
Abstract
Macrophages are a primary contributor to the orchestration and severity of the foreign body response. As phagocytes and antigen-presenting cells, macrophages engage foreign objects, producing chemokines, degrading enzymes, and proinflammatory cytokines, including tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6). Encapsulated islet transplantation (EIT) is a return of function therapy in which donor insulin-secreting cells are encased in a biomaterial and implanted into a diabetic patient to regulate blood glucose levels. However, the foreign body response by macrophages to the encapsulated islet allograft may cause rejection. Recent studies have shown that substrate stiffness affects macrophage activity, which can inform EIT capsule design. However, due to the dysregulation of glucose maintenance in diabetic patients, varying from normoglycemic to hypoglycemic or hyperglycemic conditions, it is imperative to determine if glucose dysregulation affects macrophage mechanosensitivity to EIT biomaterials. This study explores the relationship between glucose metabolism and mechanosensitivity and the ultimate impact on proinflammatory macrophage function in static hyperglycemic and normoglycemic conditions. Using a 2-dimensional (2D) polyacrylamide model of 3-order magnitude in stiffness, 2, 15, and 274 kPa Young's moduli, the effect of glycemic condition on the mechanosensitive characteristics of unstimulated and proinflammatory RAW264.7 macrophage function in vitro using lipopolysaccharide (LPS) was examined. Hyperglycemic conditions were found to impact macrophage response to substrate stiffness significantly. Notably, TNF-α secretion was significantly reduced as substrate stiffness increased in LPS-stimulated hyperglycemic conditions, whereas normoglycemic macrophages held similar secretion across all stiffnesses. Stiffness-influenced differences in cytokine secretion were also induced in IL-6 secretion by hyperglycemic conditions. Hyperglycemic conditions promoted a biphasic trend in IL-6 cytokine secretion and gene expression by proinflammatory macrophages with significantly decreased production when cultured on 15 kPa compared to production on 2 and 274 kPa. Although hyperglycemic conditions drastically increased IL-10 secretion, stiffness-influenced differences were not shown when compared to the same glycemic condition. Furthermore, under LPS stimulation, lactate secretion had an inverse relationship to TNF-α secretion. However, no significant stiffness-influenced difference was demonstrated in glucose transporter 1 (GLUT1) expression, glucose uptake, or GAPDH. These findings suggest that hyperglycemic conditions enhance the mechanosensitivity of proinflammatory macrophages and should be explored further.
Impact statement
The work presented increases our understanding of the effect of glycemic condition on macrophage mechanosensitivity related to substrate stiffness. This has ramifications on the design of material-based therapies, such as encapsulated islet transplantation, for type 1 diabetic patients who experience glycemic dysregulation.
Introduction
Macrophages are innate immune cells and primary contributors to the foreign body response to tissue-engineered therapies. 1 As phagocytes and antigen-presenting cells, macrophages secrete degrading enzymes, chemokines, and proinflammatory cytokines, like tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), to protect the body from the invader. Due to this functionality, macrophages can hinder the effectiveness of tissue-engineered therapies, leading to immune rejection. One tissue-engineered therapy of interest is encapsulated islet transplantation (EIT). EIT is a tissue-engineered therapy that targets exogenous insulin dependence and regulates glucose levels in Type 1 diabetic patients. 2 This therapy consists of encapsulated whole donor islets in a semipermeable polymeric membrane, specifically the insulin-secreting β cells from islets.
The capsule protects the cells from leukocytes, while promoting islet function and viability. While researchers have seen success in diabetic animal models, EIT therapies are not a commercially relevant, long-term solution for insulin dependence due to rejection by the patient's immune system, which results in continued reliance on exogenous insulin. 3 Macrophages remain at the EIT site for the entire duration of the response, promoting intercellular signal cascades, which result in complete encapsulation of the transplant with fibrous tissue, named pericapsular fibrotic overgrowth (PFO). 4 PFO blocks the implanted islets from receiving oxygen, glucose, and nutrients, leading to loss of function and apoptosis. Altering the biochemical and mechanical characteristics of the capsule biomaterial may promote better viability for EIT.
This capsule can range in stiffness based on material and composition. However, the most common material used for EIT, alginate, can vary in stiffness from 1 to 100 kPa. 5 Although the role of mechanotransduction on the immune function of macrophages and other leukocytes is still being elucidated, recent studies have shown that macrophages are mechanosensitive. Substrate stiffness has been demonstrated to affect macrophage characteristics and function.6,7 Our laboratory has shown that substrate stiffness alters macrophage cell area, proliferation, actin organization, and migration speed when cultured on polyacrylamide gels with Young's moduli ranging from 1 to 280 kPa. 6
This was comparable to the findings of Blakney et al, 8 who found that substrate stiffness caused changes in gene expression of proinflammatory cytokines, TNF-α, IL-1β, and IL-6, by macrophages stimulated with lipopolysaccharide (LPS) when cultured on 130–840 kPa poly(ethylene glycol) hydrogels. Furthermore, Escolano et al found upregulated NLRP3 inflammasome formation, a multiprotein complex responsible for the final processing of IL-1β, and upregulated IL-1β and IL-6 secretion on 0.2 kPa polyacrylamide gels, compared to stiffer 33.1 kPa gels when stimulated with LPS and nigericin. 9 Although we can reasonably conclude that macrophages are mechanosensitive, we do not have a clear picture of the impact of glucose concentration on this phenomenon.
Dysregulation of glucose maintenance in diabetic patients causes blood glucose to vary from normoglycemic (70–120 mg/dL) to hypoglycemic (<70 mg/dL) or hyperglycemic (>120 mg/dL) conditions. It is understood that glucose dysregulation results in impaired immune function. Moreover, glucose metabolism plays an essential role in the function of macrophages.10,11 Long-term hyperglycemia has been demonstrated to promote increased TNF-α and IL-1β secretion. 12 The metabolic pathway begins with the uptake of extracellular glucose surrounding the cell through the cell's glucose transporter 1 (GLUT1), in the case of macrophages. 13
Increased glucose uptake due to GLUT1 overexpression resulted in increased reactive oxygen species production and proinflammatory phenotype in macrophages. 13 The glucose is then broken down through glycolysis by various enzymes to form two molecules of adenosine triphosphate (ATP) and pyruvate. 14 Lactate, a product of glycolysis, is produced by reducing pyruvate. Lactate is secreted in high amounts by innate immune cells after inflammatory stimulation and has been linked to hindering proinflammatory cytokine secretion.15,16 Furthermore, literature has demonstrated that inhibiting glycolysis using 2-deoxyglucose prevented macrophage activation in vitro and reduced inflammation in vivo.17,18
These findings suggest that glucose metabolism and mechanosensitivity may have a synergistic relationship that impacts macrophage function. However, it is imperative to determine if glucose dysregulation will affect the mechanosensitivity of macrophages as they respond to EIT biomaterials. Elucidating the role of mechanosensitivity on macrophage function in varied glycemic conditions can better inform EIT capsule design. This study explores the relationship between glucose metabolism and mechanosensitivity and its impact on macrophage function.
Using a 2D polyacrylamide gel model of 3-order magnitude in stiffness, we cultured unstimulated and LPS-stimulated RAW264.7 macrophages under normoglycemic (100 mg/dL) conditions and hyperglycemic (450 mg/dL) conditions on 2, 15, and 274 kPa polyacrylamide gels. Macrophages were cultured on the model for up to 3 days, and key aspects of glycolysis and LPS-stimulated mechanistic pathways were assessed. For glycolysis, samples were assayed for glucose uptake, lactate secretion, and gene expression of GLUT1 and the glycolysis enzyme GAPDH. Macrophages were also evaluated for TNF-α, IL-6, and IL-10 cytokine secretion and gene expression of toll-like receptor 4 (TLR4), a receptor for LPS stimulation, TNF-α, IL-6, and IL-10 (Fig. 1). Critically, this work demonstrates that hyperglycemia enhances mechanosensitivity in proinflammatory macrophages.
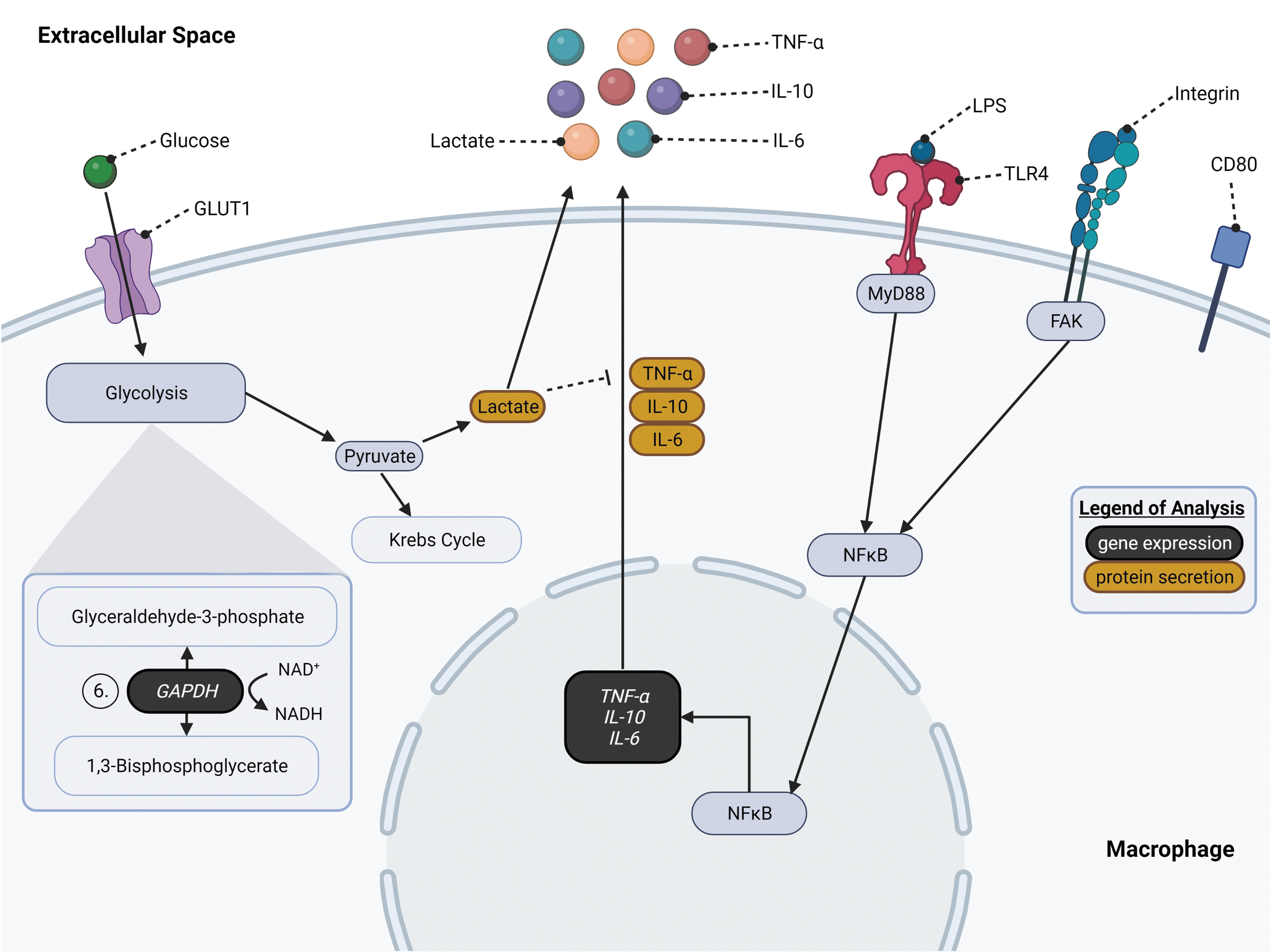
General overview of glycolysis, LPS/TLR4, and FAK signaling pathways in macrophages relevant to the cellular experimentation conducted in this study. On the left side, glucose enters the cell through GLUT1. 14 Once glucose uptake occurs, glucose undergoes glycolysis to break down the molecule into pyruvate and ATP. Pyruvate can later undergo lactic acid fermentation to form lactate, which is secreted from the macrophage. On the center-right side, LPS binds with TLR4 through LBP, MD-2, and CD14 (not shown) to form a complex leading to MyD88 (myeloid differentiation primary response gene 88)-dependent signal cascade. 29 Activation of MyD88 signals the activation and translocation of NFκB into the nucleus to activate the cytokine genes, TNF-α, IL-6, and IL-10. These cytokines are later secreted out of the cell. Under LPS stimulation, macrophages polarize to an inflammatory M1-like phenotype that can be signified by the surface marker CD80. 30 Furthermore, macrophages sense the mechanical properties, stiffness, of the ECM through the FAK pathway, which also leads to NFκB activation and translocation into the nucleus similar to MyD88-dependent signaling.31,32 This figure was illustrated using BioRender. ECM, extracellular matrix; FAK, focal adhesion kinase; GLUT1, glucose transporter 1; IL, interleukin; LPS, lipopolysaccharide; NFκB, nuclear factor kappa B; TNF-α, tumor necrosis factor-alpha.
Methods
Cell culture
Unstimulated RAW264.7 macrophages (ATCC, Manassas, VA) were cultured separately on tissue culture polystyrene based on glucose conditions before experimentation. Macrophages cultured under hyperglycemic conditions were cultured in 450 mg/dL Dulbecco's modified Eagle's media (DMEM) (ATCC). In contrast, cells in normoglycemic conditions were cultured in 100 mg/dL DMEM (Thermo Fisher Scientific, Waltham, MA). Ten percent fetal bovine serum was added to all culture media. Cells were seeded onto polyacrylamide gels of 2, 15, or 274 kPa in a six-well dish and allowed to adhere for 16–18 h at 37°C, 5% CO2. Afterward, gels with adhered cells were transferred into a new well with fresh media based on the condition to begin each study (Supplementary Fig. S1). Unstimulated adherence media samples were stored at −80°C until assayed.
Polyacrylamide synthesis and characterization
Polyacrylamide gels were synthesized according to previously established methods.6,19 Concentrations for each stiffness were as follows: 2 kPa–3% acryl and 0.1% bis, 15 kPa–8% acryl and 0.2% bis, and 274 kPa–15% acryl and 1.2% bis (Bio-Rad Laboratories, Hercules, CA). Ammonium persulfate (APS) (Thermo) and N, N, N’, N’-tetramethylethylenediamine (TEMED) (Bio-Rad Laboratories) were added to the acrylamide mixture. Gel substrates about 80 μm thick were synthesized by dispensing 30 μL of polyacrylamide on amine-activated glass coverslips (22 × 22 mm; Fisher Scientific, Pittsburg, PA) sandwiching between sterilized glass slides. Gels were cured at room temperature for 30 min and then pried from the glass slide using a sterile razor blade.
Gels were coated with 0.1 mg/mL human fibronectin (Sigma-Aldrich, St. Louis, MO) after activating with 0.5 mg/mL sulfo-SANPAH (Sigma-Aldrich) according to previously published methodology. 19 Gels were sterilized by exposure to ultraviolet light for 15 min and rinsed with sterile phosphate-buffered saline (Thermo Fisher Scientific) before cells were seeded for experimentation. To validate the stiffness of each gel, bulk polyacrylamide was synthesized using the same formulations previously outlined. Gels were sectioned into cylindrical columns using a 2:1 ratio of height to diameter as outlined by the ASTM standard for compressive testing of hydrogels.20,21 Bulk compression tests on the Instron confirmed Young's modulus of each gel (Table 1).
Young's Modulus Measurements Through Bulk Gel Compression Tests Based on Acrylamide and bis Concentrations
Cytokine secretion quantification
First, 105 RAW264.7 macrophages were seeded onto each fibronectin-coated gel to adhere for 16–18 h at 37°C, 5% CO2. After macrophages were transferred into a new well, 3 mL of hyperglycemic or normoglycemic media, based on the original glycemic condition used for culture, was added to each well. For stimulated samples, 0.5 μg/mL Salmonella typhosa LPS (Sigma-Aldrich) was included in the media. Samples were incubated at 37°C, 5% CO2, for 3 days. Supernatants were collected from each sample after each day and stored at −80°C until assayed. TNF-α, IL-10, and IL-6 were quantified using sandwich enzyme-linked immunoassay (TNF-α and IL-6: Peprotech, Inc., East Windsor, NJ; IL-10: Thermo Fisher Scientific) with protocols from their respective kits. Three biological replicates were conducted for each sample.
Proliferation of unstimulated RAW264.7 macrophages
105 RAW264.7 macrophages were seeded onto 2 and 274 kPa fibronectin-coated gels under hyperglycemic and normoglycemic conditions to adhere for 16–18 h at 37°C, 5% CO2. Gels with adhered macrophages are then transferred into a new well imaged at time points 0, 24, and 48 h. Five technical replicates were imaged for each biological sample. Three biological samples were conducted.
Real-time quantitative polymerase chain reaction
To quantify gene expression, 2 × 105 macrophages were seeded on each gel and adhered for 16–18 h. Adhered cells were transferred into a new well to begin experimentation. Fresh media were added based on glycemic condition, and cells were stimulated with 0.5 μg/mL LPS for 4 h at 37°C, 5% CO2. Macrophages were harvested for gene expression analysis using StemPro™ Accutase™ cell dissociation reagent (Thermo Fisher Scientific) for 30 min at room temperature. Cells were collected into pools by sample (n = 6 gels for each sample; ∼1.2 × 106 cells total), and cell pellets were generated by centrifugation at 1000 RPM for 10 min.
The supernatant was aspirated, and TRIZOL and chloroform reagents were added to the remaining pellet to begin RNA isolation. RNA was extracted using RNeasy Plus Mini Kit (Qiagen) according to the manufacturer's instructions. RNA concentration and purity were quantified using Nanodrop 2000 (Thermo Fisher Scientific). Subsequently, using a high-capacity cDNA reverse transcriptase kit (Thermo Fisher Scientific), RNA was reverse transcribed to cDNA. TaqMan probes and TaqMan™ Universal polymerase chain reaction (PCR) Master Mix were added to the cDNA and loaded to a 384-well plate.
TaqMan probes used are as follows: TNF-α–Mm00443258_m1, IL-10–Mm00439614_m1, IL-6–Mm00446190_m1, TLR4–Mm00445273_m1, GLUT1–Mm00441473_m1, GAPDH–Mm99999915_g1, and 18S ribosomal RNA (18srRNA)–Mm03928990_g1 (Thermo Fisher Scientific). PCR plates were run on a QuantStudio 7 Flex Real-Time qPCR system (Thermo Fisher Scientific). 18srRNA was used as a reference gene, and all samples were normalized to 100 mg/dL 2 kPa for comparison. Samples were compared to each other using the fold change of gene expression calculated using the 2−ΔΔCT method. Three biological replicates were conducted for each sample.
Western blotting
Protein expression of TLR4 was examined using the same culture method as section 2.5, 2 × 105 macrophages were seeded on each gel and adhered for 16–18 h. Adhered cells were transferred into a new well to begin experimentation. Fresh media were added based on glycemic condition, and cells were stimulated with 0.5 μg/mL LPS for 4 h at 37°C, 5% CO2. Protein isolation buffer [RIPA Buffer (Thermo Fisher, Waltham, MA)], 1:100 protease inhibitor cocktail (Sigma Aldrich, St. Louis, MO), and 20 μg/mL phenylmethylsulfonyl fluoride (PMSF) were added to inverted polyacrylamide gels in a six-well plate and was incubated on ice for 5 min.
Following incubation, cell lysate was scraped from gels using a cell scrapper and collected. Cell lysates were further incubated on ice for 1 h, with rigorous vortexing every 15 min, after which, the tubes were spun down at 1000 g for 5 min and the protein solution lysate was collected. To analyze protein content, a bicinchoninic acid (BCA) assay was run on the protein solution lysate in accordance with Pierce BCA Protein Assay Kit protocol (Thermo Fisher).
The loading weight of protein was extrapolated from absorbance readouts calibrated to a standard curve. Samples were diluted to a loading weight of 10 μg. Diluted protein samples were incubated with 4 × Laemmli buffer (Bio-Rad) and β-mercaptoethanol (Bio-Rad) for 10 min at 100°C. Following a brief microcentrifugation, protein solutions in Laemmli buffer were loaded into a precast Mini-PROTEAN TGX Gel (Bio-Rad) with a Precision Plus Dual Color ladder (Bio-Rad) in an empty well for reference. The gel was run at 120V for 1 h using the Bio-Rad PowerPac HQ in a Tris/Glycine/SDS Buffer (Bio-Rad). Before membrane transfer in a Bio-Rad Trans-Blot Turbo Transfer System, supported Nitrocellulose Membrane (Bio-Rad) and two transfer spongers were pre-soaked in Trans-Blot Turbo Transfer Buffer (Bio-Rad) for 10 min.
After transfer from TGX gel to membrane, the membrane was blocked in 1:1 phosphate-buffered saline (PBS): Intercept Blocking Buffer (Li-Cor, Lincoln, NE) for 1 h at RT. Membranes were subsequently incubated with 2 μg/mL TLR4 polyclonal antibody (Thermo Fisher) and 1:500 β-actin monoclonal antibody (BD, Franklin Lakes, NJ) for 16–18 h at 4°C. The following day, the membrane was washed 3 × with PBS with 0.1% Tween 20 (PBST) for 15 min each. The membrane was then treated with 1:10,000 Li-Cor IR-Dyes (Li-Cor) in 1:1 PBST: Intercept Blocking Buffer for 1 h at room temperature. The membrane was washed 3 × with PBS for 15 min each. The membrane's fluorescent readout was captured with a Li-Cor Odyssey CLx (Li-Cor) imager and analyzed using Li-Cor ImageStuidio Version 5 software.
Immunofluorescence imaging and quantification
After culturing RAW264.7 macrophages for 3 days, samples were fixed with 4% PFA followed by 1 × PBS washes before blocking with 4% BSA/PBS. The primary antibody (CD80), used to indicate the proinflammatory (M1) phenotype, was incubated for 16–18 h at 4°C. Samples were then washed with 1 × PBS. Secondary antibody (FITC) in 4% BSA/PBS was added to samples for a 1-h incubation at room temperature in the dark. DAPI was used to counterstain the cell nucleus. ImageJ software was used to determine the fluorescence intensity of each cell. The average CD80+ expression of each sample was quantified using the mean intensity of the FITC channel divided by the number of nuclei per image (Equation 1). Five technical replicates were done for each biological replicate. Three biological replicates were conducted for each sample.
Glucose uptake and lactate secretion
After 105 RAW264.7 macrophages adhered to fibronectin-activated polyacrylamide gels overnight and gels were transferred into a new well, fresh media were added based on glycemic condition. Stimulated cells were treated with 0.5 μg/mL LPS. Media samples were aliquoted each day for 3 days and stored at −20°C until analyzed. Glucose and lactate concentrations of samples are quantified using a YSI 2950D Biochemistry Analyzer (YSI, Yellow Springs, OH). The rate of glucose uptake and lactate secretion was determined by glucose and lactate concentration in the media over 3 days.
Statistical analysis
Statistical tests were conducted using GraphPad Prism 9 (GraphPad, San Diego, CA). Normality was verified for data groups using Shapiro-Wilks tests with n = 3 biological replicates. For immunofluorescence imaging, n = 5 technical replicates were conducted, while n = 3 technical replicates were performed for all other assays. Once completed, data groups were analyzed using analysis of variance and between pairs using Tukey tests. Statistical significance was indicated by p < 0.05.
Results
Macrophage cytokine secretion is impacted by substrate stiffness and glycemic condition when stimulated with LPS
To determine if there is a synergistic relationship between mechanosensitivity and glucose metabolism on macrophage function, we first investigated proinflammatory cytokine secretion. Seeding RAW264.7 macrophages, cultured under normoglycemic (100 mg/dL glucose) and hyperglycemic (450 mg/dL glucose) conditions, onto fibronectin-coated polyacrylamide gels of 2, 15, and 274 kPa, we examined cytokine secretion of TNF-α, IL-10, and IL-6 when LPS stimulated and unstimulated over 3 days.
Mechanosensitivity trends in TNF-α production changed based on glucose concentration and stimulation conditions. Under LPS stimulation, TNF-α production was inversely proportionate with stiffness, where increased stiffness significantly reduced production of TNF-α by the macrophages when under hyperglycemic conditions (450 mg/dL) after 3 days of stimulation (Fig. 2A). Interestingly, this inverse proportionality was not present in the normoglycemic condition (100 mg/dL) under LPS stimulation. In fact, this linear trend was only demonstrated again in unstimulated 100 mg/dL conditions (Fig. 2B).

TNF-α production by macrophages was inversely proportionate to increased stiffness where mechanosensitive.
When TNF-α production was compared based on the glycemic condition under the same stiffness, LPS-stimulated macrophages on 2 kPa substrates held the most significant variance from normoglycemic to hyperglycemic condition on day 3 (Fig. 2C). As substrate stiffness increased, the variance between TNF-α production of 450 mg/dL samples and 100 mg/dL samples became comparable. This phenomenon was only seen when cells were stimulated, with TNF-α production of unstimulated cells with comparable secretion between 450 and 100 mg/dL across all stiffnesses (Supplementary Fig. S2A).
IL-10 displayed no mechanosensitivity trend under hyperglycemic or normoglycemic conditions. IL-10 production was also comparable across all stiffnesses for stimulated (Fig. 3A) and unstimulated samples (Supplementary Fig. S3A). However, when evaluated by glycemic condition on the same stiffness, similar to TNF-α, 2 kPa samples under LPS stimulation held a significantly (p < 0.0005) higher upregulation than 15 and 274 kPa (Fig. 3C). While there is no distinctive trend in IL-10 production through comparing stiffness under the same glycemic condition, hyperglycemic conditions drastically increased IL-10 production by LPS-stimulated macrophages, but not in unstimulated macrophages (Supplementary Fig. S2B and Supplementary Fig. S3A).
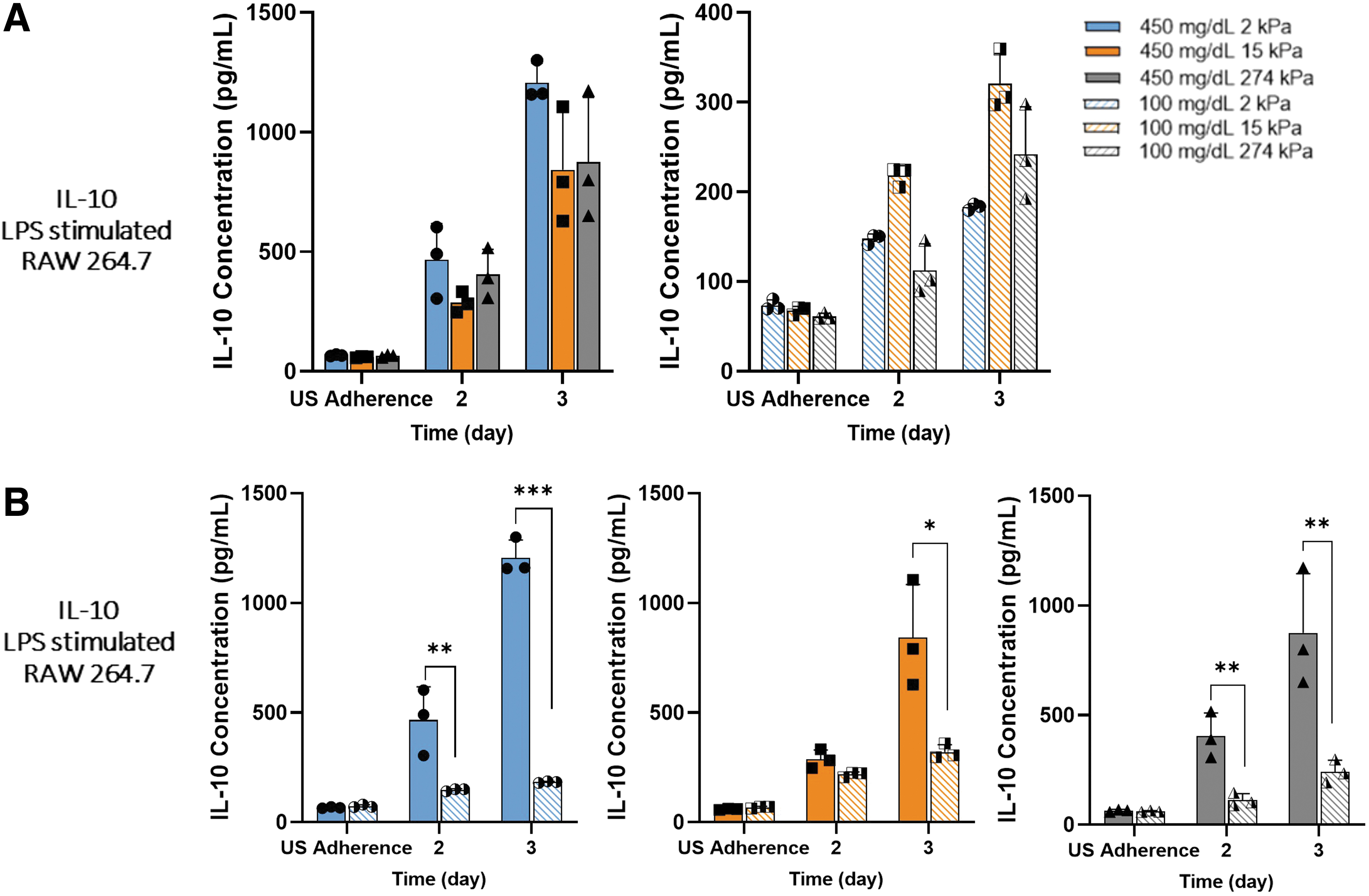
Hyperglycemia upregulated IL-10 secretion by proinflammatory macrophages most significantly on 2 kPa PA gels.
Furthermore, glucose metabolism affected the mechanosensitivity of the macrophages with IL-6 production. Mechanosensitivity differences in IL-6 production were only witnessed once stimulated under 450 mg/dL (Fig. 4A). A biphasic trend was demonstrated as substrate stiffness increased in 450 mg/dL LPS samples, in which IL-6 secretion was reduced on 15 kPa, while 2 and 274 kPa held comparable secretion levels. Hyperglycemic samples also had drastically higher IL-6 secretion than normoglycemic samples, with a similar biphasic trend as stiffness increased (Fig. 4B).
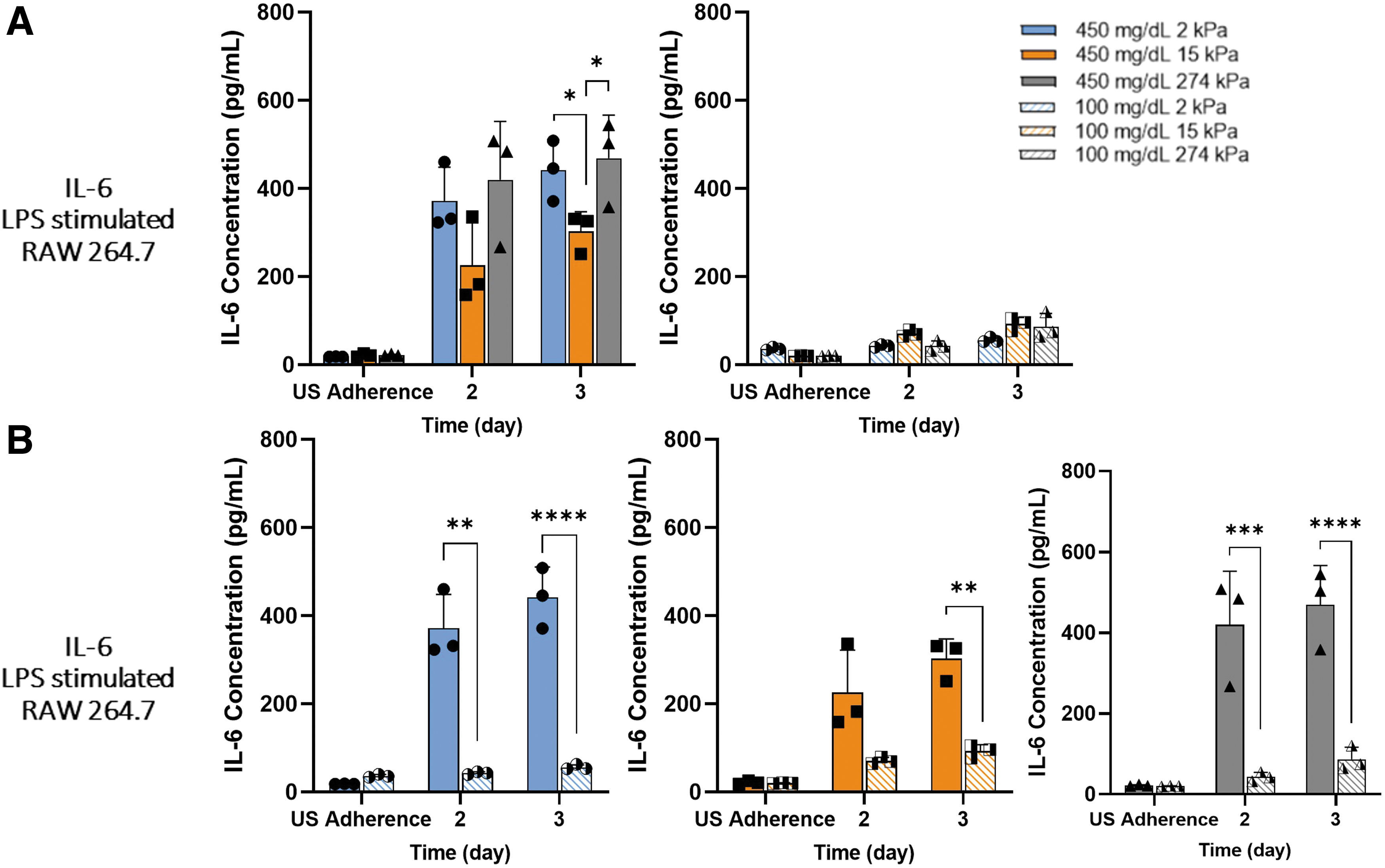
Hyperglycemia promoted biphasic mechanosensitivity trend in IL-6 secretion by LPS-stimulated macrophages.
Unstimulated macrophages produced low levels of IL-6 across all stiffness and glycemic conditions (Supplementary Fig. S2C, and S3B). A proliferation study was conducted comparing macrophages cultured on 2 and 274 kPa gels under both glycemic conditions to confirm if cell number was the cause of these findings. While there was a slight increase in cell number based on glucose, this was not significant and would not account for our findings (Supplementary Fig. S4). These results suggest hyperglycemia influences macrophage mechanosensitivity to different substrate stiffnesses through cytokine production.
Macrophage phenotype changes with response to substrate stiffness and glycemic condition
To further investigate the effect of glycemic condition and substrate stiffness on macrophage function, we assess how macrophages polarized to their proinflammatory, M1 phenotype when stimulated using the CD80 surface marker. LPS stimulation promoted enhanced M1 polarization under both glycemic conditions as expected (Supplementary Fig. S5). Four hundred fifty milligrams per deciliter samples demonstrated comparable CD80+ macrophages when stimulated across all stiffnesses (Fig. 5A). LPS-stimulated 100 mg/dL macrophages, however, held a significant decline that was seen in CD80+ cells from 2 to 274 kPa (Fig. 5B). With these results, it can be suggested that glucose can alter the polarization of macrophages in response to substrate stiffness and LPS stimulation.
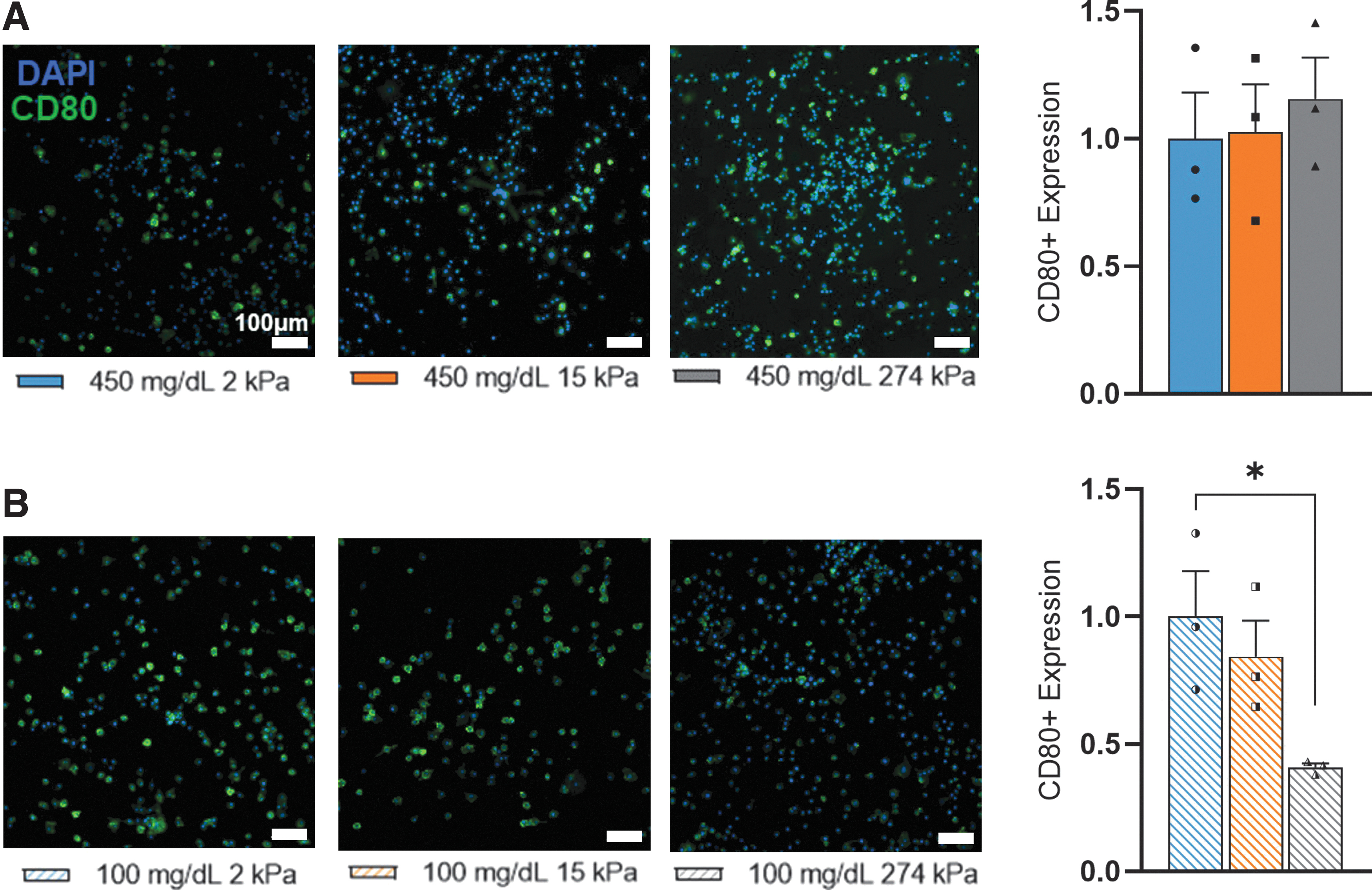
Glycemic condition altered phenotypic response to substrate stiffness by proinflammatory macrophages.
Hyperglycemic conditions and substrate stiffness altered cytokine gene expression in the TLR4/LPS signaling pathway of LPS-stimulated macrophages
To continue our investigation of the effect of glycemic condition and substrate stiffness on proinflammatory markers, we examined the TLR4 signaling pathway (Fig. 1). TLR4 held no change in gene or protein expression based on glucose or stiffness (Fig. 6A). Examining downstream, only IL-6 held similar trends in relative gene expression to protein secretion. No stiffness-influenced trends were demonstrated in TNF-α (Fig. 6B). IL-6 held a significant upregulation in gene expression dependent on glycemic condition, with 450 mg/dL samples displaying significantly higher relative gene expression than 100 mg/dL, as seen in protein secretion data (Fig. 6C).
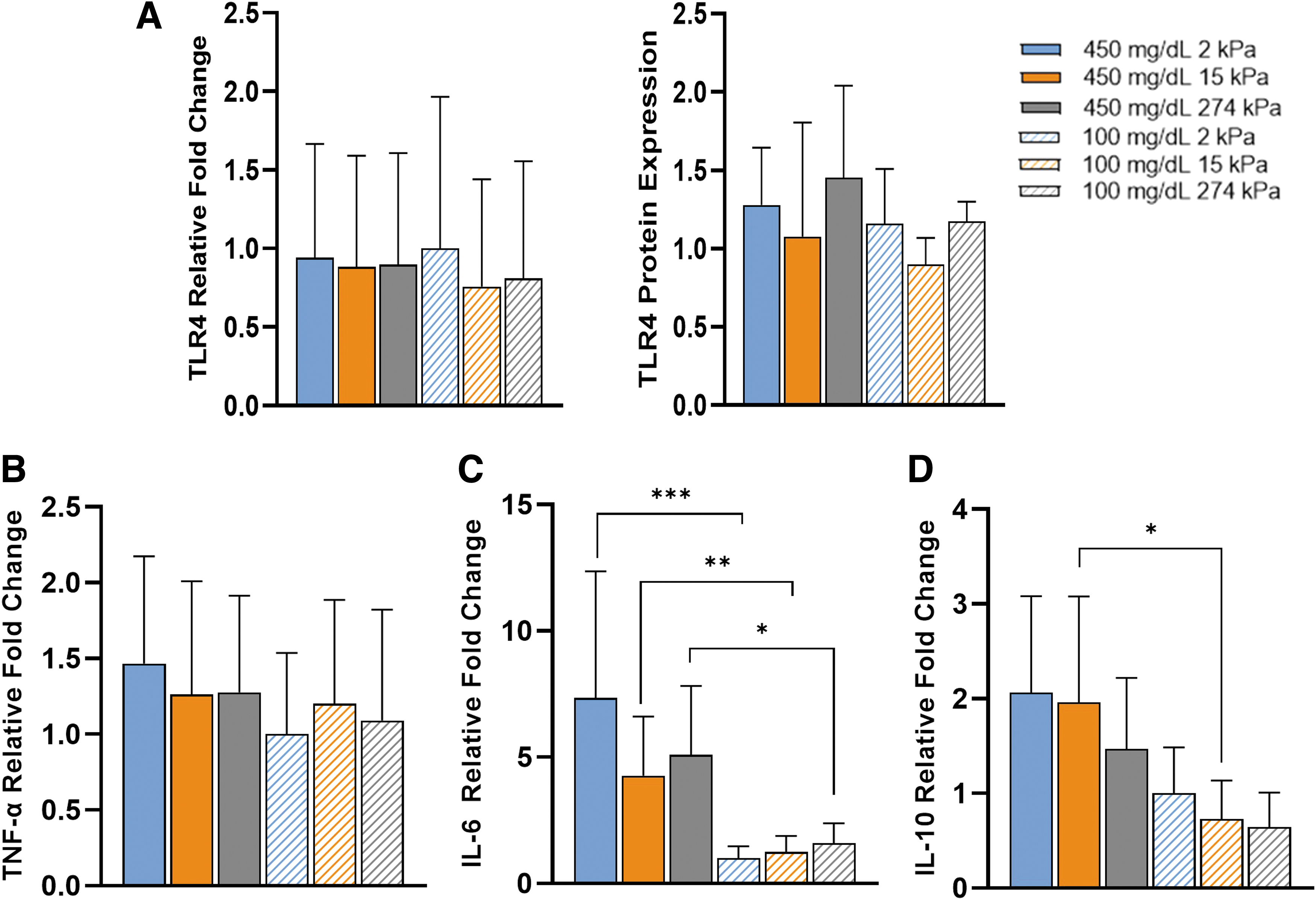
Hyperglycemia upregulated gene expression and mechanosensitivity downstream in TLR4/LPS signaling pathway of proinflammatory macrophages. Comparison of gene and protein expression of
Mechanosensitivity trends were also similar to protein secretion, although not statistically significant. Glycemic condition and substrate stiffness did alter IL-10 gene expression (Fig. 6D). Glucose affected IL-10 gene expression more than substrate stiffness, with significant upregulation in IL-10 expression between 100 and 450 mg/dL on 15 kPa substrates, similar to protein secretion. These results suggest glycemic condition and substrate stiffness alter how macrophages process LPS, especially in IL-10 and IL-6 expression.
Lactate secretion rates by LPS-stimulated macrophages are proportionate with substrate stiffness increases in hyperglycemic conditions only
Key aspects of the glycolysis pathway were investigated to determine the effect of substrate stiffness on the glucose metabolism under normoglycemic and hyperglycemic conditions. GLUT1 held no statistically significant change in relative mean expression by glycemic condition or stiffness (Fig. 7A). However, glucose uptake rates were significantly increased across all stiffnesses of 450 mg/dL compared to 100 mg/dL, in both LPS stimulated and unstimulated (Fig. 7B). Moving downstream, GAPDH held no trend in gene expression based on glucose and stiffness (Fig. 7C). Finally, lactate secretion increased with stiffness and glucose only when stimulated (Fig. 7D). Under hyperglycemic conditions, lactate secretion rates trended upward with rising stiffness and significantly increased compared to normoglycemic samples. These results indicate that mechanosensitivity in glycolysis is occurring downstream in the signaling pathway, but only in hyperglycemic proinflammatory macrophages.
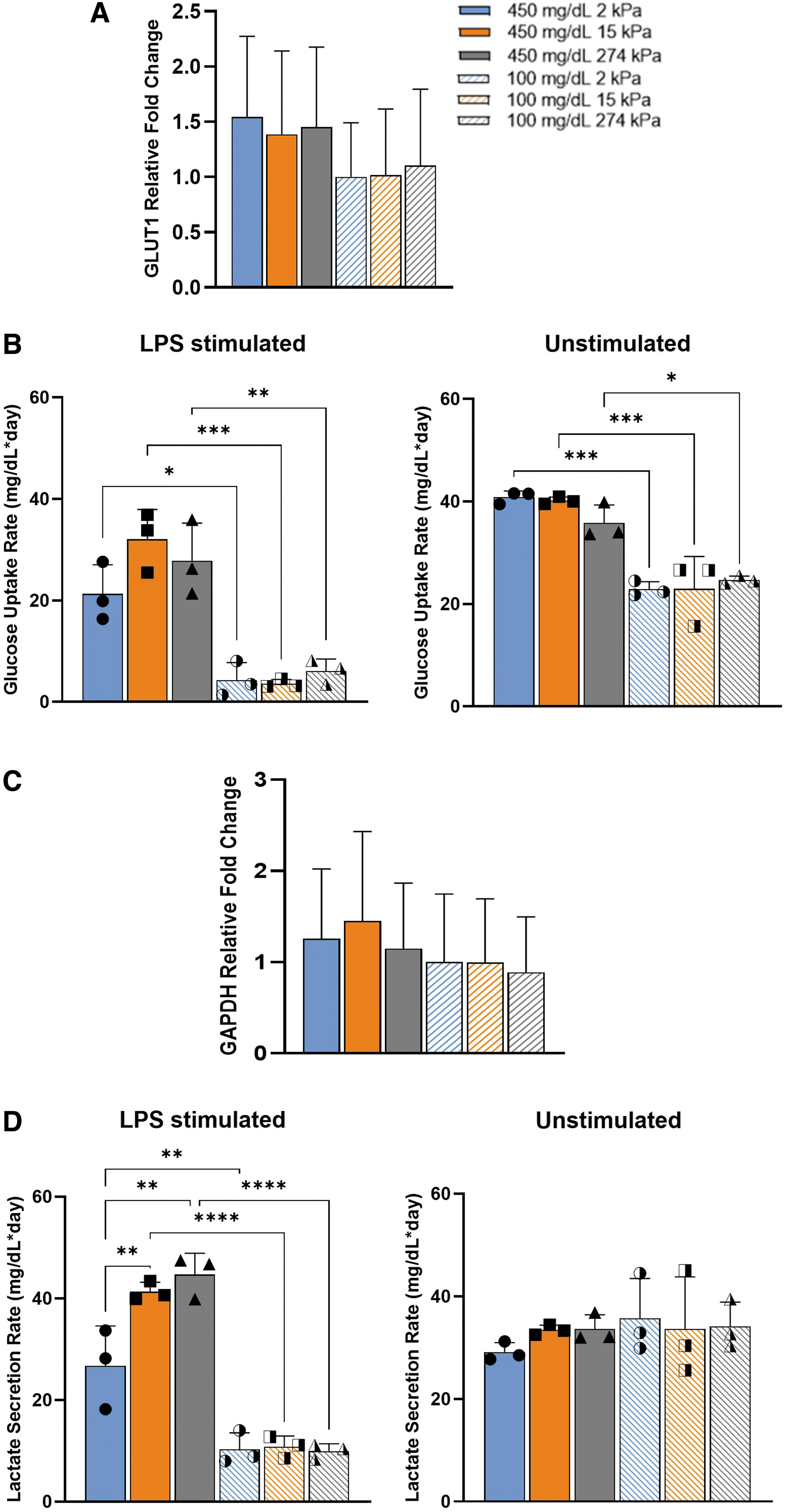
Hyperglycemia promoted mechanosensitivity in downstream glycolysis by LPS-stimulated macrophages indicated by lactate secretion. Comparison of
Discussion
In this study, we sought to determine the effects of glucose dysregulation on the mechanosensitivity seen in proinflammatory macrophage function. Using a 2D polyacrylamide model of 3-order magnitude stiffness to mimic 1–100 kPa EIT stiffness range, our results indicate that there is a synergistic effect between glycemic condition and substrate stiffness on proinflammatory macrophage function: hyperglycemic conditions enhance mechanosensitivity in proinflammatory macrophages. To our knowledge, this is the first study of its kind addressing the link between glycemic condition to mechanosensitivity in macrophage function. As such, we can only compare findings to studies that investigated glucose metabolism and substrate stiffness effects on macrophage function separately.
Although RAW264.7 cells were used in our study, hyperglycemic conditions triggered distinctive mechanosensitivity trends by substrate stiffness in cytokine secretion under LPS stimulation, as seen in literature under various murine and human cell lines and murine primary cells. More specifically, TNF-α production was inversely proportionate with increased stiffness in hyperglycemic conditions; this aligns with previous findings by Blakney et al, 8 Patel et al, 22 and Escolano et al 9
Similar to our study, Blakney et al used a higher range of substrate stiffnesses (130–840 kPa) to demonstrate a reduction in TNF-α production with increased stiffness in LPS-stimulated RAW264.7 and bone marrow-derived macrophages compared to unstimulated macrophages. 8 However, our study showed mechanosensitivity trends of unstimulated macrophages in normoglycemic conditions for TNF-α production, but not in hyperglycemic conditions as seen by Blakney et al 8 and Patel et al 22 Interestingly, the phenomenon only occurred for TNF-α secretion, while IL-10 and IL-6 secretion remained comparable when unstimulated. IL-10 did not show mechanosensitivity trends when compared under the same glycemic conditions for both unstimulated and LPS-stimulated macrophages.
However, as glucose concentration changed from normoglycemic to hyperglycemic conditions on the same stiffness, IL-10 production on 2 kPa expressed the most significant increase in secretion when stimulated with reduced significance as stiffness increased. Moreover, hyperglycemia promoted mechanosensitivity in IL-6 secretion in LPS-stimulated macrophages, although this was not the case in normoglycemic conditions. In fact, macrophages did not display any significant secretion difference based on stiffness for any of the examined cytokines in normoglycemic conditions in the presence of LPS. Overall, this suggests that glycemic condition is a driving factor in mechanosensitive response to substrate stiffness for cytokine secretion by proinflammatory macrophages.
We further explored the TLR4/LPS signaling pathway for gene expression of the TNF-α, IL-6, and IL-10, as well as TLR4. Mean relative gene expression of the corresponding cytokines that were assessed mimicked mechanosensitivity and glucose trends highlighted in cytokine secretion. Our hyperglycemic TNF-α and IL-10 gene expression findings did reflect work by Blakney et al 8 and Chuang et al 23 Interestingly, IL-6 gene expression was most distinctive by glycemic condition with a significant upregulation in expression in hyperglycemic conditions compared to normoglycemic conditions—further highlighting the synergy between glucose in mechanosensitivity and proinflammatory macrophage function.
Contrarily, Previtera et al 24 found TLR4 to be influenced by substrate stiffness with upregulated expression in response to increased stiffness. Our results did not mirror this previous finding, which may be due to differences in cell type, as their study was conducted with primary murine bone marrow macrophages. However, our findings do suggest that the mechanosensitivity is occurring downstream in the TLR4/LPS signaling pathway and glycemic condition affects this phenomenon.
Macrophage phenotype has been tied to cytokine secretion, with M1 macrophages secreting upregulated levels of TNF-α, IL-6, and IL-12, while M2 macrophages secrete more IL-4 and IL-10. 25 Okamato et al found that stiff gels attenuated proinflammatory response by macrophages after stimulation. 26 Chen et al further illustrated this phenomena, displayed downregulated CD86, another M1 marker, expression as stiffness increased corresponding with an upregulated CD206, an M2 marker, expression in bone marrow-derived macrophages. 27 While we did observe this effect through quantifying immunofluorescence imaging, M1 macrophage polarization was only inversely proportionate to increased stiffness in normoglycemic conditions with no mechanosensitivity trends seen in hyperglycemic conditions.
This was an unexpected finding as the opposite trend is expected, with distinct mechanosensitivity trends in the hyperglycemic condition and insignificant stiffness effects in normoglycemic conditions to match both our cytokine results. This may be specific to the surface marker CD80 and other proinflammatory and anti-inflammatory surface markers in conjunction will need to be assessed to determine mechanosensitivity trends by substrate stiffness on macrophage phenotype. However, these results further elucidate the effect of glycemic condition on macrophage response to substrate stiffness.
Glucose is a critical mediator to immune function, reviewed in detail by O'Neill et al 11 Further to this point, Gonzalez et al displayed that hyperglycemic conditions induced proinflammatory cytokine production in primary human monocytes, upregulating IL-6 and TNF-α. 28 Freemerman et al demonstrated a similar conclusion by overexpressing GLUT1 in RAW264.7 macrophages to increase glucose uptake. Due to this, it was important to determine if components of the glycolysis pathway are mechanosensitive. It was expected that hyperglycemic conditions caused an increase in GLUT1 expression. Our findings showed GLUT1 to be impacted more by glycemic condition than substrate stiffness, however, not statistically significant.
Also, glucose uptake and GAPDH gene expression showed similar results to GLUT1 expression. Interestingly, lactate secretion in hyperglycemic LPS-stimulated macrophages increased proportionately to stiffness opposite of TNF-α data under the same parameters. This demonstrates the role of lactate as an inhibitor of TNF-α secretion, elucidated by Ratter et al 16 However, this trend was only displayed in hyperglycemic LPS-stimulated macrophages. These findings highlight mechanosensitivity in downstream glycolysis genes in stimulated macrophages that should be further studied.
While this study used a well-studied and characterized cell population and gel model, it is fair to note that this study is limited in the following ways: this study was conducted in vitro under static glycemic conditions, which does not fully represent the fluctuation in glycemic conditions experienced by patients with diabetes. Also, while this work could be compared to mechanobiology and glucose metabolism studies using various murine and human macrophage lines and primary bone marrow-derived macrophages, the phenomenon of glycemic dysregulation on stiffness-influenced function has not been investigated. Additional studies using primary cells or a human cell line and in vivo will be needed to apply these conclusions to capsule design for EIT.
Conclusion
In this study, we were able to successfully demonstrate that glycemic condition can influence mechanosensitivity to substrate stiffness in macrophage function. This work suggests hyperglycemic conditions enhance mechanosensitivity in proinflammatory macrophage function, specifically in downstream proinflammatory gene expression, cytokine secretion, and lactate secretion. Additional studies are needed to further elucidate this phenomenon to encompass the full range of macrophage functions. In future studies, downstream TLR4 signaling events such as MyD88 expression, nuclear factor kappa B (NFκB) phosphorylation, and NFκB nuclear translocation, as well as morphology and phagocytosis, will be evaluated to clarify the effect of substrate stiffness and glycemic condition on proinflammatory macrophage function. We aim to increase our understanding of this phenomenon to be used as a design parameter for EIT and other implant therapeutics for diabetic patients.
Footnotes
Authors' Contributions
The first author, C.J., conceptualized the research, designed and conducted experimentation, prepared the article, and generated the figures. D.F. conducted glucose uptake and lactate secretion experiments. I.M.S. conducted the TLR4 Western blot. H.A.E. and J.P.F. oversaw the project, assisted in experimental development, and reviewed the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
We want to thank the A. James Clark School of Engineering, NIBIB/NIH Center for Engineering Complex Tissues (P41EB023833), NSF (CBET1856350), and NSF (1507730) for funding this project.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.