
Abstract
The lymphatic system maintains tissue fluid homeostasis and it is involved in the transport of nutrients and immunosurveillance. It also plays a pivotal role in both pathological and regenerative processes. Lymphatic development in the embryo occurs by polarization and proliferation of lymphatic endothelial cells from the lymph sacs, that is, lymphangiogenesis. Alternatively, lymphvasculogenesis further contributes to the formation of lymphatic vessels. In adult tissues, lymphatic formation rarely occurs under physiological conditions, being restricted to pathological processes. In lymphvasculogenesis, progenitor cells seem to be a source of lymphatic vessels. Indeed, mesenchymal stem cells, adipose stem cells, endothelial progenitor cells, and colony-forming endothelial cells are able to promote lymphatic regeneration by different mechanisms, such as direct differentiation and paracrine effects. In this review, we summarize what is known on the diverse stem/progenitor cell niches available for the lymphatic system, emphasizing the potential that these cells hold for lymphatic tissue engineering through 3D bioprinting and their translation to clinical application.
Impact statement
The lymphatic system undertakes many physiological functions of the body. Growing evidence suggests it also participates in different pathologies such as cancer metastasis. Lymphangiogenesis is the formation of new lymphatic vessels from pre-existing ones and is mainly limited to embryonic development or pathologic conditions. Conversely, lymphvasculogenesis is described as the formation of new lymphatic vessels from progenitor cells. The origin and formation of lymphatic vessels in the adult remains obscure. This review describes the different stem/progenitor cells niches in the body and their adequacy as cell sources for 3D bioprinting of vessels for lymphatic tissue regeneration.
Introduction: Structure of Lymphatic Vessels
The lymphatic system has been relatively understudied for decades. Of late, it has gained more attention due to its increasing implication in both pathological and physiological processes. Historically, the lymphatic system has been attributed to the fluid-tissue homeostasis upkeep, the transport of nutrients from the intestinal system to the blood circulation, and the immunosurveillance function of solid tissues. These functions are achieved by a highly complex organization of a one-way hierarchical system composed of five types of vessels: the capillaries, collecting vessels, lymph nodes, trunks, and ducts. 1 Lymphatic vessels at the periphery are formed by lymphatic capillaries, which are composed of a single layer of endothelial cells linked to one another through button-like junctions, that give the capillaries their one-way valve function. 2
Capillaries range from 10 to 60 μm in diameter and are normally nonfenestrated. Lymphatic endothelial cells (LEC) of the capillaries are lined with a noncontinuous basement membrane and connected to the extracellular matrix (ECM) by anchoring filaments. 3 Capillaries drain into structurally and functionally different, larger vessels called the collecting lymphatic vessels. While capillaries are composed of a single layer of endothelial cells linked to one-another through button-like junctions, LECs of collecting vessels are bound more tightly via zipper-like junctions. 4 In addition, collecting vessels present a continuous basement membrane and a layer of smooth muscle cells (SMC). Their main function is no longer collecting the extravasated fluid from the interstitial space but transporting it toward blood circulation through two different pumping units. 5
On the one hand, lymphatic collecting vessels contain one-way valves, 6 which favor the production and maintenance of a pressure gradient between lymphangions (a lymphangion comprises the tube segment between two valves), in turn, favoring a pressure gradient from capillaries toward lymphatic ducts. 7 On the other hand, the SMC layer of the collecting lymphatic vessels produces a peristaltic contraction that propels the lymph toward the lymph nodes. 8 The immune cells within the nodes act as a first barrier of defense against infection. 7 Lymph enters the nodes through the afferent vessels, and it is filtered by the macrophages and lymphocytes that remove foreign pathogens, cell debris, and proteins. 9 All of them drain into bigger lymphatic trunks that end up draining into the main two lymphatic ducts, and subsequently, into blood circulation. 10
Historically, theories on the formation of the lymphatic system have been controversial. In brief, Florence Sabin first proposed the centrifugal model. This model described the formation of the lymphatic system from the endothelial cells budding from the veins during embryonic development, to form the primary lymph sacs that mature into an interconnected web of lymphatic vessels. 11 On the contrary, Huntington and McClure described an opposite development model (the centripetal model), in which primary lymph sacs stem from mesenchymal stromal cells (MSCs) completely independent from veins. 12 Ever since, many studies have provided compelling arguments in defense of both models, and in fact, both of these seem to be correct. Nevertheless, postnatal lymphangiogenesis is secluded to only different pathological conditions (tumorigenesis, tumor metastasis, inflammation, and wound healing), as it rarely occurs under physiological conditions.13–15
Adult lymphangiogenesis has been described to mimic embryonic lymphangiogenesis; however, the newly formed lymphatic vessels tend to be malformed and poorly functional. 16 Even though a major portion of newly formed lymphatic vessels arises from pre-existing lymphatics, several studies have pointed out the formation of de novo lymphatic vessels from stem cells, known as lymphvasculogenesis.17,18 Many different studies have proven the presence of lymphatic endothelial progenitor cells (LEPCs) in different tissues and their implication in adult lymphangiogenesis (Fig. 1). Likewise, stem cell-based therapies have gained a lot of interest in recent years, due to their unique potential. Nevertheless, there are still many unknowns regarding the formation of adult lymphatic vessels and their implication in pathological conditions. Therefore, understanding the sources of lymphatic progenitors available in the adult body and how to make use of them for 3D bioprinting applications might be of great value for the treatment of lymphatic pathologies and related diseases.
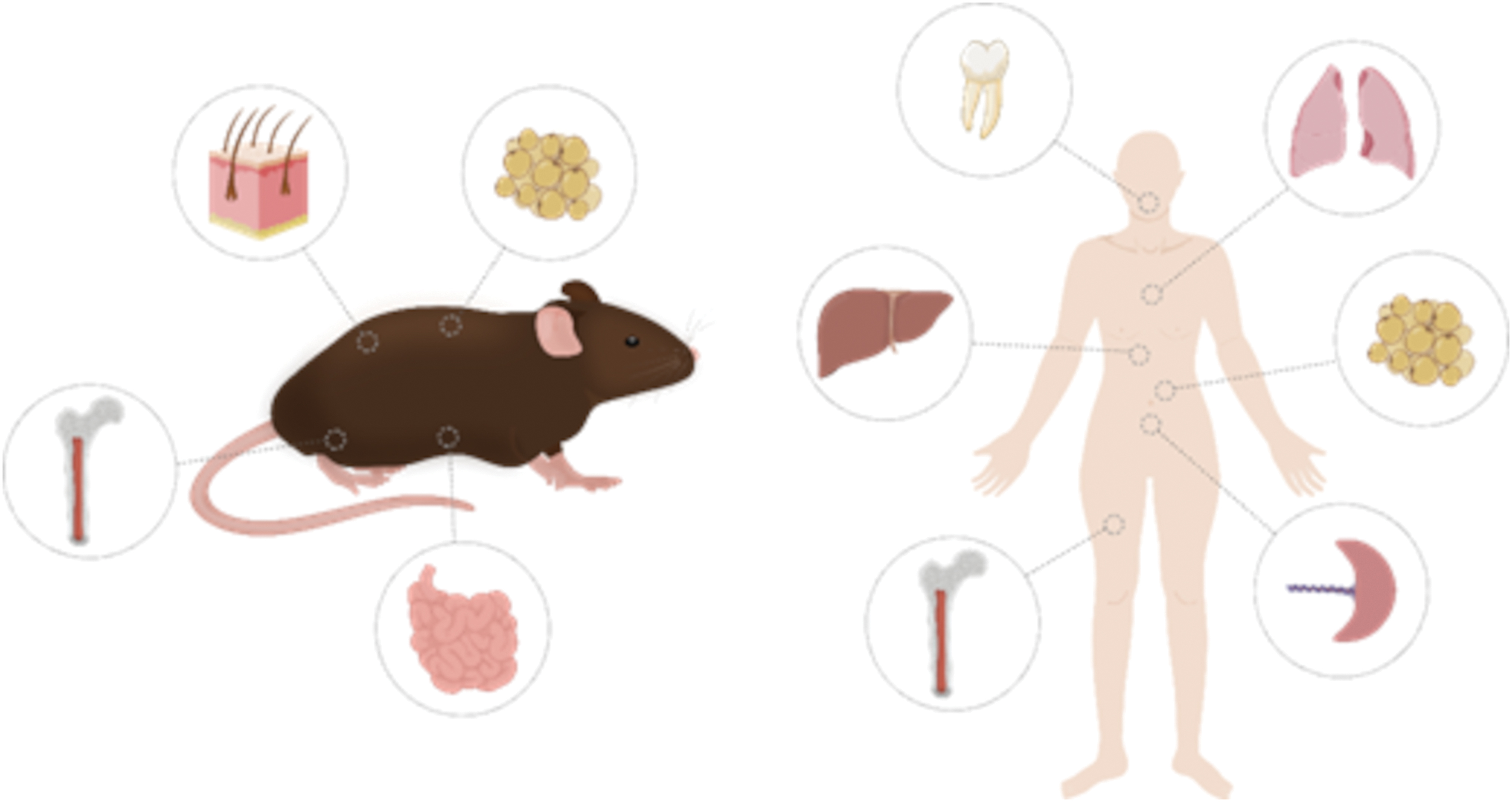
Illustration summarizing the different tissues studied as a potential source of stem cells and lymphatic endothelial progenitor cells. Color images are available online.
Method: In Vivo Lymphatic Precursors and In Vitro Differentiation
Bone marrow-derived LEPCs
Bone marrow cells actively engage in several physiologic activities, such as inflammation, hematopoiesis, and angiogenesis. 19 The role of hematopoietic cell-derived endothelial cell precursors in angiogenesis has been extensively studied. 20 More recently, the possibility of obtaining LEPCs from the bone marrow has attracted more interest.
Indeed, the bone marrow harbors cells of both hematopoietic and nonhematopoietic origins. In recent years, growing evidence shows the contribution of the bone marrow in neolymphangiogenesis, which is postulated to occur via two distinct routes: (i) activation of bone marrow-derived macrophages and their subsequent secretion of prolymphangiogenic cytokines and (ii) direct differentiation of LEPCs into LECs.17,21 In 2012, the expression of podoplanin (a marker of LEPCs) by BM-derived macrophages was first reported under inflammatory conditions. 22 Bone marrow-derived macrophages exhibit significant lymphangiogenic capacity during embryogenesis and in the presence of prolymphangiogenic cytokines, such as vascular endothelial growth factor (VEGF) A and C, and induce infiltration of new lymph vessels into breast cancer tumors.23–25
Other scientists have recently revealed that BM-derived macrophages express a number of LEPC markers (LYVE-1; Podoplanin; VEGFR-3; PROX1) and adopt a tube-like structure in the presence of tumor microenvironment-derived hyaluronan. 26 Similarly, tumor microenvironment-derived TNFα induces the differentiation of bone marrow-derived monocytes toward an endothelial phenotype through the expression of integrin α5 and adhesion of the cells to fibronectin. 27 Moreover, lymphatic differentiation of myeloid LEPCs is inducible with lipopolysaccharide (LPS), a ligand of Toll-Like Receptor-4 (TLR4). 28 Another study injected BM-derived macrophages of the M2-phenotype into myocardial ischemia-suffering rats. The injections resulted in increased lymphangiogenesis in vivo; and increased proliferation, migration, tube formation, and expression of VEGF-C in LECs in vitro. 29
Nonetheless, some discrepancies have arisen with regard to the prolymphangiogenic macrophage phenotype, which may be related to disease-specific factors. For instance, in renal fibrosis, M1 macrophages polarized and transdifferentiated into LECs through the stimulation of the VEGF-C/VEGFR-3 pathway, both in vitro and in vivo. 30
Conversely, other research groups have provided compelling evidence that BM-derived mesenchymal stromal cells (BM-MSCs) contribute to lymphangiogenesis in vivo and in vitro, via paracrine signaling of VEGF-A. 31 Injection of BM-MSC increased tumor growth and intratumoral lymphatic vessel density in vivo and proliferation and migration of LECs and immortalized LECs in vitro. Another study further confirmed that BM-MSC-derived conditioned medium promoted the expression of VEGF on human dermal lymphatic endothelial cells (HDLEC). Culturing HDLECs in BM-MSC-derived conditioned medium promoted tube formation and migration, suggesting that BM-MSCs play an active role in tumor lymphangiogenesis. 32 In addition, MSCs hold differentiation potential to multiple cellular lineages; including cells of the lymphatic system under prolymphangiogenic growth conditions.
In 2016, Igarashi et al. described that VEGF-C considerably increased MSC proliferation and migration and induced the overexpression of PROX1 and LYVE1, while precluding the expression of osteogenic markers. 33 Nonetheless, MSCs have demonstrated to exert a more prominent paracrine effect on neolymphangiogenesis by promoting the differentiation of endothelial cells toward lymphatic phenotype through the secretion of VEGF-C rather than through their own differentiation.34,35
Of note, there have been several approaches to translate nonclinical research onto patients.36,37 Recently, a randomized controlled trial on lower limb lymphedema compared autologous transplantation of BM-derived mononuclear cells (BM-MNCs) to the current gold standard, compression therapy. 38 The study showed that patients receiving autologous BM-MNC transplants presented a reduction of limb circumference, pain relief, and improvement in walking ability, in contrast to patients with compression therapy where no improvement was seen.
Umbilical cord-derived LEPCs
Human umbilical cord-derived mesenchymal stromal cells (UC-MSCs) fall into the ample definition of MSCs, nevertheless, they present significant differences in proliferation and gene expression compared to BM-MSCs. 39 UC-MSCs' main advantage in comparison with BM-MSCs is that the acquisition of the former is less invasive and easier to perform. Further, they possess a higher proliferation capacity, greater potency, and rarely produce teratomas.40–42 On the negative side, there are hardly any studies on the feasibility of using UC-MSCs and their derivatives as progenitors of lymphatic vessels.
The umbilical cord also harbors endothelial progenitor cells (EPCs). In fact, EPCs are a viable and noninvasively accessible stem cell niche. These cells play an important role in postnatal angiogenesis but their lymphangiogenic capacity remains virtually unknown. The contribution of EPCs to lymphangiogenesis has been postulated to occur via direct differentiation of the lymphatic progenitors into mature LECs. 43 Tan et al. proved that a small subpopulation of EPCs (CD34+VEGFR-3+ cells) held lymphatic potential, and could be differentiated into LYVE1+PROX-1+ cells through the stimulation of the VEGF-C/VEGFR-3 pathway in culture. 44 Simultaneously, others described in vitro differentiation potential of human cord blood progenitors into Podoplanin+VEGFR-3+LYVE-1+CD11+ cells (LEC and myeloid markers), confirming the potency of the lymphatic precursors. 45
Recently, the lymphangiogenic capacity of VEGFR-3+Pod+CD11b+ LEPCs found in the human cord blood has been confirmed. 46 Besides human cord blood, other scientists described a subpopulation of endothelial colony-forming cells (ECFC) from peripheral blood that expressed all the aforementioned lymphatic markers (high expression levels of PROX-1, VEGFR-3, LYVE-1, and Podoplanin) and lacked blood endothelial cell-specific markers (VEGFR-1, among others). 47 However, these findings need independent confirmation.
Few studies have progressed onto translating nonclinical findings to the clinic. One of the few investigated the adequacy of alginate hydrogels as delivery system of the cells, along with the lymphangiogenic capacity of the EPCs. The authors concluded that EPCs possess lymphatic and blood subpopulations that react with the growth factors in the surrounding environment, and confirmed the suitability of alginate as a supporting material for said application. 48
Adipose tissue-derived LEPCs
An alternative to obtain LEPCs are adipose tissue-derived stromal cells (ADSCs), which are adult stem cells of adipose tissue with the ability to self-renew, proliferate, and differentiate into many different lineages. In comparison with BM-MSCs, ADSCs are quite similar in terms of proliferation, morphology, potency, and senescence, 49 but ADSCs have the advantage of being more accessible and easier to obtain than BM-MSCs. Further, the number of MSC colonies routinely obtained in ADSC cell cultures are in the order of 500-fold than those usually obtained from the bone marrow. 50
Increasing evidence indicates that the involvement of ADSCs in lymphangiogenesis may take place via two different phenomena: (i) the paracrine effect exerted by growth factors released by ADSC; or (ii) direct differentiation of ADSCs into LEPCs. Some studies have given compelling evidence on the lymphangiogenic capacity of ADSCs, mainly attributed to paracrine signaling. Coculture experiments of human-ADSC (hADSCs) and human LECs with direct and indirect contact with recombinant VEGF-C, concluded that coculturing of hLECs with hADSCs presented almost the same effect as VEGF-C culture, and promoted migration and proliferation of the former, but no tube formation. 51
Recently, other scientists set reliable grounds on the paracrine effect of hADSC-conditioned medium on LECs in vitro. 52 Both of these studies along with others suggest that the paracrine effect of the hADSCs on LECs is mainly mediated by the secretion of VEGF-C and its interaction with membrane bound receptor VEGFR3.53–55 In addition, Ahmadzadeh et al. concluded that hADSC-conditioned medium had a greater effect than addition of VEGF-C, bFGF, and HGF factors, even though the detected levels of these factors in the conditioned medium were very low. 56
Other studies have put the focus on direct differentiation of ADSCs into LECs, instead of relying on the paracrine effect they exert. 57 Older studies regarding differentiation of ADSC into LEPC were, in general, focused on the in vivo injection of ADSCs.58,59 Others found that implanted podoplanin-positive ADSCs not only increased lymphatic capillary density and reduced lymphedema but also augmented the recruitment of bone marrow-derived M2-phenotype macrophages. 60 Other approaches complemented in vivo cell injection of ADSCs with transplantation of vascularized lymph node to hindlimb lymphedema-induced mice. The results indicated that complementing vascularized lymph node transfer with ADSCs significantly promotes neolymphangiogenesis, compared to lymph node transfer only. 61 Another study described how the overexpression of PROX1 promotes the differentiation of ADSCs into LECs, by promoting the expression of LEC-specific markers. 62
Recently, other authors described a subpopulation of podoplanin-positive ADSCs that (i) upregulated LEC-specific cellular markers and possessed endothelial-like morphology in vitro, and (ii) induced lymphangiogenesis in vivo. 63 The authors attributed to podoplanin-positive ADSCs the ability to exert paracrine signaling and direct differentiation into LEPCs; whereas podoplanin-negative ADSCs were limited to paracrine signaling by secretion of MSC factors.
In summary, in vitro differentiation of ADSCs into LECs has followed different experimental approaches: (i) differentiation by culturing with prolymphangiogenic growth factors (such as VEGF-C156S, bFGF/FGF2, and CCBE1, among others)30,64,65; (ii) inducing the differentiation with external growth factors and concomitant inhibition of other inhibitory pathways 58 ; (iii) culturing with proinflammatory interleukins 66 ; (iv) coculturing with mature LECs51,63; and (v) overexpression of prolymphangiogenic transcription factors with microRNA transcripts.62,67
Several studies have investigated the feasibility of using so-called adipose-derived regenerative cells (ADRCs) for cellular therapy of breast cancer-related lymphedema (BCRL).68,69 The first pilot study tested the safety and feasibility of the procedure with 1-year follow-up after the implantation. Their results indicated an improvement in the quality of life of patients, with no quantitative decrease in lymphedema. 70 In 2021, the same group published a phase I clinical trial, in which 10 BCRL patients underwent ADRC injection. At 4-year follow-up, the patients had improved upper extremity function and BCRL symptoms. Nonetheless, the authors highlighted the need for a randomized controlled trial. 71
Dermis-derived LEPCs
Over 20 years ago, seminal articles by Kriehuber et al. and Breiteneder-Geleff et al. kick-started lymphatic tissue research by isolating for the first time podoplanin-positive and -negative cells from dermal suspensions, confirming stable lineage differences between both cell fractions.72,73 Ever since, several studies have suggested the adequacy of the dermis as a potential cell source for many tissue engineering applications, the engineering of lymphatic tissue being one among others.
As opposed to the aforementioned cell sources, the majority of the studies regarding the suitability of the dermis as a potential source of lymphatic cells have focused on using mature LECs harvested from different skin surgery by-products (such as circumcision or mastectomies) and subsequent cell proliferation in vitro. Although tissue engineering of lymphatic tissue has not received much experimental consideration, HDLEC from children foreskin's have become the “gold standard” cell source in practice.56,74–77 Nevertheless, to our knowledge transplantation of HDLECs into animals to regenerate the diseased area has barely been addressed experimentally. One of the few such studies demonstrated that podoplanin-positive cells of rat dermis improved secondary lymphedema in nude rats' tails. 78 These findings suggest that human dermal LECs can induce lymphangiogenesis.
Other LEPC sources
Other researchers have focused on quite distinct tissue sources for LEPCs. These sources are (i) human dental pulp stem cells (Pod+LYVE1+PROX1+VEGFR-3+), 79 (ii) human or animal lung-derived stem cells (Pod+CD31+ cells),35,80 (iii) animal mesentery-derived stem cells (Pod+LYVE1+CD31+ cells),81–83 and (iv) renal stem and lymphoid lineage cells.30,84
From a clinical application perspective, these tissue sources do not pose of great value due to the invasiveness of the harvesting procedures, hence, attempts of standardizing these procedures in the clinic are quite improbable.
Experiment: Requirements for Lymphatic Vessel Development
There are important aspects of the lymphatic system's anatomy and physiology that must be taken into consideration when developing anatomically accurate lymphatic vessels in vitro by 3D bioprinting.
Stem cell requirements
Stem cells play a pivotal role in tissue engineering and 3D bioprinting, especially when the objective is to fabricate intricate structures like lymphatic vasculature. The use of terminally differentiated cells such as LECs may also be envisaged, but it entails the risk of adapting their cellular phenotype to expansion in cell culture. Instead, the selection of stem cells and their passaging tailored to this objective depends on several critical criteria, which determine their adequacy for the particular application. First and foremost, stem cells possess by definition a remarkable ability to self-renew and differentiate into daughter cell types. 85 Specifically, certain adult stem cell types exhibit the tendency for differentiating into LECs, as previously discussed.
When considering the use of adult stem cells in tissue engineering, it is crucial to take into account the phenotypic stability of differentiated cells. While no relevant tumorigenity has been detected in thousands of patients treated with diverse types of adult stem cells, it is also true that undifferentiated MSCs may adopt protumorigenic roles on the tumor microenvironment.86,87 For this reason, safety of potential lymphatic vessel transplants has to be guaranteed.
Furthermore, when developing tissue constructs derived from stem cells, cells with a high potential for expansion and proliferation are needed to achieve the required organ mass. 88 In addition, an important aspect to consider is the immunogenicity of stem cells. Avoiding immunogenic reactions to implanted grafts is primordial, making stem cells an attractive option due to their potential for autologous (patient-specific) use, thereby minimizing the risk of rejection. 89
Another crucial aspect related to the physiology of lymphatic vessels is their interaction with the surrounding ECM. Anchoring filaments play a central role in facilitating the entry of lymph into the vessels. 3 Therefore, stem cells intended for this application must not only have the capacity to adhere to the biomaterial scaffold but also possess the ability to modify the scaffold as needed to meet the functional requirements of the tissue. This aspect of modifying the scaffold ties directly into the functional integration of the lymphatic vessels, a key challenge in tissue engineering. 90 Achieving seamless integration with the implanted tissue is a critical challenge, and the adaptability of stem cells in modifying the scaffold contributes significantly to overcoming it.
Structural and morphological requirements
The lymphatic system is an extremely intricate system with enormous morphological and structural variation among the different tissues that conform it. Its high stratification plays a key role in maintaining the correct functioning, hindering the development of anatomically and physiologically accurate lymphatic vessels in vitro. In Table 1, we summarize the most important structural and morphological aspects to take into consideration to develop anatomically accurate vascular tissue constructs.
SMC, smooth muscle cell.
Hydraulic pressure, interstitial flow, and shear stress
The lymphatic system drains the fluid build-up from the interstitial space, resulting from the filtration of plasma in the capillaries, toward the vascular system. Several studies have investigated the nature of this phenomenon, as it is crucial for the homeostasis of the body as well as the morphogenesis and pathophysiology of the lymphatic system. 91 Under steady state, the hydraulic pressure of the interstitial fluid has been described to be slightly below atmospheric pressure (−5 to 0 cm H2O). 92 Due to the difficulty of the measurement, there is little information of lymphatic capillaries, but for more prominent lymphatic vessels (such as collecting lymphatic vessels), hydraulic pressures have been described to be positive.
These findings would suggest that the pressure gradient would oppose the lymphatic drainage; however, this does not occur. 93 Possibly, transient cyclical pressure changes the interstitial-intraluminal pressure balance, inducing a negative gradient favoring lymphatic drainage. Once the steady state is reached again, the gaps between the endothelial cells that compose initial lymphatic endothelium close, avoiding back-flow of the lymph.94–96
In addition to the extrinsic forces that initiate lymphatic drainage, there are intrinsic forces that arise from intraluminal pressures and the forces exerted by the lymphatic vessels, which enable lymph transport throughout the body. The lymphatic system, as the vascular system, has a lower pressure in the distal areas, and when the lymph flows toward the medial tissues, the pressure within the vessels increases. 97 Due to the nature of the lymphatic system, the extrinsic forces would not be able to overcome such pressures, and thus intrinsic forces come into play. On the one hand, the SMC layer periodically contracts, propelling the lymph.
Likewise, the lymphatic vessels are composed of lymphangions, which with the help of valves enable the flow from one to the other. So when the lymphatic smooth muscle contracts, the intraluminal pressure of the lymphangion overcomes the transvalve pressure, and propels the lymph from one lymphangion to the adjacent one. 98 The contractions in distal lymphatics range 2–4 cm H2O pressures, whereas in medial lymphatics, these increase up to 10–20 cm H2O. 99 Therefore, the interplay between the periodical contractions and the valvular system stratification enable the lymph flow against the gradient with little to no backflow.
Besides, interstitial flow arising from the Starling forces has been described to enhance lymphatic tissue regeneration. 100 Several studies have shown that before the formation of lymphatic capillaries, the interstitial tissue fluid accumulation leads to fluid flow, which promotes LEC migration to the area and morphogenesis of the capillaries. 101 In connection with interstitial flow, LECs also encounter varying levels of shear stress, dependent on their specific locations within the body and the conditions present. 102 Recent research has elucidated that the influence of shear stress on lymphatic physiology, affecting factors such as cell morphology and migration, is mainly driven by the dynamic flow patterns of shear stresses. 103 Several studies have reported that the elevated sensitivity to shear stresses is conferred by the presence of VEGFR3 on the cellular membrane of LECs. 104
In addition, collecting lymphatic vessels exhibit unique responses to shear stress, modulating nitric oxide release in response to shear stress and thus affecting SMC contractility. 105
To summarize, to develop fully functioning lymphatic vessels in vitro, it must be taken into great consideration that not all the lymphatic vessels are the same, but they differ from each other in a hierarchical manner. Likewise, the extrinsic and intrinsic forces that act upon them confer the functionality of the vessels. Therefore, the vessels must be able to absorb lymph from the interstitium, with the help of the anchoring filaments, as well as be able to propel said lymph using the peristaltic propulsion exerted by the SMC layer and the pressure gradients between contiguous lymphangions.
Advantages and Limitations of 3D Bioprinting Techniques
Lymphatic tissue is structured in a hierarchical manner, and its proper functioning relies on its anatomical arrangement. Among the available methods for creating engineered lymphatic tissue vessels in vitro, 3D bioprinting offers a range of advantages that make it most suitable for this purpose. Nonetheless, the effectiveness of 3D bioprinting depends on the specific technique used, as each approach presents advantages and limitations.
The inkjet bioprinting, also known as droplet-based bioprinting (DBB), is a bioprinting technique that utilizes thermal, piezoelectric, or electromagnetic technology to deposit bioink droplets and create the desired structures. These methods offer high cell viability in a rapid and cost-effective manner. 106 Inkjet bioprinting typically achieves a printing resolution ranging from ∼20–300 μm. However, it is crucial to maintain the bioink viscosity below 10 mPa/s because higher viscosities create excessive resistance to droplet formation, potentially leading to nozzle clogging.107,108 Nevertheless, this viscosity limitation can affect the structural integrity of the construct, particularly when patterning 3D vascular structures. 109 To prevent the collapse of subsequent layers, it requires rapid polymerization or structural support for the bioink, which reduces the choice of materials. Furthermore, the cell density within the bioink is an important consideration in DBB, as it influences droplet size and splitting time. 109
Although cell viability remains high with these methods, there are important considerations to take into account, especially with piezoelectric bioprinting. The frequencies used in piezoelectric bioprinting (15–25 KHz) have been reported to potentially harm cell membranes and lead to cell lysis. 110 In contrast, thermal inkjet bioprinters briefly reach very high temperatures (300°C). However, cellularized bioinks only sense a 4°C–10°C increase. 111 A significant issue associated with inkjet bioprinting is the relatively small droplet size. When not printed within a hydrophilic solution, these small droplets can lead to drying, resulting in cell dehydration. 112 In addition, the small droplet size, coupled with the reduced viscosity of the materials, can pose challenges when attempting to create intricate 3D structures.
Extrusion-based bioprinting utilizes pneumatic, piston, or solenoid-driven mechanisms to extrude bioinks through a nozzle onto a printing substrate. 113 This method is widely used due to its accessibility, compatibility with high-viscosity bioinks, and rapid multilayer printing capabilities. 114 The method is based sequential deposition of bioink filaments layer by layer to create a lattice-like macroporous structure. The technique allows the printing of larger high viscosity 3D constructs with enhanced mechanical integrity, although it poses a risk of shear stress-induced cell damage due to increased nozzle pressure. 115 Nevertheless, extrusion bioprinting generally offers lower resolution, typically exceeding 100 μm, hindering capillary-like construct formation. However, it is the most widely used method for vascular 3D bioprinting.
Despite resolution limitations, capillary networks are often formed within the filaments postprinting through vasculogenesis and angiogenesis, while larger vessel-like channels can be directly or indirectly printed. 116 The typical approach involves the fabrication of vascular channels by a sacrificial layer, which is later removed for its endothelialization. 117 In certain approaches, cells have been introduced into the sacrificial layer, where they attach to the inner surface of the tube once it is removed. 118 Supporting cell types such as pericytes, SMCs, and fibroblasts have been incorporated to enhance vessel stabilization and maturation.119,120 Furthermore, coaxial extrusion is well suited for microvasculature printing, enabling core-shell printing for rapid cross-linking and fabrication of perfusable tubular constructs. This approach offers precise control over channel size and composition, making it versatile for various applications. 121
Embedded 3D bioprinting offers a solution to challenges faced by conventional extrusion bioprinting methods. Extrusion bioprinted substrates are typically deposited in an open-air environment, limiting the complexity of printed structures and causing issues especially with soft hydrogel bioinks (with stiffness less than 100 kPa). Embedded bioprinting addresses these concerns by printing directly into a physical support matrix, providing structural support during printing, enabling omnidirectional extrusion, and reducing gravity-induced deformations. 122 The hydrated support matrix also helps maintain cell viability by controlling environmental factors such as pH, temperature, and sterility. 123 Thixotropic hydrogels, which change viscosity under shear stress and recover afterward, are ideal support matrices. Approaches utilizing this technique for creating vascular structures have relied on either depositing the bioink into a support bath that can be removed once the printing is complete, or depositing a sacrificial bioink into a support bath that, when removed, leaves the hollow cavities behind. 124
Light-assisted bioprinting utilizes a powerful light source to convert liquid bioink into the desired 3D structure and comprises two main categories: laser-based bioprinting and digital-light processing (DLP) bioprinting. The former can be divided into two subtypes: additive or subtractive. Laser-based bioprinting, also known as laser-induced forward transfer, involves a high-intensity laser, a ribbon containing the bioink medium, and a receiving substrate. 125 The laser focuses on a metallic plate within the ribbon, delivering sufficient energy to vaporize the bioink, which is then ejected as a droplet onto the receiving substrate. Typically, biopaper or a specific hydrogel serves as the support material. This method consistently achieves high cell viabilities (≥90%) due to precise control over parameters such as pulse energy, frequency, bioink viscosity, layer thickness, and donor-receiver distance, all of which influence resolution.126,127
The laser technology offers exceptional resolution (up to 50 μm), affording precise control over cell dispersion within printed structures and design flexibility. Further, subtractive laser-assisted bioprinting techniques are a commonly used techniques based on the photoablation of the substrate by a high-intensity laser, which when excited produces the etching of the material leaving a hollow space behind. It is a very interesting method for capillary bioprinting because the in situ photoablation of the materials allows to create capillary-like structures without the need for removal of sacrificial layers or postprocessing. However, photoablation techniques are quite slow in nature and limited to capillary-like networks, as to obtain a bigger vessel, higher intensity is required, which is inversely proportional to cell viability. 108
DLP bioprinting, or projection stereolithography (SLA), uses a digital-micromirror device to focus light onto a plate, polymerizing the bioink layer by layer. This technology allows rapid printing of high-resolution constructs, with resolution determined by the microscale focal size of each micromirror. 128 As the field has rapidly advanced, traditional SLA methods have evolved to incorporate two-photon or multiphoton polymerization techniques. This approach is particularly intriguing due to its exceptionally fine resolution, demonstrated in the creation of capillary-like structures with lumens ranging from 5 to 20 μm.129,130 Recently, volumetric bioprinting has emerged as a promising method for rapidly producing intricate structures. This technique involves projecting multiple 2D light patterns from various angles onto a container filled with photosensitive resin, gradually forming the desired construct. 131 However, despite its potential, this technique is relatively new and thus, progress in vascular 3D bioprinting remains limited. 132
Notably, the absence of mechanical forces during bioink extrusion in DLP methods leads to high postprinting cell viabilities, exceeding 95%. 133 Both laser-based bioprinting and DLP techniques can accommodate a wide range of bioink viscosities (1–300 mPa/s). 134 However, it is worth noting that the cost associated with both methods restricts their use to a select few research groups.
Discussion: Applications in 3D Bioprinting and Lymphatic Tissue Engineering
Considerable efforts have been put into developing functional tissue engineered constructs of many tissues and organs. However, engineered constructs cannot be fully viable unless they are provided of blood and lymphatic vascularization.4,135 Although attempts to produce vascularized constructs have been thoroughly studied, the lymphatic system has not received the same attention, thus, highlighting the imperative need for developing functional tissue engineered lymphatic vessels.
A major challenge in lymphatic tissue engineering is to reproduce the highly intricate morphology of the tissue structure and its cellular conformation. Most of the studies cited in this article have focused on cellular therapy for lymphatic etiologies, relying on the intricate biological cues of lymphatic progenitors to innately produce viable lymphatic vessels. Even though the results shown indicate a plausible path to follow, many challenges remain, some of which could be solved by combining the current knowledge with lymphatic tissue bioprinting and/or engineering approaches.
Tissue engineering encompasses the three main categories that make up tissues; these being, the cells, the supporting ECM and the biochemical cues. The typical tissue engineering approach would comprise a cell-loaded biomaterial in combination with prolymphangiogenic biochemical factors to trigger the growth and expansion of lymphatic vessels. The few scaffold-based tissue-engineering approaches taken, in vitro and in vivo, have mostly relied on the innate capacity of LECs to assemble naturally into lymphatic vessels. These have covered the main strategies in tissue engineering, such as coculturing of LECs with other cell types (i.e., fibroblasts or SMCs) 136 ; on culturing LECs in a 3D matrix composed of biocompatible scaffolds or hydrogels 137 ; decellularization of lymphatic vessels 138 ; or use of scaffold-free constructs. 78
Although the aforementioned techniques have attempted to recreate the complexity of the lymphatic vascular system, 3D bioprinting is a far more interesting approach mainly because of its repeatability, controllability, and reproducibility. 113 Due to its working principle, 3D bioprinting allows for creating 3D biomimetic tissue with specific positioning of multiple cell types and biochemical factors in a highly controlled architecture that resemble more accurately the structure of the native tissue. To be more specific, a significant challenge in lymphatic tissue engineering involves withstanding the mechanical stresses exerted by functioning tissues. Three dimensional bioprinting, as previously explained, offers the ability to precisely arrange cells within the ECM and customize scaffold properties to match native tissues. 108 Maturation is a crucial aspect of tissue-engineered constructs, particularly in the context of vessels such as lymphatic ones, where flow is essential.90,100 3D bioprinting enables the rapid flow of medium through printed tubular structures shortly after printing.
Moreover, when combined with the deposition of supporting cells, it has the potential to greatly enhance tissue maturation.119,120 It's worth noting that there are relatively few strategies focused on 3D bioprinting of lymphatic vessels, and this is an area of active research we are currently pursuing. One notable study created an organ-on-a-chip model of both blood and lymphatic vasculature using bioprinting. In this study, through a combination of microfabrication and additive manufacturing, they constructed a model consisting of four tightly sealed layers: two polydimethylsiloxane layers in the middle and two external poly(methyl methacrylate) layers. Within this system, bioprinted vascular and lymphatic vessels were embedded in a GelMA matrix with the aim of simulating the tumor microenvironment. 139
Lymphatic tissue engineering in all its extent could potentially provide new therapeutic approaches to many different diseases and conditions that affect millions of people. For instance, a wealth of data show that lymphangiogenesis plays an important role in the dissemination of solid tumors; however, the specific mechanism that enables such dissemination is poorly understood. In our view, a functional tumor microenvironment model containing vascular and lymphatic vessels as well as the key biomolecules would greatly benefit our understanding of tumor dissemination behavior. As another relevant example, postmyocardial infarction lymphangiogenesis occurs as means of providing immune cells to promote regeneration into the diseased area. 140 However, the influx of new lymphatic vessels leads to the malformation of collecting vessels resulting on a prolonged edema in the injury site. 141
In this case, lymphatic tissue engineering could help patients by providing fully operative lymphatic vessels and thus avoiding the malformation of collecting vessels. Other applications of lymphatic tissue engineering would be skin grafts. In this case, regardless of the ample research and knowledge on the topic, current skin grafts are far from native skin. Current skin engineering approaches have not included a fully functioning lymphatic vascular system. Finally, with the discovery of the lymphatic system in the brain, many neurological diseases have been described to interplay with the lymphatic system (Alzheimer's disease and Multiple Sclerosis among others).142,143 The interested reader is referred to a recent review by Alderfer, Hall, and Hanjaya-Putra that further explains the most recent advances in lymphatic models to study immune cell interaction and drug delivery through the lymphatic system. 144
Clinical Applications
As described above, there are many different tissues that can be exploited for sourcing stem cells; however, their clinical applicability depends on practical and logistic issues as well as their in vitro behavior.145,146 Even though adult stem cells are ubiquitously spread along the body, some applicability limitations lie beyond the potency or the differentiation yield of the cells, such as the invasiveness of the extraction procedure or donor-site characteristics. Of the previously described tissue sources, the most reliable for clinical purposes are the bone marrow, adipose tissue, and dermis.
Bone marrow-derived MSCs pose great regenerative value for cellular therapies; nonetheless, the harvesting of these cells may lead to excruciating prolonged donor-site pain, potential infection, and tedious recovery. Therefore, in comparison with other tissues that require less invasive procedures and better recoveries, such as adipose tissue, the bone marrow is in disadvantage. 145 Another source for stem cells is the dermis, in this case, the cells could be harvested from the human dermis from children's foreskin or from the by-product of skin surgeries. Postnatal circumcision is an extended surgical procedure that is performed on daily basis, nonetheless, this allogeneic cell source could induce some immunological response, that in fact, could induce a detrimental effect rather than a healing one. In addition, skin grafting is a very invasive technique that should be reconsidered as the cellular quantities, and viability obtained from this tissue source is quite scarce.
On the contrary, adipose-derived stem cells are advantageous to other tissue types, as adipose tissue is a copious by-product of many therapeutic and cosmetic procedures. Excluding the abundance and accessibility of the tissue, ADSCs also present favorable therapeutic characteristics. For instance, ADSCs are suitable for long-term subcultures, possess high multilineage potential, and eagerly expand in vitro. 147 Moreover, ADSCs behave as host-response modulators exhibiting a greater immunomodulatory capacity than BM-MSCs in vitro, as they suppress the cytotoxic activity of natural killer cells and the differentiation of monocyte-derived dendritic cells, besides limiting the proliferation of lymphocytes.148–150 The main limitation regarding adipose tissue is their dependency toward certain donor characteristics (i.e., age) as their multilineage potential and their proliferative capacity are known to be compromised, at least in vitro. 151
A recent study characterized the potential of dermal MSCs versus ADSCs in wound healing with interesting results. They compared the proliferative capacity, stemness, isolation efficiency, and genetic stability in culture, as well as the capacity of wound closure and their paracrine efficiency on fibroblasts and keratinocytes. 152 Their results demonstrated that even though both cell sources had low mutation rates over culture time and showed similar stem cell characteristics; ADSCs were more efficiently isolated (100 × better), were more proliferative in vitro, and induced a better paracrine effect. 152 Hence, taking these results into consideration, we conclude that from a clinical and practical perspective, adipose-derived stem cells hold greater power and are in fact more adequate to be used for cellular therapy and regenerative medicine than the rest of the tissue sources.
In addition, conventional in vitro tissue models have been confined to a 2D microenvironment, which has been valuable for enhancing our general understanding but falls short in representing in vivo conditions accurately. On the contrary, while conventional 3D cultures offer improvements by better replicating in vivo structures, they still have limitations in terms of incorporating multiple cell types and faithfully recapitulating the unique spatial arrangement of the ECM. 153 Bioprinting emerges as a solution to address these unmet needs. Three dimensional bioprinting shows great promise for advancing the clinical applications of LEPCs in several ways.
An important challenge in drug development stems from inadequate models that fail to accurately represent human diseases. 154 One significant enhancement that can be introduced into advanced 3D disease models is the incorporation of a lymphatic system. This addition not only would help in better understanding the origins of various diseases but also contribute to the development of more effective treatments. Further, bioprinting enables the precise and intricate fabrication of 3D scaffolds, potentially mimicking the natural microenvironment of lymphatic tissue. 120 These scaffolds could offer structural support for the growth and organization of LEPCs, enabling the development of functional lymphatic networks.
Conclusions
Lymphatic tissue engineering has tremendous potential for a variety of applications, such as tissue and organ replacement, disease modeling, drug testing and biologically relevant modeling. Nevertheless, its development is in need of adequate in vitro models of lymphatic vasculature. Lymphangiogenesis in the adult occurs in pathological states, giving rise to newly formed lymphatic vessels from preexisting ones. Likewise, several studies have described the process of de novo lymphatic vessel formation arising directly from stem cells, called lymphvasculogenesis. Nevertheless, there are many gray areas in the literature regarding the formation of lymphatic vessels and their implications in different pathologies. Stem cell-based therapies have gathered significant attention in recent years. Hinging on their potential, research has covered the potential sources for LEPCs in vivo. In this review, we have discussed the potential tissue sources for LEPC isolation with a focus on their future use on 3D bioprinting.
In addition, we have also discussed the specific requirements for lymphatic tissue engineering, and the adequacy of bioprinting as a potential solution to lymphatic vessel deficiencies. Besides, we have analyzed the potential clinical application of the different tissue sources discussed. In brief, adipose tissue- and bone marrow-derived LEPCs have demonstrated their contribution to lymphangiogenesis through different routes. While both of them possess the capacity of directly differentiating into LEPCs, they also contribute through unique pathways, these being paracrine signaling of prolymphangiogenic factors for the former and macrophage polarization for the latter. Other tissues, such as the umbilical cord blood or the dermis, have also been considered. Several studies have described the presence of a small subset of LEPCs in the EPC and ECFC found the blood; however, few studies have investigated their potential for this application.
Dermis-derived LECs have become a practical choice for in vitro studies; nonetheless, their translation to in vivo models has been limited. Furthermore, the ample variety of 3D bioprinters available along with their unique benefits make them a very interesting method for tissue engineering of lymphatic vessels. Even though some techniques are restricted to methodological limitations, such as extrusion bioprinting on developing complex capillary networks, different 3D bioprinting methods can be used in combination to meet the requirements. In conclusion, there is a considerable gap in lymphatic tissue engineering, and its exploitation can benefit a wide variety of applications.
Authors' Contribution
I.A.A. drafted the original article. I.J., H.L., and A.I. revised it and contributed corrections and discussion. All authors approved the final version before submission.
Footnotes
Disclosure Statement
The authors state no conflict of interest.
Funding Information
I.A.A. is supported by a predoctoral fellowship from Fundación Jesús Gangoiti Barrera. Work related to this review is covered by project 2020111004, funded by the Department of Health of the Basque Government, and projects PI22/01247 and PT20/00030, funded by Instituto de Salud Carlos III (ISCIII), and cofunded by the European Union.