
Abstract
Tissue-engineered corneal epithelium transplantation is effective treatment for severe limbal stem cell deficiency (LSCD), while epithelial terminal differentiation, tans-differentiation, and insufficient stem cell during construction affect the quality of tissue-engineered corneal epithelium. In this study, we applied SB203580 in the culture medium to downregulate the p38 mitogen-activated protein kinase (MAPK) signaling pathway during construction of tissue-engineered corneal epithelium. With application of SB203580, tissue-engineered corneal epithelium showed enhanced strength and condensed structure. The expression of progenitor cell markers ATP-binding cassette sub-family G member 2, tumor protein p63, keratin 14, and Wnt family member 7A was increased, differentiation markers keratin 12, paired box 6, keratin 10, and keratin 13 and trans-differentiation markers actin alpha 2, smooth muscle and snail family transcriptional repressor 1 was decreased, while cell junction markers claudin 1 and cadherin 1 was increased in the tissue-engineered corneal epithelium. The Wnt/catenin beta 1 signaling pathway was upregulated in the epithelium after p38 MAPK inhibition. Transplantation of tissue-engineered corneal epithelium treated with SB203580 to rabbit LSCD model showed faster wound healing and improved epithelial quality. We conclude that downregulation of p38 MAPK signaling pathway helps maintain the stemness and prevent terminal differentiation and abnormal differentiation of corneal epithelial cells during epithelium construction process, and thus can improve the quality of tissue-engineered corneal epithelium.
Impact statement
Downregulation of p38 MAPK signaling pathway helps maintain the self-renewal of limbal stem cells and prevents terminal differentiation and abnormal differentiation of corneal epithelial cells. Small molecules modulating p38 MAPK signaling pathway ameliorate tissue-engineered corneal epithelium.
Introduction
The integrity and transparency of cornea play a critical role for normal vision. The outermost layer of cornea consists of a stratified nonkeratinized squamous epithelium that protects against fluid loss, microbial invasion, ultraviolet radiation, and physical or chemical trauma. 1 The homeostasis of corneal epithelium is contributed by limbal stem cells (LSCs), which are located at the limbus and represent the ultimate source of self-renewing corneal epithelium. 2 Severe ocular surface diseases, such as chemical burns, 3 thermal burns, 4 Stevens-Johnson syndrome, 5 and iatrogenic injury caused by ocular surgeries, 6 can cause direct damage of the LSCs and/or destruction of their microenvironment. This pathological change was named as limbal stem cell deficiency (LSCD), which may result in conjunctivalization, neovascularization, and chronic inflammation of the ocular surface, and eventually lead to severe vision loss or blindness.7,8
In recent three decades, tremendous efforts have been made to treat LSCDs. Tissue-engineered corneal epithelium transplantation has proved effective to restore the population and function of LSCs in severe LSCD cases.9–11 After that, numerous studies have been conducted to optimize the construction of tissue-engineered corneal epithelium. Among those, selection of suitable seed cells, development of biocompatible cell carrier, and modification of culture medium have been research hotspots in this field.12,13 However, terminal differentiation, tans-differentiation of corneal epithelial cells, as well as insufficient LSCs during construction still remain hurdles of current corneal epithelium tissue engineering protocols, 14 which will eventually affect the outcome of surgery.
P38 mitogen-activated protein kinase (MAPK), a family member of the mitogen-activated protein kinases, mediates a wide variety of cellular behaviors in response to extracellular stimuli and implicated to play an important role in inflammatory responses. 15 It was also found as a major modulator of cell differentiation processes of various cell types such as intestinal epithelial cells, 16 pulmonary alveolar epithelium,17,18 and blood vessel endothelial cells. 19 Previous study found that p38 MAPK signal was downregulated during ex vivo expansion of limbal epithelium on intact amniotic membrane compared with those cultured on plastic surface 20 ; however, the significance of p38 MAPK signal on the renewal of limbal epithelial stem cells was not illustrated. We have previously noted that human limbal explants cultured at air-liquid interface could induce abnormal epidermal differentiation through activation of p38 MAPK signaling pathway; addition of the p38 inhibitor abolished abnormal differentiation without affecting limbal epithelial proliferation.21,22
Our previous study also showed that p38 inhibitor applied as a culture medium additive could help ex vivo expansion of skin epithelial progenitor cells. 23 We then propose that controlling the activation of p38 MAPK signaling pathway at a relatively low level may be beneficial for the quality of tissue-engineered corneal epithelium.
To test this hypothesis, we applied SB203580 (SB), the specific inhibitor targeting p38α and p38β of p38 MAPK signaling pathway, to the construction of tissue-engineered corneal epithelium. Indeed, we found that inhibition of p38 MAPK signaling pathway could help maintain the stemness of corneal epithelium, protect the corneal epithelium cells against terminal differentiation, prevent abnormal differentiation and trans-differentiation, and thus improve the quality of tissue-engineered corneal epithelium.
Methods
Nine New Zealand white rabbits were used to establish the tissue-engineered corneal epithelium. For each experiment, three rabbits were sacrificed by carbon dioxide inhalation and the limbal tissues were harvested by removal of central corneas with a 9.25 mm diameter trephine; then the residual limbal tissues were cut into 12 blocks about 2 × 2 mm in size. They were incubated with 2 mg/mL Dispase II (Roche, Basel, Switzerland) in supplemental hormonal epithelial medium (SHEM), made of 0.5% DMSO, 0.5 μg/mL hydrocortisone (both Sigma-Aldrich, St. Louis, MO), DMEM/F12, 10 ng/mL mouse EGF, 10 μg/mL ITS, 5% FBS, and 1% Penicillin-Streptomycin (all Gibco, Carlsbad, CA) at 4°C for 12 h.
The epithelial cell sheets were isolated and cultured on the surface of inserts (BD, Franklin Lakes, NJ) in a six-well plate with SHEM medium (i.e., control group) or SHEM medium with 10 μM SB203580 (Yeasen, Shanghai, China) (i.e., SB group). The medium was changed every 2 days. After a 10-day cultivation, the epithelium was further cultured under airlifting manner for 5 days to promote stratification, as previously described (Fig. 1A). 24
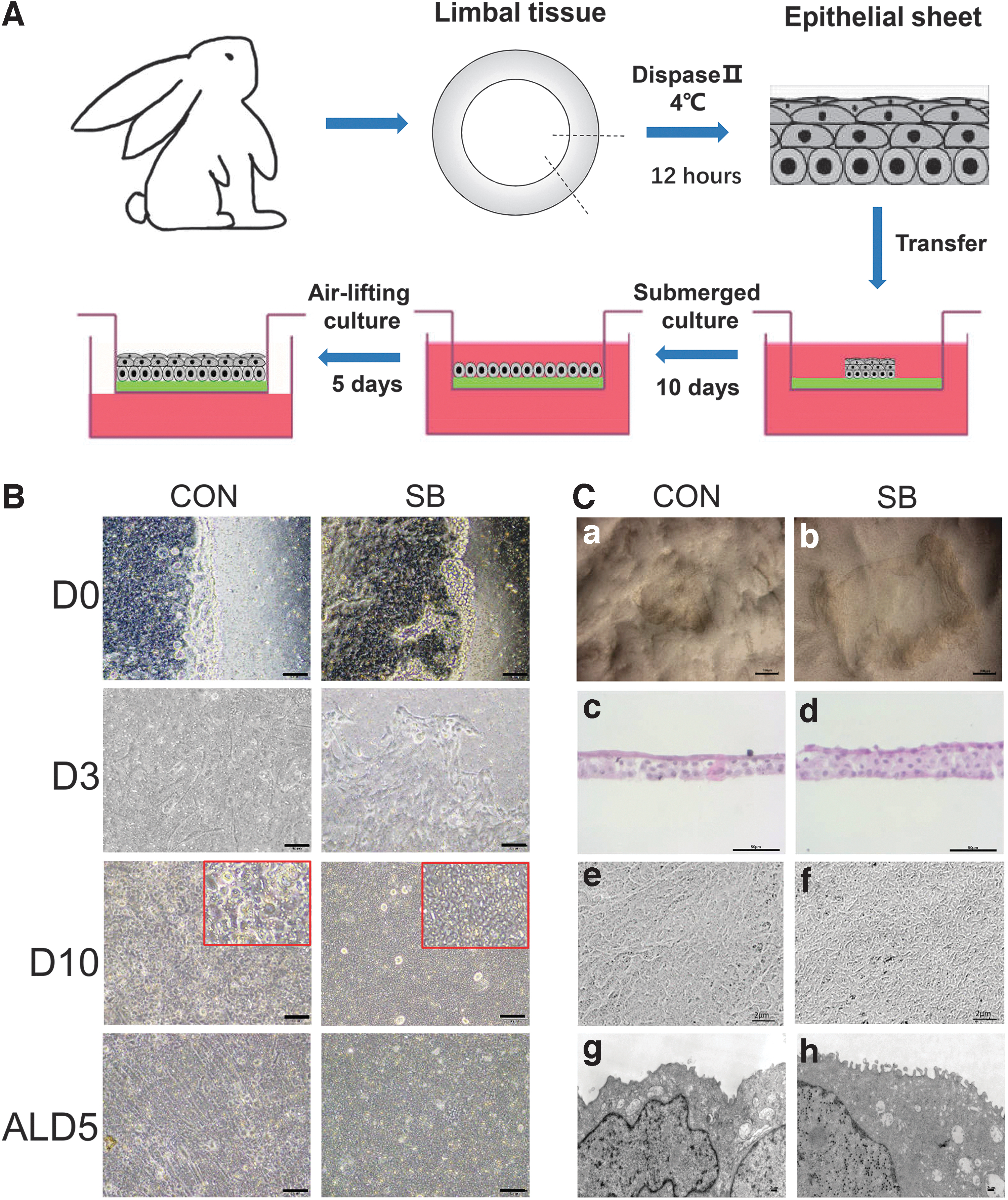
The construction of tissue-engineered corneal epithelium.
Twelve Fujian yellow rabbits were anesthetized through intramuscular injection of ketamine hydrochloride (0.1 mL/kg) (Dunhua City Shengda Animal Medicine, Dunhua, China) and 0.1% pentobarbital sodium (2 mL/kg) (Xiamen Minbo Biotechnology, Xiamen, China). One eye of each animal was subjected to a 2 mm wide and 0.2 mm deep limbal lamellar keratectomy and central corneal epithelial scraping to generate LSCD model. The animals were randomly divided into three groups with four rabbits in each group. SB group (SB) animals (n = 4) were transplanted with tissue-engineered corneal epithelium generated with SB, control group (CON) animals (n = 4) were transplanted with tissue-engineered corneal epithelium generated without SB, and animals without transplantation (n = 4) were considered no treatment group (NT). The lateral normal eye from each animal was defined as normal group.
The tissue-engineered corneal epithelia were mounted on the recipient corneal stroma surface with basal side down using Porcine Fibrin Sealant Kit (Sealant-pharma, Hangzhou, China) as previously reported, 24 and the whole cornea was covered with a bandage contact lens to protect the transplanted epithelium. Postoperatively, the eyes were administered TobraDex eye gel and TobraDex eye drops (both Alcon, Fort Worth, TX) once a day. The eyes were observed and photographed under a slit-lamp microscope (Kanghua Science & Technology, Chongqing, China) on day 0, 7, 10, and 14. The animals were sacrificed by carbon dioxide inhalation 2 weeks after the surgery and the corneal and limbal tissues were harvested for histological examination.
Experiment
Animals
The Female New Zealand white rabbits and Fujian yellow rabbits (weighing 2–2.5 kg, aging 3–4 months) were obtained from Shanghai SLAC Laboratory Animal Center and Fujian Lianjiang Yuhuashan Natural Eco-Agriculture Experimental Farm, respectively. The study received the experimental procedure approval of the Animal Ethics Committee of Xiamen University (Approval No. XMULAC20170014).
Hematoxylin and eosin staining
Enucleated rabbit eyes were fixed freshly in prepared 4% paraformaldehyde (Sigma-Aldrich) in phosphate-buffered saline (PBS) and embedded in paraffin. Corneal sections of 5 μm thickness were stained with hematoxylin and eosin (H&E) (both Sigma-Aldrich) using standard protocol. 25 The images were captured by BX60 microscope (Olympus, Allentown, PA).
Western blot analysis
At the end of cultivation, epithelial cell sheets were collected by cell scraper, and extracted in a cold RIPA buffer (Sigma-Aldrich), containing phosphatase and protease inhibitors cocktails (Thermo Fisher Scientific, Waltham, MA). Protein concentration was measured by bicinchoninic acid assay (Thermo Fisher Scientific). Equal amounts of protein extracts (20 μg) were subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to PVDF membranes (Roche). Membranes were blocked in 5% BSA for 2 h. After that, the membranes were incubated overnight at 4°C with primary antibodies for p38, phosphor-p38 (p-p38), MAPK activated protein kinase 2 (MAPKAPK2), phosphor-MAPKAPK2 (p-MAPKAPK2), heat shock protein family B (small) member 1 (HSPB1), phosphor-HSPB1 (p-HSPB1), catenin beta 1 (CTNNB1), phosphor-CTNNB1 (p-CTNNB1), cadherin 1 (CDH1), snail family transcriptional repressor 1 (SNAI1), claudin 1 (CLDN1), paired box 6 (PAX6), keratin 12 (KRT12), keratin 10 (KRT10), keratin 13 (KRT13), actin alpha 2, smooth muscle (ACTA2), or actin beta (ACTB).
After three washes with Tris buffered saline containing 0.05% Tween-20, the membranes were incubated with secondary antibody for 1 h at room temperature (RT). The results were visualized by enhanced chemiluminescence reagent (NCM Biotech, Suzhou, China) and recorded by the transilluminator (ChemiDoc XRS System; Bio-Rad). Densitometric analysis of Western blot results was performed by Image J software (National Institutes of Health, Bethesda, MD). Table 1 shows information of antibodies applied in the Western blot procedure.
Antibodies Applied for Western Blot
Scanning electron microscopy
The samples were fixed in cacodylate-buffered 2.5% glutaraldehyde for 1.5 h, dehydrated in alcohol series (30%, 50%, 70%, 80%, 95%, and 100%) each for 15 min, and dried by the critical point method. After that, tissues were sputter-coated with gold-palladium and examined with JSM-6390LV scanning electron microscope (Jeol, Tokyo, Japan).
Transmission electron microscopy
The samples were fixed in glutaraldehyde for 4 h, washed three times in cacodylate buffer for 15 min and postfixed in 1% osmium tetroxide for 90 min. After fixation, samples were dehydrated in acetone (30%, 50%, 70%, 95%, and 100%), embedded in Spurr's resin and cut into ultrathin sections with an ultramicrotome (EM TP; Leica). Sections were stained with aqueous uranyl acetate, lead citrate and examined with HT-7800 transmission electron microscope (Hitachi, Tokyo, Japan).
Immunofluorescence staining
Rabbit peripheral cornea and limbal tissues were fixed in optimal cutting temperature compound at −80°C overnight. Longitudinal sections of 6 μm thickness were cut and stored at −80°C. Immunofluorescence staining was performed as we previously reported. 26 In brief, corneal sections were fixed with 4% paraformaldehyde for 20 min, permeabilized with 0.2% Triton X-100 in PBS for 20 min, and blocked with 2% BSA for 2 h at RT. After that, corneal sections were incubated overnight at 4°C with primary antibodies for CTNNB1, CLDN1, CDH1, KRT12, PAX6, ACTA2, SNAI1, kertain 14 (KRT14), and kertain 15 (KRT15). After four washes with PBS, the sections were incubated with secondary antibody for 1 h at RT. Following four washes with PBS, slides were mounted with DAPI (Vector Labs, Burlingame, CA) and imaged using Zeiss LSM 880 confocal microscope (Carl ZEISS, Jena, Germany). Table 2 shows antibodies applied in the immunofluorescent staining procedure.
Antibodies Applied for Immunofluorescent Staining
Total RNA extraction and quantitative real-time polymerase chain reaction
Total RNA was extracted from tissue-engineered cell sheets by using TRIzol Reagent (Invitrogen, Eugene, OR). RNA quantity was measured by a Du 800 Nucleic Acid/Protein Analyzer (Beckman Coulter, Indianapolis, IN). Reverse transcription was performed with Oligo18T primers and reverse transcription reagents. Quantitative real-time polymerase chain reaction (PCR) was performed with mRNA primers listed in Table 3. PCRs were performed on a BIO-RAD CFX-96 Real-Time system with TB Green Premix Ex Taq (TaKaRa, Shiga, Japan) according to the manufacturer's protocol. 27 All reactions were performed in triplicate and the average cycle threshold values greater than 40 were treated as negative. The results were normalized by ACTB as internal control.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Clone culture of corneal limbal epithelial cells from tissue-engineered epithelium
Epithelial sheets cultured from the rabbit limbal tissues were digested with 2 mg/mL Dispase II at 4°C for 12 h, followed by 0.05% trypsin/0.53 mM EDTA digestion at 37°C for 15 min to render the epithelial sheets into single cells. A colony formation assay for epithelial progenitor cells was performed using the seeding density of 200 cells/cm2, as previously reported. 28 After 10 days of culture, the cells were fixed in 4% paraformaldehyde for 20 min and stained with 0.5% crystal violet (Sigma-Aldrich) in methanol for 30 min at RT. The cells were imaged, and the number of colonies was counted under an inverted microscope.
Statistical analysis
Statistical analysis was performed using GraphPad Prism Software (San Diego, CA). Summary data were reported as mean ± SD. Group means were compared using the appropriate version of Student's unpaired t-test. Test results were two tailed, where p < 0.05 was considered statistically significant.
Results
Construction of tissue-engineered corneal epithelium
The rabbit limbal epithelia were expanded in SHEM medium (control group, CON) or SHEM medium with SB203580 (SB group, SB) under submerged manner shown in Figure 1A. After 3 days of culture, criss-crossed single spindle cells were found migrating out from the cell sheet in control group, while the corneal epithelial cells migrated outward in sheets in SB group.
Epithelial cells in both groups reached confluence of 4.67 cm2 inserts with 0.4 μm pore size in six-well plants on day 10, and the size of cells was spherical and homogeneous in SB group, compared to larger and more disheveled cells in CON group under 100 × magnifications. After that, epithelial tissues were switched into airlifting culture manner, and prominent stratification was noticed in both groups after 5 days. During the construction of tissue-engineered corneal epithelium, irregular morphology and fibroblast-like cells could be found in control group, while cells were more regular and no fibroblast-like cells were observed in SB group (Fig. 1B).
After 2 weeks of culture, we used spatula to remove the tissue-engineered corneal epithelium from the insert and found it was easy to get intact cell sheets from SB group (Fig. 1C-a), while the cell sheets always broke into fragments in control group (Fig. 1C-b), indicating the cell sheets from SB group has higher strength characteristic. H&E staining of epithelial cross-sections showed that cell sheets from control group contained three to four layers of cells, while there were five to six layers in SB group. Both groups contained flattened cells with polarity in superficial layer, wing cells in middle layer, and small cells with high nuclei-cytoplasmic ratio in basal layer (Fig. 1C-c, d). Scanning electron microscopy images (Fig. 1C-e, f) and transmission electron microscopy images (Fig. 1C-g, h) showed that the tissue-engineered corneal epithelia from SB group contained more microvilli on the surface compared with control group.
P38 MAPK and Wnt/CTNNB1 signaling pathways in tissue-engineered corneal epithelium
Two constructions of tissue-engineered corneal epithelium expressed a similar level of total-p38, while p-p38 showed much higher expression in control group compared with that of SB group (Fig. 2A). We also identified decreased expression of p-MAPKAPK2 and p-HSPB1, downstream molecules of p38 MAPK signaling pathway, in SB group (Fig. 2A, B).
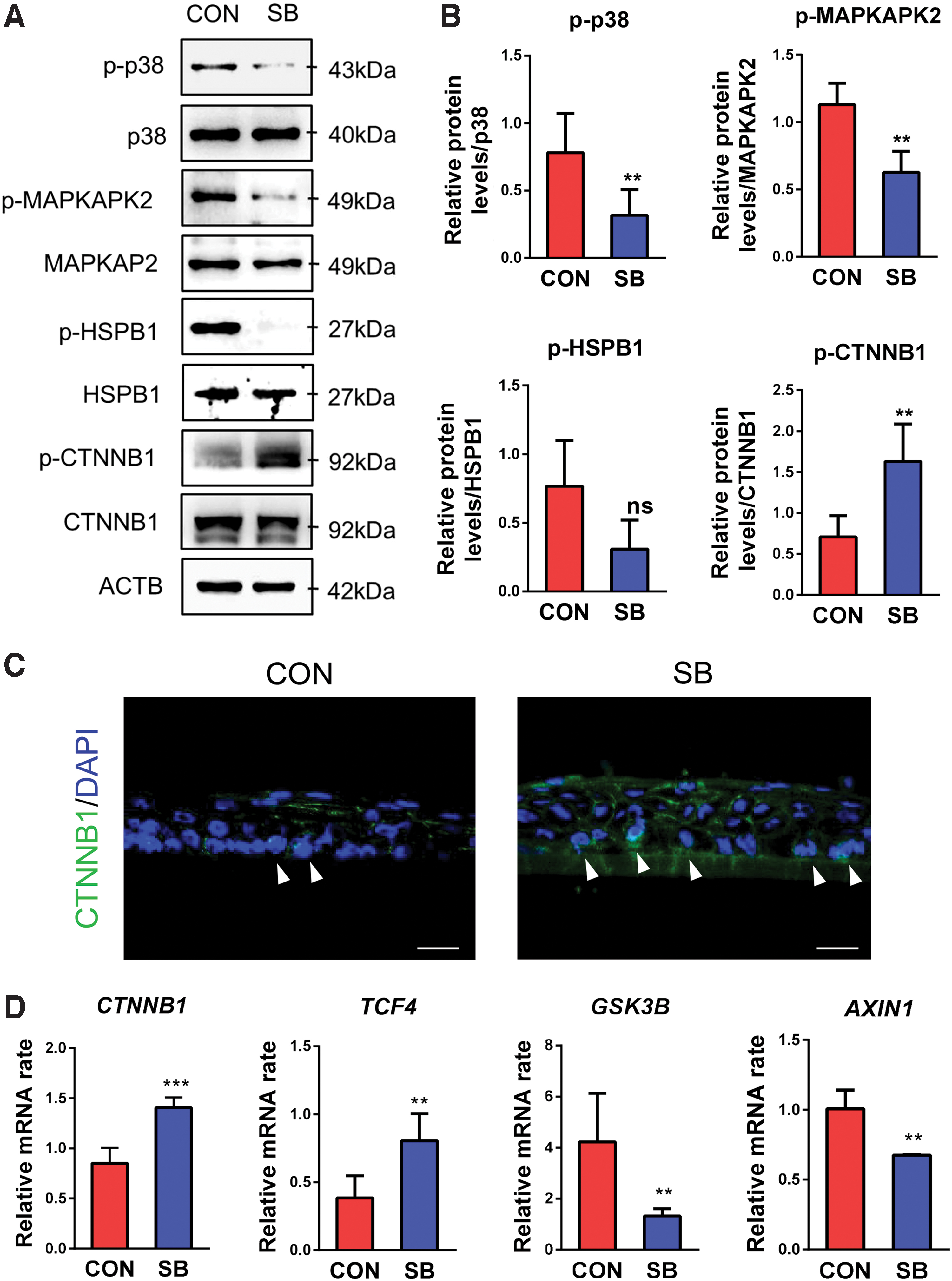
Effects of application of SB203580 on p38 MAPK and Wnt activation in tissue-engineered corneal epithelium.
Wnt/CTNNB1 signaling pathway is highly involved in self-renewal of various epithelial stem cells, including corneal epithelial progenitor cells. 2 We investigated the status of Wnt/CTNNB1 signaling pathway in tissue-engineered corneal epithelium and found that the phosphorylated protein level of CTNNB1 (p-CTNNB1) (Fig. 2A, B) and mRNA expression of CTNNB1 (Fig. 2D) were both upregulated in SB group. Immunostaining revealed more basal layer cells in the tissue-engineered corneal epithelium-expressed CTNNB1 in the nucleus after addition of SB203580 to culture system (Fig. 2C), indicating activation of Wnt/CTNNB1 signaling pathway. Transcription factor 4 (TCF4), the downstream molecule of the Wnt/CTNNB1 signal, was significantly activated in SB group (Fig. 2D). Moreover, glycogen synthase kinase 3 beta (GSK3B) and axin1 (AXIN1) gene expression was downregulated in SB group (Fig. 2D). These results indicated that SB203580 could inhibit p38 MAPK signaling pathway in cooperation with the high activity of canonical Wnt/CTNNB1 signaling pathway in the construction of engineered corneal epithelium.
Cell junction formation of the tissue-engineered corneal epithelium
Immunostaining showed that CDH1 was strongly expressed in the superficial layer, while weakly expressed in the basal and suprabasal layers in control group. In contrast, it was expressed in the full thickness of cultivated cell sheets in SB group (Fig. 3A). The other major component of cell-cell junction complex, CLDN1, showed weak expression in control group, while it was heavily expressed in the cell border of SB group (Fig. 3A). The scanning electron microscopy images clearly demonstrated low density of desmosomes and relatively big intercellular space in tissue-engineered corneal epithelium from control group (Fig. 3B-a, c), while there was high density of desmosomes and no obvious intercellular space presented in SB group (Fig. 3B-b, d).
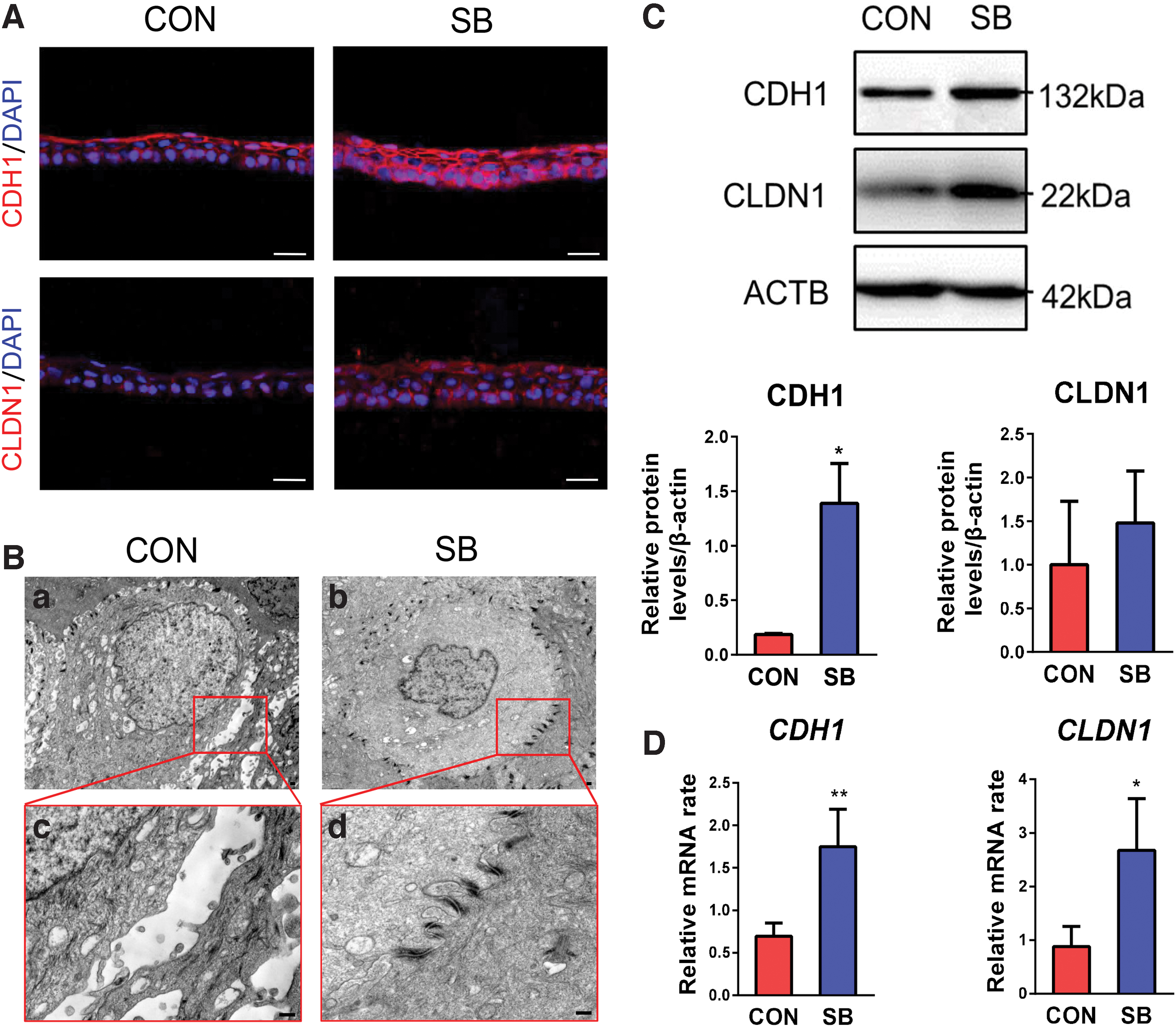
The cell junction of tissue-engineered corneal epithelium.
Western blot analysis confirmed the expression of CDH1 and CLDN1 proteins (Fig. 3C), and quantitative RT-PCR revealed their mRNA expression in tissue-engineered corneal epithelium (Fig. 3D). These results indicated that the cell junction formation was stronger and more regular in tissue-engineered corneal epithelium constructed with p38 inhibitor. This may partially explain the better integrity of epithelial sheets that could be harvested in the end of culture, as shown in Figure 1C.
Cell phenotype of the tissue-engineered corneal epithelium
The terminal differentiation marker of KRT12 was expressed in one to two superficial layers of tissue-engineered corneal epithelia in SB group compared to CON group in which nearly full cells expressed KRT12 (Fig. 4A), and the KRT12 mRNA and protein expression were much lower in SB group (Fig. 4B, C). PAX6, the transcription factor, which is highly expressed in the corneal epithelium and weakly expressed in the limbal epithelium, showed strong expression in epithelium from control group, while was weakly expressed in SB group (Fig. 4A). Western blot and qRT-PCR further confirmed lower expression of PAX6 in SB group (Fig. 4B, C). These results indicated that the general phenotype of tissue-engineered epithelium is similar as peripheral corneal epithelium in control group, while it is close to limbal epithelium in SB group.
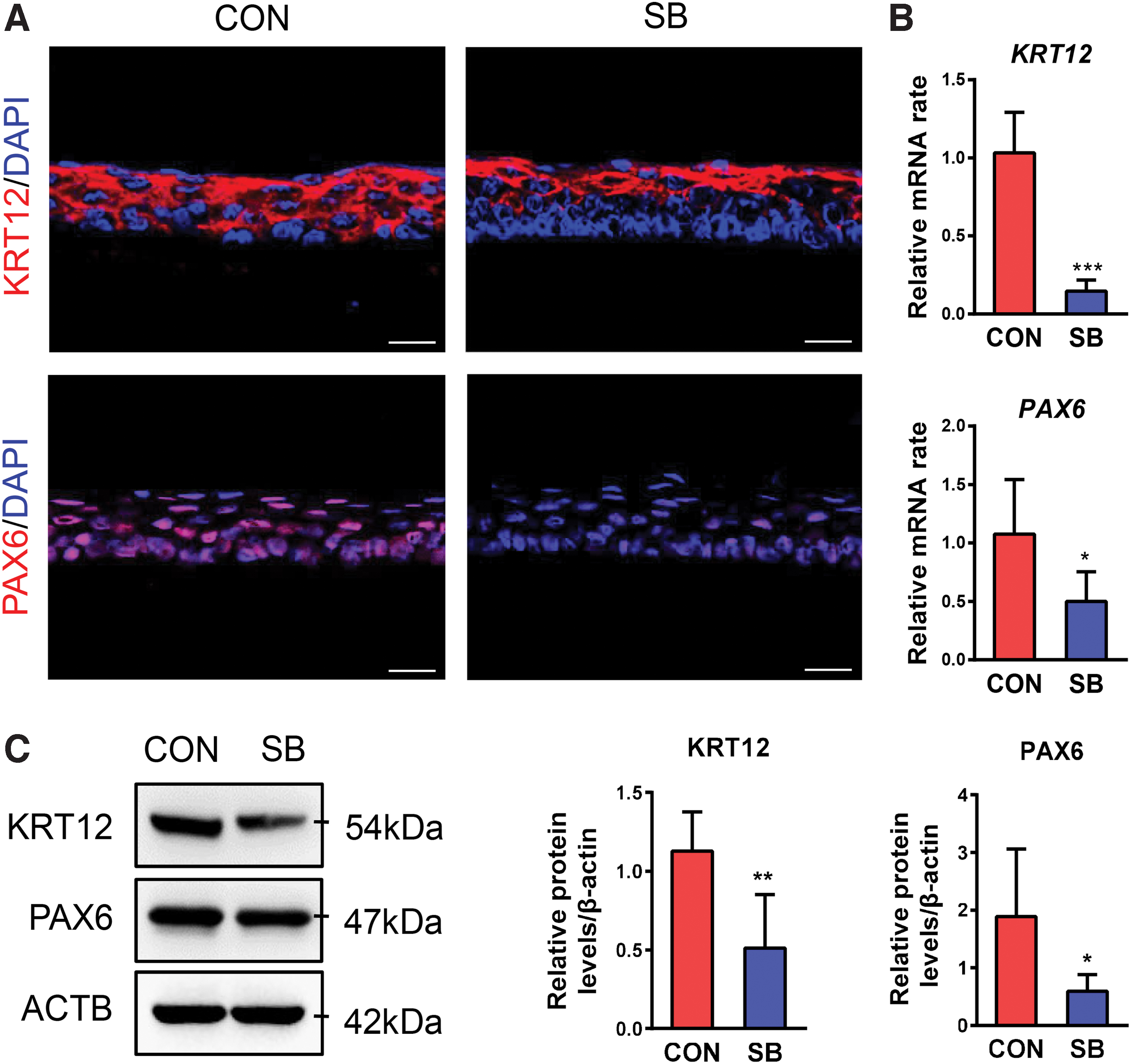
The cell phenotype of tissue-engineered corneal epithelium.
It was reported that activation of p38 MAPK signaling pathway was involved in epithelial mesenchymal transition (EMT). 29 We found ACTA2 and SNAI1 were expressed in some basal epithelial cells from control group, while there was no expression in SB group (Fig. 5A). Similar pattern was confirmed by Western blot and qRT-PCR (Fig. 5B–D). Meanwhile, abnormal differentiation marker KRT10 and terminal conjunctival differentiation marker KRT13 both decreased at protein levels and mRNA levels in SB group (Fig. 5B–D). These results indicated that cells in tissue-engineered corneal epithelium showed a lower level of terminal differentiation, abnormal differentiation, and trans-differentiation in the cell sheets cultured with SB203580.
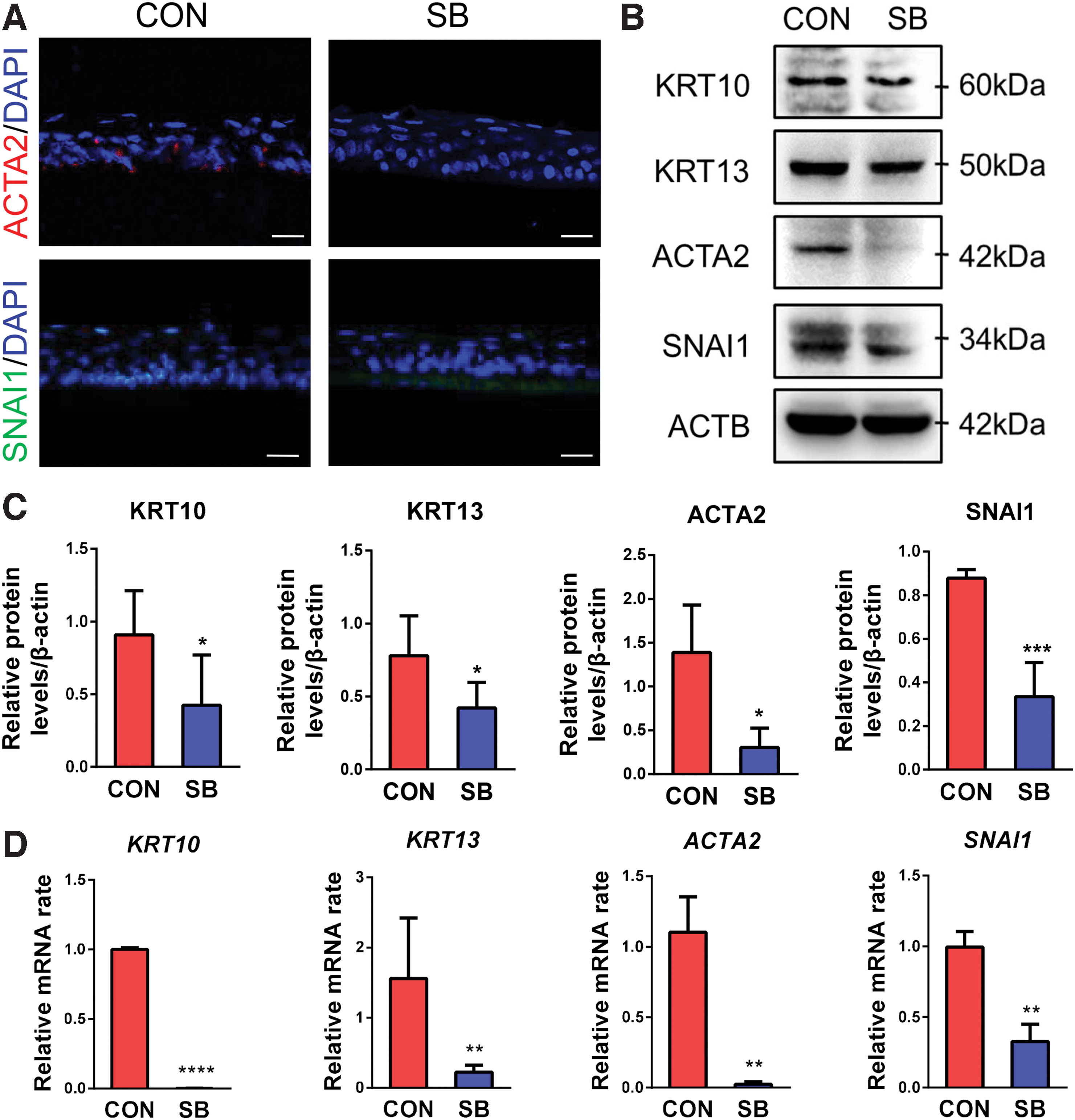
The abnormal differentiation of tissue-engineered corneal epithelium.
The stemness of tissue-engineered corneal epithelium
The replenishment of stem cell pool is the crux of tissue-engineered corneal epithelium during clinical treatment. In the view of low differentiation of corneal epithelium in SB group, we compared the characteristic of stemness in tissue-engineered corneal epithelial cell sheets. KRT14 is considered a progenitor cell marker of stratified epithelium such as skin and cornea, 30 mainly located in the basal layer and suprabasal layer of the epithelial sheet in control group. However, KRT14 expression expanded to almost full thickness of the epithelium in SB group (Fig. 6A). The gene expression of KRT14 was much higher in SB group compared with control group (Fig. 6B), which was comparable to the immunofluorescence staining. qRT-PCR results revealed that other epithelial stem/progenitor cell markers, such as ATP binding cassette subfamily G member 2 (ABCG2), tumor protein p63 (TP63), and Wnt family member 7A (WNT7A), were all higher in SB group (Fig. 6B).

The stemness of engineered corneal epithelium.
Furthermore, we harvested the cells from tissue-engineered corneal epithelium after 15 days of cultivation and co-cultured with 3T3 fibroblast feeder layers in SHEM medium for 10 days. The phase-contrast images and crystal violet staining showed that colony size was generally bigger and the shape was regular in cells from SB group (Fig. 6C), stands for characteristic of holoclone or meroclone, as previously described. 31 The colony-forming efficiency (CFE) was much higher in epithelium generated from SB group compared with that of control group (Fig. 6D). These results supported the notion that p38 inhibitor SB203580 could help expansion of the progenitor cell pool and maintain the stemness of engineered corneal epithelium.
Ocular surface tissue remodeling after transplantation of tissue-engineered corneal epithelium
To determine the efficacy of these epithelia on ocular surface reconstruction, we transplanted the tissue-engineered corneal epithelium on the ocular surface of rabbit LSCD model, which presented characteristics such as persistent corneal epithelial defect, stromal edema, neovascularization, chronic inflammation, and abnormal expression of KRT12, PAX6, KRT14, and KRT15 after surgery, as shown in Supplementary Figure S1. Two weeks after the operation, the corneal surface epithelial defect was completely healed in both groups based on the fluorescein staining (Fig. 7A). However, the wound closure was faster in corneas transplanted with epithelium from SB group (Fig. 7B). H&E staining showed regular structure and small size basal epithelial cells in the limbus and central cornea after transplantation of epithelium from SB group, while the limbal epithelial was less organized in control group (Fig. 7C).
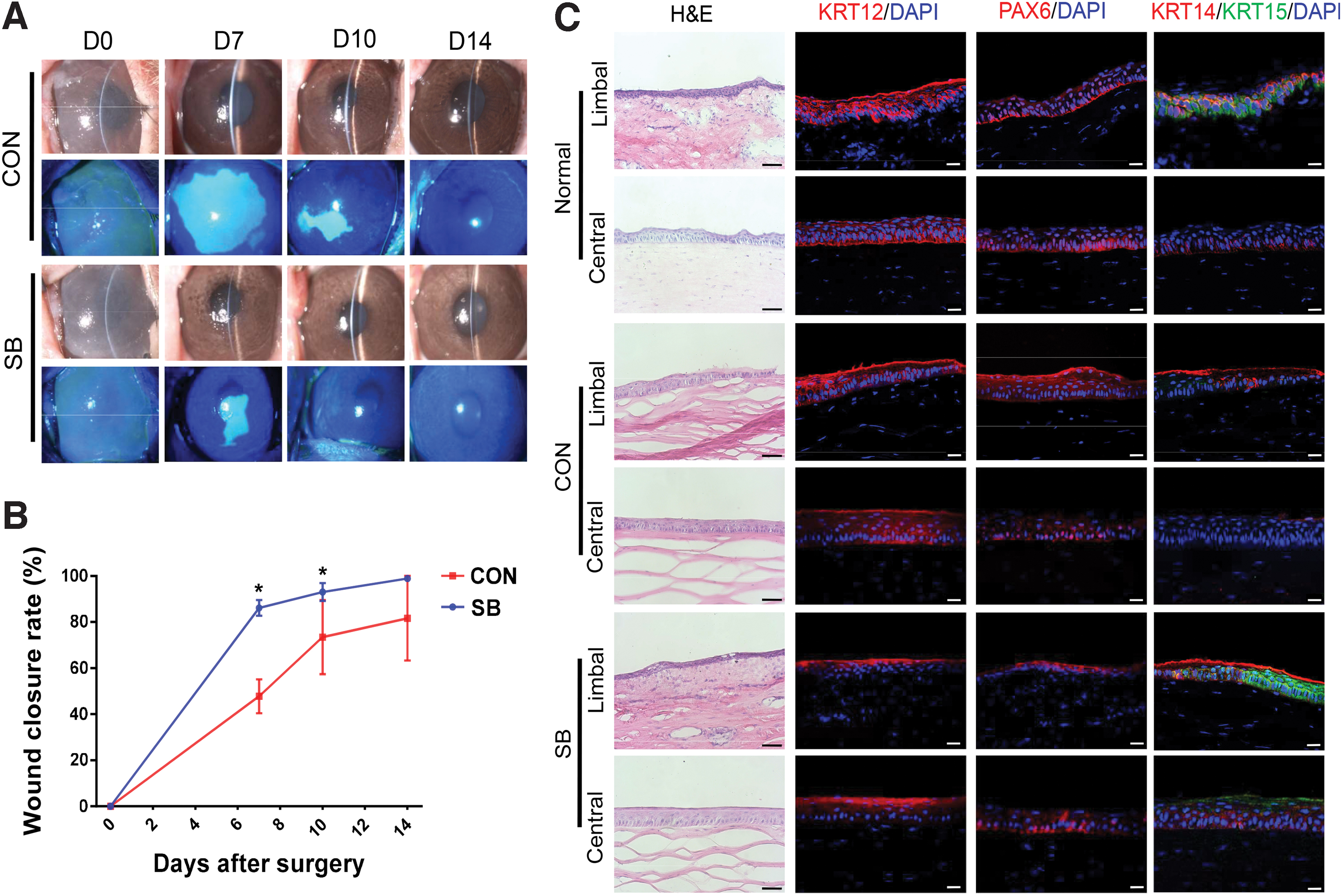
Rabbit ocular surface reconstructed with tissue-engineered corneal epithelium.
KRT12 and PAX6 expression were present in full thickness throughout the central and limbal corneal epithelium in control group, while KRT12 located exclusively to the superficial epithelium, and PAX6 was weakly expressed in the epithelium of SB group (Fig. 7C). The cells in the limbus of SB group highly expressed KRT15, while it was almost negative in control group (Fig. 7C). There were obviously more KRT14-positive cells in both limbal and central cornea with tissue-engineered epithelium from SB group, compared with that from control group (Fig. 7C), indicating the reconstructed limbal and corneal epithelia contain more cells that maintain progenitor phenotype in SB group. As a result, the outcome of ocular surface reconstruction was overall better after transplantation of tissue-engineered corneal epithelium generated from SB group.
Discussion
For the first time, we applied p38 MAPK signaling pathway modulator in corneal epithelial tissue engineering. With the presence of p38 MAPK inhibitor SB203580 in the culture medium, the tissue-engineered corneal epithelium showed better quality in general, and was successfully transplanted on the ocular surface of a rabbit LSCD model.
We clearly observed that corneal epithelial cell sheet grew on the culture insert membrane in a more organized pattern with the presence of p38 inhibitor. As a result, the epithelial sheet was more compact, the cell shape was more homogeneous, and the cell size was smaller. With the upregulation of CDH1 and CLDN1, p38 MAPK inhibitor could enhance gap junction and tight junction, compared to the loosened cell sheet in conventional culture medium. Previous study also found enhanced gap and tight junctions of hepatocytes by SB203580 treatment both in vivo and in vitro. 32
The tissue-engineered corneal epithelia from SB group contained more microvilli on the apical surface, indicating that p38 inhibition facilitated normal structure of corneal epithelial cells. These results supported the notion that tissue-engineered epithelium exhibited higher strength and tension in SB group, thus was easy to be harvested as an intact cell sheet from polyethylene terephthalate membrane, and was much easier to handle during surgery.
The purpose of LSCD therapy is to replace damaged corneal epithelial cells, mobilize the LSCs in limbal niche, and reconstruct the environmental homeostasis. 8 The cells from SB group maintained the shape of basal limbal epithelial cells, with strong expression of ABCG2, WNT7A, TP63, and KRT14, indicating their progenitor phenotype. CFE of the tissue-engineered epithelium, which is a golden standard of epithelial stem cell indicator, was <20% in control group, while was nearly 80% in SB group. This dramatic difference indicated that inhibition of p38 MAPK signaling pathway could promote the self-renewal of corneal LSCs, increase the stem cell pool, and maintain the stemness of epithelial progenitors in the tissue-engineered corneal epithelium. Providing sufficient LSCs is the main purpose of transplantation of tissue-engineered corneal epithelium. Animal study using rabbit LSCD model further confirmed the improved quality of tissue-engineered corneal epithelium with the application of p38 inhibitor.
Airlifting culture is a common maneuver in epithelial tissue engineering procedure to promote stratification of the epithelium. The airlifted condition not only promoted epithelial stratification, migration, and proliferation but also induced abnormal differentiation of corneal epithelial cells into skin phenotype. 33 Our previous study on limbal explant culture first demonstrated this phenomenon, and we found that the activation of p38 MAPK signaling pathway was involved in this process. 22 Application of p38 inhibitor largely prevented abnormal differentiation.
In this study, we also shifted the cell culture from submerged manner to airlifting manner on day 10 and further cultured for 5 days to promote stratification of the epithelium. We found the stratification process was not interrupted by the presence of SB203580. However, abnormal differentiation was significantly prevented by SB203580, and the cells went through normal lineage commitment into limbal epithelial phenotype. This ensured normal phenotype of ex vivo constructed cell sheet for clinical transplantation.
EMT is another common phenomenon that happens in limbal explants cultured at air-fluid interface.34,35 This study also confirmed the presence of EMT in the tissue-engineered epithelium after airlifting culture for 5 days. Based on our previous protocol of separating limbal epithelium from underneath stroma, it is not likely to have any stromal fibroblast contamination in the culture system; 35 therefore, the ACTA2 and SNAI1-positive cells in the tissue-engineered corneal epithelium demonstrated those cells underwent EMT. However, the application of p38 inhibitor in the culture system dramatically reduced mRNA and protein expression of ACTA2 and SNAI1, the key regulators of EMT, indicating SB203580 could help maintain normal phenotype of limbal epithelium by inhibiting the abnormal transdifferentiation.
Wnt/CTNNB1 signaling pathway plays a vital role in various cellular processes, such as stem cell self-renewal, proliferation, and differentiation.36,37 In our previous study, we found Wnt/CTNNB1 signaling pathway was activated in the ocular surface epithelium under air exposure manner. 38 WNT7A, one of the modulators of Wnt/CTNNB1 signaling pathway, could be upregulated in wounded corneas, and promote proliferation of human corneal epithelial cells in vitro.39,40 Previous report also found Wnt/CTNNB1 signaling, activating with high levels of ABCG2 and ΔNp63α expression in cultured limbal epithelial cells in the presence of LiCl. 41
Interestingly, we found that Wnt/CTNNB1 pathway was upregulated with increase of WNT7A after SB203580 treatment. We propose that Wnt signal and p38 MAPK signal work together to regulate proliferation and differentiation of limbal stem/progenitor cells. Air exposure activates both signaling pathways simultaneously; inhibition of p38 signaling pathway may provoke Wnt/CTNNB1 pathway. The cross talk between Wnt signaling pathway and p38 MAPK signaling pathway in limbal stem/progenitor cells deserves further investigation.
Conclusion
Downregulation of p38 MAPK signaling pathway in corneal epithelial stem cells helps maintain stemness and protect the corneal epithelium cells against terminal differentiation, squamous metaplasia, and EMT, and enhance the cell junction during ex vivo expansion. Application of p38 inhibitor may be beneficial to the construction of tissue-engineered corneal epithelium. Small molecules that modulate signaling pathways may play important roles in tissue engineering protocols in the future.
Footnotes
Authors' Contributions
Y.M. and S.O.: conceptualization (equal); methodology (equal); data curation (equal); writing—original draft (equal); formal analysis (equal); and writing—review and editing (equal). W.L.: conceptualization (lead); methodology (lead); writing (lead)—original draft (lead); formal analysis (lead); and writing—review and editing (lead). C.Z., S.L., X.L., and M.L.: investigation (supporting). J.Y., Y.W., H.H., R.Z., Z.L., and Z.L.: resources (supporting).
Disclosure Statement
No competing financial interests exist.
Funding Information
This study is supported, in part, by the National Key R&D Program of China (2018YFA0107301 [to W.L.]), the National Natural Science Foundation of China (NSFC; No. 81870625 [to R.Z.], No. 81970773 [to W.L.], No. 82101084 [to S.O.]), and the China Postdoctoral Science Foundation (2021M69898 [to S.O.]). The funders have no role in the study design, data collection and analysis, decision on publishing, or preparation of the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.