
Abstract
Rationale:
Elevated shear stress (ESS) induces vascular remodeling in veins exposed to arterial blood flow, which can lead to arteriovenous (AV) fistula failure. The molecular mechanisms driving remodeling have not been comprehensively examined with a single-cell resolution before.
Objective:
Using an in vivo animal mode, single-cell RNA sequencing, and histopathology, we precisely manipulate blood flow to comprehensively characterize all cell subpopulations important during vascular remodeling.
Methods:
AV loops were created in saphenous vessels of rats using a contralateral saphenous vein interposition graft to promote ESS. Saphenous veins with no elevated shear stress (NSS) were anastomosed as controls.
Findings:
ESS promoted transcriptional homogeneity, and NSS promoted considerable heterogeneity. Specifically, ESS endothelial cells (ECs) showed a more homogeneous transcriptional response promoting angiogenesis and upregulating endothelial-to-mesenchymal transition inhibiting genes (Klf2). NSS ECs upregulated antiproliferation genes such as Cav1, Cst3, and Btg1. In macrophages, ESS promoted a large homogeneous subpopulation, creating a mechanically activated, proinflammatory and thus proangiogenic myeloid phenotype, whereas NSS myeloid cells expressed the anti-inflammatory and antiangiogenetic marker Mrc1.
Conclusion:
ESS activates unified gene expression profiles to induce adaption of the vessel wall to hemodynamic alterations. Targeted depletion of the identified cellular subpopulations may lead to novel therapies to prevent excessive venous remodeling, intimal hyperplasia, and AV fistula failure.
Impact Statement
Developing therapies to control vascular remodeling and angiogenesis is of critical relevance to patients undergoing vascular surgery, especially those who suffer from arteriovenous fistula failure or vessel occlusion. Clinically effective pharmacotherapies to address this are currently not available. Recent work has shown that cellular cross talk among proinflammatory heterogeneous mechanically activated subpopulations is extremely important in many disease states, but the effects in vascular remodeling remain incompletely understood. Improving our understanding of how elevated shear stress affects vascular remodeling in multiple different cell types will help develop targeted therapeutics to either alleviate or intensify vascular remodeling.
Introduction
The connection of an artery and vein serves as a successful clinical option in multiple disease settings.1–4 For example, arteriovenous (AV) fistulas are performed by connecting a vein to the arterial system to promote the vein to remodel, thicken, and become stronger. This makes permanent hemodialysis possible and is the current gold standard to establish reliable access to the vascular system. In addition, vessel occlusion leads to devastating health consequences, such as myocardial infarction or lower limb amputations in diabetic patients, 5 and AV connections (bypass surgeries) can be used to circumvent the occlusions to alleviate the symptoms of the patient. Unfortunately, up to 50% of AV fistulas and 43% of bypass surgeries fail because of complications related to severe vascular remodeling or thrombosis, which contributes significantly to patient morbidity and mortality.6–8
These failures occur because when a vein graft is suddenly exposed to the elevated (arterial) blood flow, the constant, elevated shear stress (ESS) drives unpredictable vascular remodeling of the vein,9–11 such as wall thickening and vessel arterialization. 12 Mechanical forces in arterial vessels are as high as 50 dyne/cm2, whereas veins typically experience 1–5 dyne/cm2. 13 Although we know that shear stress activates a cascade of mechanical signaling pathways, 14 the exact effects of ESS on cellular cross talk among heterogeneous subpopulations remain incompletely understood.
Our laboratory has recently investigated heterogenous cell populations involved in a variety of disease states, including skin fibrosis, foreign body response in biomedical implants, and wound healing.15–22 In this study, we used tissue engineering in an in vivo animal model to precisely manipulate blood flow in vein grafts. By creating a group where the vein graft is exposed to ESS and a group where the vein graft is not exposed to elevated shear stress (NSS), we were able to comprehensively characterize all cell subpopulations important during vascular remodeling. Specifically, heterogeneous cell populations of endothelial cells (ECs) and macrophages demonstrated the most differential responses to ESS and were primarily responsible for driving angiogenesis through an upregulation of multiple pathways and cell-to-cell paracrine signaling.
Materials and Methods
Animals
Surgeries were performed on 20 female Lewis rats (Charles River Laboratories, Wilmington, MA, USA). Animals were kept on a 12-h dark/light cycle and had free access to food and water. All surgeries were performed using inhalation anesthesia with isoflurane (5% and 2.5%) in pure oxygen. Until postoperative day (POD) 2, animals were treated with heparin (80 IU/kg intra venous) und buprenorphine (0.05 mg/kg subcutaneously; Bayer). If animals displayed signs of pain, additional buprenorphine was given beyond POD 2. The animals were euthanized at the determined endpoints by intracardial injection of pentobarbital under deep anesthesia. The rate of thrombotic events usually ranges at around 20%, and grafts that showed a negative manual patency test (“milking test”) 23 at explantation were excluded from further analysis.
Microsurgical AV loop operation
These surgeries were performed using a 16× magnification (OPMI pico, Carl Zeiss). On the medial thighs of the rats, the saphenous artery and vein were exposed and dissected. An ∼20mm long vein graft was harvested from one side and end-to-end anastomosed between the saphenous vein and the artery of the contralateral side. Graft patency was controlled through the manual patency test (“milking test”). The AV loops were then placed into a polytetrafluoroethylene (PTFE) isolation chamber filled with acellular scaffold (MatriDerm®). By closing the lid of this isolation chamber, a defined spaced is created, which prevents ingrowth of surrounding vessels into the scaffold. As a control (NSS), anastomoses were created on the saphenous vein, and then it was placed into an isolation chamber with two opposed openings, eliminating the effect of the ESS on the vein graft.
Single-cell barcoding, library preparation, and single-cell RNA sequencing
After 7 (n = 10 biological replicates) or 14 days (n = 10 biological replicates), the venous tissue was explanted, microdissected, and enzymatically digested in 20 mL of collagenase type 1 (ThermoFisher) in phosphate-buffered saline (PBS) at a concentration of 5 mg/mL. 24 Subsequently, we minced and rotated the cell digest for 1 h at 37°C and then used a vortex mixer to further disrupt the remaining collagenous tissue. Using a 100 and 70 um nylon cell filter (ThermoFisher Scientific), we filtered the cells and quenched the digest in 10% fetal bovine serum Dulbecco’s modified Eagle’s medium. The solutions were spun at 300 g for 8 min at 4°C in a centrifuge to pellet the cells and then resuspended into a final concentration of 1,200 cells/uL in 0.04% bovine serum albumin (Sigma-Aldrich) in PBS, this in accordance with the maximum capture concentration short of overloading, as per specifications from 10X Genomics. This cellular suspension was submitted for droplet-based microfluidic single-cell RNA sequencing (scRNA-seq) using the 10× Chromium Single-Cell platform (Single Cell 3’ v3, 10X Genomics) at the Stanford Functional Genomics Facility (SFGF). To process on the Chromium Controller, we combined a droplet of the suspension, reverse transcription master mix, and partitioning oil. Reverse transcription was performed at 53°C for 45 min. The copy DNA (cDNA) was amplified for 12 cycles in total (Bio-Rad C1000 Touch thermocycler) with the cDNA size selected using SpriSelect beads (Beckman Coulter) and a ratio of 0.6 of SpriSelect reagent volume to sample volume. We used the Agilent Bioanalyzer High Sensitivity DNA chip to analyze cDNA for the purpose of quality control. The cDNA was then fragmented in the proprietary fragmentation enzyme blend for 5 min at 32°C, followed by end repair and A-tailing at 65°C for 30 min. cDNA was double-sided size selected using SpriSelect beats. Ligation of sequencing adapters to the cDNA occurred at 20°C for 15 min. The cDNA was then amplified using a sample-specific index oligo as a primer, followed by a second cycle of double-sided size selection using SpriSelect beads. An Agilent Bioanalyzer High Sensitivity DNA chip was used to analyze the final libraries for qualitative control purposes. cDNA libraries were sequenced on a HiSeq 4,000 Illumina platform aiming for 50,000 reads per cell.
Data processing, FASTQ generation, and read mapping
The Cell Ranger (10X Genomics; version 3.1) was used to convert the base calls into reads, implementing mkfastq, and then aligned against the rn6 (rat genome) using Cell Rangers count function with SC3Pv3 chemistry and 5,000 expected cells per sample. 25 Cell barcodes representative of quality cells were delineated from barcodes of apoptotic cells or background RNA based on the following thresholds: having less than 10,000 total transcripts, at least 200 unique transcripts, and less than 10% of their transcriptome of mitochondrial origin.
Data normalization and cell population identification
Unique molecular identifiers (UMIs) from each cell barcode were retained for all downstream analysis. The raw UMI counts were normalized with a scale factor of 10,000 UMIs per cell and subsequently natural log-transformed with a pseudocount of 1 using the R package Seurat (version 3.1.1). 26 Highly variable genes were identified, and cells were scaled by regression to the fraction of mitochondrial transcripts. Aggregated data were evaluated using uniform manifold approximation and projection (UMAP) analysis over the first 15 principal components. 27 The SingleR toolkit (version 3.11) was used to ascribe automated cell-level annotations.
Generation of characteristic subpopulation markers and enrichment analysis
Using Seurats’s native FindMarkers function, cell-type marker lists were generated with a log fold change threshold of 0.25 using the receiver operating characteristic (ROC) test to assign predictive power to each gene. Gene set enrichment analysis (GSEA) was performed on the 100 most highly ranked genes for each cluster of this analysis against pathway databases in a programmatic manner using EnrichR (version 2.1). 28 The search tool for retrieval of interacting genes (STRING) was used to acquire the protein–protein interaction networks using the top differentiating genes. 29
Immunofluorescent staining and confocal laser microscopy
Tissue was explanted and fixed in 4% paraformaldehyde and dehydrated in 30% sucrose in 1× PBS for 48 h. The tissue was then incubated in optimal cutting temperature compound (OCT; TissueTek, Sakuta Finetek) for 24 h at 4°C and then cryoembedded into tissue molds on dry ice. Frozen sections of 7 µm thickness were generated on a cryostat perpendicular to the longitudinal axis of the AV loop.
Antigen retrieval was performed using 0.01M sodium citrate buffer in PBS (Abcam). This was followed by blocking for 2 h in 5% goat serum (Invitrogen) in PBS. Sections where then incubated in the KLF2 polyclonal antibody (PA5-115479, ThermoFisher) and CD206 (MRC1) polyclonal antibody (PA5-114370, ThermoFisher) diluted 1:100 in 5% goat serum overnight at 4°C. Secondary antibodies were applied for 2 h at room temperature (goat anti-rabbit Immunoglobulin G [Heavy chain + Light chain], highly cross-adsorbed secondary antibody, Alexa Fluor plus 488, ThermoFisher). A Zeiss LSM 880 confocal laser scanning microscope at the imaging facility at the University of Arizona was used for imaging. High-resolution images of the tissue cross sections were obtained with 25× magnification, and automatic tile scanning was used to generate a scan of the entire tissue. The ZEN Black software (Carl Zeiss) was used to stitch the individual images together. Four region of interests of 400 × 400 µm were defined on four locations around the main vessel, including the endothelium. Quantification of the immunofluorescent staining was determined using MATLAB code developed in previous image analysis studies by the authors. 30
Statistics
Data are shown as mean ± SEM and were analyzed with Prism 8 (GraphPad). A Student’s t-test (unpaired and two-tailed) was used for comparisons of two groups. Normality was assessed using the D’Agostino and Pearson tests. The statistical methods used for scRNA-seq analysis are described in the specific sections previously.
Study approval
All animal experiments were conducted at the Hagey Research Laboratory at Stanford University, CA, USA, according to the Animal Welfare Act and were approved by the Administrative Panel on Laboratory Animal Care (APLAC 8778).
Results
We used an established AV loop model to expose vein grafts to ESS in vivo and explanted the vessels after 7 (n = 13) and 14 days (n = 14).32–34 To specifically interrogate how different cell types respond to ESS, we performed scRNA-seq on these cells using the 10X Genomics Chromium platform (Fig. 1A, B). A total of 6,622 cells were captured, with 3,550 at day 7 and 3,072 at day 14 (Fig. 1C). We identified 2,315 macrophages, 1,580 fibroblasts, 1,001 neutrophils, 611 ECs, 544 venous smooth muscle cells (VSMCs), 358 T cells, 195 dendritic cells (DCs), and 18 mast cells.
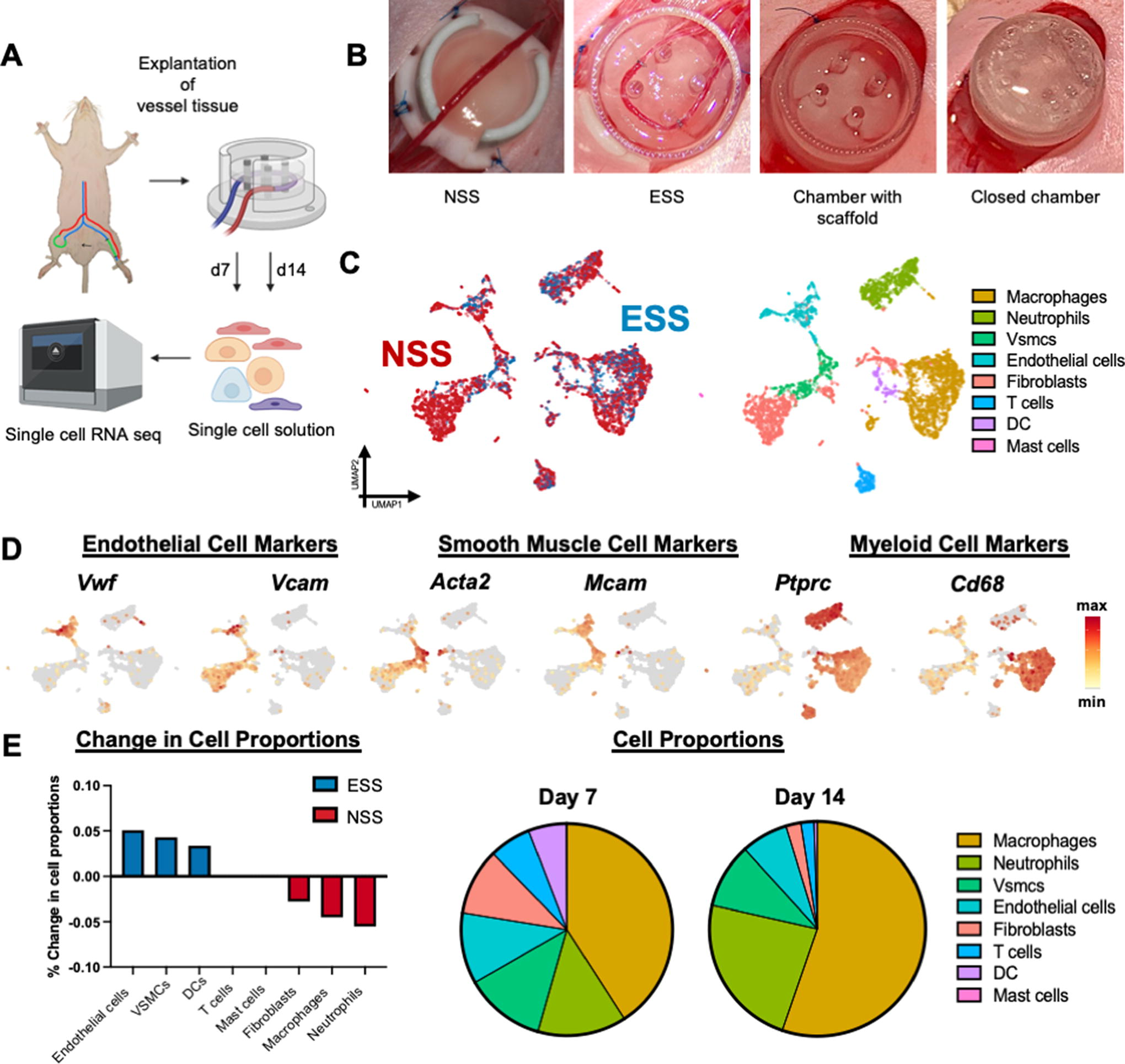
The presence of ECs was confirmed by the expression of canonical EC markers such as Vwf, Cd143, and Cd34 (Fig. 1D); VSMCs were identified by the expression of Acta2 and Mcam; myeloid cells by the expression of Ptprc and Cd68; fibroblasts by Pdgfra and Col1a1; neutrophils by Lcn2 and Ngp; T cells by Cd4; DCs by Ccr7 and Flt3; and mast cells by Cd63 35 (Fig. 1D, Supplementary Fig. S1). ESS promoted the largest increase in cell proportions in the ECs (Fig. 1E), whereas macrophages interestingly composed the highest proportion of total cells at both time points (Fig. 1E). Thus, we decided to interrogate the effects of shear stress on ECs and macrophages more closely.
ESS induces differentiation of distinct endothelial colony forming cell subpopulations in ECs
Unsupervised clustering of the ECs using the Seurat package revealed five heterogeneous and transcriptionally distinct EC subpopulations (Fig. 2A). ECs exposed to normal shear stress (NSS) were found to express heterogenous populations, which were relatively evenly distributed at about 11–27% representation per cluster (Fig. 2B). The “capillary-like” (C) cluster expressed Fabp4, which has been shown to be expressed mainly in the ECs of capillaries and not arteries (Fig. 2C, D). 36 The “lymphatic” (L) cluster was enriched with expression of Lyve1, a gene shown to be expressed in the lymphatic endothelium (Fig. 2C, D). 37 The angiogenic-fibrotic (AF) showed high expression of multiple genes associated with increased vessel formation, Pdgfrb, 38 regulation of angiogenesis, Tagln, 39 and binding to the extracellular matrix and cell viability, Crispld2 40 and Col6a1 41 (Fig. 2C). NSS also increased EC antiproliferation genes, such as Cav1, 42 Cst3, 43 and Btg1, 44 and the gene Emp1, 45 which negatively regulates cell growth. NSS promoted regulation of fibrinolysis (S100a10) 46 and cell morphogenesis (Lgals3) (Fig. 2E). 47 These clusters likely represent the native EC steady-state phenotypes in our system.
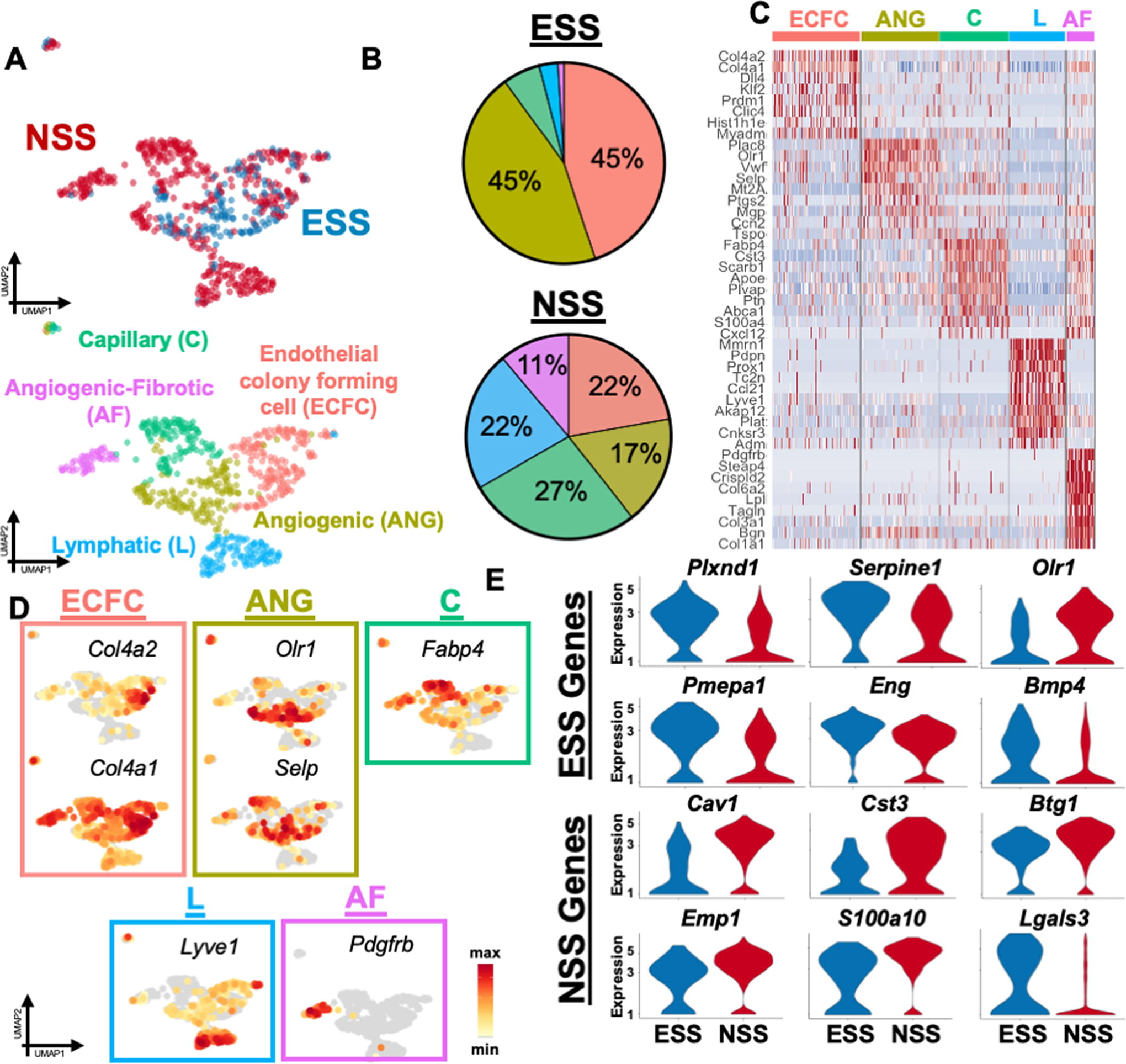
Interestingly, ESS shifted ECs toward a more homogeneous distribution of “endothelial colony forming cells” (ECFC) and “angiogenic” (ANG) cellular subpopulations (Fig. 2B). The ECFC subpopulation expressed extracellular matrix and basement membrane genes Col4a1 and Col4a2 (Fig. 2C, D), 48 which are the characterizing genes of the ECFC population and also involved in the differentiation of VSMCs. 49 These cells possess an extremely high ANG and proliferative potential and are necessary for the outgrowth of new vessels. 50 This cluster also showed upregulation of Kcne3, which is highly expressed in tip cells during angiogenesis. 51 Cells in cluster ANG expressed known markers for angiogenesis such as Olr1, Selp, and Ccn2 (Fig. 2D). Olr1 is responsible for the expression of adhesion molecules and is involved in the stimulation of proangiogenic markers, such as nuclear factor ‘kappa-light-chain-enhancer' of activated B-cells (NF-KB) and vascular endothelial growth factor (VEGF), in ECs and macrophages, 52 as well as Selp in stalk cells during angiogenesis. 53 Ccn2 has been shown to promote tube formation 54 and is necessary for pericyte adhesion. 55 Elevation of shear stress also induced an overall upregulation of mechanosensing genes Plxnd1, 56 Serpine1, 57 and Olr1, 58 as well as genes involved in the Transforming growth factor beta (TGF-ß) pathway (Pmepa1, Eng, Bmp4) (Fig. 2E).59–61 Genes promoting angiogenesis (Cxcl16) and VSMC proliferation (Nt5e) were also highly upregulated within the ESS group (Fig. 2E).62,63
Overall, these findings demonstrated that the elevation of shear stress promotes a differentiation of ECs into two homogenous subpopulations that are responsible for angiogenesis and cross talk between macrophages and VSMCs. Specifically, ESS upregulated the mechanical and angiogenesis pathways that guided macrophages to promote VSMC proliferation. In contrast, NSS promoted a more heterogeneous distribution of EC subclusters, with a preference for specific subclusters that instead inhibit angiogenesis and regulate cell organization.
ESS enriches activity of focal adhesion and TGF-β pathways in ECs
We next evaluated the biological function of the ECFC and ANG clusters and performed GSEA (Fig. 3A, B). The ECFC cluster most differentially upregulated the Mitogen-activated protein kinases (MAPK) pathway (Fig. 3A), a key mechanotransduction pathway that has been shown to induce cell proliferation and transmission especially in venous remodeling of hypoxic pulmonary hypertension.64–67 ANG ECs most differentially regulated the TNF signaling pathway (Fig. 3B), which is responsible for venous remodeling and lymphangiogenesis after inflammation. 68 This pathway is also involved in the migration of VSMCs, which interestingly depends on MAPK pathway activation. 69
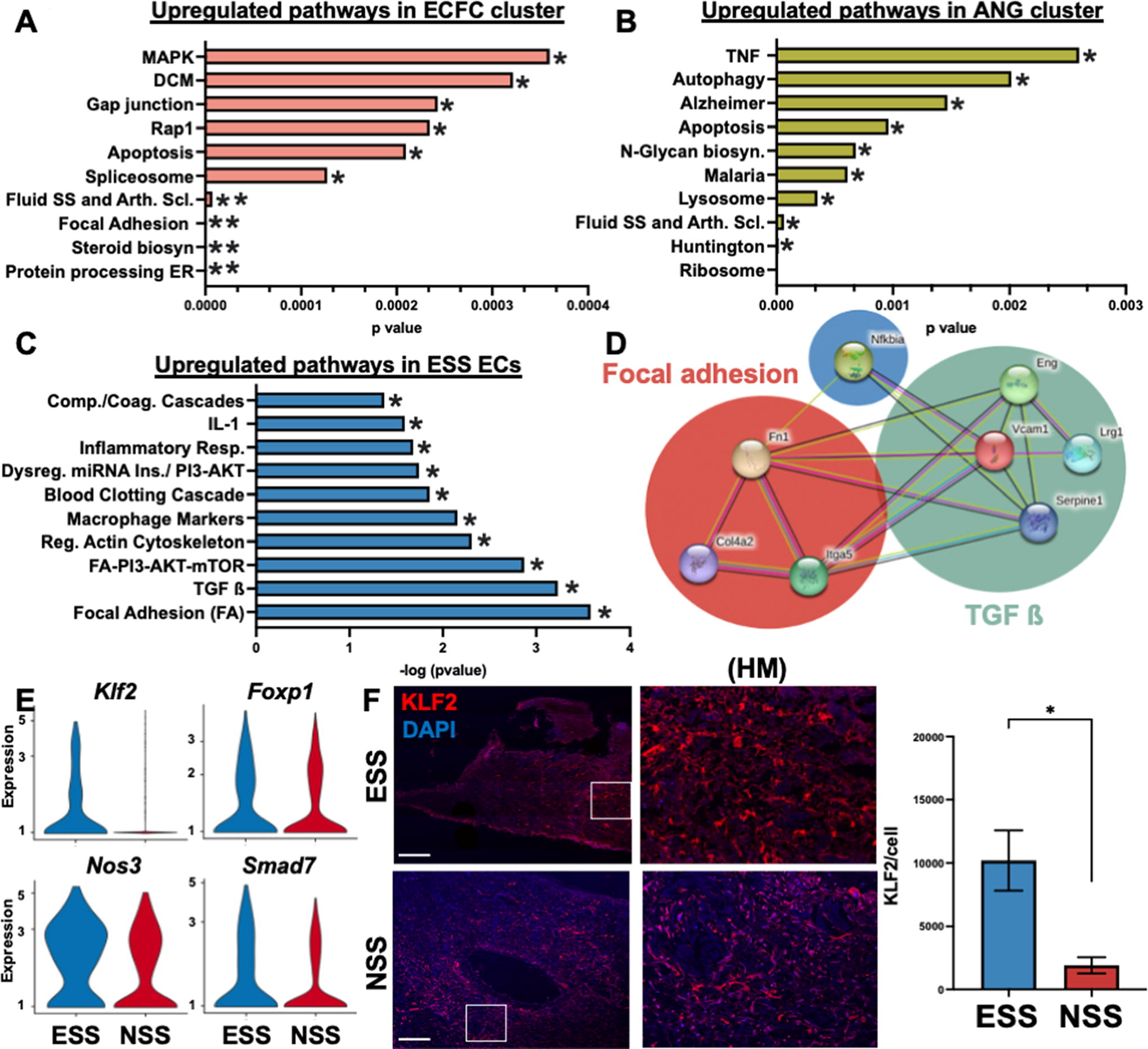
We then used the top 100 differentially expressed genes (DEGs) based on average log2 fold change to identify a variety of highly upregulated genes in the ESS ECs compared with NSS ECs. Pathway analysis of these genes performed with EnrichR showed an enrichment of the focal adhesion pathway, TGF-ß signaling pathway, and PI3-K-mTOR pathway (Fig. 3C). Using the STRING analysis tool on these genes, we created a protein–protein interaction network (Fig. 3D) and saw that there is a strong interaction between the genes involved in the TGF-ß signaling and the focal adhesion pathway, especially between Vcam1, Itga5, and Col4a2.
ESS promotes upregulation of EC protective factor Klf2
Endothelial-to-mesenchymal transition (EndMT) describes the process in which ECs transition into a more mesenchymal-like state. 70 We identified that ESS upregulated Klf2, which is a well-established inhibitor of vascular homeostasis that protects ECs from undergoing EndMT (Fig. 3F), 71 and other genes that inhibit EndMT, namely Klf2, Nos3, Foxp1, and Smad7. Klf2 inhibits EndMT through Nos3 and also upregulates Foxp1 and Smad7 to inhibit TGF-ß.72,73 To confirm these findings, we performed immunofluorescent staining and confirmed upregulation of the protein KLF2 in the ESS group compared with the NSS group on a protein level (Fig. 3F, Supplementary Table S1). These findings demonstrate that ESS overall promotes ECs to adapt to changes in shear stress by inhibiting EndMT through a KLF2 mechanism.
ESS promotes upregulation of distinct clusters of mechanically active macrophages to differentiate into a pro-inflammatory state
While previous work has identified the importance of ECs in vascular remodeling,74,75 few have investigated the importance of immune cell contributions during these functions. Specifically, although it has been noted that inflammation generated by immune cells is necessary for promoting angiogenesis,76–78 the exact transcriptional contributions are still not well understood. In our data set, we observed that macrophage subpopulations surprisingly comprised the highest cell proportions at both time points (Fig. 1E, Fig. 4A). Comparison of both immune cell defining (Ptprc, Cd68) and canonical “M2” anti-inflammatory macrophage markers (Arg1, Socs3, Cd200, Cd163) showed a similarly even distribution along the ESS and NSS groups (Fig. 4B). Using unsupervised clustering of the macrophages, we observed four distinct heterogeneous cellular clusters (Fig. 4C). Again, we observed that myeloid cells exposed to ESS demonstrated a more homogeneous distribution of the subclusters compared with NSS (Fig. 4D). Specifically, NSS promoted a distinct upregulation of an anti-inflammatory, and thus antiangiogenic, cluster that was barely evident in the ESS group (Fig. 4D). This cluster showed upregulation of the complement factor genes Cq1a, Cq1b, and Cq1c, which have been previously defined to be involved in mediation of EC adhesion and spreading, 79 as well as the anti-inflammatory Mrc1 (Cd206) (Fig. 4E).80,81 Using immunofluorescent staining, we confirmed that Mrc1 showed significantly higher protein expression in the NSS group (Fig. 4F, Supplementary Table S2). NSS cells also exhibited significantly higher expression of other genes associated with anti-inflammation such as Lyve1, 82 Clec10a, 83 and homeostasis (Fcgrt). 84 A very small percentage of NSS cells demonstrated an upregulation of alternative proinflammatory genes and genes involved in platelet aggregation (Treml1, Ly6g6f, Gp9),85–87 indicating the presence of an alternate path of inflammation in the absence of elevation in shear stress.
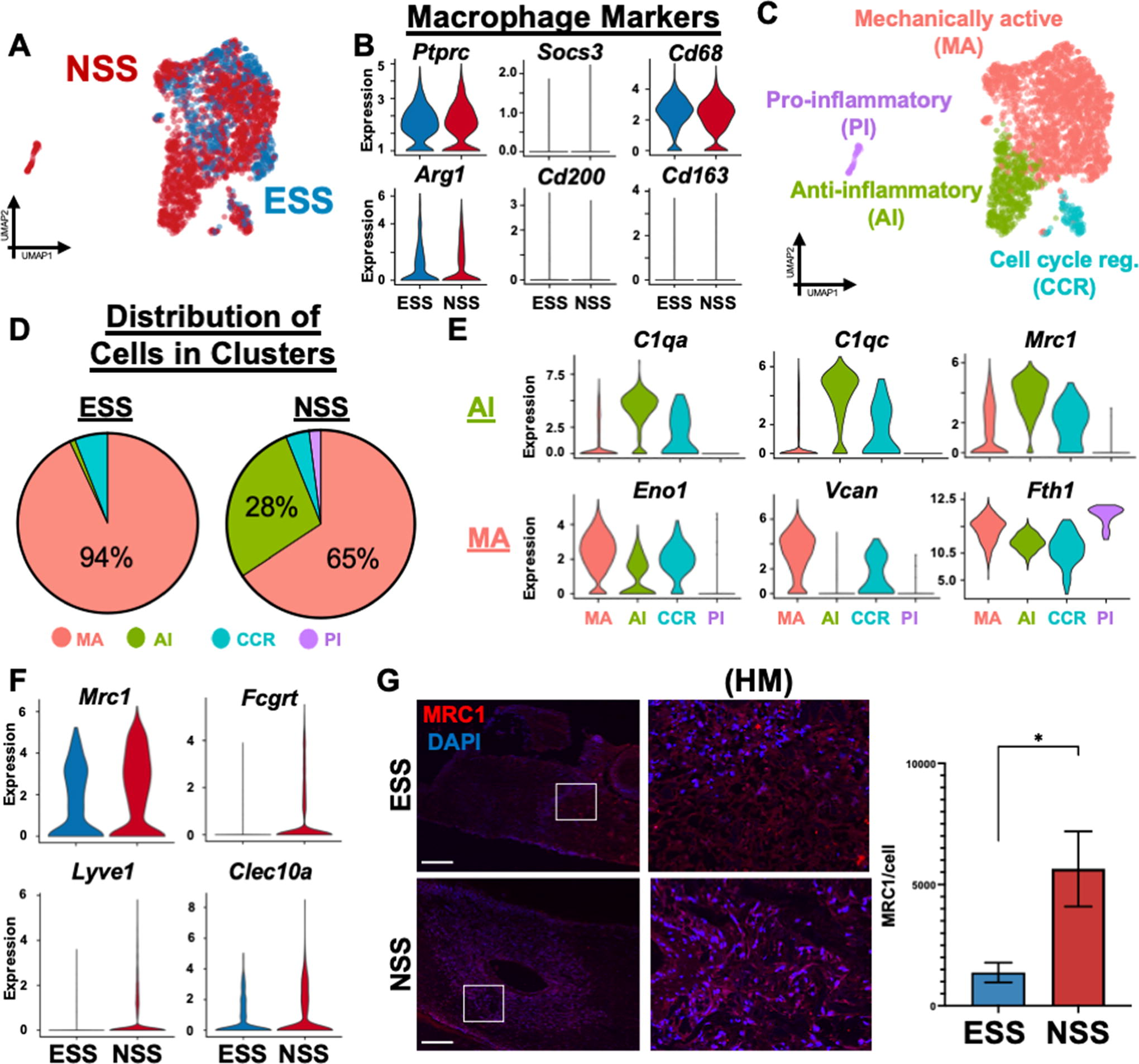
In contrast, ESS differentially drove >90% of all macrophages toward an almost homogenous expression of the “mechanically active” (MA) cluster (Fig. 4D). The MA cluster showed upregulation of genes involved in vascular remodeling (Slpi), 88 neointimal formation after arterial injury (S100a11), 89 and adhesion to the extracellular matrix (Fn1). 90 Fth, which was also upregulated in MA, has previously been shown to have a higher expression in M1 inflammatory macrophages, 91 with a protective, enhancing effect on angiogenesis. 92 It also exhibited an upregulation of Eno1, which contributes to the production of proinflammatory mediators, 93 as well as Vcan, which is part of the early inflammatory response and also involved in VEGF-dependent angiogenesis (Fig. 4E). 94 ESS also promoted 10% of macrophages to differentiate into a “cell-cycle regulation” cluster, which showed upregulation of multiple genes involved in regulation and proliferation within the cell cycle (Cdca3, Cenpw, Sgo2, Spc24).95–98
Overall, these findings show that elevation of shear stress promotes a more MA subpopulation of macrophages, which promote an enhancing effect on angiogenesis. In contrast, NSS induced a distinct anti-inflammatory phenotype of macrophages that were more involved in homeostasis and cross talk between ECs for mediating adhesion and spreading.
Discussion
AV shunts are commonly used in patients requiring recipient vessels for reconstructive microsurgery and1,2 to provide access for hemodialysis 99 and have been used for soft-tissue and bone tissue engineering because of their proangiogenic capacity.33,34,100–104 After an AV shunt is placed, venous tissue is exposed to ESS on an arterial level. This change in fluid dynamics induces multiple changes and adaptations of the venous vessel wall. In a clinical setting, the arterial-like levels of shear stress drive intimal hyperplasia and venous wall remodeling.105,106
Multiple studies have shown that elevated (arterial-like) blood flow in veins promotes changes in angiogenesis and vessel formation within the vessel wall.107–109 Vascular ECs have traditionally been identified as the primary cell type that react to increased shear stress and the main cell type responsible for paracrine signaling to various other cells.110–112 Myeloid cells, especially macrophages, are involved in acute inflammation after soft tissue injury and have recently been identified as being mechanically sensitive in the context of a number of biological functions, such as including touch, hearing, balance, and proprioception.113–115 However, none of these studies fully examined the depth of transcriptional changes and cellular signaling pathways activated or inhibited after exposure to ESS. Advances in scRNA-seq provide us with the opportunity to investigate the vascular response to ESS for the first time in depth and using an in vivo nondisease model.
In our study, we used AV shunts in a tissue-engineered construct to rapidly induce neoangiogenesis, efficiently creating a vascularized scaffold with cellular infiltration without the addition of growth factors. We observed that ESS most differentially influenced the transcriptional profiles of ECs, driving them toward a homogenous cell population within ECFC and ANG subpopulations. Specifically, the ECFC cluster cells were primarily driven by the MAPK mechanotransduction pathway, which is highly involved in cell proliferation in the early stages of angiogenesis. ECFC cluster cells also demonstrated increased involvement in gap junctions, which are highly important for cell–cell communication and transmittance of chemical and electrical signals 116 and corroborate our previous findings identifying upregulation of connexin 43 from ESS. 10 In contrast, a normal level of shear stress promoted heterogeneous cell populations such as those involved in many kinds of different locations such as capillaries or lymphatic vessels, or negatively regulated proliferation or cell growth.
ESS ECs also demonstrated increased expression of focal adhesion-PI3-Akt-mTOR pathways, as well as TGF-ß, which critically activates EndMT.117,118 During EndMT, ECs de-differentiate into a more stem-like phenotype, losing primary EC functions such a bloodstream-to-tissue exchange regulation or intercellular growth signaling.119,120 Although excessive EndMT leads to eventual graft failure and thrombosis,121,122 a complete lack of EndMT causes shunt immaturity, eventual failure, and lack of use for hemodialysis. Although multiple in vitro studies have tried to induce EndMT,123–126 studying EndMT in vivo has been challenging because of a lack of reliable animal models, 119 furthering our incomplete understanding of these paradoxical relationships. Using our animal model, we were able to reliably induce EndMT in vivo with ESS and carefully interrogate the cellular response. We found that ECs balance EndMT by promoting Foxp1, Nos3, and Smad7 to inhibit EndMT, as well as Krüppel-like-Factor 2 (Klf2), which specifically has a protective function on the vessel wall.71,127
Along with strong changes in EC subpopulations, we also observed that ESS had a strong transcriptional effect on myeloid cells. Although we have recently investigated mechanosensitive macrophages in wound healing,24,128 the mechanisms driving vessel formation remain incompletely understood. Our data indicate a previously unappreciated importance of myeloid mechanosensitivity during angiogenesis, 129 showing that an elevation of shear stress in the vein graft promotes the upregulation of a distinct cluster of MA macrophages, which may communicate with ECs to drive vessel sprouting via VEGF signaling. 130 A normal level of shear stress instead promotes an anti-inflammatory, and thus an antiangiogenic cluster of macrophages primarily defined by the expression of C1qa, C1qc, and Mrc1, which interact with neighboring macrophages 131 and ECs. 79
A limitation of this study is the lack of insight into the heterogeneity of venous remodeling within the graft wall. When blood flow is altered, mean pressure, shear stress, flow velocity, vascular resistance, and the Reynolds number also changes. In the context of the AV loop, the altered components are mainly flow velocity and shear stress. 9 However, recently published literature has demonstrated that intraluminal pressure also modulates endothelial function and vascular remodeling, 132 and the venous graft matures heterogeneously when exposed to these alterations. Future research should include spatial transcriptomics of the different segments of the venous graft within the AV loop model to identify transcriptomic changes related to regions of low shear stress (inner curve) and high shear stress (outer curve).
Overall, we have comprehensively characterized the cellular effects of mechanical signaling on venous remodeling and found that the response of EC and macrophage subpopulations to sudden changes in mechanical flow should be carefully considered for patients requiring procedures with AV connections (Fig. 5). Our findings provide a better understanding of how to use shear stress to manipulate and shift the heterogenous cell populations of the vessel walls across various clinical and translational applications. These findings could be applied to several vascular disease states, such as promoting the maturation of venous fistulas to improve vascular access for hemodialysis or preventing shunt or graft failure in cardiovascular surgery. Improving our understanding of these interconnected cellular relationships could improve overall patient survival and quality of life.
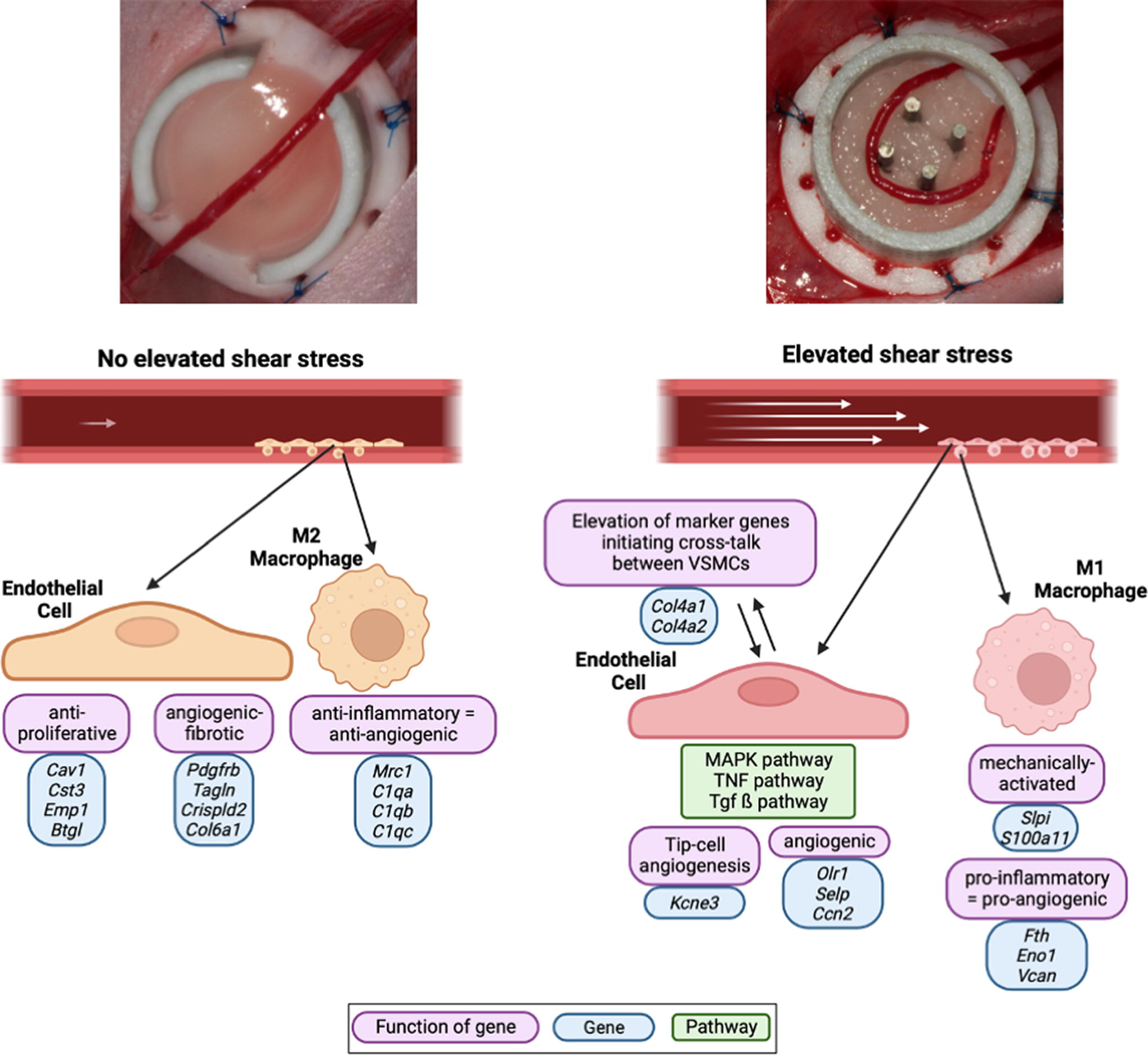
Schematic of the proposed cellular mechanism of action showing how increased shear stress drives cellular cross talk and proangiogenic pathway upregulation, while no elevated shear stress promotes an anti-inflammatory, more fibrotic state.
Footnotes
Acknowledgments
The authors thank Theresa Carlomagno for her administrative support.
Data availability
The scRNA-seq data discussed in this publication have been deposited in National Center for Biotechnology Information’s Gene Expression Omnibus 31 and will be made accessible through a Gene Expression Omnibus series accession number.
Authors’ Contributions
K.S.F., D.H., K.C., and G.G.: designed the study. K.S.F.: performed the animal experiments. K.S.F., D.H., D.S., and K.C.: performed the single-cell RNA sequencing experiments. K.S.F., D.H., E.T.Z., and K.C.: performed the scRNA-seq analysis. D.S., B.L., W.H., A.B.K., M.G.M.P., H.K., A.C.H., and M.G.: performed histological staining. K.S.F., B.L., S.M.M., and J.G.: performed the confocal imaging. K.S.F., K.C., and G.G.: wrote the article. D.H., D.S., J.H., and U.K.: helped revise and edit the article.
Disclosure Statement
The authors have declared that no conflict of interest exists.
Funding Information
Single-cell sequencing was supported by the SFGF with funds from the National Institutes of Health (S10OD018220 and 1S10OD021763). Confocal imaging was supported by the National Cancer Institute of the National Institutes of Health under award number P30 CA023074.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.