
Abstract
Patient-oriented applications of cell culture include cell therapy of organ failure like chronic renal failure. Clinical deployment of a cell-based device for artificial renal replacement requires qualitative and quantitative fidelity of a cultured cell to its in vivo counterpart. Active specific apicobasal ion transport reabsorbs 90–99% of the filtered load of salt and water in the kidney. In a bioengineered kidney, tubular transport concentrates wastes and eliminates the need for hemodialysis, but renal tubule cells in culture transport little or no salt and water due to dedifferentiation that mammalian cells undergo in vitro thereby losing important cell-type specific functions. We previously identified transforming growth factor-β (TGF-β) as a signaling pathway necessary for in vitro differentiation of renal tubule cells. Inhibition of TGF-β receptor-1 led to active and inhibitable electrolyte and water transport by primary human renal tubule epithelial cells in vitro. Addition of metformin increased transport, in the context of a transient effect on 5′-AMP-activated kinase phosphorylation. These data motivated us to examine whether increased transport was an idiosyncratic effect of SB431542, probe pathways downstream of TGF-β receptors possibly responsible for the improved differentiation, evaluate whether TGF-β inhibition induced a range of differentiated tubule functions, and to explore crosstalk between the effects of SB431542 and metformin. In this study, we use multiple small-molecule inhibitors of canonical and noncanonical pathways to confirm that inhibition of canonical TGF-β signaling caused the increased apicobasal transport. Hallmarks of proximal tubule cell function, including sodium reabsorption, para-amino hippurate excretion, and glucose uptake increased with TGF-β inhibition, and the specificity of the response was shown using inhibitors of each transport protein. We did not find any evidence of crosstalk between metformin and SB431542. These data suggest that the TGF-β signaling pathway governs multiple features of differentiation in renal proximal tubule cells in vitro. Inhibition of TGF-β by pharmacologic or genome engineering approaches may be a viable approach to enhancing differentiated function of tubule cells in vitro.
Impact statement
Cell therapy of renal failure requires qualitative and quantitative fidelity between in vitro and in vivo phenotypes, which has been elusive. We show that control of transforming growth factor-β signaling can promote differentiation of renal tubule cells grown in artificial environments. This is a key enabling step for cell therapy of renal failure.
Introduction
Cell and tissue culture holds great promise for medicine far beyond its origin in viral culture. Our area of interest, artificial and biohybrid vital organs, is vitally dependent on the function of cultured cells. Unfortunately, progress has been hampered by the dedifferentiation of epithelial cells in ex vivo culture, a phenomenon described as cell culture stress. In the body, renal tubule epithelial cells are essential for homeostasis of fluid volume, electrolyte concentrations, endogenous organic solutes, and xenobiotics. Renal proximal tubule cells selectively and actively transport solutes bidirectionally between luminal filtrate and basolateral capillaries.
Tubule cells cultured in laboratory conditions rapidly lose transporter expression, apical brush border microvilli, and revert to glycolytic energy generation. The importance of the renal proximal tubule in removal of protein-bound uremic toxins (PBUTs), reabsorption or excretion of drugs, and as an important site of dose-limiting toxicity for new molecular entities has led to a variety of efforts to improve the fidelity of in vitro cell culture to the in vivo counterpart.
Our group has been working to develop cells and culture techniques that will allow a bioreactor of epithelial cells to perform concentration and excretion functions in an artificial kidney.1–8 We showed that cultured renal tubule cells exposed to an inhibitor of transforming growth factor-β (TGF-β) signaling in combination with metformin increased transporter expression and inhibitable apicobasal transport. 7 That work was encouraging as it showed that tubule cells could be induced to transport salt water in vitro. We wanted to gain further insight into the mechanisms and effects of TGF-β inhibition. Proximal tubule cells primarily reabsorb salt via the sodium-proton exchanger 3 (NHE3) and also reabsorb glucose via the sodium-glucose cotransporter 2 (SGLT2). The proximal tubule excretes organic anions, including uremic toxins, through basolateral organic anion transporters.
TGF-β is representative of a superfamily of cytokines involved in regulation of cell proliferation and apoptosis across multiple organs, including the kidney. 9 Ligand binding to the TGF-β receptor 2, a transmembrane serine-threonine kinase, phosphorylates TGF-β receptor 1 (TGF-βR1), triggering a series of downstream events that are grouped into “canonical” pathways involving phosphorylation of receptor-regulated Smads, and a wide range of “non-canonical” pathways. The end consequences of TGF-β signaling are both tissue and context-specific. To confirm that the effect of SB431542 on apicobasal transport was indeed due to TGF-βR1 inhibition rather than an idiosyncratic off-target of SB431542, we tested two inhibitors of TGF-βR1, also known as Activin-like Kinase 5 (ALK5), SB431542, and A8301.
To evaluate whether canonical (SMAD) or noncanonical effects of TGF-β inhibition underlie the differentiation response we previously observed, 7 we subsequently tested an inhibitor of SMAD3 and inhibitors of phosphoinositide-3 kinase, the mammalian target of rapamycin, p38 mitogen-activated protein kinase MAPK, and TGF-β-activated kinase 1. We assessed two other functions of tubule cells, glucose uptake and organic anion excretion, to test whether the effects of TGF-β inhibition were limited to sodium transport or included additional measures of proximal tubule function. Finally, as the beneficial effect of metformin was unanticipated, we examined whether SB431542 also affected an effect of metformin AMPK phosphorylation. Conversely, we tested whether metformin affected canonical TGF-β signaling by measuring the effect of metformin on SMAD phosphorylation.
In this work, we show that canonical TGF-β inhibition is necessary and sufficient for renal proximal tubule cells to actively transport fluid volume, reabsorb glucose, and excrete organic anions in vitro. Thus, we demonstrate that there may be comparatively simple approaches to fostering differentiation of tubule cells on conventional cell culture substrates that are commercially available, simple to handle, and integrate well with microscopy and cell culture tools.
Materials and Methods
Cell culture
Primary human renal tubule cells were purchased from Lonza (Cat CC-2553; Basel, Switzerland) or Innovative Biotherapies (Ann Arbor, MI). Cells were used up to fourth passage. Cells were seeded onto 1.12 cm2 permeable supports (Transwell, Corning, Oneonta, NY). Expression of claudin-2, claudin-7, aquaporin-1, and megalin was measured by polymerase chain reaction (PCR) to confirm proximal tubule origin of the primary cells. All other materials were purchased from Sigma (St. Louis, MO) unless otherwise indicated. Cells were cultured in 50:50 Dulbecco's modified Eagle's medium:F12 with hydrocortisone, triiodothyronine, ascorbic acid, insulin, transferrin, and selenium, with 5% fetal bovine serum in the basolateral media only [4].
Small molecule inhibitors of TGF-β inhibitor type 1 (SB431542, 10 μM, A8301, 1 μM, both from Cayman Chemical, Ann Arbor, MI), SMAD3 (SIS3, 5 μM, Cayman Chemical), as well as LY294002 (1 μM, inhibitor of phosphoinositide 3-kinase, Cayman), rapamycin (40 nM, inhibitor of the mammalian target of rapamycin, or mTOR, Cayman), SB203580 (1 μM, inhibitor of p38 mitogen-activated protein kinase or p38 MAPK, Cayman), takinib (2 nM, inhibitor of TGF-β-activated kinase 1 or TAK1, Cayman), and metformin (200 μM, Sigma) were added to cell culture media for the indicated experiments. Texas red isothiocyanate-conjugated dextran (TRITC dextran; 10,000 kD, No. D1868; ThermoFisher, Waltham, MA) was added in some experiments to evaluate monolayer confluence. TRITC-Dextran was added to the apical compartment of the cell culture insert to a final concentration of 100 μg/mL, and 24 h later, aliquots of apical and basolateral media were sampled.
Fluorescence at 550 nm excitation and 571 nm emission was measured to quantify dextran concentrations in apical and basolateral compartments. Cells were grown on a shaker table (73 rpm; equivalent average fluid shear stress 2 dyn/cm2) in 95% air 5% carbon dioxide at 37°CCelsius. Media was changed every 72 h.
Transport measurement
Primary proximal tubule cells (Lonza) at 19–42 weeks of stable culture with and without SB431542 had media changed to identical media with or without addition of tenapanor (1 μM; AdooQ Bioscience LLC). Twenty-four hours later, apical and basolateral media were aspirated and weighed. A blank well with the porous membrane occluded with epoxy was used as an evaporative control. The difference between initial volumes and final volume as estimated by weight was taken as the transported volume.
RNA isolation and real-time PCR
Total RNA was isolated using the Micro or Mini RNeasy kits (Qiagen, Hilden, Germany). RNA quality was determined by measuring absorbance at 260 and 280 nm on a Nanodrop Spectrometer. First-strand cDNA was synthesized from total RNA using the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA) according to the manufacturer's instructions. Real-time PCR was performed on triplicate samples using SsoAdvanced Universal SYBR Green Supermix (Bio-Rad) and a Bio-Rad CFX96 Real-Time PCR System. Data were normalized to human GAPDH mRNA levels as an endogenous control.
Relative expression (RE) levels are expressed relative to static control using the ΔΔCt formula (RE = 2 − [(Ct(gene, test sample) − Ct(GAPDH, test sample)) − (Ct(gene, static sample) − Ct(GAPDH, static sample))]), in which CT is the threshold cycle number. Bio-Rad CFX Manager Software version 3.1 was used to determine CT numbers and relative expression levels. PCR primer sequences were from PrimerBank database 1 and oligonucleotides were synthesized by the Vanderbilt University Molecular Biology Core.
Western blotting
Protein concentration was quantified using Pierce BCA assay (ThermoFisher). Samples were prepared at a concentration of 10 μg protein per well with 4 × protein loading buffer (LICOR, Lincoln, NE) supplemented with 1% DTT (Abcam, Cambridge, United Kingdom). Prior to running gels, samples were denatured at 95°C for 5 min. Samples were loaded into 4–20% gradient polyacrylamide gels and ran for 35 min at 200 volts at room temperature. Gels were then transferred to nitrocellulose membranes via wet transfer for 1 h at 100 volts at 4°C. Membranes were incubated with primary antibodies (1:500–1:1000) overnight at 4°C, then washed three times for 5 min with 1 × TRIS with 0.1% Tween 20. Membranes were incubated with secondary antibodies (1:25,000–1:50,000) for 1 h at room temperature then washed three times for 15 min with 1 × TBS.
Membranes were imaged with an Odyssey XF Imaging System (LICOR). Protein expression was quantified using ImageStudioLite or Empiria Studio (LICOR) and normalized to β-actin using the built-in normalization function in Empiria Studio. Antibodies to phospho-AMPK, phospho-Smad2, total Smad2, and SLGT2 were purchased from Cell Signaling (Danvers, MA). Antibodies to NHE3 and β-actin were purchased from Santa Cruz Biotechnologies (Dallas, TX). Antibodies to total AMPK were purchased from Invitrogen (Waltham, MA). Antibodies to OAT1 were purchased from ProteinTech (Rosemont, IL).
Liquid chromatography
Chromatography was performed on an Agilent 1200 series high-performance liquid chromatography (HPLC) system. Concentrations were determined using a diode array detector (Agilent G1315C, Santa Clara, CA) at 245 nm. An Aeris 3.6 μm, 150 × 4.6 Widepore XB-C18 (Phenomenex) column was used for separation. A phosphate buffer was prepared from Sodium Phosphate Dibasic Heptahydrate (Fisher Chemical, Ottawa, ON) and Sodium Phosphate Monohydrate (Fisher Chemical) and adjusted to a pH of 4.1. HPLC grade methanol (34860; Sigma-Aldrich) was added to equal 10% (vol%/vol%) for the mobile phase in an isocratic method. The method had a flow rate of 1 mL/min, injection volume of 15 μL, and a total run time of 8 min. Retention times for para-amino hippurate (PAH) and acetaminophen were 1.5 and 2.1 min, respectively. Powdered PAH (A1422; Sigma-Aldrich) was used to prepare standards and quality controls (QC). Acetaminophen (MP Biomedicals, LLC, Solon, OH) was used as the internal standard.
Samples were filtered with 30K centrifuge filters (Sartorius, United Kingdom). Standards were prepared at 5, 10, 30, 50, 70, 100 μg/mL [QCS at 20, 60, 80] levels in 30% methanol and distilled water. The working range for the method was 1—100 μg/mL PAH. Three QC levels per run were prepared directly into cell media.
Glucose measurement
Fresh cell culture media was added to both apical and basolateral compartments for 24 h. After 24 h, 100 μL of cell culture media from each sample was added to 1 mL Glucose Assay Reagent (Glucose [HK] Assay Kit, Sigma) and incubated at room temperature for 15 min. Approximately 2 μl from each sample was then placed on the spectrophotometer (Nanodrop, Thermo Scientific) and absorbance measured at 340 nm.
Statistical analysis
All statistical inference testing was performed on results from prespecified experiments. Differences in means were assessed by Welch's t-test with two tails and assumption of unequal variance using R (The R Foundation for Statistical Computing). Statistical significance was assigned at an alpha threshold of 0.05.
Results
Volume transport
Sodium reabsorption is an essential function of the renal tubule cell. We previously showed that treatment of renal tubule cells with SB431542, a TGF-βR1 inhibitor, increased apicobasal transport. We wished to determine if this was related to changes in gene expression of the predominant apical sodium-proton exchanger of the proximal tubule, NHE3. Further, we also examined whether the increased transport was an idiosyncratic response to SB431542 or was mediated by inhibition of TGF-βR1/ALK5 by other agents as well. We therefore tested a different TGF-β inhibitor, A8301, or SIS3, an inhibitor of the canonical TGF-β pathway protein Smad3.
Cells expressed claudin-2 mRNA at a 1570 ± 350—fold higher level than claudin-7. Cells also expressed mRNA for aquaporin-1 and megalin. Together, along with high-level expression of NHE3, OAT1, and SGLT2, these expression profiles suggest that the isolates we used were primarily proximal tubule cells. Cells grew to confluence in 10 days to 2 weeks. Cells formed confluent monolayers as measured by TRITC-conjugated dextran concentrations in the basolateral compartment <2% of apical concentrations after 24 h.
We compared apicobasal fluid volume transport (μL/cm2/day) as the indicator of phenotype between control cells, cells were treated with the TGF-βR1 inhibitor SB432542, the TGF-βR1 inhibitor A8301, or with an inhibitor of Smad3 phosphorylation, SIS3 (Fig. 1). Cells were cultured with SB431542 (107.1 ± 14.2 vs. 38.8 ± 11.9, p = 0.033) or SIS3 (98.2 ± 4.5 vs. 38.8 ± 11.9; p < 0.0001) or A8301 (98.2 ± 4.5 vs. 38.8 ± 11.9; p < 0.0001; Fig. 1 for all), all transported more fluid than corresponding control cells.
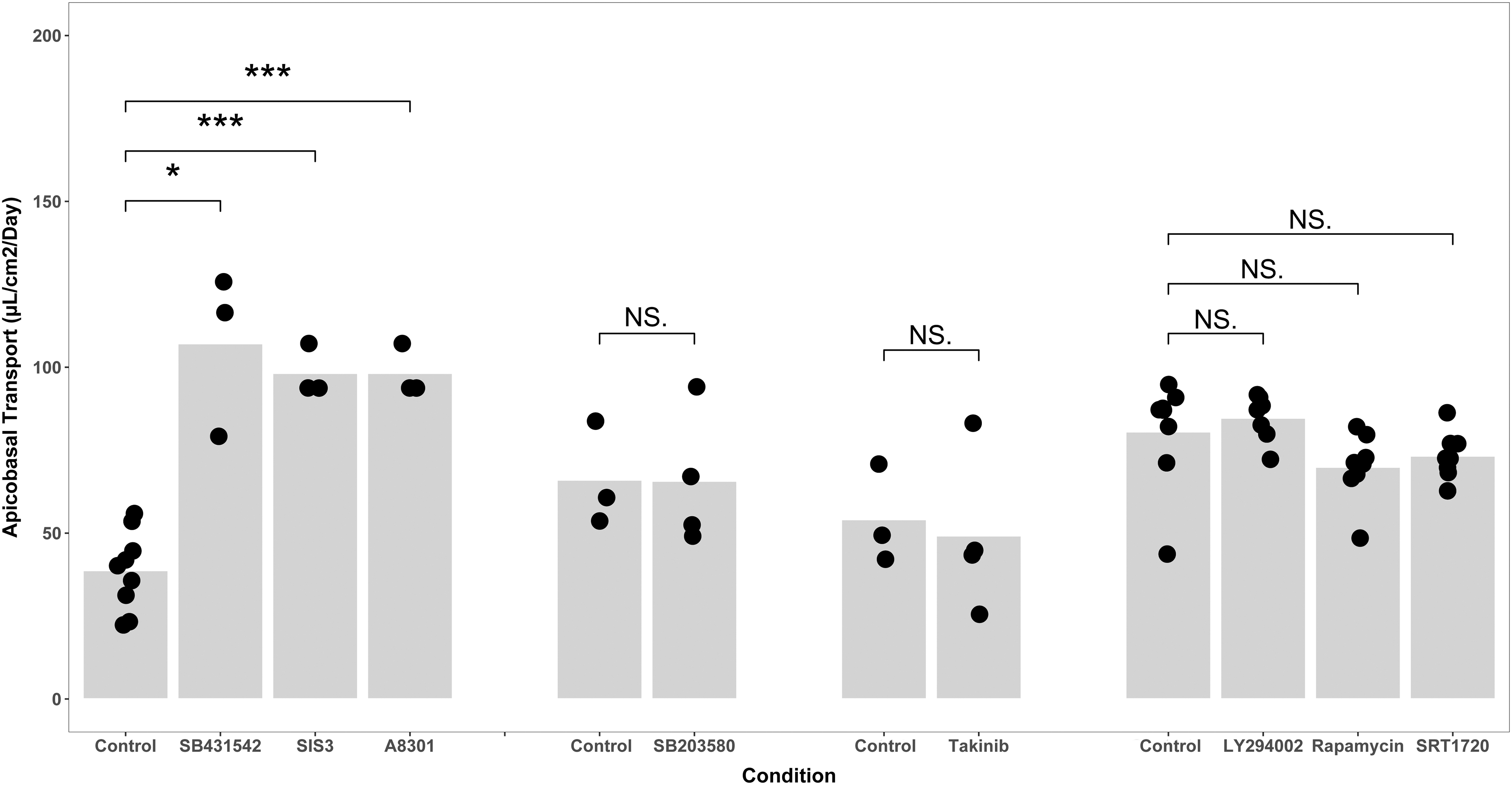
Inhibition of canonical TGF-β signaling increases fluid transport by renal tubule cells in culture. The results of four separate experiments are depicted in four groups above as the absolute transport values in the control groups vary from isolate to isolate. Primary renal tubule cells increased their fluid transport when either of two inhibitors of the TGF-β receptor or an inhibitor of Smad3 phosphorylation were added to media (left, *p < 0.05, *** p < 0.0001 by Welch's t-test). There were no significant differences in transport between cells cultured with inhibitors of noncanonical TGF-β and cells cultured in control media, which is 50:50 DMEM:F12 with hydrocortisone, triiodothyronine, ascorbic acid, insulin, transferrin, and selenium, with 5% fetal bovine serum in the basolateral media only (rightmost three groups). Data points indicate the number of biological replicates for each condition. DMEM, Dulbecco's modified Eagle's medium; TGF-β, transforming growth factor-β.
We further probed TGF-β signaling by measuring transport in primary renal tubule cells incubated with inhibitors of noncanonical TGF-β signaling, LY294002 (1 μM, inhibitor of phosphoinositide 3-kinase), rapamycin (40 nM, inhibitor of the mammalian target of rapamycin, or mTOR), SB203580 (1 μM, inhibitor of p38 mitogen-activated protein kinase or p38 MAPK), and takinib (2 nM, inhibitor of TGF-β-activated kinase 1 or TAK1). Cells retained barrier function as measured by inulin leak rates, but none of the noncanonical pathway inhibitors increased transport (Fig. 1). These data confirm that indeed canonical TGF-β signaling mediates the change in phenotype we observe.
We previously observed that metformin-induced AMPK activation enhances the effects of SB431542 in vitro. 7 To develop insight regarding the mechanisms of increased transport, we compared NHE3 expression between control cells and cells incubated with a combination of metformin and SB431542 (“M+SB”) at the mRNA and protein level. While SB431542 alone did not significantly change NHE3 mRNA or protein expression, cells cultured with M + SB expressed NHE3 mRNA at a 3.8-fold higher level compared to control (p = 0.027, Fig. 2), and expressed NHE3 protein by western blot at an approximately six-fold higher level than cells grown in control media (p = 0.032, Fig. 2). We compared apicobasal transport between renal proximal tubule cells grown in control media, media with added M + SB, and media with M + SB and the NHE3 inhibitor, tenapanor. 10
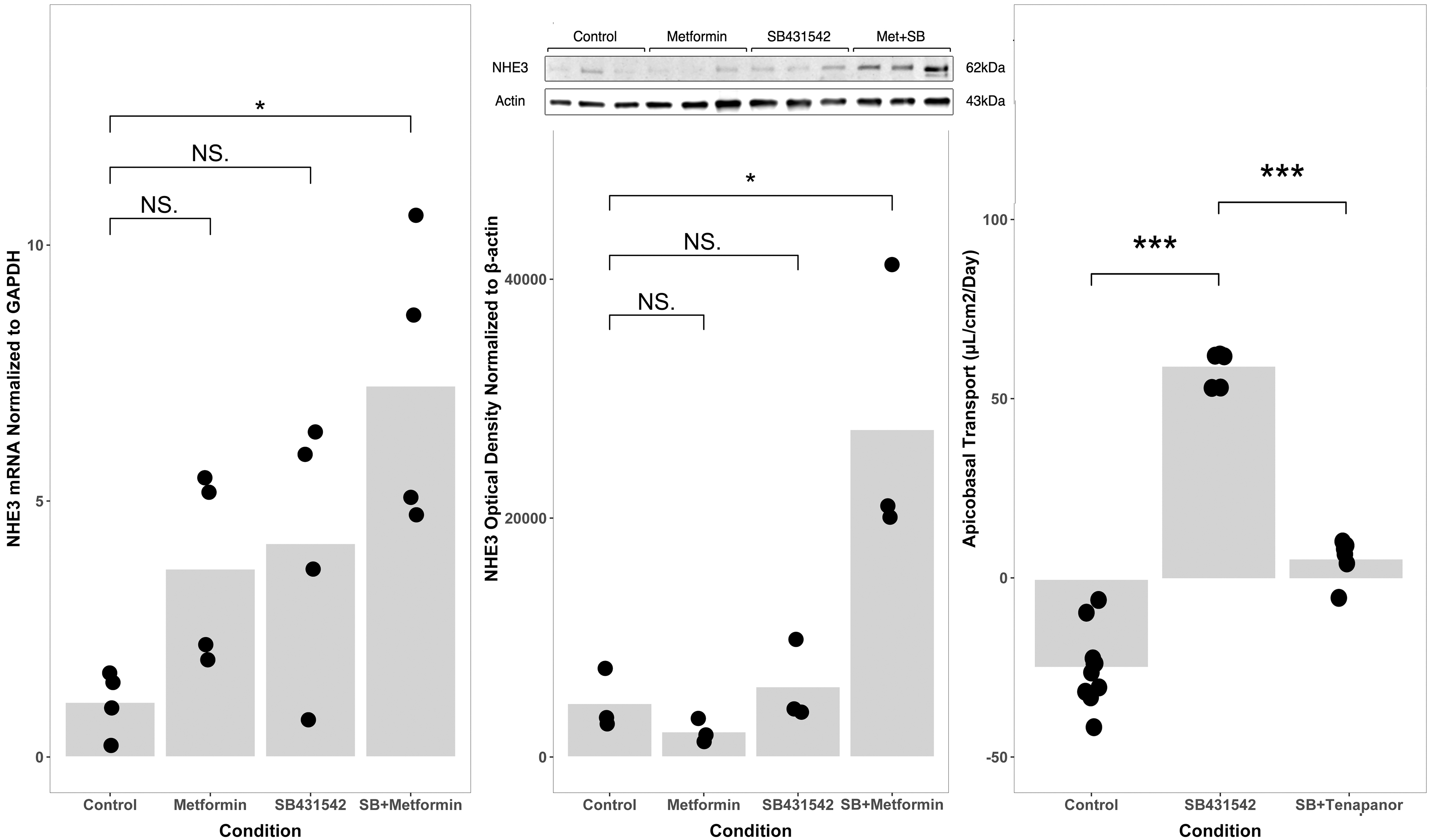
TGF-β inhibition increases NHE3-mediated apicobasal active transport. Primary renal proximal tubule cells were cultured in control media (50:50 DMEM:F12 with hydrocortisone, triiodothyronine, ascorbic acid, insulin, transferrin, and selenium, with 5% fetal bovine serum in the basolateral media only), media with added metformin, added SB431542, or both. NHE3 gene expression (left) was significantly higher in cells cultured with both (“SB + Metformin”, *p < 0.05 vs. control) while other comparisons were not statistically significant. NHE3 protein expression by western blot (middle panel inset) was significantly higher in the SB + Metformin group than in controls (middle panel; *p < 0.05) while other comparisons were not significant. Apicobasal volume transport was greater in SB431542-treated cells than in control cells (Right panel; 59.5 ± 4.6 μL/cm2/day vs. −24.8 ± 10.7 μL/cm2/day; ***p < 0.001). Addition of tenapanor, a specific inhibitor of NHE3, to cells cultured with SB431542 reduced transport to 5.7 ± 5.7 μL/cm2/day (***p < 0.001 compared to SB431542 alone). Data points indicate the number of biological replicates for each condition.
TGF-β blockade by SB431542 increased apicobasal transport (−24.8 ± 10.7 μL/cm2/day vs. 59.5 ± 4.6 μL/cm2/day; p < 0.001, Fig. 2) and transport was reduced to 5.7 ± 5.7 μL/cm2/day by addition of tenapanor to the media (p < 0.001, Fig. 2). NHE3 reabsorbs sodium by exchanging one sodium ion for one proton, so we were curious whether the proximal tubule cells altered the acidity of the apical media. Indeed, SB431542-treated cells acidified the apical media (pH 7.13 vs. pH 7.45, p = 0.010). These data show that the effect of TGF-β inhibition on transport in proximal tubule cells is specifically mediated by expression and activity of NHE3.
Glucose reabsorption
We wanted to further examine the phenotypic effects of TGF-β inhibition, testing whether other features of proximal tubule cells were enhanced in vitro by blockade of the TGF-β pathway. Proximal tubule cells reabsorb filtered glucose from the glomerular filtrate through the apical sodium-glucose cotransporter SGLT2. We compared SGLT2 expression between inhibitor and control media and measured whether tubule cells in culture depleted glucose from the apical, basolateral, or both compartments. Finally, we used an inhibitor of sodium-glucose cotransporter, phlorizin, to confirm that the effect we observed was specifically mediated by the SGLT2 transporter. Sodium-glucose cotransporter 2 (SGLT2) mRNA was expressed at a 2.3-fold higher level in cells cultured with the TGF-β inhibitor A8301 (p = 0.042, Fig. 3), and SGLT2 protein was higher by western blot (p = 0.037, Fig. 3).
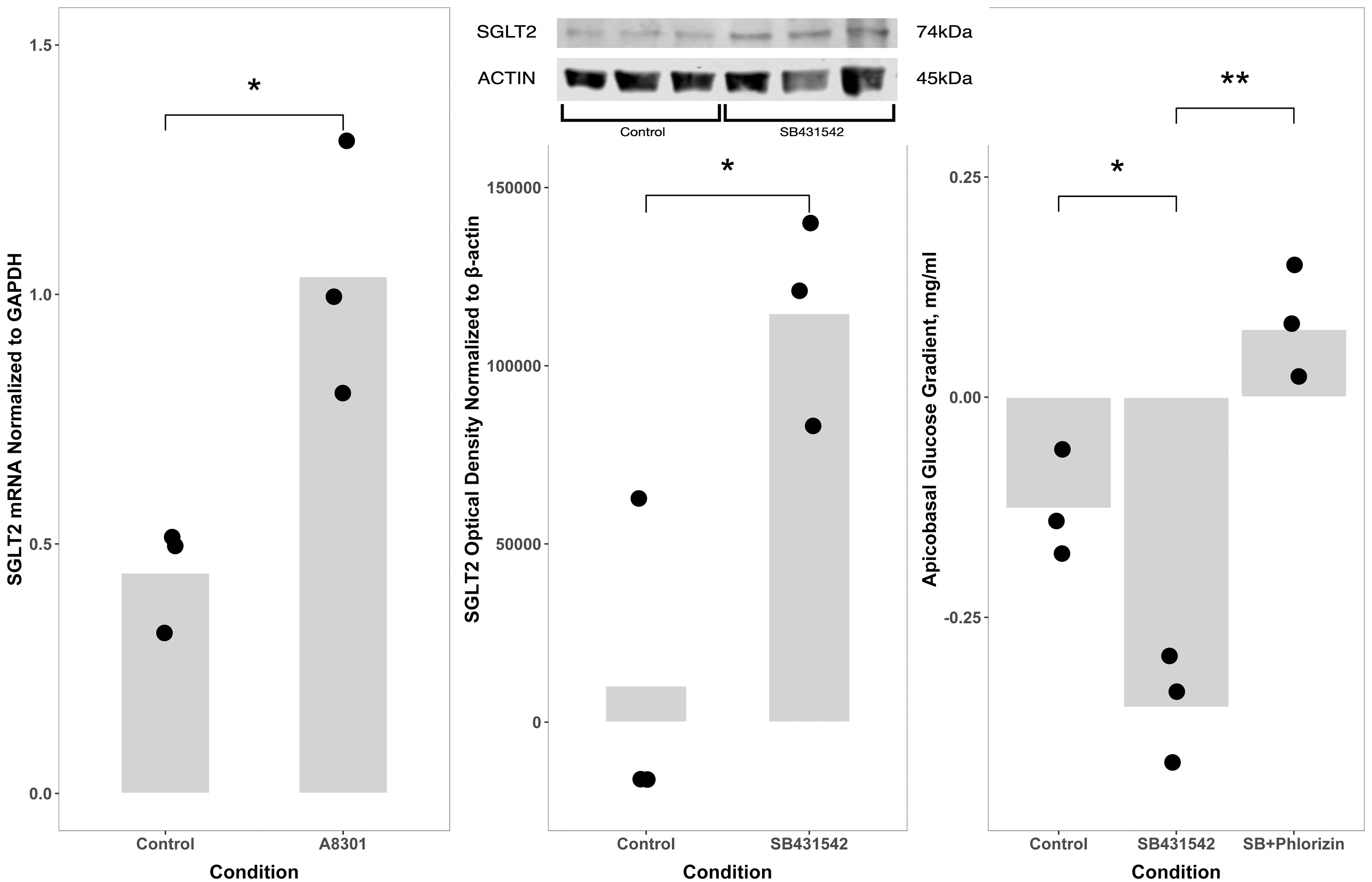
TGF-β inhibition increases glucose reabsorption by renal proximal tubule cells in vitro. Messenger RNA for the SGLT2 was expressed at a 2.3-fold higher level in cells cultured with A8301, a TGF-β receptor inhibitor, than in control cells (left panel; *p = 0.042). SGLT2 protein expression by western blot (insert, middle panel) revealed that cells cultured with SB431542, a TGF-βR1 inhibitor, expressed SGLT2 at a higher level than cells cultured in control media (50:50 DMEM:F12 with hydrocortisone, triiodothyronine, ascorbic acid, insulin, transferrin, and selenium, with 5% fetal bovine serum in the basolateral media only) (middle panel; *p < 0.05). Glucose concentrations in apical and basolateral media were measured 24 h after a media change. Cells cultured with the TGF-β inhibitor SB431542 reabsorbed glucose from apical media more than did control cells (apicobasal glucose concentration gradient after 24 h incubation: −352 ± 66 μg/mL vs. −126 ± 63 μg/mL; *p < 0.05), and addition of the SGLT inhibitor phlorizin prevented the reabsorption (−352 ± 66 μg/mL vs. 77 ± 59 μg/mL, **p < 0.01). Data points indicate the number of biological replicates for each condition. SGLT2, sodium-glucose cotransporter 2.
Cells cultured continuously for 42 weeks with the TGF-β inhibitor SB431542 reabsorbed glucose from apical media more than the control cells (apicobasal glucose concentration gradient after 24 h incubation: −126 ± 63 μg/mL vs. −352 ± 66 μg/mL; p = 0.012), and addition of the SGLT inhibitor phlorizin in a second series of experiments at 47 weeks in culture prevented the reabsorption (−352 ± 66 μg/mL vs. 77 ± 59 μg/mL; p = 0.001, Fig. 3). The observation that active, specifically inhibitable glucose transport is increased by TGF-β inhibition supports the hypothesis that the effects of TGF-β inhibition are more widespread than NHE3-mediated sodium transport alone.
Organic anion excretion
Proximal tubule cells contribute to excretion of endogenous and xenobiotic compounds from blood through high-affinity active transporters indirectly coupled to sodium gradients. Proximal tubule cells in conventional culture do not express organic anion transporters such as OAT1, so we examined whether OAT1 expression and organic anion excretion changed with TGF-β inhibition. OAT1 expression by PCR showed no trend toward increased expression with addition of SB431542. OAT1 expression by western blot, however, was significantly different between control and TGF-β-inhibited cells (p = 0.0031 SB431542 vs. control, Fig. 4). To examine function, we chose a classic OAT substrate widely used in renal physiology, PAH as an indicator of functional phenotype.
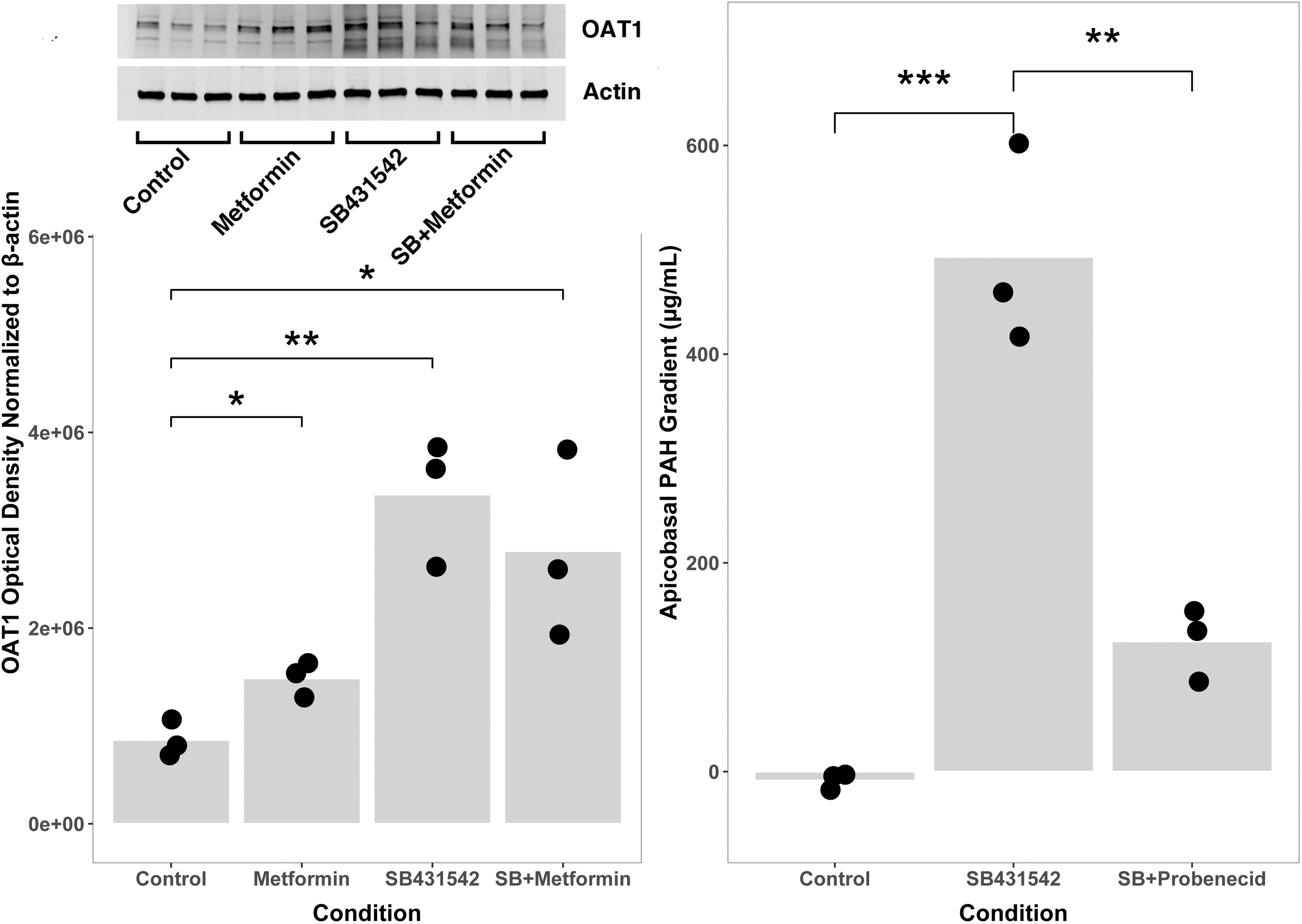
TGF-β inhibition increases OAT1 expression and function in primary renal tubule cells. Cells cultured with metformin, SB431542, a TGF-β inhibitor, or both, expressed OAT1 protein as measured by western blot (left panel inset) more than cells cultured in control media (50:50 DMEM:F12 with hydrocortisone, triiodothyronine, ascorbic acid, insulin, transferrin, and selenium, with 5% fetal bovine serum in the basolateral media only) (left panel, *< 0.05; **p < 0.01). Cells cultured with SB431542 transported PAH from basolateral to apical compartments, whereas control cells did not (apicobasal PAH gradient 493 ± 96 μg/mL in SB431542-treated wells vs. −8.5 ± 7.7 μg/mL in control wells, *** p < 0.001, right panel). PAH transport was inhibited by the addition of probenecid to media (apicobasal PAH gradient 493 ± 96 μg/mL in SB431542-treated wells vs. 125 ± 35 μg/mL in SB431542 + probenecid wells, **p < 0.01, right panel). Data points indicate the number of biological replicates for each condition. PAH, para-amino Hippurate.
We also evaluated a pharmacologic inhibitor of organic anion transport, the drug probenecid, to test the specificity of the response. Primary renal tubule cells cultured with SB431542 for 42 weeks transported PAH from basolateral to apical compartments, whereas control cells did not (apicobasal PAH gradient 493 ± 96 μg/mL in SB431542-treated wells vs. −8.5 ± 7.7 μg/mL in control wells, p = 0.00086). PAH transport was inhibited by the addition of probenecid to media (apicobasal PAH gradient 493 ± 96 μg/mL in SB431542-treated wells vs. 125 ± 35 μg/mL in SB431542 + probenecid wells, p = 0.0034). As with glucose transport, TGF-β inhibition increased organic anion transporter expression and specifically inhibitable active transport, confirming the specificity of the response to organic anion transport pathways.
Role of metformin
In our prior work, we noticed that metformin independently and additively increased active fluid transport by primary human cortical thick ascending limb cells. 7 The additive effect of the biguanide hypoglycemic agent metformin on differentiation is incompletely understood. Metformin has a mitochondrial role inhibiting Complex I, which at least transiently reduces ATP generation. Decreased ATP/ADP ratio allosterically alters AMP-activated protein kinase (AMPK) to permit liver kinase B1 (LKB1), the predominant kinase responsible for AMPK phosphorylation, to phosphorylate AMPK. Several groups have reported the influence of metformin on canonical TGF-β signaling, so we probed metformin's effects on TGF-β in these cells by comparing Smad2 phosphorylation in cells with and without metformin.11,12 We confirmed that metformin increased AMPK phosphorylation as predicted, while TGF-β inhibition had no effect on AMPK phosphorylation (Fig. 5).
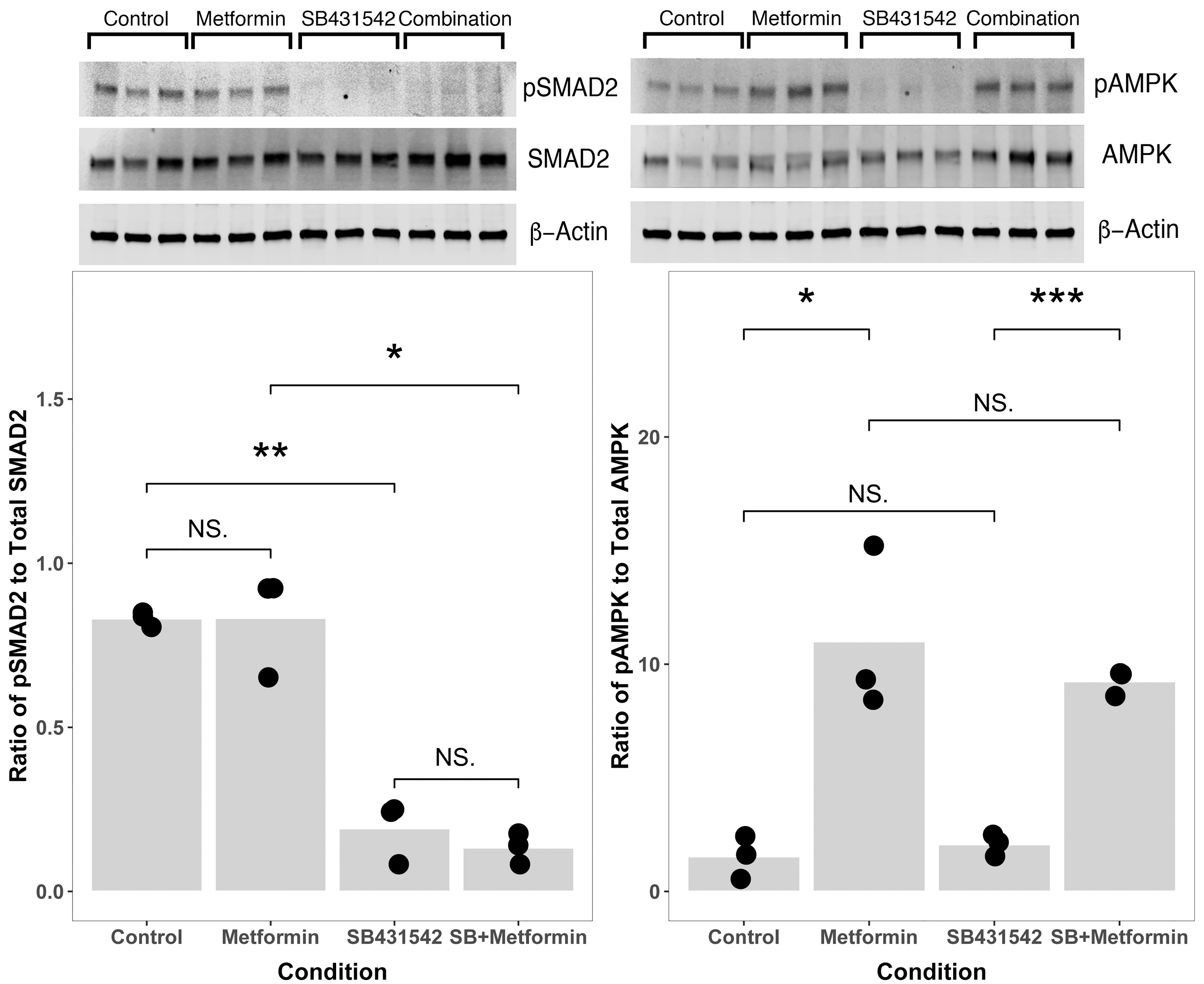
Metformin does not increase Smad2 phosphorylation in primary renal tubule cells, and SB431542 does not alter APMK phosphorylation. An inhibitor of TGF-β signaling SB43152 alone or in combination with metformin decreased Smad2 phosphorylation (left panel, *p < 0.05;**p < 0.01), but metformin alone does not significantly decrease Smad2 phosphorylation (left panel). Metformin alone or in combination with SB431542 increases AMPK phosphorylation (right panel, *p < 0.05; ***p < 0.001), but SB431542 alone has no effect on AMPK phosphorylation. The apparent additive effect of metformin and TGF-β inhibition may be attributable to independent signaling events rather than crosstalk between the two pathways. Data points indicate the number of biological replicates for each condition.
We found that for renal proximal tubule cells in vitro, metformin did not significantly affect a measure of canonical TGF-β signaling, Smad2 phosphorylation (Fig. 5). These data suggest that direct crosstalk between canonical TGF-β signaling and metformin's impact on AMPK do not explain the additive effects we observed.
Discussion
Cell culture stress
To place the current work in context, these data follow on decades of work by multiple teams defining the roles of soluble factors, genetic manipulations, and the physical microenvironment in the etiology of cell culture stress. Primary cultures of cortical epithelial cells derived from transplant discards maintain ammoniagenesis, vitamin D hydroxylation, and glucose transport when cultured under flow in the lumens of hollow-fiber bioreactors. 13 Culturing tubule cells on an orbital shaker improves aerobic metabolism and gluconeogenesis compared to static culture.14–17 In two-dimensional culture, albumin uptake by primary renal tubule cells increased with addition of apical fluid shear stress.2,3,17 Indeed, fluid shear stress affects multiple differentiation pathways, including some involved in noncanonical TGF-β signaling.15–17 Pharmacologic and genomic interventions can improve differentiation.
Small molecule beta-adrenergic stimulation increases mitochondrial number, morphology, and oxygen consumption of cultured proximal tubule cells. 18 A conditionally immortalized renal tubule cell line (ciPTEC) created with temperature-sensitive SV40 large antigen could support genetic overexpression of organic anion transporters (OATs). 19 Such cells showed greater sensitivity to nephrotoxins and increased excretion of classic PBUTs compared to cells without overexpression of OATs. Coculture of tubule cells with endothelial cells in a three-dimensional (3D)-printed convoluted channel seeded with commercially available TERT-immortalized proximal tubule cells yielded a cuboidal epithelium with a brush border, that reabsorbed glucose and albumin from apical perfusate,20,21 yet, cadaver-derived tubule cells seeded on porcine small intestine submucosa also formed a cuboidal epithelium with a mature brush border, strongly suggesting that 3D culture was not essential to tubule cell differentiation. 22
TGF-β and tubule cell function
The influence of TGF-β signaling on tubule cell differentiated function has been extensively studied. TGF-β signaling seems to be implicated in decisions between recovery as function versus recovery as fibrosis, and inhibition improved recovery in animal models of renal failure.23,24 Deletion of TGF-β receptor attenuated acute kidney injury in a mouse model, but was separately seen to worsen CKD.23–27 In previous studies, we found that primary renal tubule cells on conventional porous substrates exhibited diuretic-inhibitable apicobasal transport when cultured with SB431542, a small molecule inhibitor of TGFβR1. 7 We wanted to identify the pathways through which TGF-β was acting and learn whether TGF-β inhibition in vitro allowed renal tubule cells on conventional culture materials to display differentiated features in addition to fluid transport.
TGF-β appeared to increased transporter function through transcriptional upregulation of transporter abundance for SGLT2 and possibly OAT1, but SB431542 alone did not appear to affect NHE3 mRNA or protein concentrations, suggesting that the effects of the TGF-β pathway might occur through post-translational effects. One possibility might be that TGF-β activates protein kinase A (PKA) through an interaction of Smad3–Smad4 complex and the regulatory subunit of PKA. PKA activation phosphorylates/inhibits NHE3 and acts as a signal for NHE3 endocytosis. 28
We reinforced our conclusion that TGF-β signaling was causally related to the differentiation we observed, as multiple inhibitors of canonical TGF-β signaling reproduced the phenotype of increased transport. The role of metformin remains intriguing, as it affects transcription levels, but there was no evidence of increased Smad2 phosphorylation when metformin was added to media. AMPK is a pleiotropic enzyme that appears to regulate a variety of energy-consuming and energy-generating processes in mammalian cells. 29 AMPK activation in renal tubule cells is thought to be in the causal pathway of ischemic preconditioning through mTORC1 and the PI3/Akt pathway, reducing apoptosis of tubule cells with subsequent ischemia. 30
Conversely, AMPK activation by metformin in tubule cells with mutations in tuberous sclerosis complex gene TSC1 was associated with mitigation of the pathologic phenotype via increased apoptosis. 31 Together, these examples of metformin's interactions with renal tubule cells highlight that AMPK's role in these cells is highly tissue- and context-dependent.
Comparison of in vitro transport versus in vivo transport
TGF-β inhibition in vitro led to the appearance of functions characteristic of the in vivo renal tubule cell, including diuretic-inhibitable fluid transport, phlorizin-inhibitable glucose uptake, probenecid-inhibitable organic anion excretion, and apical acid extrusion. It is worth asking whether the magnitude of these desired phenotypes agrees to any extent with in vivo function. This is a vexing question as most in vivo or immediately ex vivo studies were performed decades ago and sometimes report results in different units and contexts than our work here. Our transport measurements in cells treated with TGF-β inhibitors correspond to about 12 μmol/cm2/day sodium transport. How does that compare to in vivo sodium transport? Giebish conducted numerous micropuncture studies of the rat proximal tubule, and measured transport ranging from 400–700 pmol/min/mm tubule. 32
Using electron microscope images to estimate tubule diameter and luminal area, Giebish's measurements in the rat proximal tubule extrapolate to 35–60 μmol/cm2/day. We observed that sodium transport within the same order of magnitude as in vivo transport was striking, especially as in vitro, our cells were not stimulated with large apicobasal oncotic gradients, catecholamines, or angiotensin, and cells were not perfused with free fatty acids, the preferred energy source for proximal tubule cells in vivo.
We measured total extraction of glucose from apical media to be 3.2 μg/mm2/day, less than the ∼10 μg/mm2/day Lewis measured in renal tubule cells continuously perfused with media in 3D culture, but much more than the ∼0.5 μg/mm2/day Lewis reported for tubule cells grown on Transwells similar to ours. 20 Some of this apparent difference may be attributable to the decrease in apical glucose concentration over the 24-h observation window. In our experiments, media was exchanged every 48–72 h, over which time the apical media glucose concentration decreased, whereas Lewis continuously perfused cells at a constant glucose concentration. The starting concentration of glucose in our media was 5 mmol/L or 900 μg/mL, and the end concentration averaged 171 μg/mL or 943 μmol/L, closer to the reported Michaelis constant (Km) of 500 μmol/L for SGLT2. 33 This suggests that the glucose transport we observed might have submaximal due to substrate concentration.
Our goal is to develop differentiated renal tubule cells for use in an artificial kidney. A major barrier to progress has been the generally accepted observation that primary renal tubule cells undergo significant dedifferentiation and may not survive in conventional artificial culture. Modulation of the TGF-β signaling pathway is sufficient to enhance differentiation in renal proximal tubule cells subject to the stresses of artificial culture. It is indeed possible for cells to perform multiple specific differentiated functions characteristic of a healthy proximal tubule cell even at over 40 weeks in culture. Collectively, our work suggests that renal proximal tubule cells cultured under similar circumstances will be able to perform key functions for an implantable artificial kidney.
There remain significant challenges between this work and a practical clinical device. Primary among these challenges is the mass of cells required for clinical effect. Humes developed protocols that, depending on donor age, led to greatly expanded number of cells from donor tissue. 34 Conditional reprogramming of epithelial cells using feeder fibroblasts and rho-kinase inhibitors may provide a pathway for cell mass production without the need for potentially oncogenic immortalization. 35 Finally, in the future, biohybrid devices may provide an early clinical opportunity for organoid-derived cells.36,37
Bridging the in vitro-in vivo gap
We find that the dedifferentiation typically observed with cell culture is not intrinsic to the in vitro environment. Indeed, the uses to which cell culture has been put over the past seven decades have not previously asked that cells in culture bear quantitative fidelity to their in vivo counterparts. We seek to bridge the gap between the natural niche and the Petri plate without resorting to the intricacies of 3D culture systems, precisely because our biologic research and manufacturing infrastructures are geared to flat-sheet conditions. In this work, we have shown that control of specific signal transduction pathways is sufficient to reverse the erosion of at least some of the functional phenotypes associated with cell culture in conventional flat inelastic scaffolds for extended periods of time. This is an important enabling discovery for regenerative medicine and tissue engineering.
Footnotes
Authors' Contributions
K.H., J.A.L., R.C.E., and H.D.L. performed the experiments as well as editing the article for accuracy. R.C.H., R.Z., S.R., and M.H.W. provided guidance for experimental design and data interpretation as well as article review and editing. W.H.F. originated the experimental design, analyzed the data, and authored the main text of the article and is primarily responsible for the content.
Disclosure Statement
W.H.F. and S.R. are founders of Silicon Kidney LLC.
Funding Information
This work was supported by generous gifts from the Wildwood Foundation and the Maria and Bernard Pargh Family Foundation, as well as the Vanderbilt Center for Kidney Disease.