
Abstract
Ischemic stroke is a devastating medical condition with poor prognosis due to the lack of effective treatment modalities. Transplantation of human neural stem cells or primary neural cells is a promising treatment approach, but this is hindered by limited suitable cell sources and low in vitro expansion capacity. This study aimed (1) use small molecules (SM) to reprogram gingival mesenchymal stem cells (GMSCs) commitment to the neural lineage cells in vitro, and (2) use hyaluronic acid (HA) hydrogel scaffolds seeded with GMSCs-derived neural lineage cells to treat ischemic stroke in vivo. Neural induction was carried out with a SM cocktail-based one-step culture protocol over a period of 24 h. The induced cells were analyzed for expression of neural markers with immunocytochemistry and quantitative real-time polymerase chain reaction (qRT-PCR). The Sprague-Dawley (SD) rats (n = 100) were subjected to the middle cerebral artery occlusion (MCAO) reperfusion ischemic stroke model. Then, after 8 days post-MCAO, the modeled rats were randomly assigned to six study groups (n = 12 per group): (1) GMSCs, (2) GMSCs-derived neural lineage cells, (3) HA and GMSCs-derived neural lineage cells, (4) HA, (5) PBS, and (6) sham transplantation control, and received their respective transplantation. Evaluation of post-stroke recovery were performed by behavioral tests and histological assessments. The morphologically altered nature of neural lineages has been observed of the GMSCs treated with SMs compared to the untreated controls. As shown by the qRT-PCR and immunocytochemistry, SMs further significantly enhanced the expression level of neural markers of GMSCs as compared with the untreated controls (all p < 0.05). Intracerebral injection of self-assembling HA hydrogel carrying GMSCs-derived neural lineage cells promoted the recovery of neural function and reduced ischemic damage in rats with ischemic stroke, as demonstrated by histological examination and behavioral assessments (all p < 0.05). In conclusion, the SM cocktail significantly enhanced the differentiation of GMSCs into neural lineage cells. The HA hydrogel was found to facilitate the proliferation and differentiation of GMSCs-derived neural lineage cells. Furthermore, HA hydrogel seeded with GMSCs-derived neural lineage cells could promote tissue repair and functional recovery in rats with ischemic stroke and may be a promising alternative treatment modality for stroke.
Impact Statement
Neural stem cells, or neural cells, are considered an ideal source of cells for the treatment of neurological related diseases and conditions. We programmed gingival mesenchymal stem cells (GMSCs) toward a neural fate using a small molecule (SM) cocktail and examined the effects of their delivery to stroke sites via a hyaluronic acid (HA) hydrogel. We found that the SMs significantly enhanced the differentiation of GMSCs into neural lineage cells. Furthermore, histological examination and behavioral evaluation demonstrated that HA hydrogel facilitated the proliferation and differentiation of GMSCs-derived neural lineage cells, promoting neurological recovery in rats with ischemic stroke.
Introduction
Ischemic stroke, the most frequent subtype of stroke, is a life-threatening form of neurological diseases with poor prognosis and high morbidity. 1 To date, there is no effective and predictable therapeutic strategy for the treatment of ischemic stroke brain injury. Recently, several preclinical and clinical studies have demonstrated that stem cell transplantation can promote the recovery of cognitive and motor functions in ischemic stroke patients.2,3 However, it is challenging to obtain sufficient numbers of stem cells suitable for clinical use. The use of human neural stem cells and primary neural cells is severely limited by the availability of suitable cell sources and their low proliferative capacity for expansion. Although human embryonic stem cells and induced pluripotent stem cells (iPSCs) possess a high capacity for neural differentiation, their tumorigenic potential, ethical issues, and safety concerns present substantial challenges to clinical application. 4
An increasing number of studies have reported that dental-derived adult stem cells, including dental pulp stem cells (DPSCs), stem cells from exfoliated deciduous teeth, stem cells from the apical papilla (SCAPs), and gingival mesenchymal stem cells (GMSCs), originating from the embryonic neural crest, possess high neurogenic potential and can serve as promising cell sources for neural tissue regeneration.5–7 In particular, GMSCs are an appealing cell source because they are easy to harvest from donor gingival tissue, and wound healing is scarless.8,9 Experimental studies on colitis have proven the unique immunomodulatory properties of GMSCs that enable them to mitigate inflammation-related tissue damage.5,8
Previous studies have demonstrated that a chemical cocktail of SMs can be used to directly reprogram fibroblasts into neural lineage cells. 10 The induced neural cells resemble neurons derived from human iPSCs or neurons generated from human fibroblasts by recombinant ectopic expression of neural lineage-specific transcription factors.11,12 Moreover, this chemical induction approach does not require the use of viral vectors, genomic modification, or genetic manipulation. Our recent study also demonstrated that GMSCs can efficiently differentiate into neural lineage cells when cultured in neural induction media containing SMs. 13 Since stem cells can differentiate into multiple lineages under different microenvironmental conditions, 14 it is critical to ensure neural lineage-specific differentiation of the transplanted cells to facilitate brain tissue regeneration after an ischemic stroke. The primary objective of the present study, therefore, involves the reprogramming of GMSCs into neural lineages with SMs prior to transplantation in a rat model of ischemic stroke.
The microenvironmental conditions of an ischemic stroke site are hypoxic and inflammatory, which is detrimental to the survival, proliferation, and differentiation of transplanted cells. 13 Therefore, protecting the transplanted cells from apoptosis and preventing nonneural lineage differentiation is crucial for promoting neural regeneration. Several studies have reported that co-transplanting stem cells with bioactive materials that mimic natural stem cell niche can prevent apoptosis of transplanted cells.15,16 Both natural and synthetic biomaterials have been used in engineered to engineer three-dimensional stem cell microenvironments that can mimic brain extracellular matrix (ECM) and support endogenous repair mechanisms.17,18 Hyaluronic acid (HA) is widely used in engineered hydrogels because of its biocompatibility, injectability, and thermo-responsive properties. Furthermore, cells encapsulated in HA hydrogels can be protected from damage during delivery from shear stress. 19 Nonetheless, the use of injectable HA hydrogels seeded with GMSCs has not been investigated as a treatment for ischemic stroke.
In the present study, we reprogrammed GMSCs into neural lineage cells in vitro before in vivo transplantation using a chemical cocktail of SMs. We then investigated the therapeutic efficacy of in vivo delivery of self-assembled HA hydrogels loaded with GMSCs-derived neural lineage cells via stereotactic injection in an animal model of ischemic stroke.
Materials and Methods
Chemical cocktail-based reprogramming of GMSCs into neural lineage cells
GMSCs were isolated from human gingival tissue as previously described. 20 Approval was obtained from the Institutional Review Board of the University of Hong Kong/Hospital Authority Hong Kong West Cluster (HKU/HA HKW IRB. IRB Reference Number: UW 17–522). Informed written consent was obtained from the patients. GMSCs were characterized by flow cytometric analysis based on their expression of CD45, CD73, CD90, and CD105, as well as multilineage differentiation assays using osteogenic, adipogenic, and neurogenic induction protocols. 21 Passages 4–10 were used for these experiments. GMSCs were seeded onto gelatin-coated 24-well culture plates (1-2 × 105 cells/well) and cultured in α-MEM (Gibco-Invitrogen, Carlsbad, CA). Upon reaching 60% confluence, the culture medium was changed to a neural induction medium (DMEM/F12: Neurobasal with 0.5% [v/v] N2, 1% [v/v] B27, 100 mM cAMP, 20 ng/mL bFGF) (ThermoFisher Scientific, Inc., Waltham, MA) supplemented with the chemical cocktail VCRFSGY (0.5 mM VPA, 3 µM CHIR99021, 1 µM Repsox, 10 µM Forskolin, 10 µM ISX9, 2 µM IBET151, 5 µM Y-27632, and 1 µM Purmorphamine) (Sigma-Aldrich, Burlington, MA) for 24 h. The induced cells were subsequently subjected to further analyses in vitro or transplanted for in vivo experiments.
Assessment of GMSCs neural differentiation
The expression of early neural markers (Nestin and Vimentin), immature neural markers (Doublecortin, DCX and Tubulin β-III, TUJ1), and mature neural markers (Microtubule associated protein 2, MAP2, and Neuronal nuclei, NeuN) in GMSC-derived neural lineage cells was analyzed by quantitative real-time polymerase chain reaction (qRT-PCR) and immunocytochemistry at 0, 2, 6, 12, and 24 h of neural induction. Briefly, total RNA was extracted using the RNeasy Mini kit (Qiagen, Hilden, Germany), and reverse-transcribed to cDNA using SuperScript VILO Master Mix (Gibco-Invitrogen), according to the manufacturers’ instructions. Then, qRT-PCR was performed with TB Green Premix Ex Taq (Takara, Shiga, Japan) using a StepOnePlus Real-Time PCR System (Applied Biosystems, Grand Island, NY). Relative gene expression was quantified using the comparative CT method (ΔΔCT method). The qRT-PCR primer sequences used for amplification of the neural marker target genes are listed in Table 1. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an endogenous control. For immunocytochemistry, cells grown on glass coverslips were fixed with 4% paraformaldehyde for 20 min and then incubated with blocking buffer (5% normal serum and 0.3% Triton™ X-100 diluted in 1×PBS) for 60 min. Subsequently, cells were incubated with anti-Nestin (1:3,200, #33475, Cell Signaling Technology, Danvers), anti-Vimentin (1:100, #5741, Cell Signaling Technology), anti-DCX (1:1,000, ab18723, Abcam, Cambridge, UK), anti-TUJ1 antibody (1:1,000, ab78078, Abcam), anti-NeuN (1:1,000, ab104224, Abcam), or anti-MAP2 (1:1,000, AB5622, Millipore, Darmstadt, Germany) antibody at 4°C overnight. The samples were then incubated with the corresponding secondary antibodies (1:200), that is, goat anti-mouse secondary antibody conjugated to Alexa fluor 488 (ab150117, Abcam) or goat antirabbit secondary antibody conjugated to tetramethyl rhodamine (TRITC; ab6718, Abcam), for 2 h in the dark at room temperature. DAPI (1:10,000, ab104139, Abcam) was used to stain the cell nuclei. The samples were imaged under a laser scanning confocal microscope (FV1000, Olympus, Tokyo, Japan) using the specific excitation/emission wavelengths for TRITC (540/570 nm) and Alexa Fluor 488 (490/520 nm) and analyzed using the FV10-ASW microscopy software. For quantitative analyses of the neural marker expression, 3 random areas per group were assessed for each marker.
Primer Sequences Utilized in Quantitative Real-Time Polymerase Chain Reaction
DCX; doublecortin; MAP2; microtubule associated protein 2; NeuN; neuronal nuclei; TUJ1; beta III tubulin.
Preparation of HA hydrogels
HA was functionalized with acrylate groups using a two-step synthesis protocol. 22 A reaction consisting of 0.017 mmol HA (Mw = 70–80 kDa) (Huaxi Furuida Biomedical Co., Ltd., Beijing, China) combined with 105.5 mmol adipic dihydrazide (ADH) (Sigma-Aldrich) at pH 4.75 in the presence of 10.41 mmol 1-ethyl-3-[3-dimethylaminopropyl] carbodiimide hydrochloride (EDC, Sigma-Aldrich) was allowed to proceed overnight and was purified by dialysis (8000 MWCO) in deionized (DI) water for 1 week. The purified intermediate (HA-ADH) was lyophilized. Then, 0.014 mmol HA-ADH was incubated with 4.4 mmol N-acryloxysuccinimide in HEPES buffer (pH 7.2) (Sigma-Aldrich) overnight and purified by dialysis in DI water for 1 week before lyophilization. HA hydrogels were crosslinked by Michael’s Addition using dithiothreitol (DTT). Lyophilized aliquots of the crosslinker were diluted in 10 μL of 0.3 M HEPES (pH = 8.0, Sigma-Aldrich) buffer immediately prior to mixing with 90 μL HA solution in 0.3 M HEPES, with or without cells.
Assessment of viability
The survival rate of GMSCs cultured with HA hydrogels was assessed using a CCK8 kit (Dojindo, Kumamoto, Japan) and the Transwell system (Costar 3422, Corning, NY). Briefly, GMSCs (2 × 105 cells/well) were cultured in 24-well plates with 30 µL HA hydrogels placed in the upper compartment. On days 1, 3, 5, and 7, the culture medium was replaced by a mixture containing CCK8 solution and incubated for 2 h. Subsequently, the reaction solution was transferred to a 96-well plate, and absorbance was measured on a microplate reader (M2, Molecular Devices, San Jose, CA) at 450 nm. The background absorbance of wells containing medium and CCK8 solution, but no cells, was also measured. Cell proliferation was represented as the mean ± SD of absorbance for 6 wells from each group. All experiments were carried out in triplicate. GMSCs (5 × 106 cells/mL) were homogeneously encapsulated into HA hydrogels and crosslinked using DTT, then cultured for 3, 5, and 7 days. Cell viability was determined using the Live/Dead assay kit (Gibco-Invitrogen). Briefly, samples of HA hydrogels embedded with GMSCs were rinsed gently with sterilized PBS and stained with a reagent mixture containing 2 µM acetoxymethyl (AM) and 4 µM ethidium homodimer-1 (EthD-1) (Gibco-Invitrogen) for approximately 30 min. Then, the samples were rinsed twice with PBS and examined using confocal laser scanning microscopy (CLSM) (Carl Zeiss LSM 710, Zeiss, Oberkochen, Germany). The images were analyzed with Image J software (version 1.49, National Institutes of Health, Bethesda, MD).
Establishment of the ischemic stroke model and cell transplantation
Ethical approval for all protocols related to animal experiments was obtained from Xuzhou Medical University (No.201909W006). A total of 100 pathogen-free, adult Sprague-Dawley (SD) male rats, each weighing 220 to 230 g (Charles River Laboratories International, Inc., Beijing, China), were maintained on a 12-h light/dark cycle with food and water available ad libitum. The rats were subjected to the middle cerebral artery occlusion (MCAO) reperfusion ischemic stroke model as previously described. 23 Briefly, a propylene filament (diameter 0.12 mm, length 30 mm, tip diameter with coating 0.35 mm) (Doccol Corporation, MA) was advanced to the ostium of the right MCAO in the circle of Willis of the rat under isoflurane (4% induction, 1% maintenance in 30% O2) anesthesia. The right MCAO was occluded for 60 min prior to reperfusion by retracting the filament to the common carotid bifurcation. The rats allocated to the sham-operated group underwent an identical surgical procedure, but the MCAO was not occluded. Rats were kept and monitored in the intensive care unit (ICU) until they were fully recovered from anesthesia. Postoperatively, rats were given Enrofloxacin (10 mg/kg) (ChemicalBook, Beijing, China) for infection prevention for 1 week and Meloxicam (2 mg/kg) (ChemicalBook, Beijing, China) for pain relief for 3 days. All the surgical procedures were conducted under anesthesia. The neurological deficits of the ischemic stroke model rats were evaluated using Zea Longa 5-point scale 2 days post-MCAO surgery. The scoring criteria were as follows: 0, no deficit; 1, irregular gait, failure to extend the forepaw contralateral to the ischemic brain fully; 2, circling to the contralateral side of the ischemic brain; 3, failure to stand steadily, falling down; and 4, no spontaneous walking, with a loss of consciousness. Rats that scored 1–3 points were included in the study. 24 Cerebral infarction in the rats was further confirmed by 2% 2, 3, 5-triphenyltetrazolium chloride (TTC) (Sigma-Aldrich) staining as previously described. 25
Eight days after MCAO, the rats were anesthetized with isoflurane (4% induction, 2% maintenance) and randomly assigned to one of six study groups (n = 12): (1) transplantation with GMSCs (GMSCs); (2) transplantation with GMSCs-derived neural lineage cells (SM-GMSCs); (3) transplantation with HA and GMSCs-derived neural lineage cells (HA-SM-GMSCs); (4) transplantation with HA; (5) transplantation with PBS; (6) sham transplantation control (Sham). Based on the in vitro data, GMSCs treated with SMs for 12 h were used in the animal experiments. Rats were placed in a stereotactic frame and injected with approximately 500,000 cells in 10 μL PBS solution or hydrogels at coordinates (L) 3.0 mm, (AP) 0.0 mm, and (V) 6.0 mm, using a 10 μL Hamilton syringe with a 22G blunt tip needle at a rate of 1 μL/min. Specifically, 2 μL boluses were injected at five depths (6, 5, 4, 3, and 2 mm relative to the skull). Following the last injection, the needle was kept in place for 3 min before removal. A bone-like polyethylene burr-hole cover was used to close the craniotomy site. The incision was then sutured, and the rats were kept warm during recovery with a heating pad and rectal probe feedback system. Rats were administered cyclosporin A intraperitoneally (10 mg/kg daily) (Woburn, MA) two days before the cell transplantation and 14 days after the surgery to prevent the rejection of the transplanted cells. The control animals were injected with PBS. The rats in the sham transplantation control group received sham surgery with no injection. Rats were intraperitoneally injected with 5-bromo-2’-deoxyuridine (BrdU) (150 mg/kg perinjection; 2 mL/kg, once per day at day 1, 3, 5 and 7 after treatment). The experimental approach for this study is shown in Figure 1.
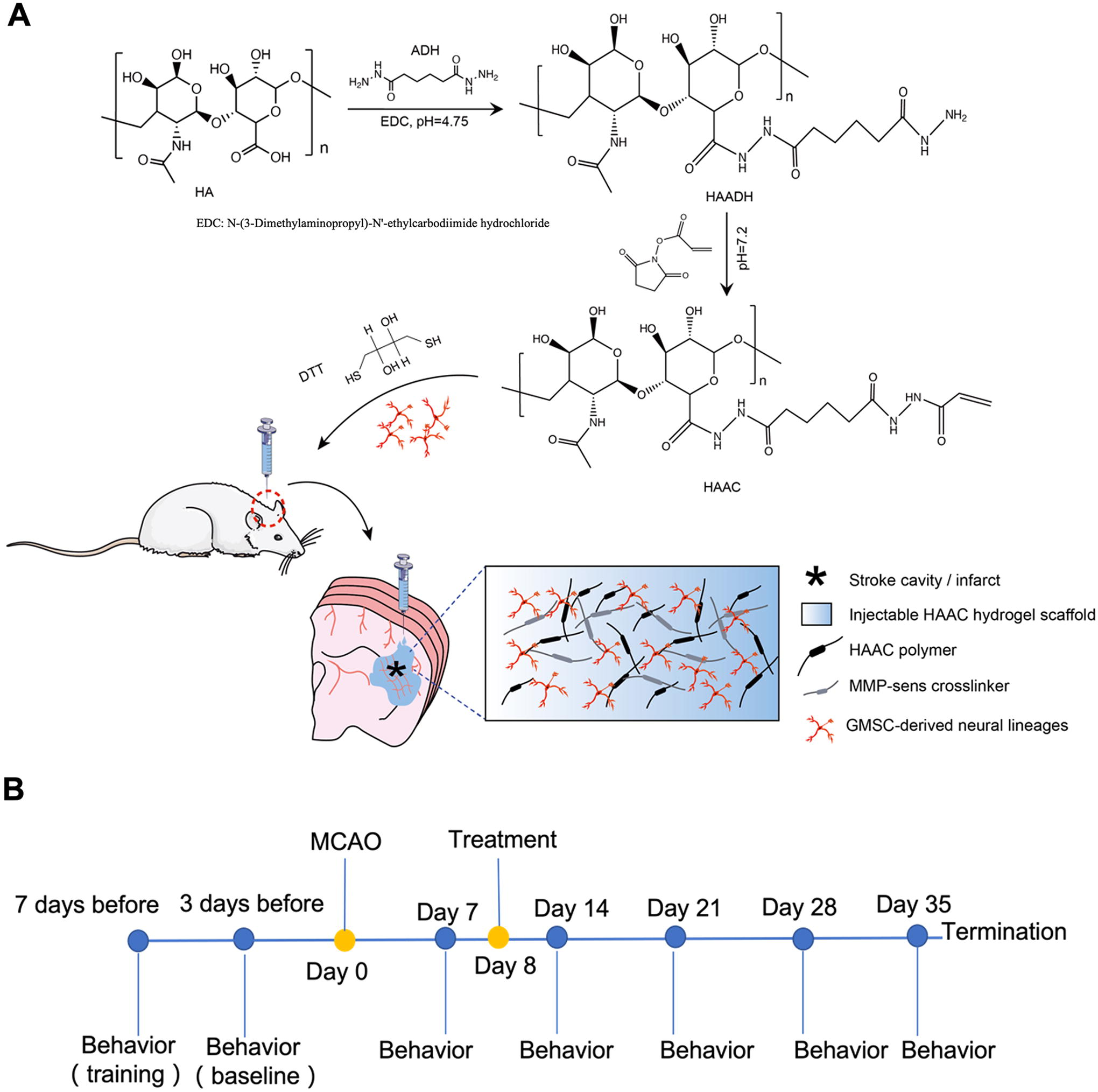
Graphic illustration of the study.
Behavioral assessments of post-stroke recovery
The behavioral tests conducted to evaluate post-stroke recovery were the grid walking foot fault test, rotarod test, balance beam test, cylinder test, and limb placing test, each performed at five separate time points (baseline before stroke surgery, 1-, 2-, 3-, 4-, and 5-week post-stroke surgery). The rats were trained on three separate days prior to recording their baseline behavior. The behavioral tests were conducted by two investigators who were blinded to the animal group allocations. The experimental timeline of procedures is shown in Figure 2.
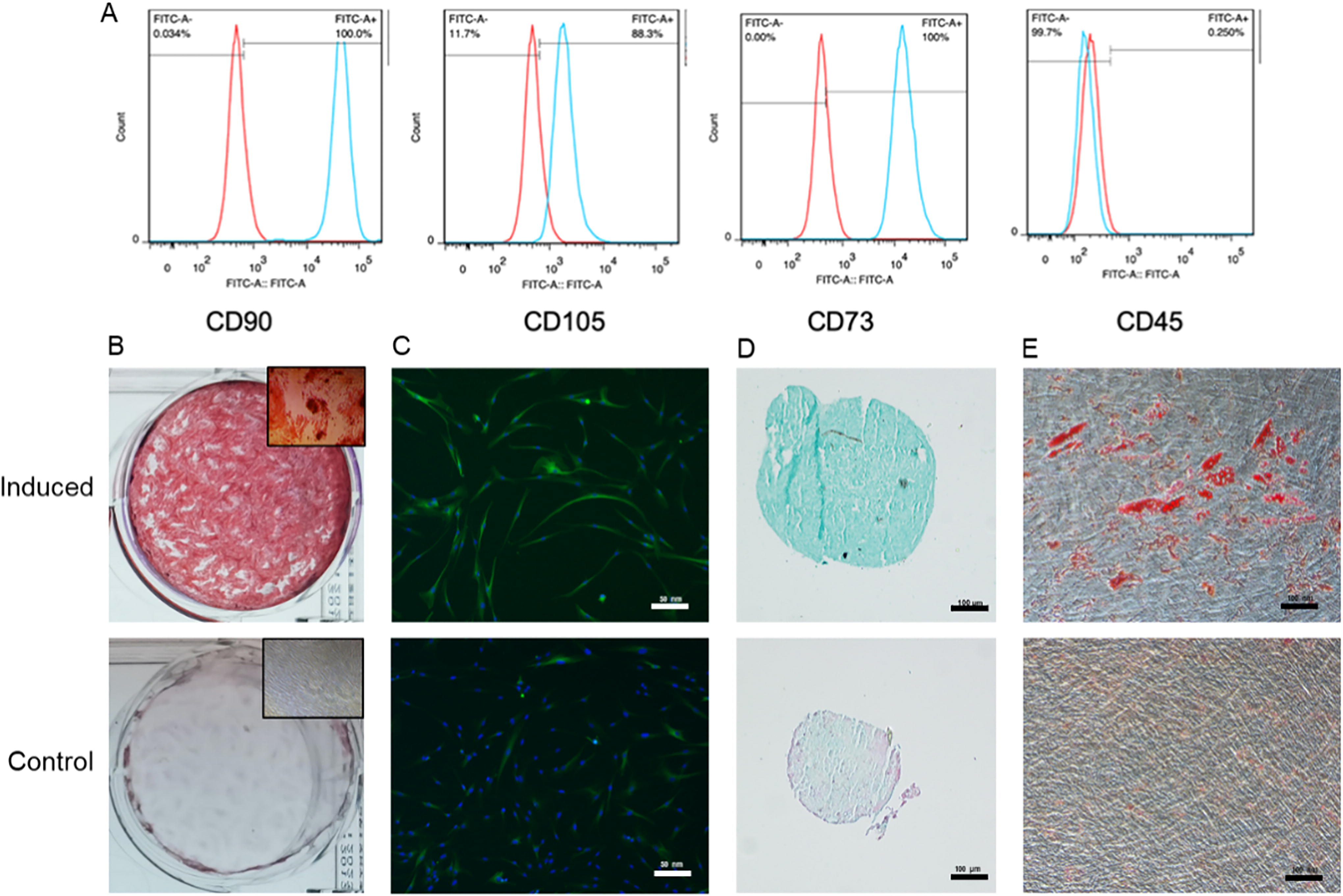
Representative histograms of cell surface markers
Grid walking foot fault test
The rat was placed on an elevated stainless-steel square grid (40 × 40 cm2 with each grid cell being 2.5 × 2.5 cm2, at a height of 50 cm) that was located in a sound-attenuated room with dim lighting for 1 minute. 26 The rat’s behavior on the grid was recorded on DVD (Icomery, Shenzhen, China). Each rat was tested three times. The data were expressed as the number of foot fault errors made by the contralateral limbs as a percentage of total steps.
Rotarod test
The rotarod test was conducted using a treadmill with a diameter of 3 cm as previously described. 24 Briefly, the rat was placed on an accelerating rotarod (velocity gradually increases from 4 to 40 rpm) for a maximum duration of 5 min, and the time until the rat fell was recorded. The test was performed on each rat three times daily, with 2-minute rest intervals between trials.
Balance beam test
The rat was placed on a beam (120 cm in length, 2 cm in width) elevated 60 cm above the ground for walking. 19 The scoring criteria were as follows: 0 = balances with steady posture; 1 = grasps the side of the beam; 2 = hugs beam with one limb, falls down from beam; 3 = hugs beam with two limbs, falls down from beam, or spins on beam > 60 s; 4 = attempts to balance on beam but falls off > 40 s; 5 = attempts to balance on beam but falls off > 20 s; and 6 = falls off, no attempt to balance or hang onto beam < 20 s. Maximal testing time was 60 s. Each animal was tested twice per time point and means were calculated.
Cylinder test
The rat was placed in a transparent cylinder (9 cm in diameter and 15 cm in height) for 5 min and its exploratory activity was recorded by camera. 27 Forepaw (left/right/both) use on initial contact against the cylinder wall after rearing and during lateral exploration was calculated. Use of the impaired forepaw was assessed and the final score (percentage) was determined using the following formula: (impaired forelimb movement + movement of both/2)/(non-impaired forelimb movement + impaired forelimb movement + movement of both).
Histology
Animals were terminally anesthetized 4 weeks after the MACO surgery by overdosing with sodium pentobarbital (50 mg/kg i.p.) and were then subjected to transcardial perfusion with saline containing 0.002% (w/v) NaNO2 and 0.002% (w/v) heparin, followed by 4% (w/v) paraformaldehyde (PFA) in 0.1M PBS (pH 7.4). Brains of sacrificed animals were removed after craniotomy and were fixed in paraformaldehyde for 24 h. The brains were then immersed in cryoprotective solution (30% sucrose in PBS with 0.01% sodium azide) for 72 h. After freezing the brain on dry ice, a sliding microtome (ThermoFisher Scientific Inc., Waltham, MA) was used to cut 30-µm coronal sections through the striatum starting at the anterior corpus callosum. Sections were stored in 30% (w/v) sucrose and 30% (w/v) ethylene glycol solution in PBS at 4°C. The tissue slides were blind labeled so as to conceal group allocation. The tissues were stained with hematoxylin and eosin (HE) and lesion topography was visualized by light microscopy. Immunohistochemistry staining of BrdU were performed using the peroxidase method. The fate of the transplanted cells within the rat brain was further examined using fluorescent double-label immunohistochemistry. After washing in PBS, sections were permeabilized in 0.4% (w/v) Triton X-100 in PBS. After blocking with 10% (v/v) goat serum and 1 mg/mL BSA, the sections were incubated for 2 h with primary antibodies specific for human nuclear antigen (HuNu) (1:200, ab191181, Abcam), followed by a primary antibody specific for either MAP2 (1:1,000, BS-1369R, Bioss) and TUJ1 (1:2,000, ab18207, Abcam). For colocalization, after washing away unbound primary antibodies, samples were incubated with appropriate fluorescent secondary antibodies for 1 h at 37°C and counter-stained with DAPI (to visualize nuclear DNA), prior to imaging under a Nikon Eclipse TiTM fluorescent microscope (Nikon Inc., Tokyo, Japan). Six random areas per group were assessed for each protein for the quantitative analysis.
Statistical analysis
Statistical analysis was performed using the IBM SPSS Statistics software, version 20.0 (SPSS Inc., Chicago, IL). All data are expressed as mean ± standard error of mean unless otherwise indicated. The data were analyzed by either One-way or Two-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. The threshold of statistical significance was set at p < 0.05.
Results
Small molecule cocktail accelerates the neural differentiation of GMSCs in vitro
GMSCs were identified as cells positive for CD90, CD73, and CD105, and negative for CD45, by flow cytometry (Fig. 2A). GMSCs demonstrated the capacity for multipotent differentiation into osteogenic, neurogenic, chondrogenic, and adipogenic lineages (Fig. 2B–E).
The SM cocktail significantly enhanced the differentiation of GMSCs into neural lineage cells, as shown by morphological changes (Fig. 3A), immunocytochemistry (Fig. 3B–D), and qRT-PCR analyses (Fig. 4A–C). The cellular morphology of GMSCs treated with SMs changed significantly from 2 h to 24 h, and there was a time-dependent increase in the percentage of cells exhibiting neurite outgrowth, the total neurite length per cell, and the number of neurites per cell (Fig. 3A). Nestin, Vimentin, DCX, TUJ1, NeuN, and MAP2 were upregulated after treatment with SMs over 24 h (Fig. 3B–D). Nestin and Vimentin, the early neural markers, increased from 0–6 h, and then decreased at 12 and 24 h. The expression of DCX and TUJ1 (markers of immature neurons), gradually increased from 0–12 h, and decreased at 24 h, whereas NeuN and MAP2 (markers of mature neural tissue) were gradually upregulated from 0–24 h. Consistent with these data, qRT-PCR analyses revealed that the expression of Nestin, TUJ1, and MAP2 were upregulated to peak levels at 6, 12, and 24 h, respectively (Fig. 4). In the other words, the result of the immunocytochemistry and qRT-PCR analyses indicated that GMSCs may be induced by SMs into an NPCs or NPC‐like cell state by 12 h, followed by further differentiation into neuron‐like cells. Thus, the cells treated with SMs for 12 h were used for the cell transplantation in vivo.
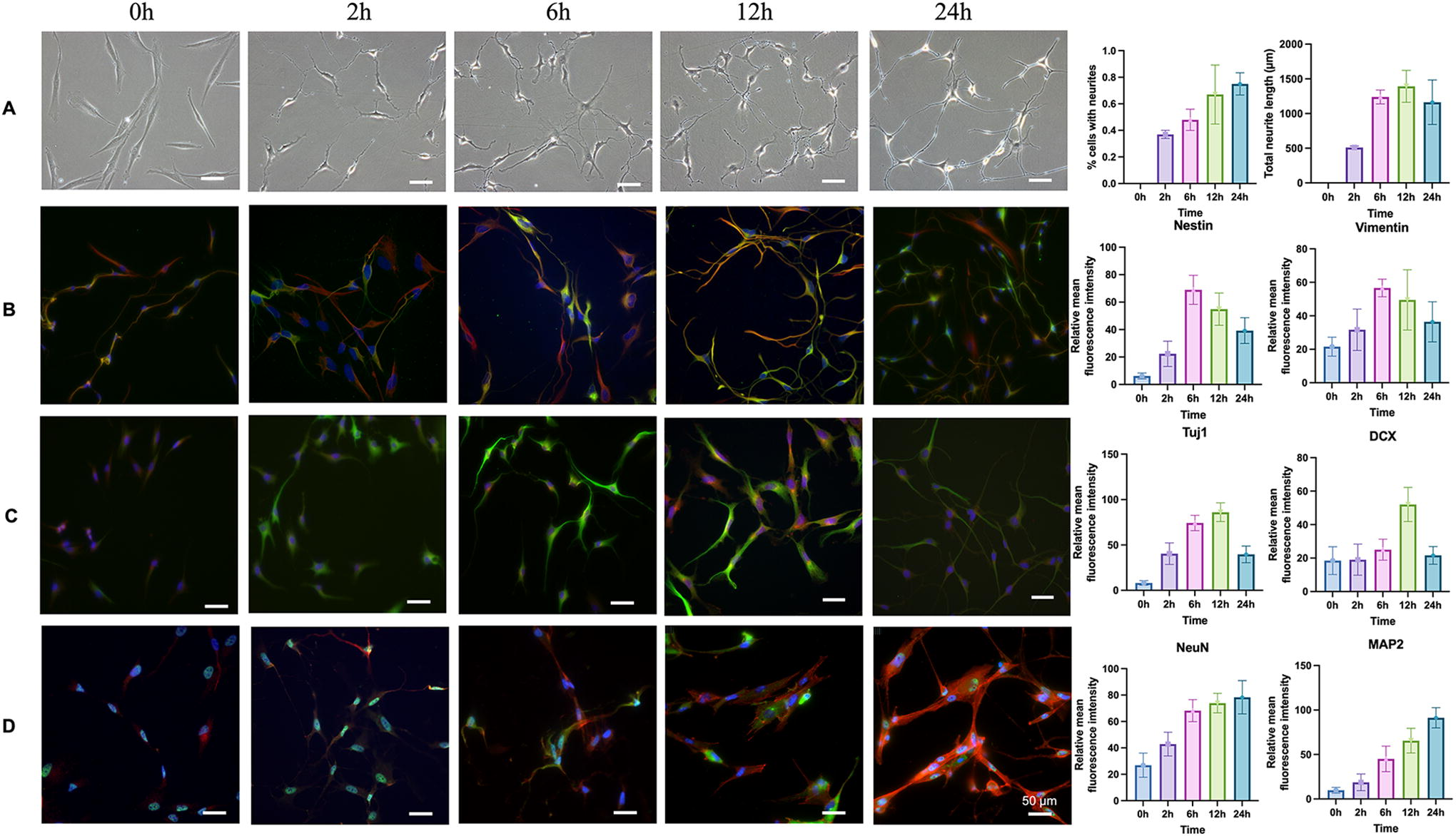
Representative images of brightfield and immunocytochemistry, and the corresponding quantitative analysis for neural markers in GMSCs treated with small molecules at different time points.
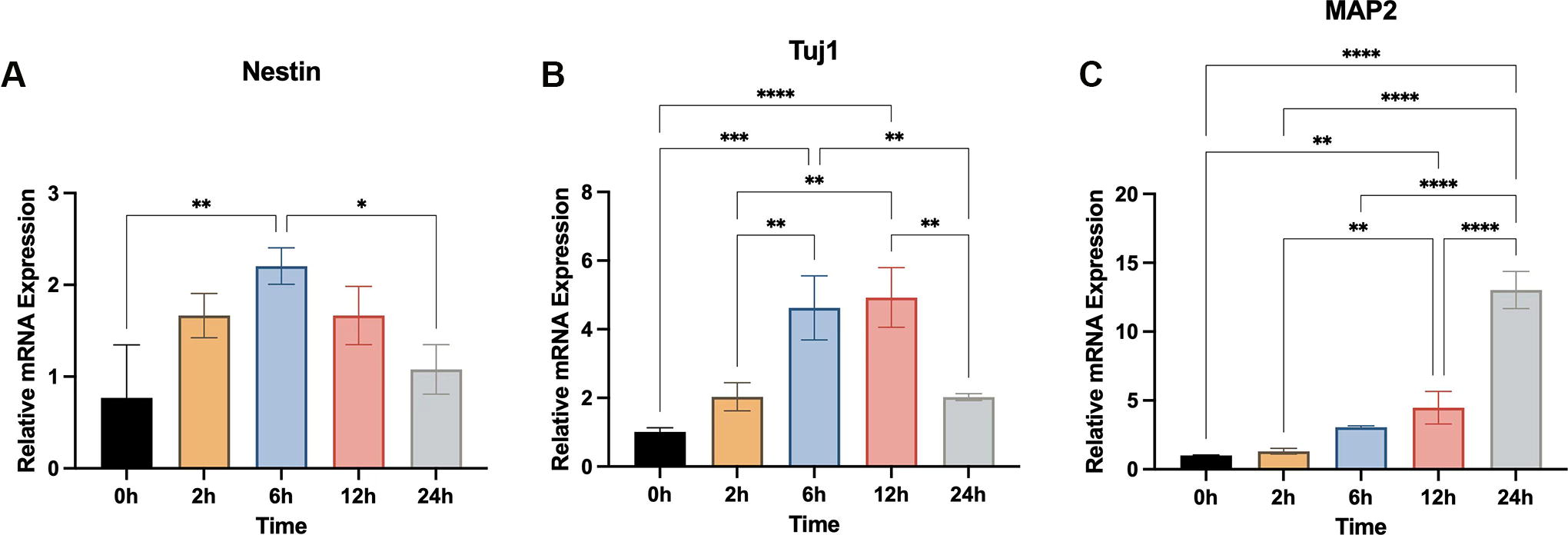
Quantitative reverse transcription-polymerase chain reaction analyses of neural marker expression by GMSCs after treatment with small molecules for different timepoints.
Viability assessment
There were no significant differences in cell viability between the control and HA hydrogels groups (p > 0.05) on days 1, 3, and 7 (Fig. 5A). CLSM images confirmed that more than 90% of the cells in both groups were still alive on days 3, 5, and 7 (Fig. 5B).
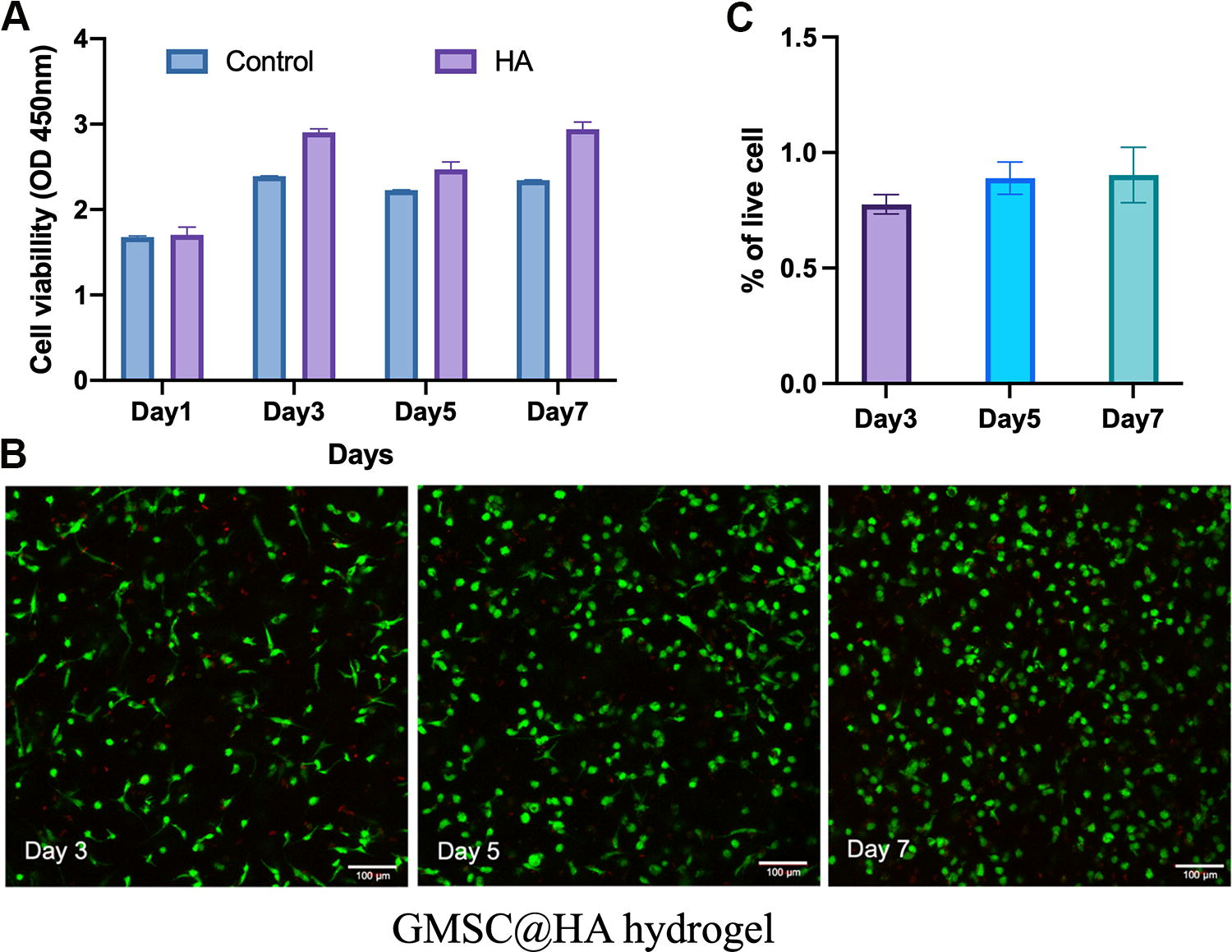
Cell viability of GMSCs cultured with HA hydrogels using the Transwell system
Transplantation of HA carrying GMSC-derived neural lineages promotes neurological recovery
A rat ischemic stroke model was established using MCAO and reperfusion method (Fig. 6A1–A3). Rats with scores of 1–3 were considered successful models based on the Zea Longa five-point scoring system. Further confirmation of cerebral ischemia was conducted by 2% TTC staining (Fig. 7B1 and B2).
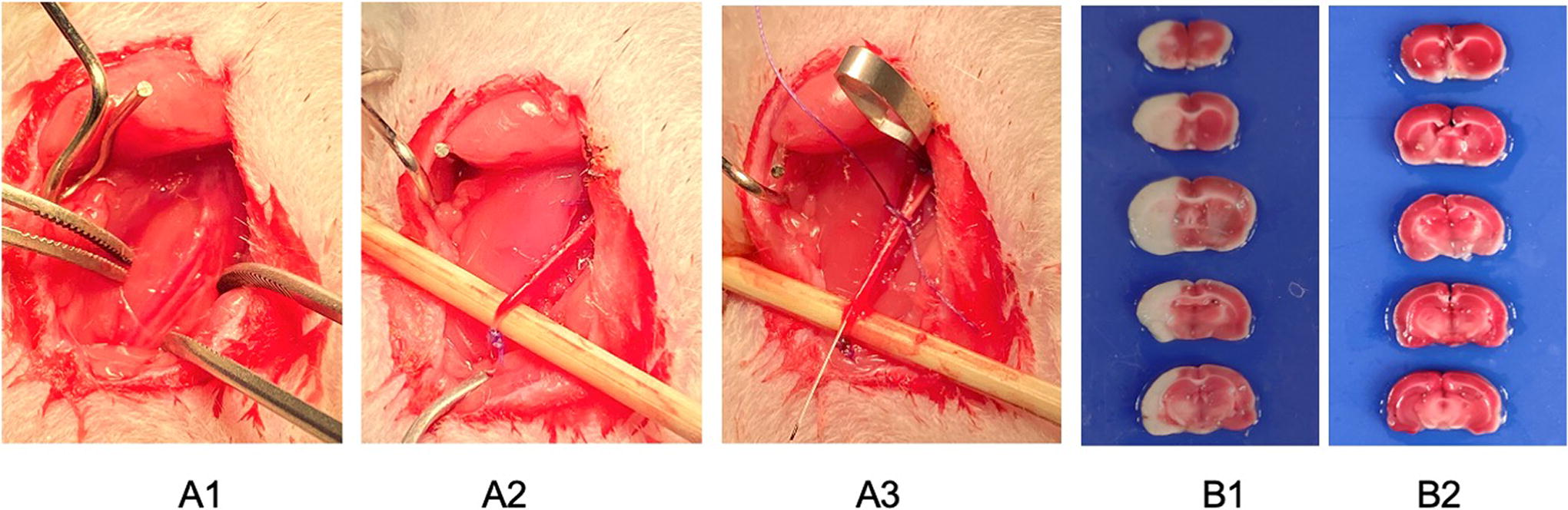
Surgery procedures of middle cerebral artery occlusion
Results from the grid walking foot fault test showed that there was a significantly lower percentage of foot fault errors in the HA-SM-GMSCs and sham control groups, compared to the PBS control, GMSCs, and SM-GMSCs groups at 3–5 weeks post- stroke surgery (all p < 0.05) (Fig. 7A). A similar trend was observed in results from the rotarod (Fig. 7B), beam balance (Fig. 7C), and cylinder (Fig. 7D) tests.
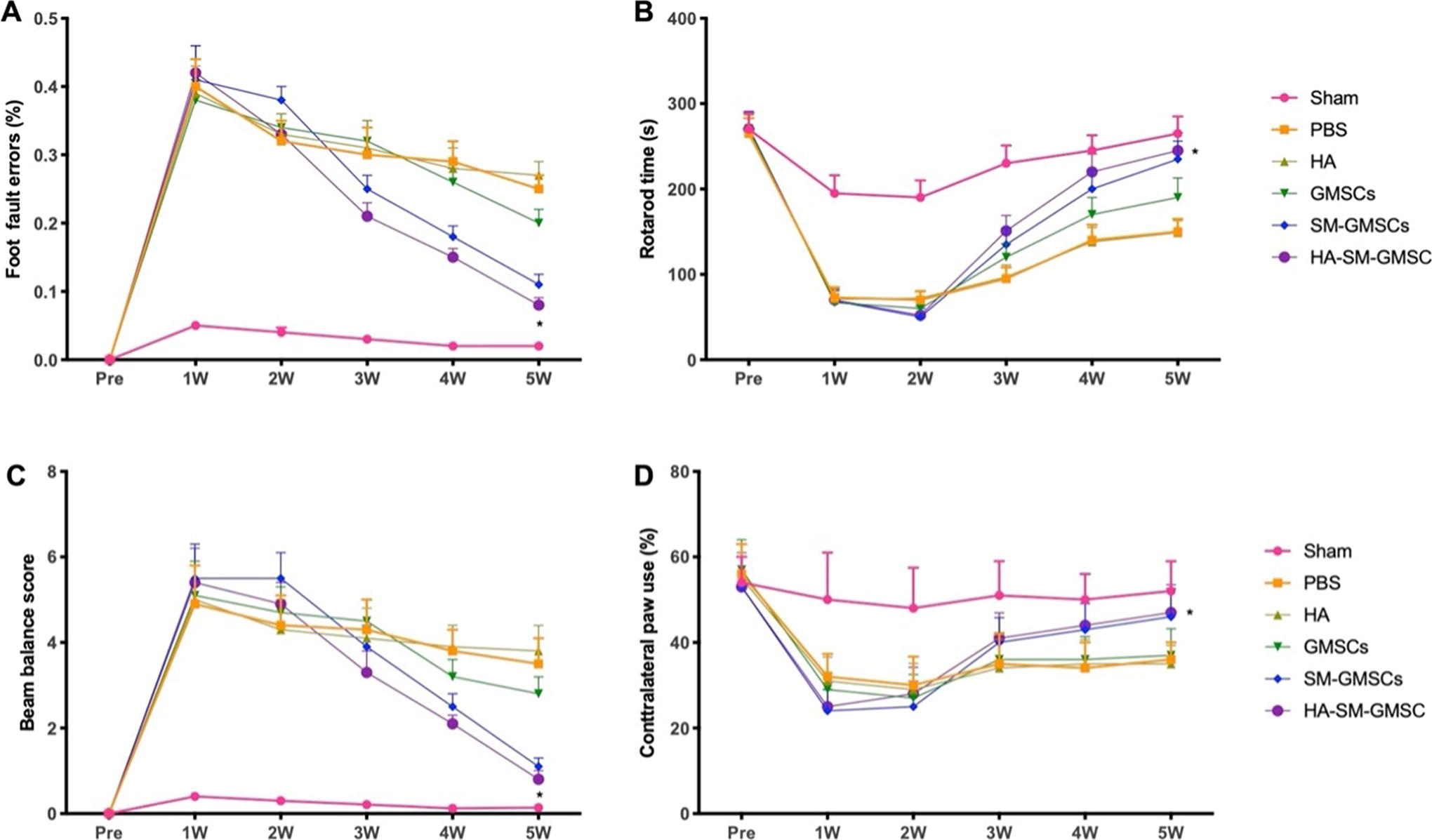
Behavioral changes of rats with ischemic stroke before and after surgery.
We also observed significantly fewer vacuoles and pyknotic cells in the HA-SM-GMSCs and sham groups compared with the PBS, HA, GMSCs, and SM-GMSCs groups at 4 weeks posttreatment (p < 0.05) (Fig. 8).
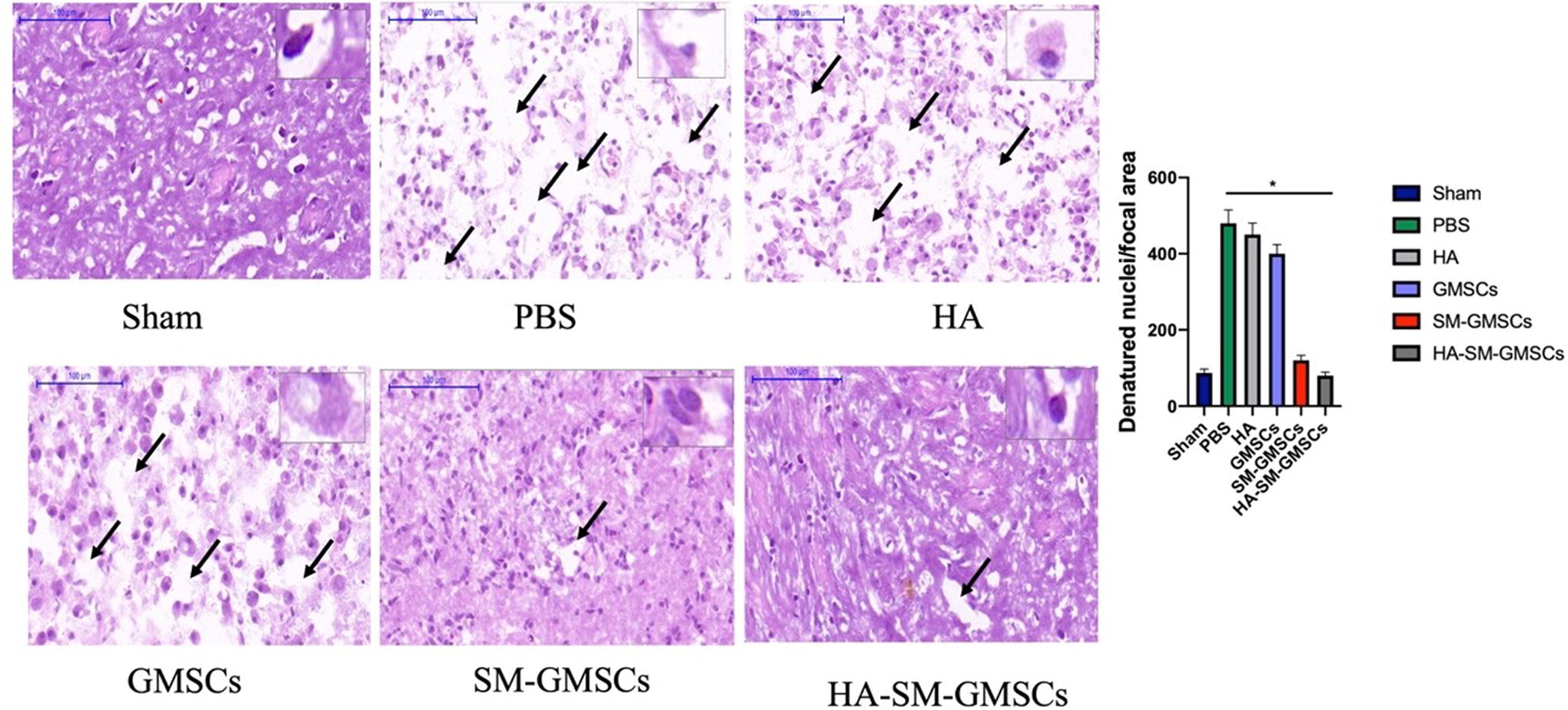
Representative HE images for sham, PBS, HA, GMSCs, SM-GMSCs, and HA-SM-GMSCs groups. (p < 0.05, HA-SM-GMSCs or sham group vs. PBS, or HA, or GMSCs, or SM-GMSCs group). GMSCs, gingival mesenchymal stem cells; HA, hyaluronic acid; HA-SM-GMSCs, hyaluronic acid-small molecule-gingival mesenchymal stem cells; SM-GMSCs, small molecule-gingival mesenchymal stem cells.
Immunohistochemical staining of brain tissues for BrdU revealed that there were significantly more BrdU-positive cells in the HA-SM-GMSCs group than in the sham, PBS, HA, GMSCs, and SM-GMSCs groups at 4 weeks posttreatment (p < 0.05) (Fig. 9).
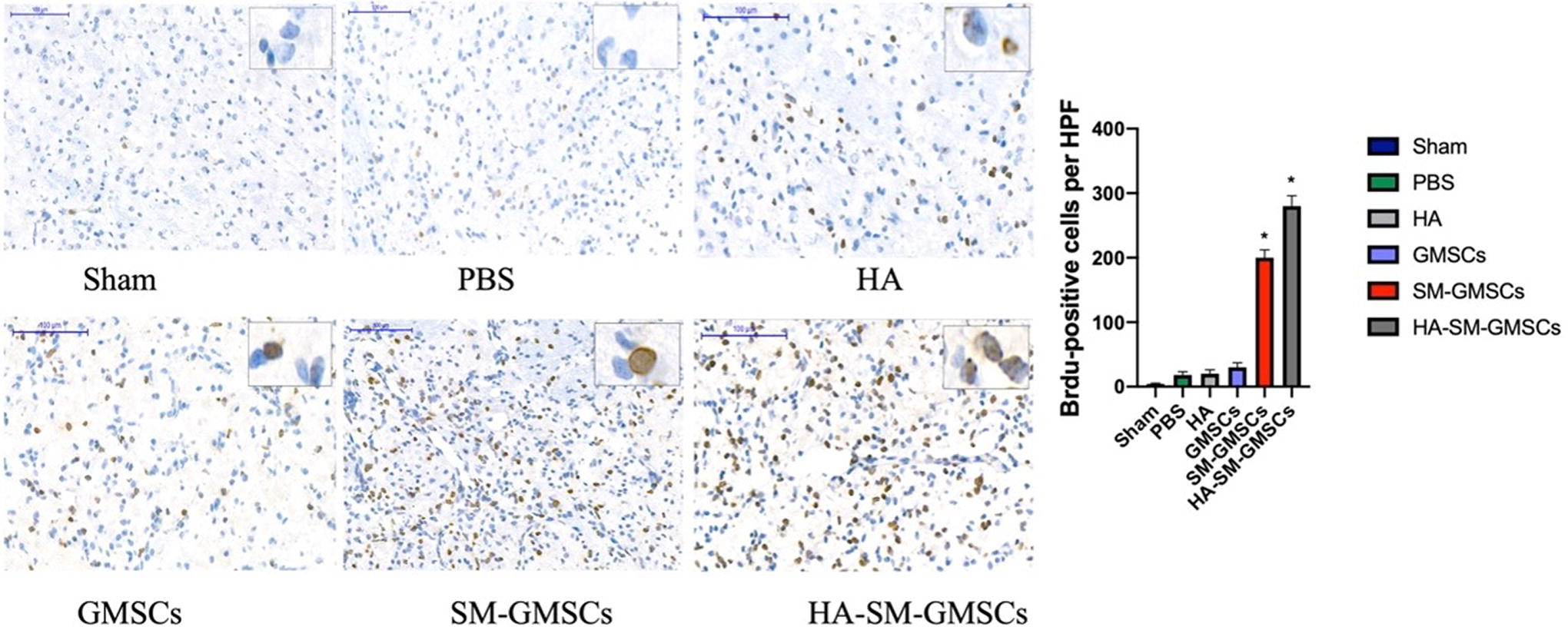
Representative immunohistochemistry staining for sham, PBS, HA, GMSCs, SM-GMSCs, and HA-SM-GMSCs groups. (p < 0.05, HA-SM-GMSCs group vs. sham, PBS, HA, GMSCs, or SM-GMSCs group). GMSCs, gingival mesenchymal stem cells; HA, hyaluronic acid; HA-SM-GMSCs, hyaluronic acid-small molecule-gingival mesenchymal stem cells; SM-GMSCs, small molecule-gingival mesenchymal stem cells.
HuNu-positive cells were detected in the HA-SM-GMSCs, SM-GMSCs, and GMSCs groups at 4 weeks posttreatment, whereas no HuNu-positive cells were observed in the sham, PBS, and HA groups (Fig. 10AFig. 10B). HuNu-positive cells also coexpressed TUJ1 or MAP2. The number of of HuNu+ TUJ1+ and HuNu+ MAP2+ in the HA-SM-GMSCs group was significantly higher compared with the SM-GMSCs and GMSCs groups at 4 weeks posttreatment (p < 0.05) (Fig. 10C).
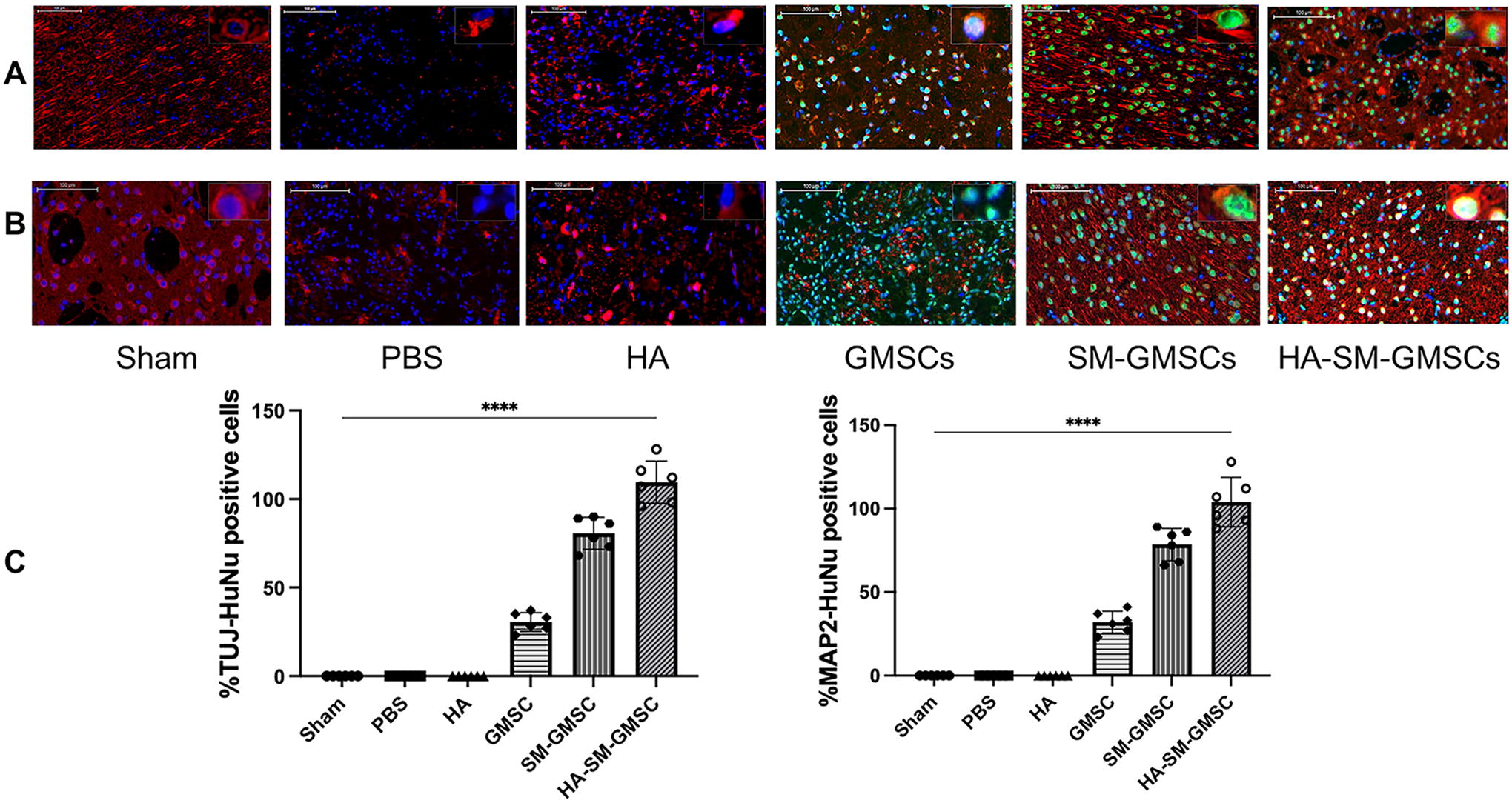
Representative immunofluorescence images for neural markers in sham, PBS, GMSCs, and SM-GMSCs groups.
Discussion
Previous studies have shown the promise of transplantation of stem or neural lineage cells as a treatment for ischemic stroke, spinal cord injury and brain trauma.28,29 Dental stem cells are typically postnatal mesenchymal stem cells (MSCs) originating from the neural crest of the embryo. They constitutively express high levels of neural markers and have been widely investigated as treatments for various neurological diseases in animal models.23,30 Some studies have reported positive results for the transplantation of undifferentiated dental stem cells in facilitating neural healing and regeneration, but there are also concerns about their ability to spontaneously differentiate into other cell lineages.31,32 The pathological and inflammatory local microenvironment of ischemic brain lesions may detrimentally affect cell survival, prevent the transplanted MSCs from differentiating into neural lineages, or even induce them towards other lineage fates, which could lead to the formation of nonneural tissue such as fibrotic scar tissue that would compromise the recovery of neural functions. 33 In this context, precommitment of the transplanted cells to the neural lineage could be a critical step to ensure that they differentiate into the desired lineage. 34
In our previous study, it demonstrated that GelMA hydrogels loaded with brain-derived neurotrophic factor (BDNF) induced SCAPs to differentiate into neuron-like cells in our previous study. 35 Our group further reported that a SM cocktail-based two-step culture protocol over a duration of 14 days could significantly enhance the commitment and differentiation of DPSCs, SCAPs, and GMSCs toward the neural lineage fate. 14 These studies highlight the importance of neural growth factors, or SMs, in regulating stem cell differentiation toward neural lineages. Unfortunately, in our previous study, we observed substantial cell death following long-term induction with SMs. 36 The neural induction period varies in different studies.36,37 It is possible to alter the induction period by modifying the SMs compounds that play an essential role in neural induction. In the present study, we deliberately shorten the induction period. We found that a cocktail of eight SMs rapidly and efficiently reprogrammed GMSCs into neural lineage cells after 12 h of treatment in a single step. The induced cells comprised a mixture of neural progenitor cells (NPCs) and neural cells, as demonstrated by the expression of markers for early (Nestin and Vimentin), immature (DCX and TUJ1), and mature (MAP2 and NeuN) neurons. The elevated expression levels of immature neural markers suggest that a short induction period may yield more NPCs than a longer induction period, which would produce more mature neurons. In other words, the expressions of immature neural markers were significantly higher when treated with SMs for 12 h than that of 24 h. This is consistent with our previous study that showed the expressions of mature neural markers were significantly upregulated after treated with SMs for 14 days. The use of NPCs would be more favorable for neural tissue regeneration as they are more proliferative than mature neural cells. Moreover, the low proliferative capacity of MSCs has been reported to be one of the most intractable challenges hindering their clinical applications. 38 Therefore, in this study, we only treated GMSCs for 12 h prior to transplantation to ensure a higher fraction of extremely proliferative NPCs in the transplanted cells.
There are a number of experimental animal models of stroke that resemble human strokes. 39 Among them, MCAO is one of the most widely used and established stroke induction methods in rats and mice, which is mostly likely mimics the human strokes. 40 Hence, MCAO has been applied in the present study. As reported, the days postinfection (dpi) varies from 1 to 28 days.41–43 Based on our pilot study, we found that the mortality rate of the rats was high when brain stereotactic injection was performed earlier than 8 days after MCAO. In addition, if cell transplantation was performed soon after the acute phase of an ischemic stroke, which might increase the risk of infection and additional complications. If intracerebral therapy was initiated at a later time after ischemic stroke was established, progressive self-repair might confound extrapolation of treatment outcomes from rat model systems to humans. Therefore, the optimal timing of the intracerebral cell injection will need to be optimized in future studies. In vivo examination revealed that GMSC-derived neural lineage cells successfully migrated and colonized the injury site. Rats receiving neural lineage cells derived from GMSCs showed significantly better functional recovery than those receiving only undifferentiated GMSCs, thus suggesting that chemically preprogramming GMSCs toward neural lineage fate is clinically beneficial. HE staining revealed that GMSCs-derived neural lineage cells consistently reduced the volume of cerebral infarction to a greater extent than undifferentiated GMSCs. Consistent with these findings, rats in the SM-GMSCs group exhibited better recovery than those in the GMSCs group. These results thus suggest that GMSC-derived neural lineage cells are a superior treatment compared to GMSCs. This was further supported by significantly higher expression of both neural markers (TUJ1 and MAP2) and BrdU in the brains of rats in the SM-GMSCs group compared to GMSCs group. Collectively, the limited availability of neural cells for transplantation can be addressed by predefining the neural lineage of prereciprocal GMSCs through SM-based reprogramming. Moreover, we found that the cell proliferation in the SM-GMSCs group was higher than in the GMSCs group. One of the possible reason is that some of the SMs, such as, ISX-9, Y-27632 and purmorphamine could promote cell proliferation and increase the number of immature neurons or NPCs.44–46 However, this certainly requires more comprehensive investigations in future studies.
The direct transplantation of cells into infarcted brain tissues after stroke frequently results in substantial cell death. 47 To overcome this problem, we encapsulate the engrafted cells within the HA hydrogels to protect them from the harsh environment of the infarcted brain tissues. More neural lineage cells were found at the infarct site in the HA-SM-GMSCs group than in the SM-GMSCs group. In agreement, the HA-SM-GMSCs group demonstrated significantly better functional recovery than the SM-GMSCs group. Immunohistochemical staining revealed a significantly higher number of HuNu+TUJ1+ and HuNu+MAP2+ cells in the HA-SM-GMSCs group than in the SM-GMSCs group. Additional BrdU-positive cells were also present in the HA-SM-GMSCs group, suggesting that cells in the HA hydrogels have significantly higher neurogenesis capacity. 48 Similar to other self-assembling peptides and acellular extracellular matrix hydrogels, HA hydrogels can create a conducive microenvironment in the brain of rats with ischemic strokes.49–51 In particular, the mechanical properties of HA hydrogels are similar to that of brain tissue, which minimizes tissue inflammatory response and local scarring.27,52 Further advantages of HA hydrogels include easy handling and in situ gelling after transplantation in the body. 53 Therefore, HA hydrogels loaded with neural cells derived from GMSCs provide a favorable microenvironment that mimics the native ECM and facilitates the replacement of lost brain tissue by supporting the migration, differentiation and proliferation of transplanted cells, which facilitates functional recovery. It should be noted that there was also some functional improvement with HA hydrogels alone and PBS control, suggesting that the ischemic stroke rats had some capacity for self-healing. Ischemic lesions can induce axonal sprouting, and induce cell migration and differentiation. 54 Although the differences were not significant, HA hydrogels alone outperformed PBS in promoting healing, as demonstrated by behavioral tests and histological examinations. One possible explanation could be that HA hydrogels provide a better microenvironment to support the regenerative potential of endogenous adult stem cells.
It is speculated that reported that the SMs reprogrammed GMSCs into the neural lineage through simultaneous downregulation of dental cell type specific genes accompanied by the consequent up-regulation of endogenous neural transcription factors within these cells. Further studies are needed to elucidate the specific role of each component of the SMs cocktail in transforming GMSCs into neural lineage cells. It is worth noting that limited behavioral improvement is observed in the first few days, whereas large behavioral recovery is observed at later time points in all groups. This could be because the grafted cells are more affected by the host environment at earlier time points and require a certain period to adapt to the host environment before they can function appropriately may be due to both the postinjury host environment and intrinsic properties of the cells. 55 However, in this study we were unable to confirm this hypothesis because the rats were sacrificed at a later time point. Lastly, We did not attempt to directly distinguish between host and grafted BrdU+ cells in vivo, which is one of the limitations of this present study. This also will be the subject of our future investigations if we are able to get the markers of cell proliferation to distinguish between the host and grafted cells.
Conclusions
In summary, this study demonstrates that a cocktail of SMs can rapidly and efficiently induce differentiation of GMSCs neural lineage cells in vitro. Furthermore, intracerebral injection of a self-assembled HA hydrogels loaded with GMSCs-derived neural lineage cells in a rat model of ischemic stroke could promote recovery of neural function and reduce the ischemic damage. The HA hydrogels provide a supportive microenvironment that allows the transplanted cells to survive, proliferate, and differentiate into neural cells. These results demonstrate that HA hydrogels loaded with GMSCs committed to the neural lineages via SM treatment, could be a promising alternative therapeutic modality for ischemic stroke. However, further studies are needed to investigate the underlying molecular mechanisms responsible for the positive therapeutic effects of transplantation of GMSCs-derived neural lineage cells in animal models of ischemic stroke.
Footnotes
Acknowledgment
The authors gratefully acknowledge the assistance of the staff at the Laboratory Animal Center of Xuzhou Medical University for their help with the animal experiments.
Authors’ Contributions
S.J., C.Y., and T.Z.: contributed to conceptualization, data curation, formal analysis, writing—original draft, writing—review and editing, and visualization. K.J.H., M.B., Q.C., and J.L.: were involved in data curation and formal analysis. B.C.H. and L.W.L.: participated in writing—review and editing and funding acquisition. P.W.: contributed to conceptualization, writing—review and editing, and funding acquisition. C.Z.: provided conceptualization, writing—review and editing, supervision, and funding acquisition.
Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was supported by the