values may differ between experiments due to different culture conditions or the use of a different cell type. Finally, we can refine the model, for example, based on discrepancies between the model predictions and the experimental observations.
Typical refinements include the inclusion of local differences in the growth rate in the cell culture, for example due to contact inhibition, or the effects of cell motility. A mathematical model describing such effects could take the form of a partial differential equation (Fig. 2d), which can describe the dynamics of system in time and in space, that is, over different locations in the culture dish. In this example, the mathematical model described how the variable (i.e., the cell density) changes in time at different locations of the culture dish. In addition to the contributions of cell growth and growth limitation, the model has a third contribution due to cell motility, in this study, described as diffusion. Other techniques to model a cell culture spatially include agent-based or cell-based models, in which cells are described individually, allowing for further contributions of cell shape and cell heterogeneity.10–13
An extensive description of how computational models can be constructed and which model types are available is beyond the scope of this article. For more details, we refer the reader to these excellent introductory articles14–16 and tutorials. 17 In this study, we would like to focus on the integration of computational modeling with (wet lab) experiments, an essential aspect to bring forward TERM research.
As mentioned above, computational models are simplified representations of complex processes and thus only imperfect approximations of reality. To quote George Box: “Essentially, all models are wrong, but some are useful.” 18 As such, the aim of computational modeling is not to build the most detailed model, but to learn about the biological system at hand through different cycles of model construction, calibration/fitting, prediction, and refinement alternated in an empirical cycle with (wet lab) experiments and data analysis5,19,20 (Fig. 3). First, if the model structure (e.g., the form of the equations such as the logistic growth function above) is already known, one may need to determine the parameter values (e.g., the growth rate, maximal density) to tune the model to the concrete (wet lab) conditions (model calibration). This can be done by setting the model parameters to experimental measurements and by estimating any remaining parameters of unknown value by fitting the model outputs to a set of experimental data (e.g., Scholma et al. 19 ).
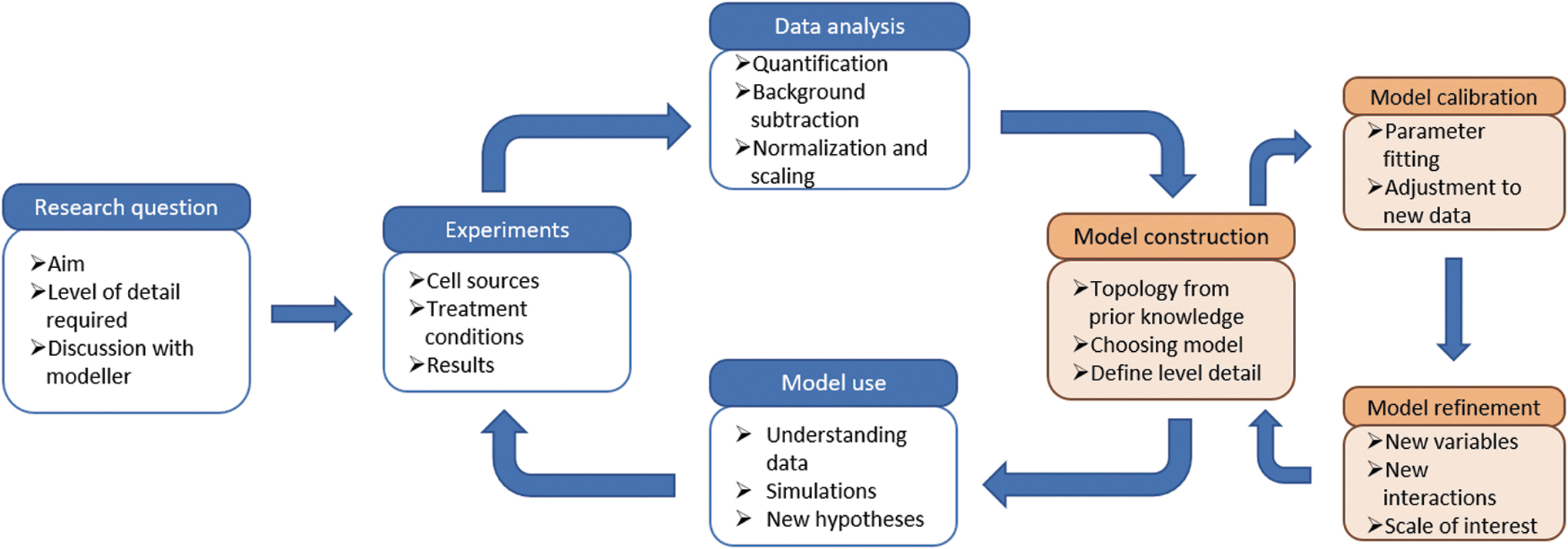
Workflow describing the integration of computational models in TERM. Starting from a research question, dedicated experiments and data provide input for a computational model. In an integrated, but partially separate empirical cycle, the model is optimized and validated, after which it can be used to interpret the data, do dynamical simulations, and generate new hypotheses that lead to new in vitro or in vivo experiments, and as such close the integrated feedback loop. Adapted from King et al. 17 TERM, Tissue Engineering and Regenerative Medicine.
The type of experimental data depends on the model and research question. If the aim is to predict protein concentrations as a function of time, then time series data are necessary to determine the rate values. If the aim is to understand heterogeneous growth in a bioreactor, then spatial data alongside molecular data will be important. Often, not enough data are available or of insufficient quality to determine the parameter set. Hereto various parameter estimation and optimization algorithms exist, see Refs. 21 and 22 for more details.
Second, the calibrated mathematical model and its analysis will lead to testable predictions that can be explored experimentally. From the example in Figure 2, we can conclude that “the time to reach the maximal density increases with the maximal density and the size of the culture dish, and decreases with the growth rate,” which could be experimentally explored by using cell lines with different growth rates and maximal densities. This process where the model outputs are systematically compared with independent wet lab observations to evaluate their quantitative and qualitative agreement with reality is called model validation. Model validation may be a huge challenge if the model outputs are not directly measurable through in vitro, in vivo, or ex vivo testing. For example, primary cells taken from cartilage quickly dedifferentiate when in culture23,24 and cells cannot be harvested at any time during development, especially in the case of research in human cells and tissues.
In addition, donor variability may greatly influence the outcome of the experiment.25,26 Researchers tackle these issues by using well-defined cell lines or primary cells used at an as-low-as-possible passage number.27–31 More often, researchers make use of literature data not used to generate the model.20,32–34 Novel data generation and literature resources are both appropriate for model validation, which is a crucial step to either determine the validity of the computational model or to identify missing mechanisms that need to be included before novel model predictions can be generated. 35 These novel model predictions can then be used to generate new testable hypotheses that can be prioritized before being tested in wet lab experiments (Fig. 3).
Scale of interest
A further consideration to make is the level of required detail. If we are interested, for example, in how culture media may help optimize the proliferation rate of cells, r, or how better design of the culture system can lead to higher cell densities, K, characterizing the proliferation of cells using the logistic equation may be just what we need. However, at some point we may find that culture medium A leads to a higher growth rate than culture medium B, and we may be interested in the mechanism underlying this difference. We may hypothesize that differences in growth factors or metabolite concentrations between the two culture media are responsible for this. A mathematical modeler could then construct a detailed model of the intracellular regulation networks or the metabolic networks,36–38 and attempt to systematically interfere with the nodes of the network to see which ones, if any, have an effect on the growth rate.
Once one of the nodes turns out to be a promising target, the modeler might choose to investigate it in more detail to identify potential ways to interfere with the effect pharmacologically, for example, by constructing a detailed model of a signal transduction network or by constructing receptor–ligand binding models at chemical detail level. Thus, the level of detail required for a suitable model is given by the question at hand. Typically, a researcher starts with a simple model and adds more detail as the analysis proceeds and more detailed questions need to be answered.
Multiscale modeling
In many cases, often the most interesting ones, after having considered the population level, the single cell level, and the molecular level of the system in isolation from one another, we may come to realize that the levels affect one another. In the hypothetical cell culturing system, a simple example would be contact inhibition. As the cells fill up the available space in the culture dish, they will touch one another and activate pathways such as the Hippo signaling pathway, mediated by E-cadherin 39 to reduce the proliferation rate. In a simple population-level model, it may suffice to capture the resulting local reduction in growth rate through the logistic growth term (Fig. 2d). A more detailed approach would model individual cells and introduce a simple model of the underlying signaling network inside each individual cell. Such approaches can give particularly useful insights into morphogenesis at different time scales.40–42
Bridging the gap between disciplines
For model construction, calibration, and validation, it is essential that both the experimentalist and modeler understand the biological question being asked and agree on the type of output needed to answer the question at hand and allow wet lab validation. When starting to model, the best way is to discuss with a computational modeling scientist to decide on the best modeling tool for your research question. This contact is preferably made before starting the experiments that will give you the first data to model, as the computational model may need alternate conditions, for example, time points, positive and negative controls, different concentrations of inputs, design variations, and so forth.
What Have Computational Models Contributed to TERM?
To date, computational models have been generated for addressing many (remaining) challenges in tissue engineering, including how cell fate choices are made,28,43–45 how cells interact to form a tissue,46–48 how the shape of the tissue dictates tissue function,49,50 how nutrients are supplied and waste products removed.32,51 In this section we will give an overview of how computational models have contributed to TERM. Since it is impossible to give an exhaustive review, we will focus on some notable examples from different tissues and at different scales, structured according to what the computational model has contributed to advancing the TERM field.
Computational models have increased our fundamental understanding of tissue development and remodeling
Dynamic models of signal transduction in which one can simulate processes as well as predict novel mechanisms play an important role in systems biology and in TERM. Signal transduction pathways allow cells to sense changes in their environment to which they can respond by changing gene transcription, metabolism, cell migration, proliferation, etc., ultimately changing cell function and/or cell fate. Therefore, the proper functioning of these signaling networks is crucial for adaptation and survival under a variety of conditions. The network regulating these processes is called a gene regulatory network and this can be captured in a dynamic computational model.
Traditionally, signal transduction was viewed as a linear process in which an input (growth factor, cytokine) was recognized by specific receptors on the cell surface that transduce the signal through an intracellular cascade (kinases, small molecular messengers) to an output element (transcription factor, cytoskeletal proteins, etc. 21,52). However, gene regulatory networks are characterized by cooperation between multiple pathways that integrate to regulate the many cell functions. To understand the influence of these complex and often nonlinear interactions, and simulate them together, computational models are very helpful tools.
Many small network models exist that describe one or two specific pathways in specific organisms,20,31,53 and others, as reviewed by Klipp and Liebermeister. 54 Recently, different models describing the gene regulatory network regulating cell fate decision making have been developed (e.g., Refs. 44,45). These mathematical models have contributed to a better understanding of cellular fate decisions and can be used to develop more effective in vitro stem cell differentiation protocols (reviewed in Spector and Grayson 55 ).
Besides unraveling signal transduction, computational models have also improved our understanding of, for example, cardiovascular tissues and their remodeling. Collagen fibers are the main load-bearing components of cardiovascular tissues. Establishing a functional collagen network is therefore of utmost importance for successfully tissue engineering cardiovascular tissues, such as blood vessels and heart valves. Computational models have substantially contributed to understanding the mechanisms of collagen remodeling in engineered cardiovascular tissues, because they allow for investigating different hypotheses and predicting their outcomes for complete tissues such as blood vessels and heart valves. Traditionally, there has been much focus on understanding and predicting how mechanical factors may drive collagen remodeling from a phenomenological point of view.
Specifically, by hypothesizing that collagen fibers align with or in between principal stress or strain directions, computational models have been able to predict the establishment of a native-like collagen architecture in blood vessels and heart valves.56–60 These results from computational models have nicely illustrated the importance of mechanical factors in regulating cardiovascular remodeling and provided some first tools to analyze how collagen remodeling can be steered in engineered cardiovascular tissues. Over the years, more biological phenomena have been added to computational models to further improve our understanding of cardiovascular development and adaptation.
One of the important phenomena that was recently added to computational models of mechanomediated vascular regeneration is the response of the immune system to synthetic scaffolds.61,62 In a recent study, Drews et al. demonstrated that a computational model incorporating both immuno-driven and mechano-mediated tissue formation was first of all able to predict early stenosis development in tissue-engineered vascular grafts as seen in a clinical trial, and, second, also able to predict the spontaneous reversal of stenosis over time. 63 This unexpected model-generated prediction was successfully verified in a preclinical study, 63 showing how computational models can contribute to the generation of new hypotheses and design of future experiments in the field of cardiovascular tissue engineering.
Another relevant aspect that is increasingly added to computational models is the impact of the cell behavior on tissue development and remodeling. 64 Cytoskeletal remodeling and contractility have been modeled in the context of TERM to understand cell (re)alignment, tissue compaction, and collagen remodeling due to cellular forces in engineered tissues as a function of the mechanical environment that the tissues are subjected to. For example, based on the known remodeling of the actin cytoskeleton in response to mechanical stimuli, and the impact thereof on collagen remodeling and contraction, computational models were able to capture the resulting cell and tissue organization in in vitro environments.65–69 Loerakker et al. subsequently applied these models to predict potential differences in the remodeling of tissue-engineered heart valves in the pulmonary and aortic position. 70
In the area of bone tissue engineering, computational models have focused on capturing the impact of the cell behavior on bone development,71,72 fracture healing,73–78 and the adaptive bone remodeling response.79–82 Recent developments focus on including the inflammatory phase of fracture healing or the immune response after tissue construct implantation. 83
Cells can sense and modify the extracellular matrix while being simultaneously in contact with other cells. This mechanobiological interplay between cell–cell and cell–matrix interactions is complex and has an essential role in the formation and maintenance of healthy functioning tissues. Several computational models have focused on capturing parts of this process, including cell–matrix adhesion,84,85 cell–cell adhesion,86–88 single-cell migration,89–93 collective-cell migration,94–96 and vascular pattern formation due to mechanical cell–cell communication. 97 These mathematical models can also be used to understand and predict the (heterogeneity of) cellular infiltration of scaffolds used for tissue engineering. Zahedmanesh and Lally, for example, developed a multiscale computational model to understand the influence of scaffold compliance and loading regime on the growth of vascular smooth muscle cells. 98 By including the transport and consumption of oxygen and nutrients in their model, Soares and Sacks were able to reproduce and understand the influence of static and dynamic mechanical conditioning protocols on in vitro tissue formation. 99
Computational models have identified key regulatory proteins, cytokines, and mechanical stimuli
To enable efficient use of TERM strategies for cartilage and bone, for example, it is important to define factors regulating tissue development and maintenance, as well as factors associated with disease progression and tissue healing. Recently, network models have been used to describe both cartilage formation of the growth plate28,43,100 and the articular cartilage.44,45 Specifically, these models aimed to describe and predict the molecular mechanisms regulating cartilage hypertrophy. Cartilage hypertrophy is a prerequisite for bone length growth. 101 In cartilage tissue engineering, however, hypertrophy during in vitro chondrogenesis of mesenchymal stem cells is followed by formation of transient calcifying cartilage, 102 with lesser mechanical and articular cartilage-specific properties. 103 Schivo et al. used a combination of literature, conventional molecular biology (quantitative polymerase chain reaction), and biophysical methods (fluorescence recovery after photobleaching [FRAP]) to validate new model predictions. This has led to the development of a model describing the development of osteoarthritis and identification of the wingless-related integration site (WNT) antagonists DKK1 and FRZB in the prevention of IL1b-induced cartilage degeneration by modulating the effect of Il1b in human primary chondrocytes. 45
In addition to soluble factors, such as growth factors and cytokines, mechanical factors play an important role in tissue development and maintenance. The effect of mechanical changes, such as matrix composition, viscoelastic collagen stress, matrix stiffness, and excessive strains in cartilage have been investigated by computational (numerical) models and provide insight in the factors regulating tissue stiffness and loading in healthy and osteoarthritic joints.104,105
Computational models have estimated protein concentrations and mechanical parameters that are experimentally inaccessible
It is well known that mechanical stimulation can regulate cellular behavior, for example, differentiation, proliferation, matrix production, and mineralization and as such mechanical stimuli are often applied in tissue engineering experiments. For example, perfusion bioreactors are used to apply fluid-induced wall shear stress on cells, but it is difficult to measure the actual mechanical stimulation cells are exposed to during an experiment. Zhao et al. used multiscale computational dynamics to quantify the microfluidic environment within scaffolds with highly irregular pore geometries. 106 Similarly, Ellis et al. have used a continuum mathematical model to calculate the spatiotemporal glucose and lactate concentrations within a bioreactor for culturing organoids, information which is challenging to obtain experimentally. 107 Their analysis showed that the bioreactor can be divided in two regions, depending on whether the effect of nutrient replenishment from the inlet has been experienced, which can be used to inform bioreactor operating conditions, ultimately improving the quality, reproducibility, and number of bioreactor-expanded organoids.
Computational models have informed and optimized experimental designs (3Rs)
Computational models have been extensively used for the optimization of scaffold topology and architecture108–111 as well as bioreactor-based processes,106,107 reviewed extensively in Refs. 112–116 as these have been proposed to be key components for the engineering of clinically relevant tissue-engineered implants. Similarly, computational models are often used to optimize the design of TERM components and processes such as biomaterials117–119 and microfluidic devices.120,121 For example, to improve cell viability after implantation of tissue engineering constructs, Pedraza et al. developed and optimized hydrolytically activated oxygen-generating biomaterials. 122 More specifically, the oxygen gradients within cell-loaded agarose constructs with varying total cell loads, external oxygen tension, and the presence or absence of the oxygen-generating biomaterial (modeled with first-order reaction rate kinetics) were predicted with COMSOL Multiphysics. From these calculations, they could optimize the dose, geometry, and surface/volume ratio of the oxygen-generating biomaterial.
Li et al. identified optimal oxygen delivery condition for matrix production by human chondrocytes in scaffold-free pallet cultures. 123 Similarly, others have focused on optimizing the release of ions or growth factors from biodegradable scaffolds.124–126 For example, calcium phosphate materials are widely used for dental and orthopedic applications and their degradation products, Ca2+ and Pi ions, are believed to affect bone cell chemotaxis, proliferation, and differentiation. However, the ion release rate from these scaffolds depends on dissolution and diffusion, which in turn depend on the scaffold composition and geometry. Manhas et al. have developed a finite element method to investigate the local Ca2+ ion release from CaP-based scaffolds. 127 Combining this work with a computational model of Ca2+ influence on osteogenic cell behavior and bone formation, 128 allowed for CaP scaffold optimization. 129
Besides optimizing tissue engineering constructs for oxygen transport and release rate, 130 others have optimized the macroscopic pore shapes of scaffolds to control the kinetics of tissue deposition, for example, bone formation. For example, Bidan et al. developed a model to predict the behavior of osteoblasts on curved surfaces. 131 They show that in cross-shaped pores the initial overall tissue deposition is twice as fast as in square-shaped pores, opening new avenues to improve the speed of bone ingrowth into porous scaffolds. The computational model of bone regeneration developed by Carlier et al. has shown that particular cell patterns in bone tissue engineering constructs are able to enhance bone regeneration compared with uniform cell distributions. 132 The model could be used to optimize cell gradient patterns for bioprinting.
Next to optimizing the cell gradient pattern, others have focused on understanding how mechanical loads are distributed throughout polymer fiber scaffolds. For example, Chen et al. used a homogenization theory to derive the bulk mechanical properties of a cylindrical construct of the fiber composite material for a range of fiber spacings and determined the local mechanical environment experienced by the embedded cells. 133
Others have used computational models to determine the optimal medium refreshment medium and initial growth factor concentration, where the goal is to maximize cell growth while minimizing the total experimental cost. 134 They show that multiple optimal operating points exist, depending on the desired cell number and costs. However, when calibrated for a particular cell type of interest, this in silico framework can calculate the optimal operating conditions, contributing to a cost-efficient production of cell-based TE products.
In the context of tissue-engineered heart valves, computational analyses 135 revealed that the geometrical design of these valves may play an essential role in the progressive development of valvular insufficiency due to cell traction-mediated leaflet retraction, a phenomenon that has often been observed in preclinical studies.136–138 The computational model demonstrated that the in vivo strains in tissue-engineered heart valves with a commonly used geometry are in fact facilitating leaflet retraction and predicted that the use of a more curved geometry would allow for obtaining superior strain profiles with respect to preventing leaflet retraction. 135 The performance of tissue-engineered heart valves with the superior geometry as predicted by the computational model was subsequently tested in a preclinical study, and showed, for the first time, clinical-grade performance of living, fully biological tissue-engineered heart valves during a full year of implantation. 49
What Can Models Contribute to the Future of TERM?
The examples mentioned above clearly illustrate how computational modeling has contributed to advancing the field of tissue engineering. Based on these encouraging results, we believe that modeling should be increasingly integrated with experimental research in the field, to improve the understanding of the relevant (biological) phenomena underlying tissue development and adaptation, predict experimental outcomes, and design future experiments.
One of the most important challenges in tissue engineering is to identify which combinations of scaffold parameters (and culture conditions in case of in vitro tissue engineering) will induce functional tissue development and subsequently long-term functionality and adaptation in response to the individual's needs. Given the countless scaffold parameters (e.g., geometry, stiffness, porosity, structure, material) that influence this complex process in combination with the costs and time associated with long-term in vitro and in vivo experiments, it is practically impossible to identify such optimal designs using an experimental approach alone. Computational models can tremendously contribute to overcoming this challenge as, after proper validation, they can predict experimental outcomes and thereby provide an efficient means to identify promising scaffold designs, particularly when integrated with formal optimization methods. 110
Obviously, such computationally identified designs should still be tested experimentally to verify their performance. Nevertheless, using computational models in the search for optimal scaffold designs will help to reduce the number of required experiments and avoid inefficient trial-and-error approaches. Finally, within the context of clinical translation, we expect that computational predictions of scaffold performance and tissue development, adaptation, and function will become highly relevant for obtaining regulatory approval. Computational simulations can complement the data from bench testing, animal studies, and clinical trials that are used to approve medical devices, 139 and offer the possibility to efficiently perform “in silico” clinical trials on virtual patients with diverse characteristics.140,141
New developments are required to enable broad implementation of integrated computational–experimental approaches in the tissue engineering field. First, integrating computational and experimental research requires multidisciplinary knowledge and/or collaboration of scientists with different expertise. This will only be successful when individual researchers are willing and able to cross the borders of their own discipline. In addition, when developing computational models, researchers from the modeling community should be (more) aware of the opportunities, uncertainties, and limitations of experimental approaches. This will make sure that models are properly connected to the (parts of) experiments they are supposed to cover and facilitate the adoption of computational models by researchers originally doing only experimental research. The performance of computational and experimental research within single research groups would provide the most ideal environment to foster integration of the two approaches. In addition, dedicated MSc or PhD programs, which train students in both computational and experimental approaches would foster such interdisciplinary integration.
In addition to computational advances, the TERM field has developed and adopted novel experimental techniques and technologies that can benefit the calibration and validation of future mathematical models. In particular, quantitative (microscopy) techniques allow characterizing the spatiotemporal dynamics of proteins, cells, and tissues, as well as the biomechanical properties thereof in physiology and pathology. 142 Initiatives are also being taken to achieve community consensus regarding the testing protocols and standards for the material characterization of biological tissues (e.g., https://c4bio.eu/). Considering the advent of new (micro)fabrication technologies and advanced (automated) analytic equipment, high-throughput screening platforms also provide an ideal strategy to analyze thousands of combinations of interactions among biomaterials and cells. 143
Similarly, various omics approaches are generating unprecedented amounts of data that characterize the structure, function, and dynamics of cells and tissues.142,144,145 Initially, a lot of omics data were generated without a clear common protocol, making integration of different data sets challenging. In recent years, standardization has been suggested to allow better integration and reuse of data sets. 146 In addition, these data sets have served the development of mathematical methods to understand and integrate these data. For example, tools such as DoRothEA and ANANSE are used to combine various omics data sets, including ChIP-seq, ATAC-seq, and RNA-seq analyses to predict key transcription factors for cell fate determination.147–152
Using these transcription factor networks one could infer the upstream signals (both chemical and mechanical) that regulate the activity of the identified transcription factors, which is important for controlling cell fate in TERM products. As such, machine learning and other data science methodologies are continuously being improved to deal with the large, incomplete data sets resulting from these high-throughput and omics methodologies. Importantly, data science methodologies alone ignore the fundamental laws of physics and can propose nonphysical solutions. 153 Therefore, exciting efforts are being undertaken to integrate data science methodologies with mathematical modeling approaches to create robust predictive models that integrate the underlying physical principles while being able to explore the massive design spaces that characterize the TERM field.153–156
As such, the development of high-throughput and more quantitative techniques results in more data as well as community standards, which will not only enable improved model calibration but also a more rigorous verification and validation of the computational models, ultimately leading to more model credibility. 35
In summary, the field of tissue engineering is a fascinating and complex multidisciplinary research field with many factors determining the ultimate performance of tissue-engineered constructs. For this to be successful, we submit that scientists from different disciplines should work together to understand and eventually control tissue development thereby ensuring long-term functionality. Within this scope, we highlight that computational models have huge potential for advancing the field, especially when integrated with experimental research, due to the ability of models to explain and predict experimental results and efficiently optimize scaffold designs. In this way, computational models may help realize TERM's ultimate goal: a cure instead of care.
Footnotes
Authors' Contributions
J.N.P., S.L., R.M.H.M., and A.C. contributed to the design and conception of the review, drafted the article, contributed to article revision, read, and approved the submitted version.
Acknowledgment
Disclosure Statement
No competing financial interests exist.
Funding Information
J.N.P. gratefully acknowledges the Dutch Arthritis Foundation grant no. 17-2-402, and the Dutch Research Council (NWO) grant no. OCENW.GROOT.2019.079. S.L. acknowledges funding from the European Research Council (ERC) under the European Union's Horizon 2020 Research and Innovation Program (ERC StG MechanoSignaling [grant agreement no. 802967]). R.M.H.M. is supported by Nederlandse Organisatie voor Wetenschappelijk Onderzoek grant NWO/ENW-VICI 865.17.004.