
Abstract
Volumetric muscle loss (VML) injuries represent a majority of military service member casualties and are common in civilian populations following blunt and/or penetrating traumas. Characterized as a skeletal muscle injury with permanent functional impairments, there is currently no standard for rehabilitation, leading to lifelong disability. Toward developing rehabilitative strategies, previous research demonstrates that the remaining muscle after a VML injury lacks similar levels of plasticity or adaptability as healthy, uninjured skeletal muscle. This may be due, in part, to impaired innervation and vascularization of the remaining muscle, as well as disrupted molecular signaling cascades commonly associated with muscle adaptation. The primary objective of this study was to assess the ability of four pharmacological agents with a strong record of modulating muscle contractile and metabolic function to improve functional deficits in a murine model of VML injury. Male C57BL/6 mice underwent a 15% multimuscle VML injury of the posterior hindlimb and were randomized into drug treatment groups (formoterol [FOR], 5-aminoimidazole-4-carboxamide riboside [AICAR], pioglitazone [PIO], or sildenafil [SIL]) or untreated VML group. At the end of 60 days, the injury model was first validated by comparison to age-matched injury-naive mice. Untreated VML mice had 22% less gastrocnemius muscle mass, 36% less peak-isometric torque, and 27% less maximal mitochondrial oxygen consumption rate compared to uninjured mice (p < 0.01). Experimental drug groups were, then, compared to VML untreated, and there was minimal evidence of efficacy for AICAR, PIO, or SIL in improving contractile and metabolic functional outcomes. However, FOR-treated VML mice had 18% greater peak isometric torque (p < 0.01) and permeabilized muscle fibers had 36% greater State III mitochondrial oxygen consumption rate (p < 0.01) compared to VML untreated mice, suggesting an overall improvement in muscle condition. There was minimal evidence that these benefits came from greater mitochondrial biogenesis and/or mitochondrial complex protein content, but could be due to greater enzyme activity levels for complex I and complex II. These findings suggest that FOR treatment is candidate to pair with a rehabilitative approach to maximize functional improvements in VML-injured muscle.
Impact statement
Volumetric muscle loss (VML) injuries result in deficiencies in strength and mobility, which have a severe impact on patient quality of life. Despite breakthroughs in tissue engineering, there are currently no treatments available that can restore function to the affected limb. Our data show that treatment of VML injuries with clinically available and FDA-approved formoterol (FOR), a beta-agonist, significantly improves strength and metabolism of VML-injured muscle. FOR is therefore a promising candidate for combined therapeutic approaches (i.e., regenerative rehabilitation) such as pairing FOR with structured rehabilitation or cell-seeded biomaterials as it may provide greater functional improvements than either strategy alone.
Introduction
Volumetric muscle loss (VML) injuries are characterized by the large-scale loss of tissue due to traumatic event or necessary surgical removal.1–4 The extent of this loss overwhelms the remaining muscle's ability to repair and regenerate, leading to a skeletal muscle dysfunction, limitations in mobility, and overall decline of patient quality of life. 5 As it stands, there exists no long-term support for skeletal muscle tissues after VML injuries due to a lack of clinical standards of care to address the damage or to systematically address the functional decline after trauma.4,6 Several studies in the regenerative medicine field have responded with the development of approaches that replace and/or stimulate the de novo growth of skeletal muscle tissue within the VML injury defect. However, these approaches have not been rigorously demonstrated to rescue function.7–11
An alternative approach to replacing or regenerating the lost muscle is to focus efforts on maximizing the quality and/or quantity of the remaining muscle after a VML injury. The remaining muscle typically represents a larger mass than that which could possibly be regenerated and, while limited, portions of the remaining muscle maintain partial innervation and vascularity.12,13 A significant hurdle to treatment is that the contractile properties and metabolic function (i.e., oxygen consumption and mitochondrial enzyme activities) of the remaining muscle are significantly impaired following VML injury.14–19 This decline in contractile and metabolic function is due to a combination of factors, including the loss of contractile units and replacement with fibrotic tissue, severe inflammatory response, and structural damage to the mitochondrial network.1,2,18,20–26 Therapeutic modalities to enhance muscle function after VML will need to address these underlying pathophysiologies to advance toward a clinical standard.
Metabolic function is a component of skeletal muscle physiology that is often overlooked in regenerative medicine therapy, but importantly, metabolism plays a critical role in meeting the energy demands associated with muscle repair and regeneration27–30 and single hindlimb VML injury influences whole body metabolism in mice, highlighting the significance of muscle metabolism on whole body homeostasis. 17 Previously, a forced overexpression of the transcription factor PGC-1α (peroxisome proliferator-activated receptor gamma, cofactor 1 alpha) partially rescued contractile and metabolic function of muscle tissue after a VML injury. 18 PGC-1α drives metabolic gene expression and this approach was a proof-of-concept demonstrating that addressing metabolic deficits in VML-injured muscle is worth further investigation as a therapeutic strategy.
However, virogenetic manipulation is not currently a feasible approach for a clinical setting. Thus, our goal was to evaluate alternative approaches to produce a similar functional rescue in remaining muscle metabolic function with clinical feasibility in mind. To this end, four pharmacological adjuvants (5-aminoimidazole-4-carboxamide riboside [AICAR], pioglitazone [PIO], sildenafil [SIL], and formoterol [FOR]), with previous history of improving muscle function, were tested herein.
We chose these four agents based on the likelihood that they would address the sustained deficiencies in strength and mitochondrial metabolic function that is characteristic of VML.14–19 AICAR is an adenosine monophosphate (AMP) analog capable of acting on the AMP-dependent protein kinase. Use of AICAR has been shown to prevent cachectic muscle wasting, 31 decrease inflammatory signaling in skeletal muscle models of diabetes, 32 and improve contractile properties in fukutin-deficient mice. 33 PIO is a peroxisome proliferator-activated receptor (PPAR) agonist that modulates insulin sensitivity34–36 and can ameliorate mitochondrial dysfunction caused by diseases like diabetes, metabolic syndrome, and polycystic ovary syndrome.34–39 SIL is a competitive phosphodiesterase-5 inhibitor that prevents the breakdown of cGMP by modulating nitric oxide, a known regulator of skeletal muscle function.
Use of SIL has been reported to improve mitochondrial metabolism, 40 improve Ca2+ storage and use in muscle fibers, 41 and reduce exercise-induced skeletal muscle damage in models of muscular dystrophy.42,43 FOR is a bronchodilator present in long-lasting inhalers that binds to β2-andrenergic receptors (β2AR), a type of G-protein coupled receptor that is abundant in skeletal muscle. It has been successfully used to improve the oxidative capacity of skeletal muscle, 44 muscle force production, 45 increase mitochondrial biogenesis and function,46–49 prevent muscle wasting,50,51 and promote muscle regeneration and hypertrophy.50,51 Taken together, there was sufficient evidence to believe these four treatments offered the most promising outcomes to be tested on VML injury.
Considering this breadth of evidence in improving skeletal muscle strength and metabolism in other disease models, we hypothesized that the administration of these drugs would lead to functional improvement in VML-injured muscle. In lieu of advancing the molecular signaling events associated with these pharmaceuticals, the primary objective here was to test the efficacy of these agents to improve contractile and metabolic function in the remaining muscle after VML injury.
Materials and Methods
Animals
Male C57BL/6 mice were housed at 20–23°C on a 12-h light/12-h dark cycle, with food and water provided ad libitum. At the time of randomization to experimental groups, all mice were between 10 and 11 weeks of age (presurgical mass 26.76 ± 1.75 g). All procedures were approved and performed in accordance with the guidelines and regulations of the Institutional Animal Care and Use Committee at the University of Georgia.
Experimental design
VML-injured mice underwent unilateral VML injury and then were randomized into one of the following experimental groups: AICAR (n = 9), PIO (n = 15), FOR (n = 15), SIL (n = 10), or no treatment (VML, n = 15). A cohort of injury-naive mice (n = 9) was included as uninjured controls and did not receive any pharmaceutical intervention. Uninjured mice were statistically compared to VML untreated to validate the injury model compared to previous studies in terms of muscle mass, strength, and metabolic function. Administration of pharmacological adjuvants for the experimental VML groups started 24 h postinjury and continued to 2 months. This duration of treatment was selected in the hope of surpassing the point of sustained inflammation, which has been observed up to 28 days following VML injury. 24
Drugs were incorporated directly into normal mouse chow (TestDiet, Richmond, IA) in the FOR (0.3 mg/kg/day), SIL (1 mg/kg/day), and PIO (20 mg/kg/day) groups, and animals were allowed to feed ad libitum. The AICAR group received daily subcutaneous injections (250 mg/kg aqueous solution) for the duration of the experiment. Doses were chosen after review of previous work on the effect of each of the drugs on muscle and key references are summarized in Supplementary Table S1.45,46,52–65 At the study endpoint, muscle mass, muscle strength, and metabolic function were assessed.
Surgical creation of VML injury
A dose of buprenorphine (1.2 mg/kg) was given preoperatively. A VML injury was performed unilaterally on the posterior compartment of anesthetized (isoflurane 1.5–2.0%) mice as previously described.14,18 Briefly, left hindlimbs were prepared by removing hair and the skin was aseptically prepared. An incision was made in the posterior of the limb to expose the muscle. Fascia and hamstrings were separated from the gastrocnemius muscle through blunt dissection. A small metal plate was inserted behind the gastrocnemius and soleus muscles and a 4-mm biopsy punch was used to remove a 23.9 ± 3.7 mg portion. Skin was re-approximated using 5–0 Vicryl sutures. buprenorphine (1.2 mg/kg) was given at 12 and 24 h, and meloxicam (2.0 mg/kg) was given at 24, 48, and 72 h postprocedure.
In vivo muscle strength
Peak isometric torque of the left hindlimb plantarflexor muscles (soleus/gastrocnemius/plantaris muscles) was assessed in vivo as previously described.66,67 Briefly, mice were anaesthetized using 1.5–3.0% isoflurane at an oxygen flowrate of 0.4 L/min. The left hindlimb was depilated and aseptically prepared before the peroneal nerve was severed. The foot was placed in a custom foot-plate attached to a servomotor (Model 300C-LR; Aurora Scientific, Aurora, Ontario, Canada), and platinum-iridium needle electrodes (Model E2–12; Grass Technologies, West Warwick, RI) were placed percutaneously on either side of the sciatic nerve to elicit isolated contraction of the hindlimb plantarflexors. Peak isometric torque was measured at stable body temperature (37°C) with the ankle joint maintained at 90° by a magnetic stabilizer clamp on the knee.
Peak isometric torque was defined as the greatest torque measured during a 250-ms stimulation using 1-ms square wave pulses at a frequency ranging from 150 to 200 Hz. Stimulation parameters were optimized between 0.6 and 3.0 mA (Model 701C; Aurora Scientific) and were determined by 0.1 mA step increases until an increase in amperage resulted in no further increase in torque produced. Fatigability of the plantarflexors was assessed with 90 submaximal contractions performed over 3 min using 250-ms stimulations at 60 Hz stimulation frequency. Following functional measurements, mice were euthanized by carbon dioxide inhalation and cervical dislocation and hindlimb muscles were processed for additional analyses.
Oxygen consumption of permeabilized muscles
High-resolution respirometry (Oroboros O2k) of permeabilized muscle fibers was used to assess mitochondrial oxygen consumption rates as previously described.18,68 Briefly, the medial portion of the gastrocnemius muscle, adjacent to the injury site, was dissected into fiber bundles and permeabilized with saponin as previously described. 27 Mitochondrial leak respiration was accomplished by the addition of glutamate (10 mM), malate (5 mM), and succinate (10 mM). State III respiration was accomplished by adding ADP (5 mM) after leak respiration had been recorded. After state III respiration was reached, cytochrome C (10 μM) was added to assess mitochondrial membrane quality to ensure no damage was obtained during the muscle fiber bundle dissection and permeabilization steps.
Any test where a cytochrome C rate exceeded a 10% increase over the state III respiration rate was deemed damaged and excluded from further analysis. Finally, carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (1 μM) was added to obtain an uncoupled respiration rate. Baseline rates (i.e., the rate before any substrates are added) were subtracted out from all other rates and rates were normalized to wet weight of muscle fiber bundles added to chamber and citrate synthase (CS) values to account for differences in mitochondrial content. 18
CS activity
Mitochondrial content was analyzed by CS enzyme activity as previously described. 18 Briefly, following muscle fiber collection for oxygen consumption experiments, the remainder of the gastrocnemius muscle was chopped into fine pieces and mixed. Approximately 40 mg of muscle was weighed and further homogenized in 33 mM phosphate buffer (pH 7.4) at a muscle to buffer ratio of 1:40 using a glass tissue grinder. Homogenate was incubated with 5,5′-dithio-bis(2-nitrobenzoic) acid (DTNB, 0.773 mM), acetyl CoA (0.116 mM), and oxaloacetate (0.441 mM) in 100 mM Tris buffer (pH 8.0). Activity of CS was monitored from the reduction of DTNB over time by measurement of absorbance at 412 nm.
Assessment of mitochondrial respiratory complex activity
Activity for each mitochondrial respiratory complex was measured using a protocol modified from Thome et al. 69 Complex I activity was measured in 50 mM potassium phosphate buffer, 3 mg/mL bovine serum albumin (BSA), 240 μM potassium cyanide, 0.4 μM antimycin A, 50 μM decyl-ubiquinone, and 80 μM 2,6-dichlorophenolindophenol (DCPIP). β-nicotinamide adenine dinucleotide (NADH) oxidation was measured through the reduction of DCPIP at 600 nm. Complex II activity was measured in buffer containing 10 mM potassium dihydrogen phosphate, 2 mM ethylenediaminetetraacetic acid (EDTA), and 1 mg/mL BSA at pH 7.8 and supplemented with 0.2 mM ATP, 10 mM succinate, and 0.08 mM DCPIP.
Following a 10-min incubation, at 37°C, the assay was initiated by the addition of oxidized decyl-ubiquinone (0.08 mM) and reduction of DCPIP followed at 600 nm. Succinate dehydrogenase (SDH) activity was measured as described previously.17,68,70 Muscle homogenate was sequentially incubated with 16.2 mM sodium succinate and 0.32 mM sodium cyanide before adding assay buffer containing 0.327 mM aluminum chloride, 0.327 mM calcium chloride, and 0.021 mM cytochrome c. SDH activity was monitored by the reduction of cytochrome c at 550 nm. β-hydroxyacyl CoA dehydrogenase (β-HAD) activity was measured by incubating muscle homogenate in a buffer containing 100 mM triethanolamine, 0.451 mM NADH, and 5 mM EDTA, as described previously.30,71,72 Acetoacetyl CoA (0.1 mM) was used to start the reaction. Activity of β-HAD was monitored from the oxidation of NADH over time as determined by its absorbance at 340 nm.
Quantitative real-time polymerase chain reaction
RNA was isolated from frozen tibialis anterior (TA) muscles from FOR-treated and untreated VML-injured muscles using an RNeasy kit (QIAGEN). cDNA was generated using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Universal SYBR Green Supermix (Bio-Rad) and the following sequence-specific primer were used to assess mRNA levels for the following: PGC-1α and PGC-1β were evaluated to assess mitochondrial biogenesis; COXIV, ND-1, ND6, and NDUFB6 were probed to assess the electron transport chain; and Fos was probed due to previous studies showing FOR treatment modulating its expression 73 (Supplementary Table S2). Expression was normalized to the expression of the 18S gene Relative gene expression was calculated using the 2-ΔΔCT method.
Statistical analysis
All data are represented as mean ± standard deviation. A two-tailed, unpaired t-test was used to validate the injury model (i.e., VML untreated) against uninjured. A one-way analysis of variance was used to determine differences between VML-untreated and VML-injured mice receiving a pharmacological agent with Dunnett's test for post-hoc analyses. Statistically significant differences are indicated when the p-value was ≤0.05, and statistical trends are noted if a p-value was between 0.05 and 0.10. All statistical analyses were performed on Prism statistical software (Version 8, GraphPad, San Diego, CA).
Results
Drug treatment effect on body mass and muscle quantity
VML-injured muscle suffers from a permanent deficit in muscle mass as well as impaired contractile and metabolic function compared to uninjured muscle.14,18 Herein, untreated VML mice had 22% less gastrocnemius muscle mass, 36% less peak isometric torque, and 27% less maximal oxygen consumption rate compared to uninjured mice (Fig. 1).
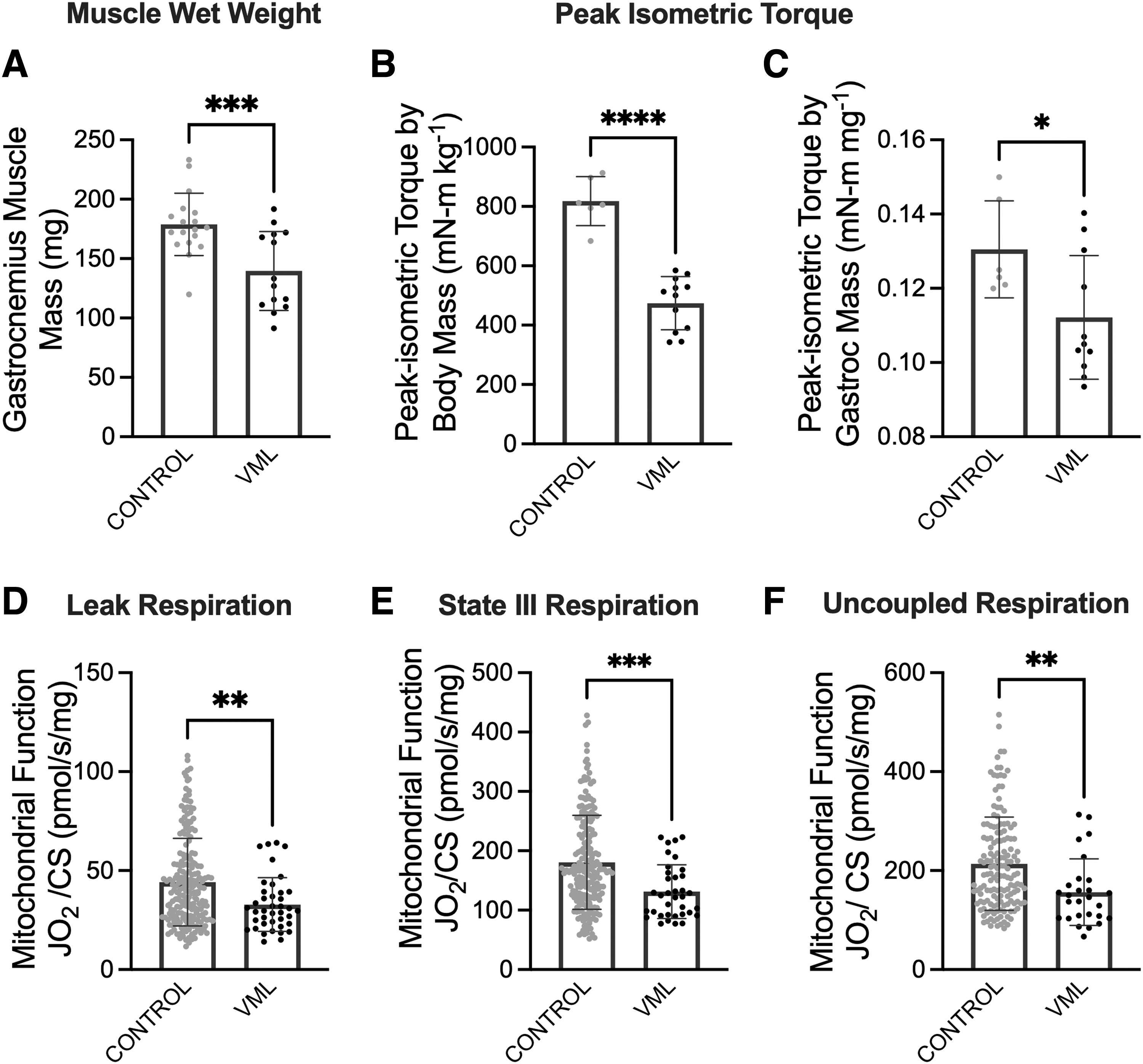
VML-injured tissue has a contractile and metabolic deficit compared to uninjured control limbs.
We first evaluated the effectiveness of our chosen treatments at increasing muscle mass 60 days following VML injury in comparison to a VML injury left untreated. FOR-treated mice had both 20% larger VML-injured gastrocnemius muscles (Dunnett's p = 0.0015) and a 23% higher gastrocnemius muscle to body mass ratio (Dunnett's p = 0.0012; Table 1) compared to untreated VML. SIL-treated mice had an 18% higher gastrocnemius muscle to body mass ratio (Dunnett's p = 0.0471). These data show that several candidate pharmacological agents resulted in muscle-specific mass changes. We next sought to determine the extent to which these changes affected VML-injured muscle function.
Mouse Body Mass and Gastrocnemius Muscle Mass
Values are mean ± SD.
Different from VML.
AIC, AICAR; ANOVA, analysis of variance; FOR, formoterol; OWA, one-way ANOVA; PIO, pioglitazone; SD, standard deviation; SIL, sildenafil; VML, volumetric muscle loss.
Strength, fatigue, and contractile properties following treatment
Contractile function was assessed by measuring peak isometric torque of the plantarflexors. Normalized to body mass (mN•m kg−1), FOR-treated mice produced ∼18% greater peak isometric torque compared with untreated VML mice (Dunnett's p = 0.0024, Fig. 2A). When normalized to gastrocnemius muscle mass (mN•m mg−1), however, there were no significant differences across treated and untreated VML-injured mice (Dunnett's p = 0.0526, Fig. 2B). Peak isometric torque tracings were also evaluated for changes in contractile properties with injury and treatment (Fig. 2C). Rate of contraction development was ∼29% greater in FOR-treated mice compared to that of untreated VML mice (Dunnett's p = 0.0157, Table 2).

Effect of pharmacological adjuvant administration on the contractile function of VML-injured limbs.
Contractile Properties
Values are mean ± SD.
Different from VML.
There were no differences across treated and untreated groups for all other contractile properties, including time-to-peak contraction, half-relaxation time, and submaximal torques at 40 and 60 Hz relative to peak (Table 2). These data suggest strength gains in FOR-treated mice are driven by greater muscle mass.
A contractile fatigue protocol was used to assess muscle fatigue. Percentage of torque loss was calculated by comparing the torque generated at the first contraction versus the final contraction (90th; Fig. 2D). Only AICAR-treated mice had less torque loss compared to untreated VML mice, suggesting less fatiguability (−48% vs. −61%, Dunnett's p = 0.0004; Fig. 2E).
Oxygen consumption of treated muscle fibers
Metabolic function was assessed by oxygen consumption rate of permeabilized gastrocnemius muscle fibers at leak, State III, and uncoupled respiratory states (Fig. 3A). Oxygen consumption rates are shown normalized to CS activity to account for mitochondrial content (Dunnett's p = 0.0347, Table 3). 74 The oxygen consumption rate during leak state respiration was ∼41% greater in FOR-treated mice than it was in untreated VML controls (Dunnett's p = 0.0007, Fig. 3B). Oxygen consumption rates during State III respiration were 36% greater in FOR-treated mice compared to untreated VML controls (Dunnett's p < 0.0001, Fig. 3C). Oxygen consumption rates during uncoupled respiration was 53% higher in FOR-treated mice compared to untreated VML controls (Dunnett's p = 0.0001, Fig. 3D). The addition of cytochrome C had no significant effect on oxygen consumption rates, supporting that permeabilized muscle fiber bundles were without significant damage (Fig. 3E).

Drug treatments improved oxidative capacity following VML injury.
Mitochondrial Enzyme Activity
Enzymatic activity normalized to citrate synthase activity as a measure of mitochondrial content. Enzymatic activity is represented as micromoles of product generated per minute per gram of protein. Values are mean ± SD.
Different from VML.
Mitochondrial enzyme analysis
Further analysis of metabolic function and muscle oxidative capacity was assessed by mitochondrial enzyme activity of the two primary electron transport chain complexes for electron carrier oxidation. Complex I activity for both AICAR- and FOR-treated mice was significantly greater than VML untreated (∼31.7% greater, Dunnett's p = 0.0193, Table 3). Complex II activity was 45% greater in FOR-treated mice compared to untreated VML (Dunnett's p = 0.0188, Table 3).
To supplement these findings, mitochondrial capacity to oxidize specific substrates, namely carbohydrates and fats, was evaluated by enzyme activity of SDH and β-HAD, respectively. SDH activity was 48.7% higher in SIL-treated mice than VML untreated (Dunnett's p = 0.0029, Table 3). β-HAD activity was significantly greater in FOR-treated mice than in untreated VML mice (29.0% greater, Dunnett's p = 0.0187, Table 3). Overall, oxygen consumption rates and mitochondrial enzyme assays suggest a modest effect of FOR treatment on skeletal muscle metabolic function.
Mitochondrial gene and protein expression
Based on the data collected throughout this study, it is apparent that the treatment of VML-injured muscle with FOR helped improve contractile and metabolic function. To determine whether these improvements are attributable to changes in the expression of mitochondrial metabolism-specific genes and/or proteins, TA muscles were analyzed from the VML-injured untreated and FOR-treated mice. The TA muscle was used due to lack of leftover biological material from the injured gastrocnemius muscle. However, we chose the TA muscle from the injured limb for its proximity to the injury site and because it is subject to the inflammatory process as the gastrocnemius muscle heals. FOR treatment did not change PGC-1α or c-Fos expression compared to VML untreated controls, and furthermore, PGC-1β and COX-IV expression were significantly lower (∼24% and 26% reduction, respectively) (Table 4).
Relative Gene Expression (mRNA/18S)
Values are mean ± SD.
Different from VML.
In contrast, mRNA content for ND-6 and NDUFB6, subunits of complex I (NADH:Ubiquinone Oxireductase chain protein 6 and Subunit B6) was 45% greater with FOR treatment compared to VML untreated. No change was observed in the expression of NADH dehydrogenase subunit 1 (ND-1) (Table 4). Finally, OXPHOS protein immunoblot data yielded no significant difference between FOR-treated and untreated VML mice at any individual respiratory complex (complexes I–V; Supplementary Data S1; Supplementary Fig. S1).
Discussion
There currently exists a therapeutic desert to address long-lasting skeletal muscle functional deficits after a VML injury. This is due, in part, to a vast majority of VML-related research focusing on regeneration of the lost tissue as opposed to functional restoration of the remaining muscle, and to a low percentage of published VML studies that include clinically relevant outcome measurements of muscle function. 11 To address this clinical need gap, we used functional outcomes of muscle strength and metabolic function to evaluate four FDA-approved pharmacological adjuvants' effects on the remaining muscle after a VML injury.
This study represents the first step toward identifying regenerative rehabilitation approaches, 75 those that combine a regenerative medicine approach that can modulate cellular physiology, such as those described herein, with physical rehabilitation to enhance skeletal muscle function after a VML. Our primary finding was that FOR had the most evidence for a beneficial effect, and there was minimal evidence for efficacy of AICAR, PIO, and SIL in a mouse model of VML. However, these functional adaptations did not represent any significant changes in gross anatomical structure (Supplementary Data S1; Supplementary Fig. S2).
The treatments evaluated throughout this study (i.e., AICAR, PIO, FOR, and SIL) have a track record of augmenting skeletal muscle strength in animal models of disease and injury.31,33,37,46,47,50,60,63 VML injury results in a permanent loss of muscle strength even when normalizing for changes in body mass and muscle mass, as shown in this study and previously reported.14–16,18 FOR's effect on VML-injured muscle strength was largely diminished when strength was normalized to gastrocnemius muscle mass, the primary plantarflexion muscle. Combined with the fact that FOR treatment was associated with larger remaining gastrocnemius muscle masses, this indicates FOR's benefit for VML-injured muscle strength in maintaining and/or enhancing the quantity of muscle (i.e., mass) as opposed to influencing its quality.
FOR may exert its effects on muscle quantity by increasing protein synthesis, decreasing protein degradation, and/or improving viability of remaining muscle fibers by enhancing neuromuscular junction stability.50,76,77 For example, skeletal muscles from young and old rats treated with FOR following a traumatic freeze injury had greater incorporation of [U- 14 C] phenylalanine (a marker of protein synthesis), and this was associated with greater muscle force production. 50 Also, FOR treatment in dystrophic mice that are characterized by progressive rounds of fiber damage and repair, was associated with fewer ubiquinated proteins, suggesting a decline in protein degradation. 76
VML is also characterized by an accumulation of collagen proteins (i.e., fibrosis), 78 and FOR in its vaporized form can influence the collagen protein turnover.79,80 A functional measurement associated with collagen protein within VML-injured muscle, passive stiffness about the ankle joint, 14 was not different between VML untreated and FOR-treated mice (Supplementary Data S1; Supplementary Fig. S3); however, we cannot rule out the possibility that fibrotic tissue changes within the VML-injured muscle (e.g., type of collagen and orientation) influenced the transmission of contractile force. 81
Another option, most intriguing in the context of VML, is FOR's potential signaling effects on cyclic AMP,82–84 which plays an important role in maintaining acetylcholine receptors at the neuromuscular junction.85,86 The remaining muscle fibers after a VML injury are characterized by greater denervation and polyinnervation indicative of neuromuscular junction instability.12,87 Furthermore, within the remaining muscle, there is a greater proportion of smaller, slow muscle fibers (i.e., less contractile force) that may implicate a preferential loss of larger, faster muscle fibers after injury.17,19 Further investigation into FOR's effects on remaining muscle innervation and fiber-type distribution will likely yield insight into the benefit of FOR treatment on muscle mass observed in this study.
Similar to contractile function, each evaluated drug—AICAR, 31 PIO,37,38,56 FOR,44,46 and SIL 88 —has previous evidence of improving mitochondrial respiration in skeletal muscle in a variety of disease models. As reported in this study and previously, VML injury results in a significant and lasting deficit in mitochondrial oxygen consumption rate.14,17,18 Lower oxygen consumption rates do not explicitly implicate a dysfunction, as bioenergetic factors related to ATP supply and demand, as well as the proportion of muscle fiber-types comprising the muscle can influence respiration. 89 However, previous research into the structure of the mitochondrial network following a VML injury shows disorganization near the injury site that stretches into the remaining muscle,18,90 and this lends support to an interpretation of mitochondrial dysfunction following VML.
The effectiveness of FOR to modulate mitochondrial function, content, and structure has been shown widely across tissues and disease/injury models such as diabetic renal failure, traumatic brain injury, and skeletal muscle after a spinal cord injury, ischemia-reperfusion, and cancer-induced cachexia.45,47,91–94 Herein, FOR treatment led to significant improvement in State III and maximal (uncoupled) respiration of permeabilized muscle fibers from the remaining muscle after a VML injury. We believed that FOR would activate signaling cascades that directly activated the transcription factor PGC-1α and maintained high transcription of this gene for the duration of treatment as this was supported by at least one study that utilized FOR after a spinal cord injury. 45
However, our data did not support this hypothesis. After further literature review, we saw that data concerning PGC-1α activation with FOR treatment was ambiguous.44–47 Importantly, literature has reported an increase in PGC-1α transcription in acute stages of treatment (∼8–24 h), which then wanes to basal levels.95–98 Therefore, FOR-induced mitochondrial biogenesis by PGC1α is either not supported as a mechanism of improvement in our VML model or its levels have stabilized at 2 months of treatment.
Another potential mechanism of FOR-induced improvements in mitochondrial respiration is related to electron transfer efficiency and mitochondrial complex protein content. Our results were mixed in this regard as we did detect greater gene expression for complex 1 subunits Nd6 (mtDNA) and Ndufb6 (nDNA), as well as greater complex I enzyme activity, but immunoblots for total complex I protein content showed no change. It remains a possibility that improvements in mitochondrial function with FOR are independent of biogenesis and protein content, and may reflect a greater capacity to deal with cellular stress. For example, in a model of traumatic brain injury, FOR treatment was associated with better mitochondrial-Ca2+ buffering, which may reflect an enhanced ability of the mitochondrial network to adapt to acute cellular stresses such as changes in calcium homeostasis. 92
Currently, the benefit of FOR treatment on mitochondrial respiration in VML remains unclear; however, toward the goal of validating pharmacological agents to combine with rehabilitation in future studies to address VML, FOR does have promising functional outcomes.
The improvements in functional capacity following FOR administration hold promise that VML-injured muscle is capable of metabolic adaptations, despite the adverse cellular environment following injury. Our data suggest that FOR treatment leads to the improvement of contractile and mitochondrial function by the maintenance of remaining muscle mass and improvement of overall oxidative capacity, possibly at complex I. Administration of physical rehabilitation following VML injury has been shown to improve contractile function.14,18 Therefore, further studies should investigate the integration of rehabilitative approaches with FOR treatment in the hopes of magnifying the improvements conferred by drug administration or rehabilitation alone. In conclusion, this work provides a thorough evaluation of FDA-approved adjuvants to improve muscle function after VML and names a therapeutic candidate that can be easily translated for in-clinic use.
Footnotes
Authors' Contributions
J.M.-F., S.M.G., and J.A.C. designed the experiments. J.M.-F., A.G.S., A.S.N., M.N.L., E.T.H., E.A.W., E.E.N., S.M.G., and J.A.C. performed the experiments, and collected and analyzed the data. All authors have read and approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
Funding support from the Department of Defense, through the Clinical and Rehabilitative Medicine Research Program, FY18 Neuromusculoskeletal Injuries Rehabilitation Research Award (W81XWH-18-1-0710 to S.M.G. [UMN], and J.A.C. [UGA]).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.