
Abstract
Skeletal muscle possesses adaptability to mechanical loading and regenerative potential following muscle injury due to muscle stem cell activity. So far, it is known that muscle stem cell activity is supported by the roles of several interstitial cells within skeletal muscle in response to muscle damage. The adjacent tendon is also exposed to repetitive mechanical loading and possesses plasticity like skeletal muscle. However, the interplay between the skeletal muscle and adjacent tendon tissue has not been fully investigated. In this study, we tested whether factors released by three-dimensional engineered human tendon constructs in response to uniaxial tensile loading can stimulate the proliferation and differentiation of human-derived myogenic cells (myoblasts). Tendon constructs were subjected to repetitive mechanical loading (4% strain at 0.5 Hz for 4 h) and nonrepetitive loading (0% strain at 0 Hz for 4 h), and the conditioned media from mechanically loaded and nonmechanically loaded control constructs were applied to myoblasts. Immunofluorescence analysis revealed both an increase of myotube fusion index (≥5 nuclei within one desmin+ myotube) and the myotube diameter when conditioned medium from mechanically loaded tendon constructs was applied. Myostatin, myosin heavy chain 7, and AXIN2 gene expressions were downregulated in myotubes treated with conditioned medium from mechanically loaded tendon constructs. However, proliferative potential (number of Ki67+ and bromodeoxyuridine+ myoblasts) did not differ between the two groups. These results indicate that tendon fibroblasts enhance myotube formation by mechanical loading-induced factors. Our finding suggests that mechanical loading affects the signaling interplay between skeletal muscle and tendon tissue and is thus important for musculoskeletal tissue development and regeneration in humans.
Impact statement
The interplay between satellite cells and various types of resident cells within the skeletal muscle for muscle regeneration has been extensively studied. However, even though tendon tissue is located adjacent to skeletal muscle tissue and cells in these tissues are exposed to repetitive mechanical loading together, the interaction between muscle and tendon tissues for muscle regeneration remains to be elucidated. In this study, we report that the conditioned media from engineered human tendon tissues undergoing repetitive tensile mechanical loading enhanced myotube formation. Our in vitro findings extend the fundamental understanding of the crosstalk between adjacent tissues of the muscle–tendon unit.
Introduction
Resident muscle stem cells, called satellite cells, possess roles for skeletal muscle maintenance, postnatal muscle growth, hypertrophy, and repair after muscle injury. 1 Satellite cells are normally quiescent, however, they are rapidly activated and enter the cell cycle to become myoblasts in response to muscle injury, giving rise to progeny, and fuse with each other and/or with host myofibers to regenerate muscle.2–4 These regenerative processes of satellite cells are regulated by several intrinsic cell networks and various extrinsic types of factors, 5 and any disorganization may result in muscle dysfunction and affect severity of muscle diseases. 6
The underlying mechanism of interplay between satellite cells and various types of resident cells within the skeletal muscle for muscle regeneration has been extensively studied, such as macrophages,7,8 endothelial cells,9,10 mesenchymal progenitors, 11 and myofibers.1,12,13 In particular, the importance of resident fibroblasts in the muscle tissue for muscle regeneration has been reported in both animals14,15 and humans.16,17 Interestingly, tenogenic fibroblasts expressing scleraxis (Scx), a transcription factor of tendon fibroblasts, 18 have been discovered in adult skeletal muscle. 15 These cells support the muscle regeneration through extracellular matrix (ECM) niche remodeling, 15 suggesting that tendon fibroblasts may contribute to muscle regeneration. However, even though tendon tissue is located adjacent to skeletal muscle tissue, the interaction between muscle and tendon tissues for muscle regeneration remains to be elucidated.
A small number of studies have reported the relationships between muscle and tendon tissues on the physiological aspects of muscle development and homeostasis in vivo. Mice deficient in the tenogenic genes exhibit loss of muscle mass and diminished exercise capacity.19,20 Just as there is influence from muscle to tendon, injury of the gastrocnemius muscle induces ECM remodeling of the calcaneal tendon and alters the subsequent functional properties, 21 and proinflammatory cytokines released from muscle decrease cell viability in the tendon. 22 These studies indicate that tendon and muscle affect the homeostasis of each other's tissues. In an in vitro study, we have shown that tendon fibroblasts enhanced differentiation of myoblasts using a coculture model, 23 indicating that tendon fibroblasts release factors that enhanced myogenic progression in the absence of mechanical stimuli. In addition, it has been shown that tenogenic responses in tendon cells are upregulated by mechanical loading or exercise through cellular mechanotransduction in tissue-engineered tendons.24–28 Furthermore, muscle and tendon tissues are exposed to mechanical loading together. 27 Therefore, this evidence led us to hypothesize that the potential of tendon fibroblasts for enhancing myogenic progression would be increased by mechanical stimuli.
In this study, we have used tissue-engineered tendon constructs subjected to 4% strain at 0.5 Hz for 4 h, conditions that resulted in increased SCX gene expression, which is a typical tenogenic gene in human tendon fibroblasts. The conditioned medium from either mechanically loaded or nonmechanically loaded tendon construct was applied to human myoblasts, and myogenic progression in response to mechanical loading of tendon constructs was evaluated by immunofluorescence staining and gene expression analysis.
Materials and Methods
Ethics approval
Written and informed consent for donation of excess reconstructive tissues (semitendinosus and gracilis tendon–muscle tissues) was obtained from seven healthy males undergoing anterior cruciate ligament reconstruction (age = 30 ± 10 years [mean ± standard deviation {SD}]). All experiments for the present human study were approved by the Ethics Committee of Copenhagen (Ref. H-3-2010-070) and performed in accordance with the Helsinki Declaration II.
Cell isolation and tissue culture
Myoblasts and tendon fibroblasts were isolated from the same muscle–tendon tissue unit of each donor (Fig. 1A). Muscle tissue was minced into small pieces (<0.1 cm3) after eliminating all visible nonmuscle tissue (e.g., fat, loose connective tissue), and was digested for 1 h with 0.2% type B collagenase (Roche Sigma-Aldrich, Darmstadt, Germany; 11088815001) and 0.2% dispase 170 II (Roche Sigma-Aldrich; D4693-1G) in skeletal muscle basal medium (PromoCell, Heidelberg, Germany; C-23260) at 37°C, in 5% CO2. The muscle pieces were triturated every 15 min and then filtrated through a 100 μm cell strainer (BD Falcon, NJ, USA; 352360). After washing with human skeletal muscle growth medium (mGM: myoblasts growth medium) (PromoCell; C-23060) supplemented with 10% of skeletal muscle growth medium supplement mix (PromoCell; C-39365), 15% fetal bovine serum (FBS) (BioWest, MA, USA; S181H-500), and 1% L-glutamine penicillin–streptomycin (PS) solution (Roche Sigma-Aldrich; G6784), the filtered myoblasts were cultured in T25 flasks until there were sufficient cell numbers (≤80% confluent) in mGM.
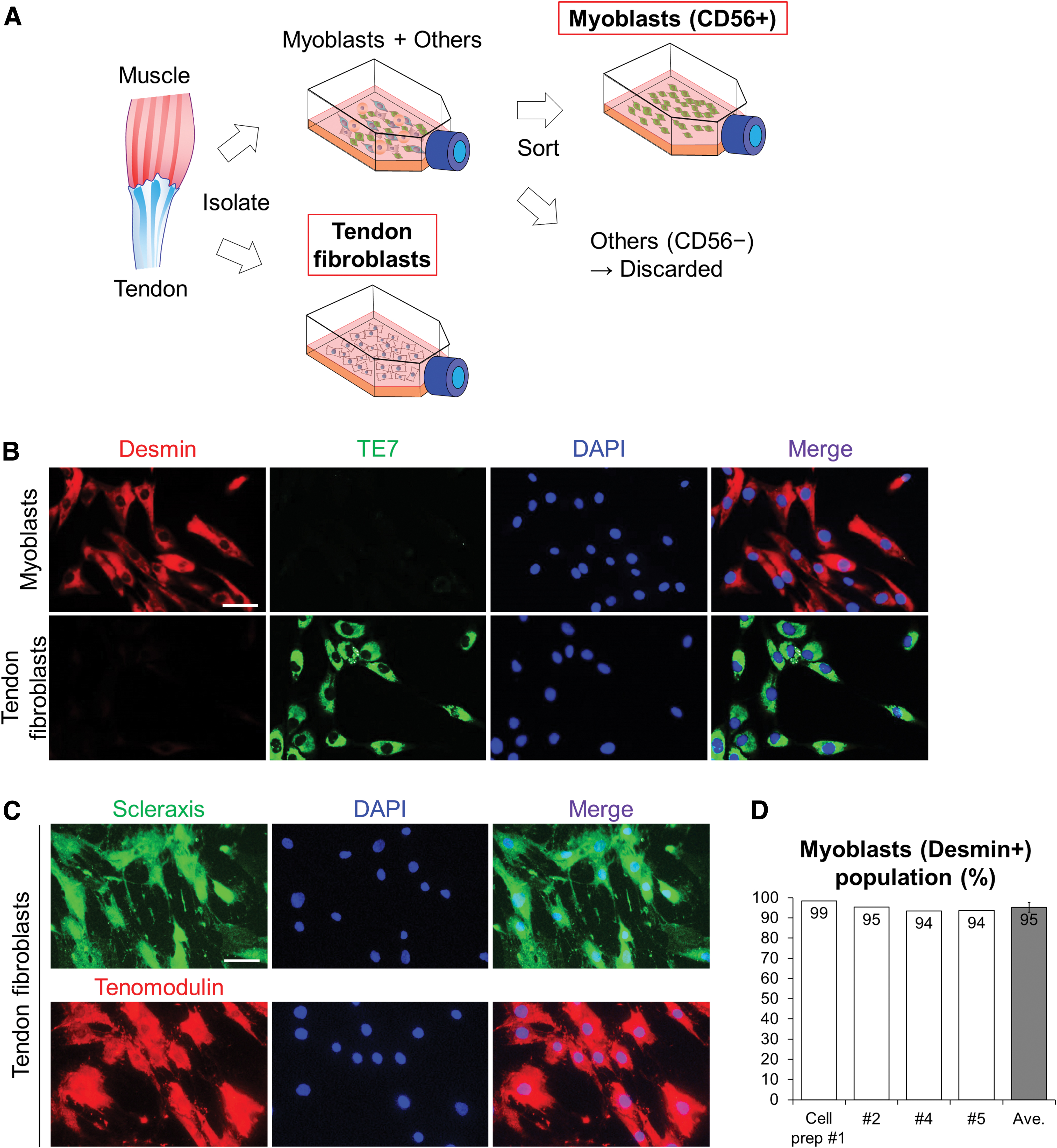
Purification of human-derived myoblasts and tendon fibroblasts.
Tendon tissue was cut into small pieces (∼0.5–1 cm3) after removing all visible nontendon tissue, the tendon pieces were digested overnight with 0.1% type II collagenase (Worthington Biochemical, NJ, USA; 43D14160) and 20% FBS in Dulbecco's modified Eagle's medium/nutrient mixture F-12 (DMEM/F12; Thermo Fisher Scientific, Hvidovre, Denmark; 21041025) at 37°C, in 5% CO2 for 17–24 h. After washing with DMEM/F12 supplemented with 10% FBS and 1% PS solution (tendon fibroblast growth medium [tGM]), tendon fibroblasts were cultured into T75 flasks until reaching sufficient cell numbers (≤80% confluent) in tGM. The mGM and tGM were changed every 2–3 days. The passage number of myoblast and tendon fibroblast was ≤6.
Purification of human myogenic cells
Human primary cultured muscle tissue-derived cell populations were purified to obtain myogenic cells (myoblasts) using anti-CD56 magnetic beads (Miltenyi Biotec SAS, Paris, France; 130-050-401) as described previously 29 (Fig. 1A). In brief, seeded myoblasts were trypsinized using trypsin ethylenediaminetetraacetic acid solution (Biological Industries, Beit Haemek, Israel; 03-054-1B) diluted in phosphate-buffered saline (PBS) (1:2) and were incubated for 1.5 min following washing with PBS. mGM was added immediately after cell detachment to protect cell surface epitopes from the trypsinization damage. After centrifugation for 6 min at 600 g, the cell pellet was mixed with 170 μL of magnetic cell isolation (MACS) buffer (Milteneyi Biotec; 130-091-221) and 35 μL of the anti-CD56 magnetic beads and incubated at 5°C for 15 min. The cell suspension was diluted with 5 mL MACS buffer, and centrifuged for 6 min at 600 g. The pellet was resuspended in 1 mL MACS buffer, and the solution was passed through a preseparation filter (Miltenyi Biotec; 130-041-407) and a large cell column (Miltenyi Biotec; 130-042-202) held in a MiniMACS separator (Miltenyi Biotec; 130-090-312).
The CD56– cells were washed away with MACS buffer, and the magnetically trapped CD56+ cells in the column were quickly collected into a CD56+ fraction tube after removal from the magnet. The CD56+ cells were recovered in mGM until sufficient cell numbers (≤80% confluent) and then used for the experiments.
Preparation of engineered human tendon constructs
Tendon constructs were engineered from the human primary cultured tendon fibroblasts using the Flexcell Tissue Train Culture System (Flexcell International Corporation, Burlington, USA; FX-6000TT), referring to previously published methods.30,31 Flexcell six-well plates (Flexcell International Corporation; TT4001U) with untreated silicone culture surfaces and foam anchors were used. Cells were seeded using a linear trough loader (Flexcell International Corporation; TT-4000TL) to generate an initially 2-mm-wide, 15-mm-long construct. Tendon fibroblasts were suspended in a mixture containing 50 μL of human plasma fibrinogen (Roche Sigma-Aldrich; F3879-1G) at 20 mg/mL, 2.5 μL of aprotinin (Roche Sigma-Aldrich; A6103-100MG) at 10 μg/mL, 1 μL of human plasma thrombin (Roche Sigma-Aldrich; T4393-100UN) at 3 U/mL, and 150 μL of construct medium (DMEM/F12 containing 10% FBS, 20 mM L-ascorbic acid 2-phosphate [Roche Sigma-Aldrich; A8960-5G], 10 mM L-proline [Roche Sigma-Aldrich; P5607-25G]) to a final concentration of 2 × 105 cells per well.
The ∼200 μL mixture was seeded quickly into the trough of the six-well plate, ensuring adequate infiltration into the two foam anchors at the ends of the trough and incubated at room temperature for 10 min. Then, the six-well plate was placed in the cell incubator at 37°C, in 5% CO2 for 30 min. After the incubation, 5 mL of construct medium was added. The construct medium was replaced every other day. All tendon constructs were cultured for 7 days.
Mechanical loading of three-dimensional engineered human tendon construct, collection of conditioned media, and RNA samples
Tendon constructs were mechanically loaded using the Flexcell Tissue Train culture system in the cell incubator. The six-well plate with tendon constructs was placed on the loading posts, and all constructs were washed with serum-free medium (SFM: DMEM/F12 containing 1% PS) three times before 3 mL of SFM was added into each well. Mechanical loading was performed in the incubator at 37°C. The foam anchors attached to the tendon constructs in the well were pulled downward by a vacuum, resulting in mechanical loading of the tissue construct. Tendon constructs were subjected to 4% uniaxial tensile strain at 0.5 Hz for 4 h in the Load condition based on a published study. 24 Control, nonrepetitive loaded tendon constructs in the same plate were made by using a rubber stopper to block the vacuum-induced loading (Control condition). We also prepared a negative control (Neg-con condition) where SFM was incubated in an empty well of the same plate. Immediately after the loading protocol, supernatants were collected from Neg-con, Control, and Load conditions (Fig. 3). All supernatants were stored at −80°C.
After collecting supernatants, all tendon constructs in Control and Load conditions were incubated with SFM for 4 h. Then, tendon constructs were divided to separate the central portion from the two anchors at the ends of the construct. All parts of the constructs were stored at −80°C, and the central portion was used for mRNA analysis in the present study.
Incubation with conditioned media
Before applying to myogenic cells, the conditioned media were diluted 1:1 with mGM or differentiation medium (DM) (PromoCell; C-23260).
Myoblasts for the proliferation experiments were plated onto a 24-well plate containing a sterilized 12 mm glass coverslip with a cell growth area of 1.13 cm2 (VWR, Darmstadt, Germany; 631-1577) at a density of 6 × 103 cells/well. All cells were initially incubated in mGM for 12 h for sufficient adhesion before applying the conditioned media. The myoblasts were washed with mGM and then incubated in the respective conditioned media for 3 days. All conditioned media were changed at 1.5 days (Fig. 3A). At day 3, myoblasts were extracted for real-time reverse transcription–polymerase chain reaction (RT-qPCR) analysis. Other myoblasts were incubated with 10 μM bromodeoxyuridine (BrdU) (Thermo Fisher Scientific; Ma1–19213) for 6 h before the fixation on day 3 for BrdU staining.
For the differentiation experiments (Fig. 4), myoblasts were seeded onto a 24-well plate at a density of 1.5 × 104 cells/well for 12 h with mGM, and then mGM was changed to conditioned media after washing with DM. All conditioned media were changed at 2 and 4 days (Fig. 4A). At day 5, myoblasts were extracted for RT-qPCR analysis. Other myoblasts were fixed for staining.
Immunofluorescence staining
All cells for immunofluorescence staining were fixed using Histofix (Histolab, Gothenburg, Sweden; 01000) for 8 min and permeabilized for 15 min with 0.01% Triton in 0.05 M Tris-buffered saline (TBS). For BrdU staining, fixed cells were incubated in 1 M hydrochloric acid (HCl) on ice for 10 min and incubated further in 2 M HCl for 10 min at room temperature followed by washing with 0.05 M TBS. All cells were incubated with appropriate primary antibodies (see below) in blocking buffer containing 1% bovine serum albumin (Roche Sigma-Aldrich; A3912-100G) in 0.05 M TBS at 4°C overnight and visualized using Alexa Fluor goat anti-mouse 488- and goat anti-rabbit 568-conjugated secondary antibodies (Thermo Fisher Scientific; A11029 and A11036). For nuclear staining, 4,6-diamidino-2-phenylindole (DAPI) mounting medium (Thermo Fisher Scientific; P36931) was used. All images were recorded using a camera (Olympus DP71; Olympus Deutschland, Hamburg, Germany) on a BX51 Olympus microscope, and digital images were taken using the software cellSens 1.14 (Olympus Deutschland).
For each well, five nonoverlapping images (center, top, left, bottom, and right sides of the cover glass) were captured at 10 × magnification (1700 μm × 1300 μm). The acquired digital images were quantified using Fiji software (NIH, Bethesda, MD, USA; version 1.52).
The following antibodies were used: rabbit anti-desmin antibody (Abcam, Cambridge, UK; AB32362); mouse anti-TE7 antibody (Merck, Darmstadt, Germany; CBL271); rabbit anti-Scx antibody (Abcam; AB58655); goat anti-tenomodulin antibody (Santa Cruz Biotechnology, Dallas, TX, USA; SC49325); rabbit anti-Ki67 antibody (Abcam; AB16667); mouse anti-BrdU antibody (Thermo Fisher Scientific; Ma1-19213); and mouse anti-myogenin antibody (Hybridoma Bank, IA, USA; F5D).
Fusion index was determined as the number of nuclei within desmin+ myotubes with ≥5 nuclei, divided by the total nuclei numbers. Differentiation index was determined as the number of myogenin+ nuclei within desmin+ myotubes with ≥5 nuclei, divided by the total number of desmin+ nuclei. The number of nuclei and myogenin+ nuclei within desmin+ myotubes was counted manually, and total nuclei number was counted automatically using Fiji software.
Myotube diameter measurements
The myotube diameters were measured as described in a published study. 32 In brief, five images (center, top, left, bottom, and right sides) with 10 × magnification were taken per well to analyze the myotube diameters. Two diagonal lines were automatically drawn on each image, and then smallest and largest diameter of the myotubes crossing each diagonal line were selected (i.e., two smallest and largest myotubes were selected per image). The diameters of the selected myotubes were measured perpendicularly from crossing the diagonal line to the myotubes using Fiji software. The average of the 10 smallest and largest diameters from five images per well was used as the minimum and maximum myotube diameters, respectively.
Real-time RT-PCR
RNA was extracted from cell cultures or tendon constructs using 1 mL TriReagent (Molecular Research Center, Cincinnati, OH, USA) as previously described. 23 Before extraction, the constructs were mechanically homogenized in the TriReagent containing five stainless steel balls of 2.3 mm in diameter (BioSpec Products, Bartlesville, OK, USA) by shaking twice in a FastPrep®-24 instrument (MP Biomedicals, Illkirch, France) at speed level 4 for 15 s. Total RNA concentrations were determined using RiboGreen assay (R11490; Fischer Scientific). Twenty-five nanograms (cell cultures) or 1 μg (constructs) total RNA were converted into cDNA in 20 μL reactions using the qScript reverse transcriptase (qScript cDNA Supermix; Qiagen, CA, USA) according to the manufacturer's protocol (Qiagen). For each target mRNA, 0.5 μL cDNA was amplified in a 25 μL SYBR Green PCR containing 1 × Quantitect SYBR Green Master Mix (Qiagen) and 100 nM of forward and reverse primers (Table 1). The amplification was monitored real-time using the CFX96 real-time PCR machine (Bio-Rad, CA, USA).
Primer Sequences for the Real-Time Polymerase Chain Reaction
The Ct values were related to a standard curve made with DNA oligonucleotides (UltramerTM oligos; Integrated DNA Technologies, Inc., Leuven, Belgium) with a DNA sequence corresponding to the sequence of the expected PCR product. The specificity of the PCR products was confirmed by melting curve analysis after amplification. Ribosomal protein lateral stalk subunit P0 (RPLP0) mRNA was chosen for normalization. No differences were seen between the conditions for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) normalized with RPLP0 as well as the Ct values of RPLP0, validating the use of RPLP0 as a normalizing gene. Data are shown relative to geometric mean of the Neg-con condition.
Statistical analysis
Data are shown as mean ± SD and/or individual values. A comparison of data between the Control and Load conditions was conducted using the Wilcoxon matched-pairs signed-rank test for mRNA data of tendon constructs. A nonparametric Friedman test was applied to identify significant differences among the three conditions (Neg-con, Control, and Load) for immunofluorescence staining and mRNA data of myoblasts. When a significant difference was observed, the Bonferroni correction method was used. Significance and trend were set at p < 0.05 and p < 0.1, respectively. All data were analyzed using SPSS version 28.0.1.1 (IBM SPSS Statistics, Armonk, NY, USA).
Results
Purification of human-derived myoblasts and tendon fibroblasts
We confirmed the purification of human-derived myoblasts and tendon fibroblasts by immunostaining (Fig. 1). Isolated CD56+ myoblasts and tendon fibroblasts were detected by immunostaining for desmin (myoblast marker) and TE7 (fibroblast marker), respectively (Fig. 1B). Furthermore, tendon fibroblast purity was confirmed by immunostaining for SCX and tenomodulin (Fig. 1C). Myoblasts' purity in the CD56+ fractions presented 95 ± 2% (mean ± SD) as desmin+ cells (Fig. 1D).
Gene expression of loaded human tendon constructs
Some studies have reported that engineered tendon constructs after loading increased in tendon-related gene markers.24,33 However, the details of these loading protocols differed from the present study. In this study, we determined whether tendon-related gene expressions of tendon constructs subjected to 4% strain at 0.5 Hz for 4 h were upregulated by the Flexcell loading system. SCX was significantly higher in the Load condition than in the Control condition (Fig. 2D), whereas cyclooxygenases 2 (COX-2) was significantly lower in the Load condition (Fig. 2E). Collagen type I, alpha 1 (COL1A1), collagen type III, alpha 1 (COL3A1), fibronectin, decorin, transforming growth factor beta 1 (TGFB1), integrin alpha 2, mohawk homeobox (MKX), interleukin 6 (IL6), or cyclooxygenases 1 (COX-1) did not differ between conditions. GAPDH, which was measured as another unrelated constitutive gene expression, was significantly lower in the Load condition compared with the Control condition (Fig. 2F). These results indicate that the loading protocol used in the present study upregulated SCX, and it did not induce inflammatory action.
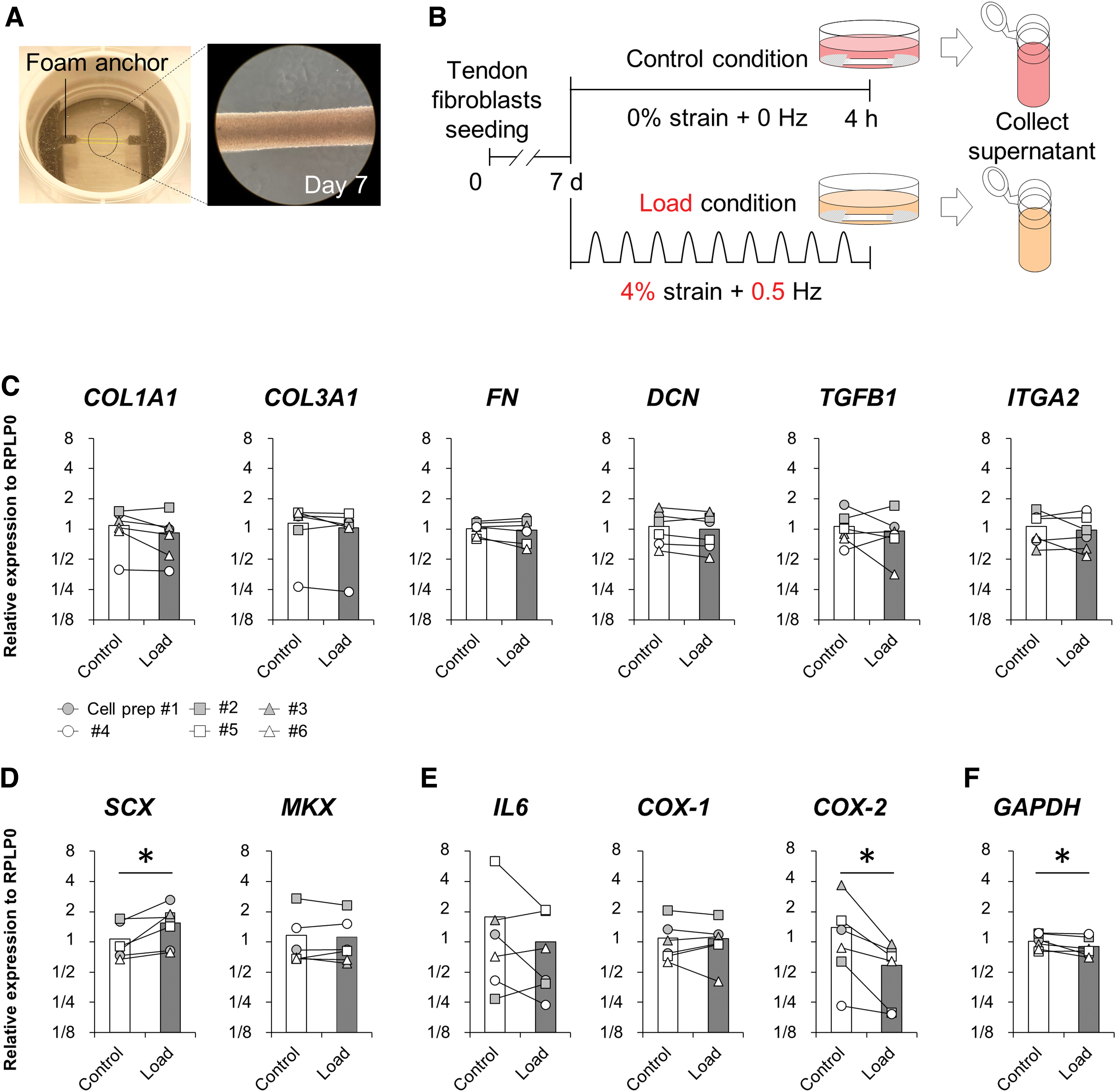
Engineered human tendon construct, protocol, and gene expression.
Conditioned media derived from mechanically loaded engineered human tendon constructs did not enhance proliferation of myoblasts
To evaluate how media conditioned by mechanically loaded tendon fibroblasts influence the proliferation of myoblasts, we compared the proliferation potential of myoblasts treated with media from Neg-con, Control, and Load conditions (Fig. 3A). We included the Neg-con condition to distinguish any proliferative potential of nonmechanically loaded tendon construct-derived factors. The total nuclei (DAPI+ cells), Ki67+ (as a marker for cells entering the cell cycle), or BrdU+ (as a marker for cells going into the S phase of cell cycle progression) nuclei number did not differ among conditions (Fig. 3B–E).
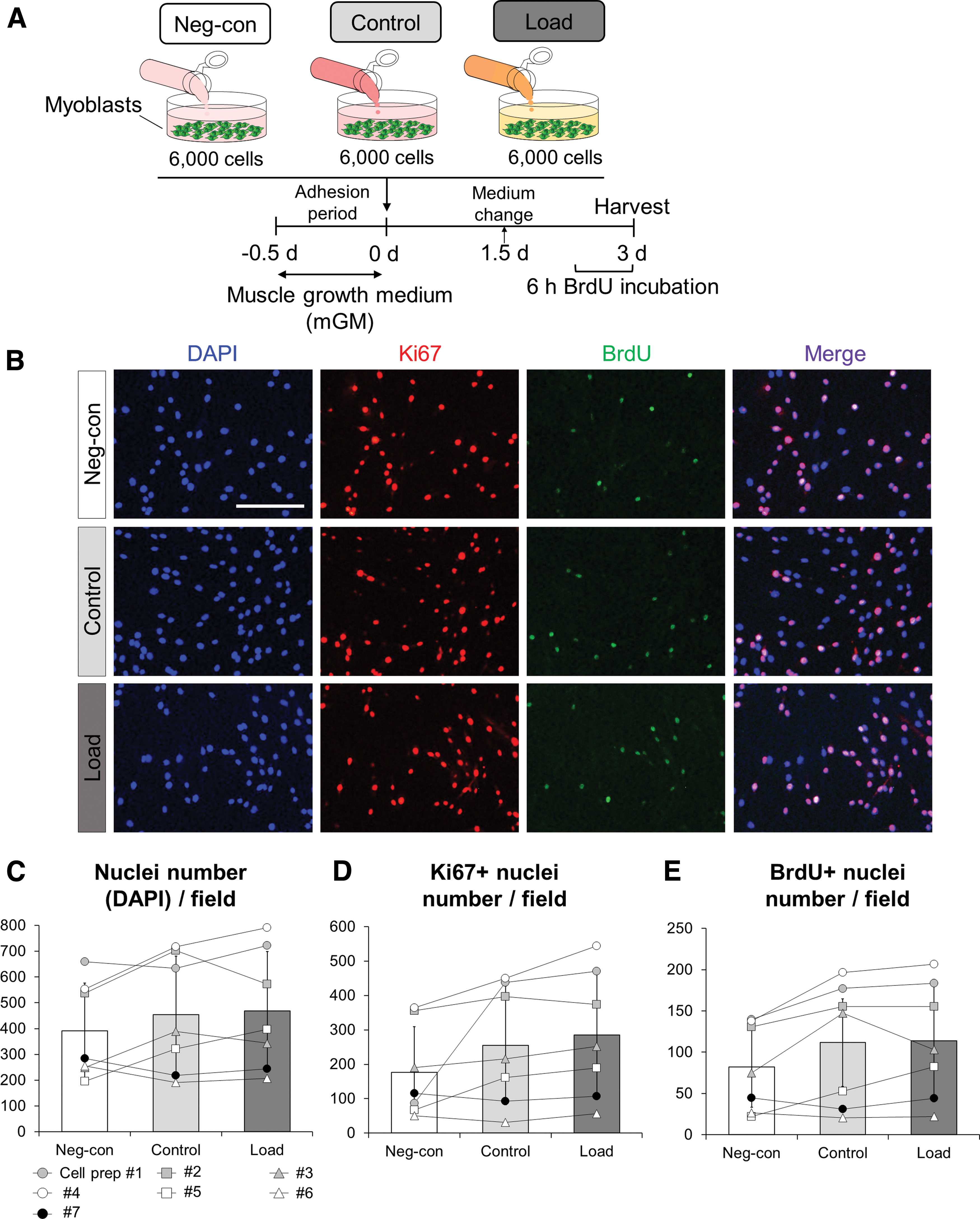
Mechanical loading of engineered tendon constructs does not influence proliferation of myoblasts.
Conditioned media derived from mechanically loaded engineered human tendon constructs enhanced differentiation of myoblasts
Satellite cells differentiate and subsequently undergo fusion with each other and the host myofibers following the activation and proliferation phases. Thus, we tested whether mechanically loaded human tendon constructs influenced the myogenic differentiation potential (Fig. 4A). Immunofluorescence analysis revealed a higher fusion index in the Load condition compared with Control condition even though no difference in nuclei number was observed (Fig. 4B, C, and F). Also, the fusion index in the Load condition had a tendency to be higher compared with the Neg-con condition (p = 0.098; Fig. 4B, F). However, there were no significant differences in the number of myogenin+ nuclei or differentiation index among conditions (Fig. 4B, D, and E). The minimum diameter of myotube in the Load condition was significantly larger than in the Neg-con condition (Fig. 4B, G). However, no difference in maximum diameter of the myotubes was found among the three conditions (Fig. 4B, H). These results indicate that conditioned media derived from mechanical loading of tendon constructs affect the fusion processes of myoblasts.
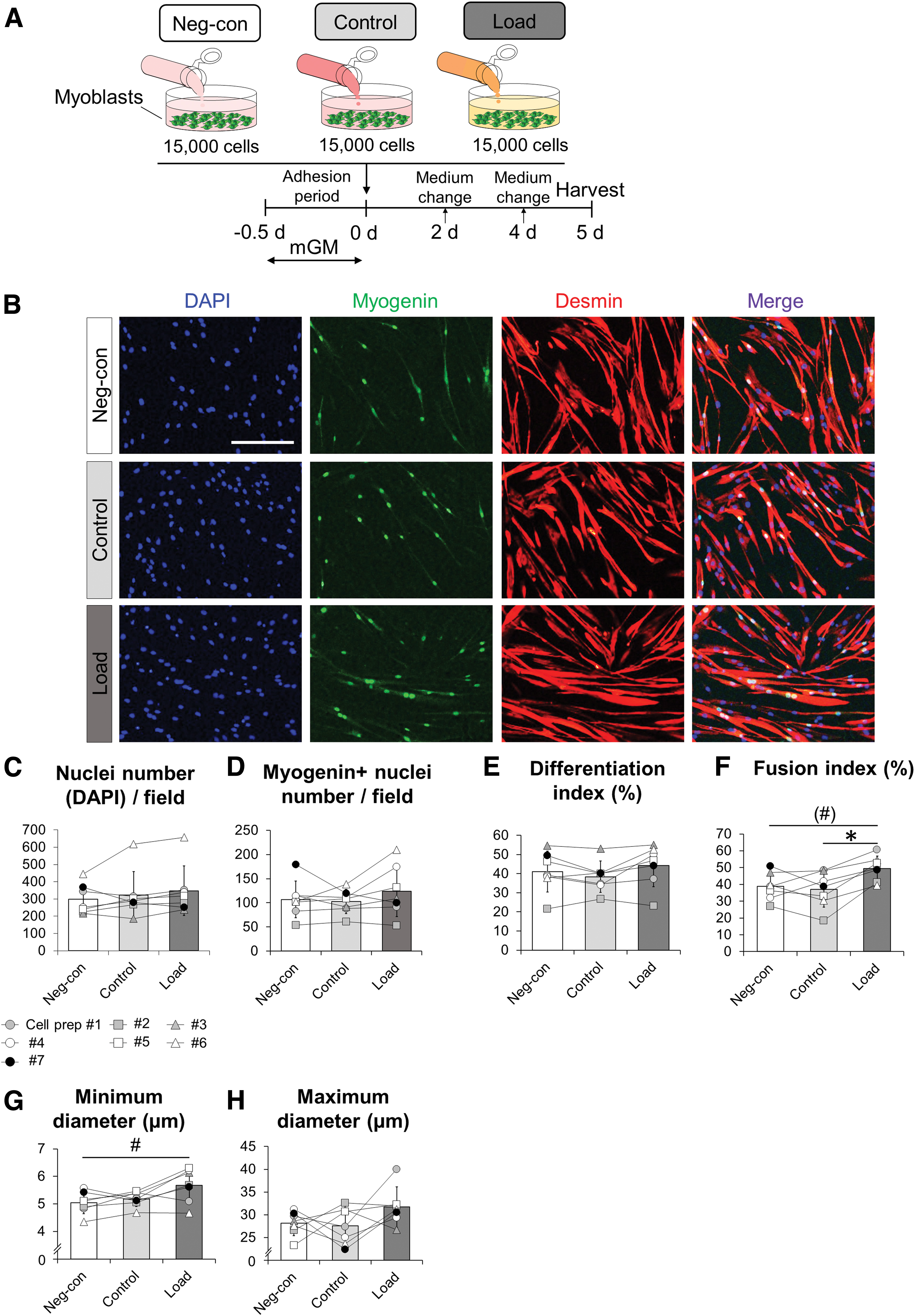
Mechanical loading of engineered tendon constructs enhances myotube formation.
Suppression of myostatin elevation and downregulation of myosin heavy chain 7 and AXIN2 in differentiated myoblasts are induced by mechanically loaded tendon construct-derived medium
Based on staining results in this study (Fig. 5), we next measured myogenic-related genes in differentiated myoblasts; myogenic differentiation 1 (MYOD1), myogenic factor 5 (MYF5), MYF6, myogenin (MYOG), myomaker (MYMK), myostatin (MSTN), myosin heavy chain 1 (MYH1), MYH2, MYH3, MYH7, and MYH8 isoforms were selected as markers of myogenic differentiation. In addition, insulin-like growth factor 1 receptor (IGF1R), axin2 (AXIN2), and chordin (CHRD) were selected as IGF1-, Wnt-, and bone morphogenetic protein signaling-related genes, respectively. GAPDH was measured as another unrelated constitutive mRNA. There were main effects for condition in MSTN, MYH3, MYH7, MYH8, and AXIN2 gene expressions, while no significances for MYOD1, MYF5, MYF6, MYOG, MYMK, MYH1, MYH2, IGF1R, CHRD, or GAPDH gene expressions were found. MSTN gene expression in the Control condition was significantly higher than the Neg-con condition (Fig. 5).
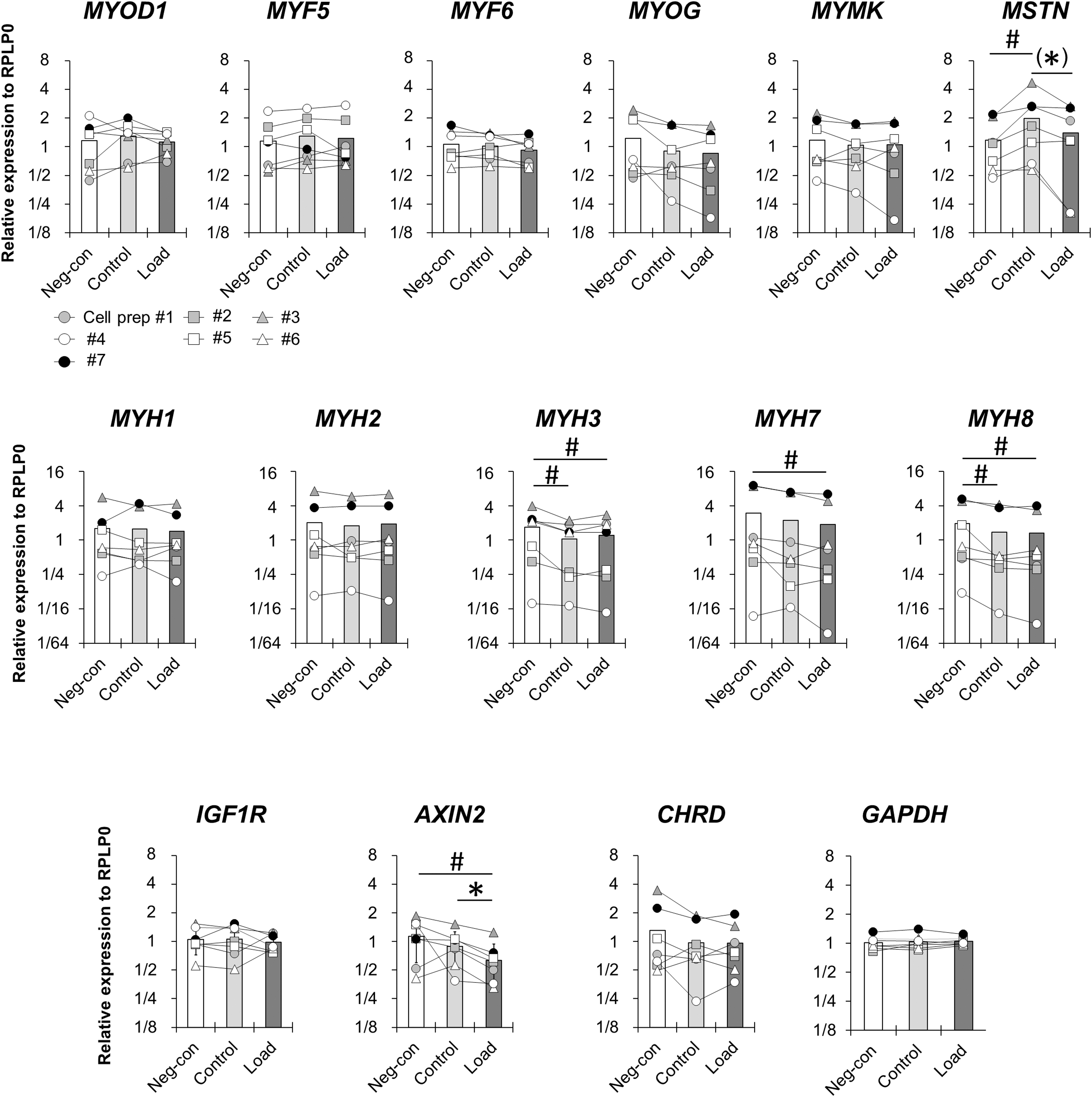
Gene expression. Real-time RT-PCR analysis of differentiated myoblast cultures in MYOD1, MYF5, MYF6, MYOG, MYMK, MSTN, MYH1, MYH2, MYH3, MYH7, MYH8, IGF1R, AXIN2, CHRD, and GAPDH. Data are normalized to RPLP0 and expressed relative to the geometric mean of the Neg-con condition. N = 7 in each condition. The symbols connected with a line among conditions correspond to myoblasts from the same patients. *p < 0.05 vs. Control, #p < 0.05 vs. Neg-con. Trends (0.05 < p < 0.1) indicated in brackets. CHRD, chordin; IGF1R, insulin-like growth factor 1 receptor; MYF, myogenic factor; MYH, myosin heavy chain; MYMK, myomaker; MYOD1, myogenic differentiation 1; MYOG, myogenin; MSTN, myostatin.
Furthermore, the MSTN in the Load condition tended to be lower than the Control condition (p = 0.091; Fig. 5). MYH7 and AXIN2 were significantly downregulated only in the Load condition compared with Neg-con condition. Additionally, AXIN2 in the Load condition was significantly lower than in the Control condition (Fig. 5). Although MYH3 and MYH8 in both Control and Load conditions showed a significantly lower value compared with Neg-con condition, there were no significant differences between Control and Load conditions (Fig. 5). These results suggest that myoblasts with a high fusion capacity in Load condition suppressed MSTN elevation and downregulated MYH7 and AXIN2. In addition, regardless of mechanical loading regimen, incubation of myoblasts with conditioned media from tendon constructs resulted in a downregulation of MYH3 and MYH8.
Discussion
The main finding of this study was that in response to mechanical loading engineered tendon constructs from human-derived tendon fibroblasts enhanced myotube formation by myoblasts but did not affect their proliferation.
Cells in skeletal muscle and tendon tissues are exposed to repetitive mechanical loading. Therefore, a perspective of muscle and tendon as a unit is necessary to understand how cells in each tissue behave under mechanical loading for maintaining their homeostasis and healing/regeneration abilities. Tendon mechanical loading-induced factors might circulate not only to adjacent tissues (skeletal muscle, bone) but also to other tissues in the body. In support of this, blood flow in tendon tissue increased following acute exercise in healthy humans,34–36 as well as in tendinopathy patients. 37 It has been shown that mechanical loading induces tenogenic responses in tendon fibroblasts and engineered tendon construct models.24–28 However, to our knowledge, this is the first study to establish how mechanical loading of tendons influences the myogenic progression of muscle stem cells.
We have used tendon constructs and a Flexcell loading system, which allowed the comparison of repetitive mechanical and nonrepetitive mechanical loading conditions. The protocol used in the present study subjected tendon constructs to a load that somewhat mimics an in vivo mechanical loading situation. 24 This loading protocol has previously been observed to promote synthesis of collagen type I and TGFB1 proteins in human tendon fibroblasts on a microgrooved substrate. 24 In the present study, we could not observe an increase in the expression of COL1A1 and TGFB1, but SCX gene expression was upregulated in the Load condition. Scx is a marker of both progenitors and differentiated tendon cells during tendon regeneration and development.18,38
At the tissue level, disruption of the fibril structure has been observed in tendons stretched beyond 4%.39,40 It has previously been demonstrated that cell viability in engineered tendon constructs subjected to overload (2 h at 8% strain, 0.1 Hz) for 7 days was not significantly different to constructs subjected to physiological load (2 h at 4% strain, 0.1 Hz), but cellular density tended to be higher in physiologically loaded constructs. 33 That study also showed a significant reduction of Scx, Mkx, Col1a1, and Col3a1 gene expression in overloaded tendon constructs. In the present study, significant decreases in these gene expressions were not observed. Furthermore, no significant elevations in inflammatory-related gene IL6, COX-1, or COX-2 expressions were found in the Load condition. From this evidence, we assume that the protocol used in the present study represents a nondamaging stimulus in the physiological range, which would be beneficial not only for myogenic progression in the muscle tissue but also for tenogenic progression in the tendon tissue.
We revealed that conditioned media from tendon constructs with mechanical loading stimulated the fusion process of myoblasts, indicating that mechanical loading-induced factors released by tendon–fibroblasts contributed to the myotube formation. A number of in vitro studies have reported that tendon fibroblasts produced several cytokines and endogenous factors in response to mechanical loading, such as IL6, TGFB1, angiopoietin-like 4, and prostaglandin E2,24,41–44 but all of the released substances have not fully been identified yet. In the present study, no elevations of MSTN, a negative regulator of myogenic progression, 45 lower expressions of MYH7, and AXIN2, a positive regulator of myoblasts differentiation 46 were observed in the Load condition. It is well known that MSTN strongly downregulates myogenic progression, 45 and in vitro, the fusion index of human primary myoblasts has also exhibited a dose-dependent reduction by MSTN recombinant protein. 47 However, MYH7 and AXIN2 expressions also decreased concomitantly in the Load condition in the present study.
MYH7 gene expression in myoblasts was promoted with treatment of a MSTN protein, 48 and mice deficient in the Mstn gene exhibited decreases in both Myh7 expression and slow-twitch myofiber distribution because of MYH7 encoding a slow-type myofiber. 49 Therefore, suppressed MSTN expression would be expected to enhance myotube formation with a decrease of slow-twitch myofiber distribution, which could result in the observed larger myotube diameters in the Load condition. Regarding the relationship between MSTN and AXIN2 on myogenic regulation, it has been shown that MSTN antagonizes axin2, which is a Wnt4-related factor. The present result may be due to axin2 acting in a negative feedback loop to maintain the normal levels in the Wnt signaling pathway following an elevation of the Wnt signal initiation. 50 Thus, conditioned media from tendon constructs subjected to mechanical loading could contain factors that not only suppress MSTN and MYH7 elevations but also induces negative feedback from AXIN2 in Wnt signaling.
As a candidate for the loading-induced factors in conditioned media based on the gene expression results, mitochondrial open-reading-frame of the twelve S rRNA-c (MOTS-c), tumor necrosis factor-α (TNFα), or IGF1 can be expected. MOTS-c is a circulating hormone that inhibits MSTN action, 51 while TNFα is a cytokine stimulating the MSTN gene. 52 IGF1 has been shown to antagonize the action of MSTN; in addition, IGF1 gene expression in tendon tissue is increased by repetitive contraction of the muscle–tendon unit. 53 However, it is unknown whether these factors are released from tendon-engineered constructs; further investigations are therefore required to identify involved factors. Lastly, caution is needed when interpreting the qPCR results of myoblasts because we did not find significant statistical differences for MSTN and MYH7 between the Control and Load condition.
We also revealed that tendon constructs themselves downregulated several myogenic-related gene expressions regardless of mechanical loading. In the present study, MYH3 and MYH8 expressions represented a lower value in Control and Load conditions when compared with the Neg-con condition. As a possibility, it might be caused by a catabolic action of factors derived from tendon constructs since IL6 is released from the tendon-derived cells as an inflammatory cytokine.41,44 To our knowledge, there is no supportive studies showing that engineered tendon constructs produce negative effectors involved in myogenic genes. It is worth considering that apparent inconsistency of the present results between immunofluorescence staining (enhanced myogenesis) and the gene expressions (reduced MYH3 and MYH8) (Figs. 4 and 5), could be caused by a delay in the onset of myotube formation between gene and protein expressions. To elucidate this issue, collecting samples with several time courses and analyzing their molecular signaling would be necessary.
We have previously shown the importance of constant crosstalk for enhancing myotube formation between tendon fibroblasts and myoblasts through fluid-mediated signaling loop during coculture in the two-dimensional culture. 23 One limitation in the present study is that we could not technically compose a coculture model that enables constant crosstalk between myoblasts and tendon constructs while under mechanical loading, which would better mimic a physiological situation.
In conclusion, we have revealed that engineered human tendon tissues undergoing repetitive tensile mechanical loading release factors that enhanced myotube formation. Our findings extend the fundamental understanding of the muscle–tendon unit across adjacent tissues for homeostasis, repair, and regeneration.
Footnotes
Acknowledgments
The authors thank Monika Bayer, Casper Soendenbroe, Anja Jokipii-Utzon (Institute of Sports Medicine Copenhagen), and Vibeke Pless (Bispebjerg Hospital Department of Dermatology) for technical support and helpful discussions.
Myogenin antibody (F5D), developed by Wright W.E., was obtained from the Developmental Studies Hybridoma Bank, created by the NICHD of the NIH, and maintained at The University of Iowa, Department of Biology, Iowa City, IA, USA.
Authors' Contributions
Y.T. designed and performed the experiments, interpreted and analyzed the data, and wrote the article. R.B.S. designed the experiments with technical support, interpreted the data, and reviewed the article. C.-Y.C.Y. designed experiments with technical support, interpreted the data, and reviewed the article. P.S. analyzed the PCR data and reviewed the article. M.K. acquired funding, interpreted data, and reviewed the article.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the Japan Society for the Promotion of Science (JSPS) Overseas Research Fellowships (1120606) and the Uehara Memorial Foundation for Overseas postdoctoral fellowships. This work was also supported by the Danish Medical Research Council (FSS) (0134-00028B), Lundbeck Foundation (R198-2015-207), and Novo-Nordisk Foundation (NNF16-0C0022846).