
Abstract
Subchondral drilling (SD), a bone marrow stimulation technique, is used to repair cartilage lesions that lack regenerative potential. Cartilage repair outcomes upon SD are typically fibrocartilaginous in nature with inferior functionality. The lack of cues to foster the chondrogenic differentiation of egressed mesenchymal stromal cells upon SD can be attributed for the poor outcomes. Continuous low-intensity ultrasound (cLIUS) at 3.8 MHz is proposed as a treatment modality for improving cartilage repair outcomes upon marrow stimulation. Bilateral defects were created by SD on the femoral medial condyle of female New Zealand white rabbits (n = 12), and the left joint received cLIUS treatment (3.8 MHz, 3.5 Vpp, 8 min/application/day) and the contralateral right joint served as the control. On day 7 postsurgery, synovial fluid was aspirated, and the cytokine levels were assessed by Quantibody™ assay. Rabbits were euthanized at 8 weeks and outcomes were assessed macroscopically and histologically. Defect areas in the right joints exhibited boundaries, incomplete fill, irregular cartilage surfaces, loss of glycosaminoglycan (GAG), and absence of chondrocytes. In contrast, the repaired defect area in the joints that received cLIUS showed complete fill, positive staining for GAG with rounded chondrocyte morphology, COL2A1 staining, and columnar organization. Synovial fluid collected from cLIUS-treated left knee joints had lower levels of IL1, TNFα, and IFNγ when compared to untreated right knee joints, alluding to the potential of cLIUS to mitigate early inflammation. Further at 8 weeks, left knee joints (n = 12) consistently scored higher on the O'Driscoll scale, with a higher percent hyaline cartilage score. No adverse impact on bone or change in the joint space was noted. Upon a single exposure of cLIUS to TNFα-treated cells, nuclear localization of pNFκB and SOX9 was visualized by double immunofluorescence and the expression of markers associated with the NFκB pathway was assayed by quantitative real-time polymerase chain reaction. cLIUS extends its chondroprotective effects by titrating pNFκB levels, preventing its nuclear translocation, while maintaining the expression of SOX9, the collagen II transcription factor. Our combined results demonstrate that healing of chondral defects treated with marrow stimulation by SD can be accelerated by employing cLIUS regimen that possesses chondroinductive and chondroprotective properties.
Impact statement
Repair of cartilage represents an unsolved biomedical burden. In vitro, continuous low-intensity ultrasound (cLIUS) has been demonstrated to possess chondroinductive and chondroprotective potential. To our best knowledge, the use of cLIUS to improve cartilage repair outcomes upon marrow stimulation, in vivo, has not been reported and our work reported here fills that gap. Our results demonstrated enhanced cartilage repair outcomes under cLIUS (3.8 MHz) in a rabbit model of subchondral injury by subchondral drilling. Enhanced repair stemmed from mesenchymal stem cell differentiation in vivo and the subsequent synthesis of articular cartilage-specific matrix.
Introduction
Damaged cartilage has limited potential for self-repair, thus necessitating repair and restoration strategies.1–6 Current treatment options that include autologous chondrocyte transplantation and implantation of tissue-engineered substitutes are expensive and of limited efficacy.7–9 From a clinical perspective, bone marrow stimulation techniques such as microfracture (MF) or subchondral drilling (SD) are widely practiced; however, outcomes are characterized by functionally inferior fibrocartilage.10,11 This is postulated to result from the following: (i) Instability of the blood clot formed during marrow stimulation; (ii) lack of chondroinductive factors near the vicinity of the defect, and (iii) a cytokine-rich proinflammatory joint environment postsurgery that likely impedes the efficient chondrogenesis of egressed mesenchymal stem cells (MSCs).12–14
To improve the efficacy of MF and SD, methods that combine marrow stimulation with orthobiologic injections or improving clot stabilization using synthetic matrices, scaffolds have been evaluated and shown to improve cartilage repair.15–20 However, clinically translatable methods to improve repair outcomes that rely on the marrow stimulation must promote the chondrogenesis of egressed MSCs in vivo, potentially in a proinflammatory joint environment postsurgery. 13
Ultrasound (US) is widely used in diagnostic and therapeutic applications and has the potential to complement existing cartilage repair therapies like MF or SD, and offers the prospect of significant clinical impact.21–23 While pulsed US (pLIUS) or continuous low-intensity ultrasound (cLIUS) has been shown to be an effective preconditioner of MSC constructs in vitro,24–29 and enhanced immunohistochemical (IHC) and mechanical properties upon the subcutaneous implantation of MSC constructs, in vivo, in a nude mouse model,30,31 there is a lack of published work investigating the ability of low-intensity continuous US (LIUS, 1–50 mW/cm2) to improve cartilage repair outcomes in vivo.
Current methods that employ low-intensity-pulsed-ultrasound (LIPUS) remain unclear because they have conflicting outcomes. For example, most in vitro or in vivo work that employs a pulsed sine wave at 1.5 MHz frequency (termed as LIPUS) to improve cartilage repair outcomes has reported conflicting outcomes.32–35 For example, when LIPUS at 1.5 MHz was employed in a canine model to improve interfacial cartilage repair of autologous osteochondral plugs, a persistent cleft remained.33,34 In a 2014 study, the LIPUS applied at 1.5 MHz failed to repair a full-thickness defect in a rabbit model. 32
In a departure from current approaches, our work focuses on identifying the in vivo beneficial frequency bandwidth where US-induced bioeffects are maximized.36–39 Using theoretical modeling as detailed elsewhere, an in vivo beneficial frequency bandwidth for chondrocytes in native cartilage was identified to be between 3.8 ± 0.3 MHz. 39 Furthermore, our published aggregate in vitro data demonstrated the chondroinductive ability of cLIUS where cLIUS was shown to induce MSC chondrogenesesis without the use of growth factors, enhanced the biosynthetic activity of MSC-derived chondrocytes and adult chondrocytes, and preserved MSC chondrogenesis in a proinflammatory environment.24,28,29,40,41
In summary, combined theoretical modeling and in vitro findings suggest that cLIUS applied at the in vivo beneficial frequency bandwidth will complement and improve repair outcomes in procedures that employ marrow stimulation for an endogenous source of MSCs. Our working hypothesis is that an optimized delivery of LIUS to a rabbit model of chondral injury by SD will significantly improve cartilage regeneration. The objective of this article is to demonstrate the efficacy of cLIUS when applied at 3.8 MHz in improving SD outcomes in a rabbit model of chondral injury.
Materials and Methods
Rabbit cartilage defect model
Bilateral defects were created on knees of the rabbits using an IACUC-approved protocol (1297) at the University of Nebraska-Lincoln (Lincoln, NE). To exclude gender-based differences in animal response to injury and repair, female New Zealand Rabbits (white), less than 8 months of age with a body weight of 4 kg or less, were purchased and acclimatized for about a week before surgery. Rabbits were anesthetized by the administration of ketamine IM (35 mg/kg) and xylazine (5 mg/kg). Gas anesthesia was maintained using a nose-cone supplying 2–3% isoflurane.
Rabbits were laid in supine position on the operating table and the knee was shaved and sterilized using betadine and 2% chlorhexidine. A medial parapatellar approach was used to enter the knee joint placed in flexion of 70° and the patella was dislocated laterally to expose the articular surface of the distal femur. The defect was created on the medial femoral condyle as follows: A 5-mm diameter, full-thickness cartilage defect was created using a biopsy punch (Miltexs, York, PA) and a curette was used to remove the cartilage and the calcified layer thoroughly.
Marrow stimulation was performed to a 3-mm depth using a surgical drill mounted with 0.9-mm K-wires that were premarked to create three 1.5-mm diameter holes in each defect site, with each hole being 2 to 2.5 mm apart on the subchondral bone (Fig. 1A). Our rationale for going 3-mm deep is based on a previous study that demonstrated a 3-mm deep hole significantly enhanced the volume of blood (∼150 μL) that could ooze from the bone marrow without penetrating the epiphyseal scar (Fig. 1B). 42 Saline was continuously flushed to minimize the heat caused by drilling. The joint was then irrigated using saline and inner dermis was sutured using biodegradable suture, while the outer dermis was closed using nylon sutures and/or metal staples, which were removed after a week. Animals were monitored for complications by research and veterinary staff.
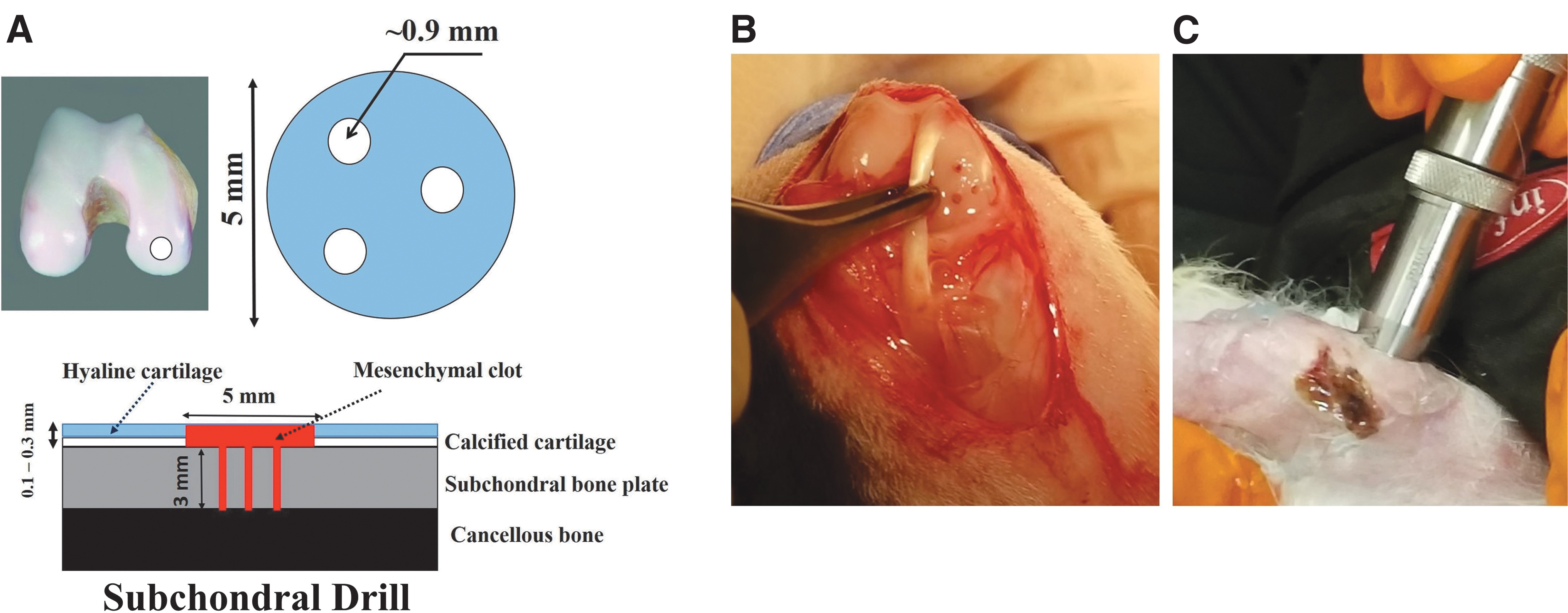
Subchondral drilling technique in a rabbit model.
Animal housing and care
Upon surgery, all animals received pain medication (buprenorphine SR) during the surgical procedure and meloxicam every 24 h for at least 2 days. After 2 days, if the animals showed any of the signs of pain or distress, buprenorphine SR or meloxicam was administered on an as-needed basis. As an antibiotic, enrofloxacin was given prophylactically once daily, starting intraoperatively and then 3 days postoperatively. Rabbits were all housed together on the floor and were allowed unrestricted activity postoperatively. All rabbits were analyzed in a long-term study for 8 weeks. A total of 12 animals were included in these studies.
US application procedure
US was applied at 3.8 MHz using a nonfocused transducer (V384/Serial No. 1118345; Olympus NDT, Inc., MA) with a radiating area of 1.27 cm2 at duty cycle of 100% (Fig. 1C). All transducers were characterized by the manufacturer before usage and tested at UAH using a custom setup at the start, at random time points, and at conclusion of the experiment to ensure transducer performance. A waveform generator (HP, 33120A) was used to generate LIUS at the required pressure amplitude of 20 kPa at 3.8 MHz.
The transducer was positioned on the skin such that the transducer face was approximately in line with the defect area. Coupling gel was applied to the skin. In the absence of exhaustive clinical literature from which to make absolute estimations of treatment time, current literature justifies 1 min of US energy per treatment head area covered. 43 Thus, US was applied for 8 min/day for 5 days/week beginning on postoperative day 3. The sites were periodically shaved to ensure contact between the transducer, coupling gel, and skin, and the rabbits were held in a baby carrier to assist with the application of LIUS.
At 8 weeks following defect creation, rabbits were anesthetized and euthanized as per the approved IACUC protocol. Following euthanasia, both stifle joints were harvested to allow visual inspection, X-ray analyses, and histopathologic testing of the repair tissue, including the surrounding cartilage, and similar tissues from the contralateral limb.
Gross evaluation
After euthanasia, bilateral knees were harvested, and the knee joint opened to expose the surgical sites for gross evaluation and digital imaging. Gross analysis included noting the general appearance of the cartilage surface.
Assessment of bone
Anteroposterior (AP) and lateral radiographs (LP) of each knee joint were obtained after euthanasia. The kilovoltage peak was 60 and the milliampere-seconds was 1. Digital X-rays were taken from a distance of 2 feet using a portable X-ray unit (EPX-F2800; ECOTRON Co., Ltd.).
Histological and immunochemical evaluation
Joints were fixed in 10% formalin and processed for histology using standard procedures: sectioned and stained with hematoxylin and eosin (H&E) and Safranin-O stains to indicate proteoglycan content across interfaces and independently graded by Dr. Oraine Snaith and Dr. Gene Siegal using the O'Driscoll scoring system. The scorers were blinded to the identity of samples. Sections were also evaluated using IHC stain for chondrogenic (collagen II [COL-II]) with specific antibodies (3H1-F9, 1:200 dilution; Novus Biologicals) using standard IHC protocols.
Analyses for proinflammatory cytokines in the synovial fluid
Synovial fluid was collected on day 7 postoperatively and analyzed by a customized Quantibody™ Rabbit Cytokine Array (RayBiotech, GA).
MSC culture in monolayer and cLIUS treatment
Human MSCs were purchased from Lonza (PT-2501; Walkersville, MD) and expanded in alpha-minimum essential medium supplemented with 10% MSC-qualified fetal bovine serum, 1 × Glutamax (Gibco), and 1 × antibiotic-antimycotic solution (Gibco) in a CO2 incubator at 37°C, 5% carbon dioxide, and 99% humidity. MSCs harvested from passage 4 to 5 were employed in all experiments.
MSCs were plated in six-well tissue culture plates (TCPs) at following seeding densities: 5 × 104 cells/well (RNA extractions) and 1 × 104 cells/coverslip/well (for immunofluorescence studies). All treatments with cytokines and/or cLIUS were conducted after 48 h of initial seeding of MSCs in TCP. The following study groups were employed: control (−cLIUS, −TNFα), cLIUS treated (+cLIUS, −TNFα), cytokine treated (−cLIUS, +TNFα), and cLIUS in the presence of cytokine (+cLIUS, +TNFα). Designated samples were exposed to 10 ng/mL of TNFα (R&D Systems, Minneapolis, MN) and simultaneously stimulated with a cLIUS regimen of 5 MHz (2.5 Vpp), with a constant pressure amplitude of 14 kPa for 10 min using an US-assisted bioreactor and a procedure detailed elsewhere.28,44,45 Upon completion of the experiment, cells were released for RNA analyses as described below.
Quantitative real-time polymerase chain reaction
Cells were released from TCPs using incubation with 300 μL of Trizol reagent per well. Homogenates from two wells served as one replicate and three such replicates were used for gene expression analysis (n = 3). RNA was extracted using PureLink RNA Mini Kit (Thermofisher) according to the manufacturer's protocol. The quantitative real-time polymerase chain reaction (qRT-PCR) analysis was carried out using Quantstudio 3 real-time PCR system (Applied Biosystems) using TaqMan® RNA-to-CT™ 1-Step Kit (Life Technologies) according to the manufacturer's guidelines.
TaqMan Gene expression assays (Life Technologies) used are as follows:
GAPDH (Hs02786624_g1), MMP13 (Hs00942584_m1), ADAMTS4 (Hs00192708_m1), NFκB (Hs00765730_m1), and SOX9 (Hs00165814_m1). The expression of mRNA transcripts was normalized to GAPDH expression and relative expression levels were calculated using the 2−ΔΔCt method.
Double immunofluorescence staining
MSCs on cover slip (CS) were fixed in 4% paraformaldehyde for 20 min and blocked with 2% goat serum in 1 × TBST (Tris buffer saline with 0.1% tween20) blocking buffer (BB) for 2 h. CS were then incubated with 1:1000th diluted rabbit anti-phospho-NFκB p65 monoclonal antibody (MA5-15160; Invitrogen) in BB overnight at 4°C. Upon washing, CS were incubated with 1:1000th diluted goat anti-rabbit IgG H&L conjugated with Alexa flour 488 (150077; Abcam) for 1 h at room temperature (RT).
For double immunofluorescence (IF) staining, CS were washed and blocked with BB for 1 h at RT and incubated with 1:1000th diluted rabbit anti-human SOX9 Mab conjugated with Alexa fluor 647 (71273; CST) overnight at 4°C. Upon washing, CS mounted on a glass slide with DAPI mounting media (P36962; ProLong™ Diamond Antifade Mountant with DAPI). Zeiss LSM 700 confocal microscope at 63 × magnification was used for imaging. ImageJ™ software (n = 30) was used to quantify fluorescent intensities.
Sample size determination and statistical analysis
Power analyses were based on published mechanical property and protein expression data obtained in vitro under cLIUS. 28 The chosen effect size was based on a 20% improvement in the modulus over the control samples and a standard deviation of ∼10% of the mean of the control samples. For example, a sample size of 6 achieves 80% power to detect an effect size of 1.4 with a significance level (alpha) of 0.05 using a paired two-sided t-test. Bilateral defects were created and each defect was considered an independent sample.
Twelve rabbits were included in this study (n = 12). O'Driscoll histological scoring of Safranin-O images obtained from rabbit joints was analyzed using Welch t-test. Significant differences in gene expression and IF image intensities were evaluated with one-way ANOVA followed by post-hoc Sidak's multiple comparison test. Statistical significance was established as follows: p < 0.05 (denoted as *), p < 0.01 (denoted as **), p < 0.001 (denoted as ***), and p < 0.0001 (denoted as ****). Data are expressed as mean ± standard deviation.
Results
In vivo evaluation of cLIUS in a rabbit model of full-thickness articular cartilage defect created with SD
Bilateral defects were created on the femoral medial condyle and left knee joint was subjected to the cLIUS treatment where the contralateral right joint served as the control (Fig. 1). During defect preparation, immediate bleeding from the underlying subchondral bone plate was always observed after drilling (Fig. 1B). When the animals were euthanized at 8 weeks postoperatively, no joint effusion, macroscopic inflammation, periarticular osteophyte, and adhesion was noticed in all treated joints. Three rabbits developed swollen joints postsurgically, which were managed by draining the synovial fluid and administration of approved analgesics. Upon the completion of the 8-week study, all the rabbits appeared to have full range of motion in both the operated knees and none of the rabbits developed apparent joint degradation or synovial inflammation.
After euthanasia, joint capsule was opened, photographed, and evaluated macroscopically (Fig. 2). In the nonstimulated control joints, defects were still obvious, with defect boundaries still visible, exhibiting noticeable color and texture differences within the surrounding cartilage (Fig. 2B). In contrast, the left knee joint that received cLIUS treatment showed uniform tissue fill in most defects with similar color and texture within the surrounding cartilage (Fig. 2D).
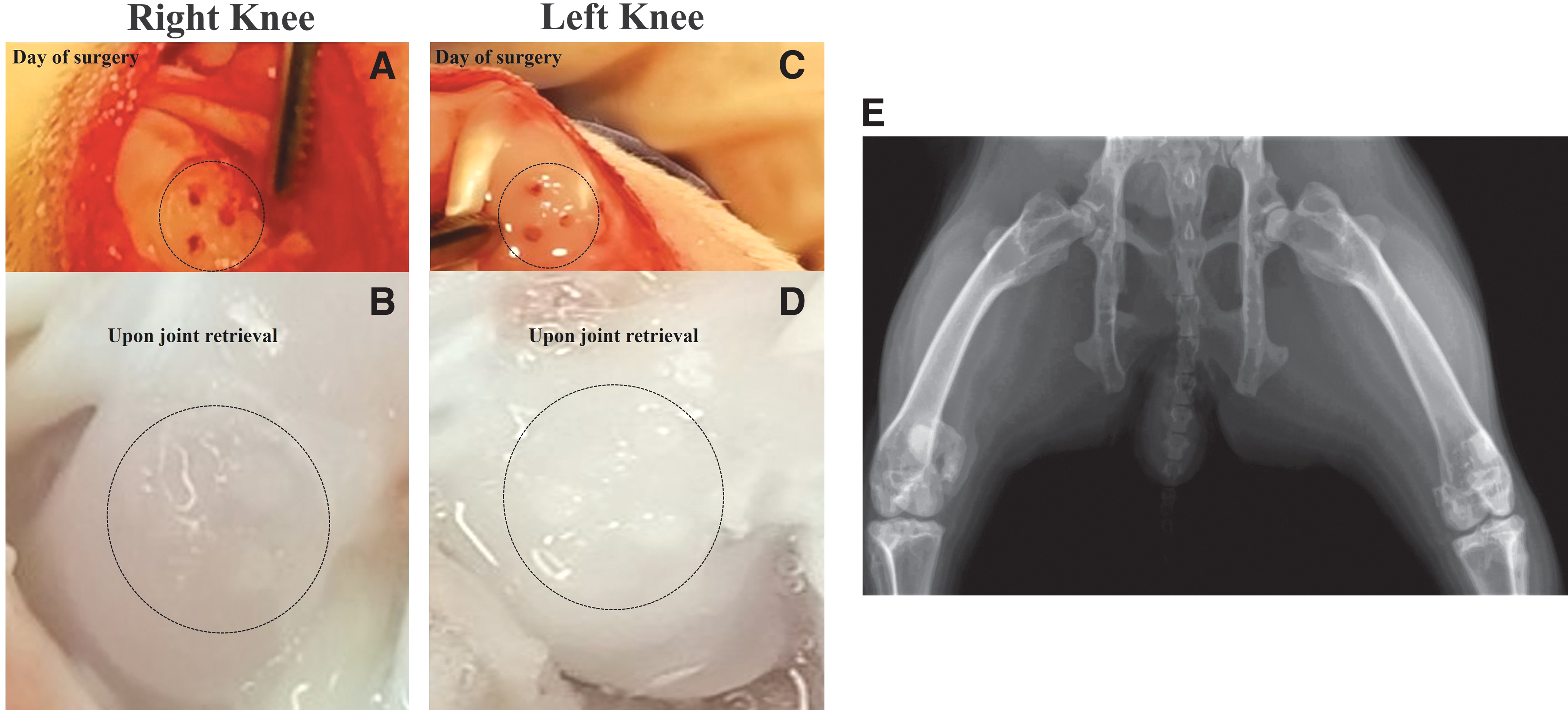
Gross visualization of the defect area. Rabbit-matched images of the defect areas are shown. Defect area is shown by a dotted circle.
Macroscopic evaluation indicated integration of the engineered tissue with the native cartilage. Previous research showed that therapeutic doses of US (<500 mW/cm2) did not have an adverse effect on bone growth or led to degeneration of cartilaginous cells in the growth plate. 46 AP and LP of each knee joint were obtained after euthanasia and analyzed. As anticipated, no adverse impact on the bone or change within the joint space was noted under cLIUS (<30 mW/cm2 at 3.8 MHz and 3.5 Vpp) employed in this study (Fig. 2E).
Representative rabbit matched staining for H&E, Safranin-O, and IHC for COL-II are shown in Figure 3. Qualitative assessment of H&E-stained histological sections of nonstimulated right knee joints revealed incomplete filling of defects, loss of cellularity, and increase in subchondral bone density as a result of the reparative process and, in some cases, an irregular cartilage surface was noted (Fig. 3A). Little to no staining for glycosaminoglycans (GAGs, Safranin-O) or type II collagen was observed in the defect area of the right knee joints (Fig. 3B, C).

Histological evaluation of rabbit knee joint sections at 8 weeks. Upon joint retrieval, rabbit joints were fixed in 10% formalin and processed for histological analyses using standard procedures. Left knee joints were treated with cLIUS and the right knee joints served as the contralateral nonstimulated control. Four to 6 μm sections were stained and presented. Images (frontal sections) at 2 × and 20 × magnifications are included and scale bars are shown in the images.
In some sections, a loss of Safranin-O staining in the superficial layer of the surrounding native cartilage was noted, indicating a loss of the extracellular matrix components. Defects in the left knee joints treated with cLIUS showed a complete fill level to the surrounding native cartilage and a positive staining for GAGs matched the surrounding undamaged cartilage (Fig. 3D, E). In addition, a rounded chondrocyte-like morphology, columnar organization, and some cell clustering were observed. IHC staining for COL-II on the left knee joints strongly highlighted the chondrocytes in the superficial and the middle or transitional zones with lesser staining in the deep radial zone. Some nonspecific staining surrounding the vascular elements was noted (Fig. 3F).
In comparison, IHC staining for collagen II of the right knee joint indicated a moderate absence of chondrocytes, some clustering and significant fissuring. Quantitative scoring of histological sections stained with Safranin-O was performed and summarized in Table 1 and the corresponding O'Driscoll histological score plot is shown in Figure 3G. Histologic scores of 8-week in vivo samples showed significantly higher scores for the groups (left knee joints) treated with cLIUS at 3.8 MHz when compared to untreated right knee joints. Synovial fluid was collected on day 7 postoperatively and analyzed by a customized Quantibody Rabbit Cytokine Array (RayBiotech), and shown in Figure 3H. Synovial fluid collected from cLIUS treated left knee joints demonstrated lower levels of IL1, TNFα, and IFNγ when compared to the synovial fluid-untreated right knee joints.
Histological Scores for Continuous Low-Intensity Ultrasound-Treated Joints and Noncontinuous Low-Intensity Ultrasound-Treated Contralateral Controls at 8 Weeks
cLIUS, continuous low-intensity ultrasound.
In vivo, early cartilage reparative processes occur in an inflammatory environment 13 ; thus, to provide an explanation for the in vivo observations reported—the ability of cLIUS to downregulate NFκB and matrix degrading proteins was evaluated in an in vitro study using hMSCs as it was not feasible to isolate rabbit MSCs that underwent surgery due to IACUC stipulations. Nevertheless, rabbit bone marrow-derived MSCs have been reported to possess a similar phenotype and chondrogenic differentiation ability to hMSCs.47,48 Catabolic cytokines activate NFκB, leading to the downregulation of SOX9, the main collagen transcription factor. 49 To demonstrate the ability of cLIUS to support the chondrogenic differentiation of MSCs in the presence of TNFα (i.e., one of the cytokines noted in the synovial fluid), the gene expression of catabolic markers (NFκB, MMP13, and ADAMTS4) and chondrocytic lineage markers (SOX9) was evaluated by qRT-PCR, and this is shown in Figure 3.
cLIUS downregulates catabolic and upregulates anabolic gene expression in MSCs exposed to TNFα
When compared to controls, inclusion of TNFα led to an upregulation in the expression of catabolic genes and NFκB and gene downregulation of SOX9 (Fig. 4). cLIUS stimulation abrogated the expression levels of catabolic genes, notably in the presence of TNFα. TNFα-treated groups that included cLIUS yielded higher gene expression levels of SOX9 and were noted to be similar to cLIUS-only treated samples. 24 Experiments in the presence of IL1β or IL6 were carried out and similar trends were noted (data not included). Collectively, it was observed that cLIUS promoted the gene expression of anabolic markers in the presence of proinflammatory cytokines by downregulating catabolic genes MMP13, ADAMTS4, and NFκB.

Gene expression analysis in MSCs exposed to proinflammatory cytokine. MSCs were seeded on TCPs and the treatment groups are as indicated. Homogenates from two wells per group served as one replicate and three such replicates were used for gene expression analysis (n = 3). Total RNA was extracted, and the gene expression of indicated biomarkers was evaluated by qRT-PCR, and GAPDH was used as a housekeeping gene. Bar graph represents mean ± 95% confidence interval; p-values indicate statistically significant differences (**p < 0.01; ***p < 0.001; and ****p < 0.0001). MSC, mesenchymal stem cell; qRT-PCR, quantitative real-time polymerase chain reaction; TCP, tissue culture plate.
cLIUS rescinds NFκB and promotes SOX9 localization to the nucleus in MSCs exposed to TNFα
The downstream transcriptional activity of pNFκB is linked to its nuclear localization. Thus double IF was used to visualize the presence of pNFκB and SOX9 in the nuclear regions and the corresponding fluorescent intensities were quantified and shown in Figure 5. In the presence of TNFα, increased levels of pNFκB and reduced levels of SOX9 in the nucleus were noted when compared to control and cLIUS samples. However, inclusion of cLIUS stimulation inverted the localization of these markers, the levels of nuclear pNFκB were decreased and SOX9 was upregulated. Analyses of cytoplasmic fluorescence levels of pNFκB and SOX9 showed similar trends (data not included). These results show that cLIUS maintains the expression of SOX9 in the presence of proinflammatory cytokines.
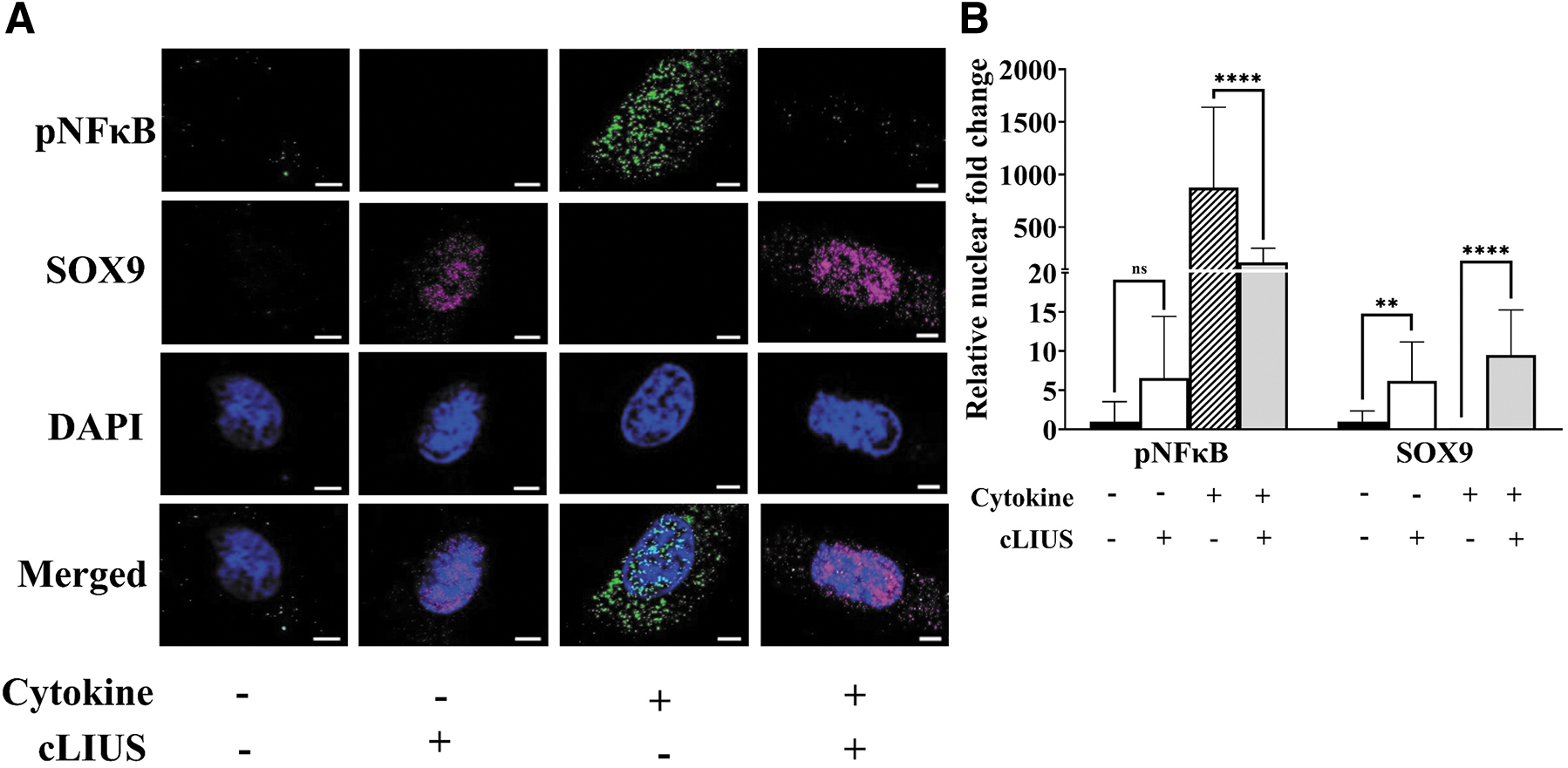
Localization of SOX9 and pNFκB in MSCs. MSCs were seeded on coverslips and the treatment groups are as indicated. Coverslips were fixed and double stained for pNFκB (green fluorescence) and SOX9 (red fluorescence) antibodies, and nuclei were counter stained with DAPI (blue fluorescence).
The combined results demonstrate that healing of chondral defects created by SD can be accelerated by employing cLIUS regimen that possesses chondroinductive and chondroprotective properties.
Discussion
The limited repair capacity of articular cartilage and subsequent degeneration of chondral injuries often lead to osteoarthritis. Bone marrow stimulation technique such as SD or MF is a widely practiced single-stage surgical procedure for the repair of cartilage defects and it is cost-effective with low patient morbidity, and involves the patients' endogenous MSCs as a cell source to stimulate cartilage repair.10,11,50 The reparative process in marrow stimulation techniques involves a clot of pluripotent MSCs that adhere to the subchondral bone. Successful outcome upon marrow stimulation is dependent on the efficacious differentiation of MSCs into a chondrogenic lineage. Our published data show that cLIUS is chondroinductive and enables chondrogenesis in the absence of exogenously added growth factors (i.e., TGFβ3); thus, cLIUS can act as an efficacious primer of MSCs. 24 Also, no upregulation of osteogenic or adipogenic markers (i.e., RUNX2, COL1A1, or PPARγ) was noted by RT-PCR or IHC.
Furthermore, cLIUS aids in the maintenance of the chondrocyte phenotype. Our promising in vitro findings regarding the beneficial effects cLIUS has on the promotion of chondrogenic differentiation and healing at the vertical interface in a full-thickness chondral defect model present an intriguing avenue of addressing this problem.24,40,41 Translation of these findings to an animal model and being able to demonstrate that a relatively noninvasive adjunct, such as cLIUS, can create and promote hyaline repair tissue. This presents a potential treatment modality for orthopedic surgeons attempting to treat patients with this challenging clinical problem.
Previous therapies with pLIUS @ 1.5 MHz failed to repair chondral defects in animal models.32–34 To progress beyond the status quo that employs pLIUS at 1.5 MHz for in vivo applications, 33 our approach was first to employ cLIUS as it couples more energy when compared to pLIUS. 44 Second, our work shifted the focus to the identification of a computationally proposed in vivo beneficial frequency bandwidth for US-induced cartilage restoration.
Briefly, theoretical models detailing the interaction of cells with cLIUS by taking into account the cellular and tissue properties were developed. 39 By doing so, the focus was shifted from an empirical approach to the identification of a beneficial frequency bandwidth where LIUS-induced bioeffects are maximized. Biot theory that helped establish the frequency dependence under cLIUS 37 was used to estimate the in vivo beneficial frequency bandwidth of chondrocytes in their native environment by considering the depth-dependent mechanical properties of the extracellular matrix and pericellular matrix.
Using a range of mechanical properties available in literature and a parametric analysis to understand the sensitivity of the estimated frequency values, in vivo beneficial frequency bandwidth for chondrocytes in native healthy cartilage was estimated to be between 3.8 ± 0.3 MHz. 39 Thus cLIUS stimulation 3.8 MHz was employed in rabbit studies. Moreover, the pLIUS (∼1.5 MHz) regimen lies outside the beneficial frequency bandwidth; hence, rabbit experiments were not scheduled with the pLIUS regimen in this study.
In general, left joints that received cLIUS at 3.8 MHz demonstrated a relative absence of chondrocyte clustering and proliferation with a focal absence in the superficial zone compared to right knee joints that served as the contralateral control. These results indicate that cLIUS when applied at the in vivo beneficial frequency bandwidth can have an enhanced effect on MSC differentiation in vivo and the subsequent synthesis of articular cartilage-specific matrix components (i.e., collagen type II and GAGs), while offering chondroprotective effects. Elevated levels of proinflammatory cytokines are noted in diseased and operated joints, drive catabolic effects that impede the chondrogenesis of MSCs, and negatively impact cartilage biosynthesis. 13 Thus, a proinflammatory environment leads to inferior cartilage repair outcomes. Hence, therapeutic strategies that mitigate catabolic responses in a cytokine-rich environment by inhibiting inflammatory pathways, including the NFκB pathway, are needed to promote effective repair processes.
In this study, bilateral defects were created to rule out animal-induced variations in response to treatment; where the right operated joint did not receive the treatment and the left operated joint received cLIUS. While this method may not represent clinical cases, however, the model helped the evaluation of the cLIUS treatment regimen by excluding animal to animal variability. Future studies will consider a larger cohort that will allow the evaluation of isolated knees. Also, future studies will include defects in other areas of the knee (i.e., femoral trochlea) and test the ability of cLIUS to promote repair.
In this study, the chondroinductive ability of cLIUS was maintained in the presence of TNFα, as noted by the increased nuclear deposition of SOX9 and its elevated gene expression in the presence of TNFα. cLIUS further extended its chondroinductive potential by downregulating the TNFα-induced catabolic responses (MMP13, ADAMTS4, and NFκB) and upregulating anabolic responses (SOX9), and similar trends were noted with other cytokines tested. Collectively, our in vitro data suggest that cLIUS elicits a coordinated response that promotes cartilaginous matrix synthesis, while simultaneously protecting the matrix from cytokine-induced degradation, 41 mitigating the catabolic effects of cytokines in the chondrogenic differentiation potential of MSCs.24,45 While the reported findings are representative of experiments performed with two different MSCs donors, the statistical analysis was conducted between replicates (not among donors). These promising results warrant detailed investigations in a clinically relevant large animal model of chondral repair, subject of ongoing investigations.
Footnotes
Acknowledgments
We thank Dr. Craig Kreikemier-Bower and the Center for Comparative Medicine (University of Nebraska-Lincoln) for assistance with animal surgeries and the Pathology Core (University of Alabama, Birmingham) for histological processing. We acknowledge the confocal-imaging facility at the University of Alabama-Huntsville.
Authors' Contributions
A.S. was responsible for research design. H.N., A.D.M., S.B., K.O., J.B.-B., and DW were responsible for research execution and were contributors to data acquisition. K.O. and J.B.-B. applied ultrasound to the rabbits. H.N., A.D.M., and H.V. were the primary contributors to the theoretical analyses that enabled this study. O.S., D.W., and G.S. were responsible for the preparation of histological specimens, scoring, and analyses. A.S., S.B., G.S., and O.S. were responsible for data analysis and interpretation. A.S. prepared the article with revisions provided by H.V., O.S., and S.B.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Health Institute grants (1R21EB025921-01A1 and 1R03AG062730-01).