
Abstract
Osteoarthritis, a degenerative disease of articular cartilage and the leading cause of disability, is preceded by acute cartilage injury in a significant proportion of cases. Current auto- and allograft interventions are limited by supply and variability in therapeutic efficacy, prompting interest in tissue engineering solutions. Cell sheet tissue engineering, a scaffold-free regenerative technique, has shown promise in preclinical and clinical trials across various cell types and diseases. Polydactyly-derived juvenile cartilage-derived chondrocyte (JCC) sheets from juvenile patients are a potent cell source for developing allogeneic therapies. JCC sheets have proven safe and effective in animal models and as an add-on therapy in a recent clinical cartilage repair study. However, JCC ex vivo expansion leads to de-differentiation, contributing to long healing times. This study hypothesized that in vitro differentiation of JCC sheets into hyaline-like cartilage constructs could accelerate cartilage regeneration without compromising implant integration. To this end, sheet integration, maturation, and healing of conventionally prepared vs. differentiated JCC sheets were compared in an established nude rat focal chondral defect model. Differentiated JCC sheets exhibit mature cartilage phenotypes prior to transplant. Both conventional and differentiated JCC sheets are reliably transplanted without additional fixation. Histological evaluation reveals that both transplant groups produced equivalent neocartilage regeneration, filling defects with mature hyaline cartilage at 2- and 4-weeks post-transplant. Notably, differentiated JCC sheets respond to in vivo signals, undergoing matrix remodeling and integration with adjacent and subchondral tissue. Given equivalent healing outcomes, the future utility of in vitro JCC sheet predifferentiation from other JCC donors with different healing capacities should be balanced against their increased culture costs over conventional sheets.
Impact Statement
This preclinical study compares regenerative outcomes of differentiated juvenile cartilage-derived chondrocyte (JCC) sheets against conventionally prepared, clinically demonstrated JCC sheets in an established nude rat focal chondral defect model. Predifferentiated JCC sheet validation provides the opportunity for cartilage cell sheet donor variability and product quality control prior to implantation but comes at a significantly increased cost of goods in culture additives (e.g., exogenous growth factors, cytokines, and chemokines). After 1 week of in vitro chondrogenesis, JCC sheets produce mature, hyaline cartilage phenotypes and are able to engraft, integrate, and mature at the defect site, ultimately producing equivalent cartilage healing to conventional JCC sheets as assessed by blinded O’Driscoll (OD) scoring. These findings motivate the broader investigation of in vitro sheet differentiation as a strategy to address known donor-derived variability in JCC potency—a cell therapy critical quality attribute essential to human translation.
Introduction
Osteoarthritis (OA), a degenerative articular cartilage disease affecting over 300 million people globally, is a leading cause of disability.1–3 Among myriad disease triggers, acute cartilage injury producing chondral defects is the etiological precursor in 12% of cases. 4 Cadaveric osteochondral allografts and osteochondral autograft transfer system (OATS), both considered current “gold standard” treatments for chondral defects,5,6 and increasingly used autologous chondrocyte implantation (ACI), matrix-induced ACI (MACI), and microfracture strategies, all have challenges and risks. 7 Thus, broad interest remains in tissue engineering strategies for cartilage therapies that reliably restore tissue integrity and halt catabolic cascades propagating disease progression. Scaffolded approaches, in which cells are seeded on or within a biomaterial (e.g., MACI on collagen mats, and many preclinical approaches), are commonly used to 1) recapitulate mechanical properties of cartilage, 2) enable 3D cell-cell interactions necessary for chondrogenesis, and 3) aid in localization of cells at target sites using structures secured using sutures or fibrin glue or via in situ gelation.8–11 Despite these benefits, scaffolds introduce known biocompatibility challenges stemming from host reactions to foreign material and degradation products that can hamper transplant integration with host tissue.9,10,12,13 Accordingly, scaffold-free approaches are increasingly investigated as evidence shows that transplant of cell-dense chondrocyte constructs exploits endogenous transplant-host signaling to regenerate cartilage defects similarly to native cartilage development while avoiding interfering biomaterials-associated challenges.9–11,13
Cell sheet tissue engineering is a clinically demonstrated, scaffold-free regenerative approach applied in 45 registered clinical trials in a variety of cell types and diseases to date. 14 Cell sheets are produced using commercial thermo-responsive cultureware that enables the release of confluently cultured cells without proteolytic enzymes (e.g., trypsin) via moderate culture temperature change from 37°C to below 32°C.15–17 This process preserves endogenous extracellular matrix (ECM) and surface proteins 18 that enable cell sheet spontaneous adhesion to and integration with biological surfaces without biomaterial support.14,16,17
The use of human chondrocyte sheets to treat OA lesions in the knee was first established in 2006 using adult chondrocytes isolated from cartilage from total knee arthroplasty. 19 However, adult chondrocyte sheet preparation requires synoviocyte feeder layer culture support and clinically relies upon autologous sourcing, ultimately limiting clinical applicability due to inconsistent quality.20,21 Human chondrocyte cell sheets are reported to be a viable cartilage regeneration strategy in two clinical studies.20,22 Most recently, juvenile cartilage-derived chondrocytes (JCC) sourced from juvenile polydactyly reconstruction surgical discards have demonstrated higher proliferative and in vitro chondrogenic potentials than adult chondrocytes,23–25 and have already been exploited as an allogeneic cell sheet alternative.22,25–27 JCC sheets were safe and efficacious in treating cartilage defects in small animal models (rat, 26 rabbit 27 ) and demonstrated promising results as add-on therapy in a recent 10-person OA patient clinical study. 22 In each study, long healing times were needed, due possibly to chondrocyte de-differentiation during expansion culture23,28,29 required for cell sheet production. All conventionally prepared human chondrocyte sheets lack mature cartilage phenotypes at the time of culture detachment.19–22,25–27 Preliminary work exploring early-stage JCC sheet-induced defect healing in an immuno-deficient rat defect model suggested regenerated neocartilage is initially fibrocartilaginous before rapidly acquiring mature hyaline-like phenotypes by 4 weeks. 30 Human mesenchymal stem cell (MSC) sheets have been stimulated in earlier work to undergo robust chondrogenic differentiation in vitro, producing thick hyaline-like tissue that maintains cell sheets’ ability to adhere to cartilage without biomaterial support.12,31,32 Thus, we hypothesize that hyaline-like JCC sheets produced by in vitro differentiation would accelerate chondral defect healing without compromising transplant integration. In this study, integration, maturation, and healing of conventional JCC sheet vs. differentiated JCC sheet transplants were compared in a nude rat model of chondral defects at 2- and 4-weeks posttransplant (Fig. 1).
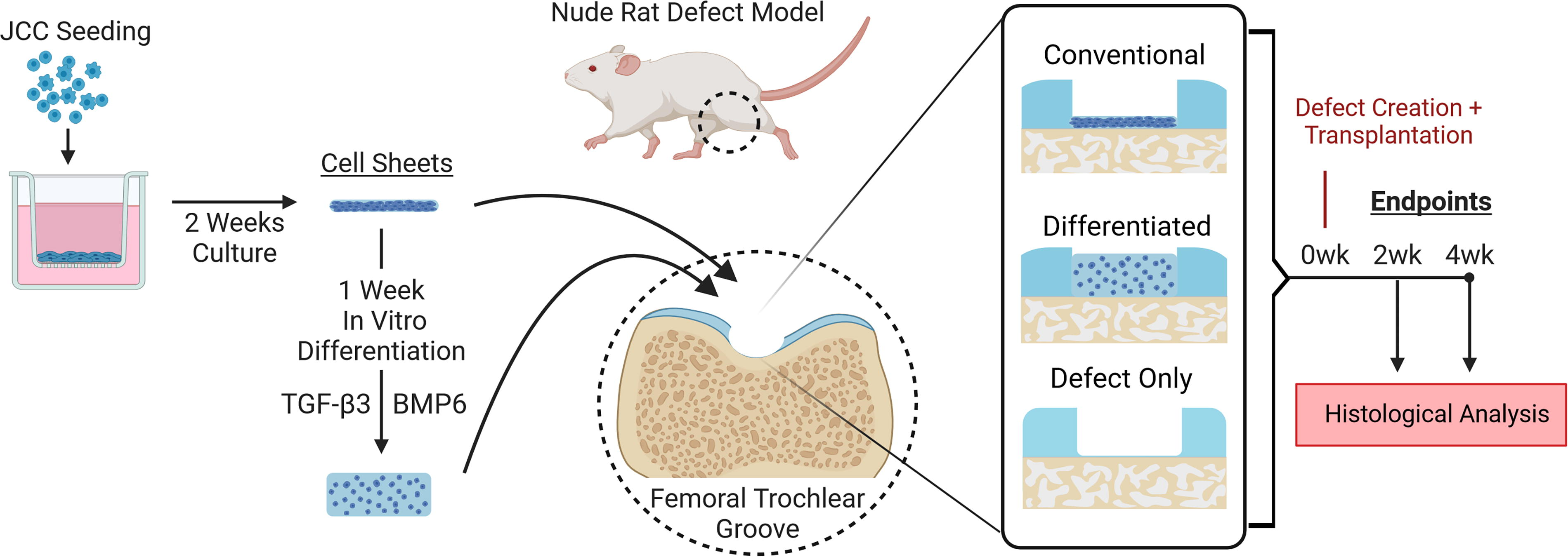
Schematic of JCC cell sheet experimental design. (created with Biorender.com). JCC, juvenile cartilage-derived chondrocytes.
Methods
Cartilage sourcing
Human articular cartilage from metacarpal and phalanges joints were dissected from an amputated polydactylous finger of a deidentified juvenile female patient (age: 14 months) enrolled at Intermountain Primary Children’s Hospital (Salt Lake City, USA). Institutional review board oversight from the University of Utah and Intermountain Primary Children’s Hospital was waived as donated tissue is a routine surgical discard and deidentified. Immediately following surgical removal, the discarded tissue was stored in saline for less than 1 hour until chondrocyte isolation began following published methods. 26
Chondrocyte isolation and cell culture
Cartilage harvested from human donor tissue was minced into <1 mm3 pieces by scalpel and then incubated at 37°C for 1.5–3 h in 5 mg/mL solution of type I collagenase (LS004197, Worthington Biochemical) in DMEM/F12 (ThermoFisher Scientific). Cells were then filtered through a 100 μm cell strainer, washed with saline, and resuspended in a chondrocyte culture medium consisting of DMEM/F12 (ThermoFisher) with 20% fetal bovine serum [(FBS), ThermoFisher] and 1% Antibiotic-Antimycotic, 100X (ThermoFisher). Isolated chondrocytes were seeded on polystyrene cultureware (CELLTREAT) at a density of 5,000–10,000 cells/cm2 and the medium was supplemented with 100 µg/mL L-ascorbic acid phosphate magnesium salt n-hydrate (Fujifilm Wako Pure Chemical) at the first medium change on day 4. Cells were collected at ∼90% confluence by TrypLE Select (ThermoFisher) dissociation, then counted and cryopreserved at passage 1 in STEM-CELLBANKER GMP grade (Zenoaq). Passage 1 thawed cells were plated at 10,000 cells/cm2 for expansion culture and passaged at 3–5 days for use in cell sheet preparation at passage 2. Banked cells used in this study were previously validated as regenerative in rodent focal chondral defects.26,33
Fabrication and differentiation of juvenile chondrocyte sheets
JCC cell sheets were prepared from cells at passage 2 seeded on temperature-responsive 6-well cell culture inserts (CellSeed). Chondrocyte culture media was changed on days 4, 6, 8, 10, and 12, and the cell sheets were detached on day 14 with forceps after 20 min at room temperature. After detachment, cell sheets were either transplanted as undifferentiated sheets or prepared for chondrogenic differentiation.12,31 Cell sheets were detached, allowed to contract, transferred onto insert culture membranes preincubated in 1 mL FBS overnight, and then centrifuged at 2,970 × g and 37°C for 10 min to facilitate rapid adhesion. After three days in fresh chondrocyte culture media, an 8-mm disc was cut from the sheet using a biopsy punch, and culture media was replaced with chondrogenic differentiation media. Cell sheets were differentiated for one week in chondrogenic differentiation media, with a media change on day 4. Chondrogenic media comprised high glucose Dulbecco's Modified Eagle Medium [(DMEM), ThermoFisher] supplemented with 200 ng/mL bone morphogenic protein 6 [(BMP6),120–06, Peprotech], 10 ng/mL transforming growth factor beta-3 [(TGF-β3), 100-36E, Peprotech], 1% insulin-transferrin-selenium (ThermoFisher), 1% penicillin-streptomycin (ThermoFisher), 1% nonessential amino acids (ThermoFisher), 100 nM dexamethasone (MP Biochemicals), 1.25 mg/mL bovine serum albumin (Sigma-Aldrich), 50 µg/mL ascorbic acid, 40 µg/mL
Nude rat cartilage defect model and juvenile chondrocyte sheet transplantation
The rodent study plan was approved by the Institutional Animal Care and Use Committee (IACUC, University of Utah, ID 20-12001). Nude rats (6 weeks old) were purchased (Charles River, Wilmington, USA) and acclimatized for a week in the animal facility. Surgery and transplant followed previously reported protocols. 26 Rats were anesthetized using isoflurane and O2 gas. The right knee joint was exposed using a medial parapatellar incision followed by lateral dislocation of the patella. A focal chondral defect with a 2 mm diameter and 300–400 µm depth was made using a small rotary grinder under a surgical stereo zoom microscope (SZX10, Olympus). Cell sheets to be transplanted were washed with saline and placed into the defect for approximately 40 min to allow for spontaneous attachment. After culture release, conventional cell sheets were cut in half with a razor blade, and single half-sheets were transplanted per defect. For differentiated cell sheet transplants, cell sheets were removed from insert culture membranes, and a 1-2 mm plug (formed using a biopsy punch) was transplanted per defect. Conventional JCC sheet transplant following published protocols,26,33 uses half sheets as they are compliant enough to fit within the defect. However, the mechanical stiffness of differentiated JCC sheets prevents half-sheets from fitting in the defect; therefore, 2 mm plugs were used for differentiated JCC sheets to fit defects. Both procedures completely covered the respective defects with JCC cell sheets. All animals received intramuscular buprenorphine (0.01–0.05 mg/kg) injections beginning 2 h prior to surgery and every 12 h for 2 days. Subcutaneous injection of carprofen (5 mg/kg) proceeded daily for 3 days postsurgery. Animals were sacrificed after 2 and 4 weeks and knee joints were surgically harvested and fixed for histological evaluation as described below.
Histological evaluation
Cell sheets prior to transplant were fixed in 4% paraformaldehyde (ThermoFisher) for 30 min at room temperature. Harvested rat knee tissue was fixed in 4% paraformaldehyde for 4 days and decalcified in RapidCal Immuno (BBC Biochemical) for an additional day at room temperature. Samples were paraffin-embedded and 5 μm transverse sections were prepared with a microtome. Slides were deparaffinized by overnight incubation in an oven at 65°C, followed by xylene washes. Sections were rehydrated by gradual ethanol replacement with deionized water. Sections were stained with combinations of 0.1% safranin O (5 min, Sigma-Aldrich), with 0.5 g/L Fast Green (5 min, Sigma-Aldrich), and Weigert’s iron hemotoxylin (4 min, Sigma-Aldrich) counterstaining. Images were taken with a BX41 microscope (Olympus) and AmScope Software. Safranin O-stained sections were graded using a modified O'Driscoll (OD) score 34 by a blinded observer (grading system detailed in Supplementary Table S1). Where heterogeneity in regenerated tissue was present, histological OD subscoring for categories I, III.A, III.B, and VI was determined for each of the distinct regions, weighted by percent of total regenerated tissue area determined using FIJI (v1.53c, NIH), and summed to produce a single subscore.
Immunohistochemistry
Sections were rehydrated as described above then antigen retrieval was performed using protease K (S3010, Agilent Technologies) for type II collagen and osteocalcin (OCN). No antigen retrieval was needed for type I collagen, human vimentin (hVim), aggrecan, or proliferating cell nuclear antigen (PCNA). Peroxidase blocking was performed using 3% hydrogen peroxide in PBS followed by nonspecific blocking with 5% donkey serum (Jackson ImmunoResearch) and 0.1% Triton-X in PBS for 1 h. Samples were then incubated overnight with primary antibodies at 4°C. Polyclonal goat anti-type I collagen (1:200, SouthernBiotech), monoclonal mouse anti-type II collagen (1:200, 2B1.5, ThermoFisher), monoclonal rabbit anti-hVim (1:200, SP20), polyclonal goat anti-aggrecan (1:100, R&D Systems), monoclonal mouse anti-OCN (1:50, G-5, Santa Cruz Biotechnology), and monoclonal mouse anti-PCNA (1:200, PC10, Santa Cruz Biotechnology) were used as primary antibodies. Normal mouse IgG2a (X0943, Agilent), normal goat IgG (NI02, MilliporeSigma), or normal rabbit IgG (X0903, Agilent) were used as isotype controls at identical concentrations as primary antibodies. Horseradish peroxidase (HRP)-conjugated donkey antigoat antibody (1:1000, 705–035-147, Jackson) was used for type I collagen and aggrecan staining. HRP-conjugated goat antirabbit antibody (1:1000, 111–035-144, Jackson) was used for hVim staining. ImmPRESS HRP horse antimouse polymer detection kit (MP-7422, Vector Laboratories) was used for type II collagen, OCN, and PCNA staining. ImmPACT DAB Peroxidase (HRP) Substrate (SK-4105, Vector Laboratories) was used as a chromogen. Brightfield images were taken with a BX41 microscope and AmScope software.
Statistical analysis
Numerical results of OD scoring are presented as means with standard deviations for each treatment group and time point. Data normality was assessed using a Shapiro–Wilk test. Statistical analysis was conducted by Welch’s ANOVA with post hoc Games–Howell test. Statistically significant differences were defined as p < 0.05 and are denoted by *.
Results
In vitro preparation and characterization of JCC sheet transplants
Similar to previous reports,22,25,26 conventional human JCC sheets harvested from thermo-responsive culture comprise about 2 × 106 cells 26 and are recovered intact as thin, macroscopically translucent, slightly contracted circular constructs (Fig. 2A). They do not stain with either safranin O or collagen type II by immunohistochemistry (IHC) (Fig. 2C, D) while staining positively for collagen type I and aggrecan (Fig. 2E, F). This indicates that constituent chondrocytes in sheets have dedifferentiated during culture as is common to monolayer cultured chondrocytes,29,35,36 (Fig. 2C–E). After one week of chondrogenic culture in vitro, differentiated JCC sheets appear opaquely white with a glassy surface (Fig. 2B) and become 10-fold thicker than conventional JCC sheets (345 ± 82 μm vs. 31 ± 3 μm, Fig. 2H). Histologically, differentiated JCC sheets acquire mature cartilage phenotypes, including matrix composition, seen in Figure 2H–K as deep, even staining for safranin O, collagen type II, and aggrecan, and chondrocyte behavior seen as condensed, nonproliferating (PCNA negative, Fig. 2L) chondrocytes within lacunae. Despite this maturity, differentiated JCC constructs continue to stain positively for collagen type I (Fig. 2J).

In vitro differentiation of JCC sheets produces hyaline-like phenotypes. Undifferentiated
Transplantation and macroscopic comparison of regeneration
Differentiated JCC sheets could be handled with forceps without risk of tearing and did not bend or fold under their own weight during manipulation. Both transplants readily and spontaneously adhere to the cartilage defect area created in vivo in rodent joints in approximately 40 min. Neither sutures nor fibrin glue were required to secure sheets into defect sites. At the time of transplant, differentiated JCC sheets alone filled the defect site reliably and restored the smooth glassy surface of the cartilage. Two weeks posttransplantation, differentiated JCC transplants visually match native cartilage color and opacity (Fig. 3I, J), but a distinct border between transplant and host cartilage is seen, suggesting incomplete tissue integration. By contrast, neocartilage in conventional JCC sheet transplants appears more translucent and fills the defect area (Fig. 3G, H). By 4 weeks, both transplant groups fill defects with visually identical neocartilage, with only some small nonunions of sheets with host cartilage seen in the differentiated JCC transplants (Fig. 3M, N, O, and P, respectively). Similar to past work in this defect model, 26 no defect healing is seen at two weeks (Fig. 3K, L) while translucent tissue covers defect sites at 4-week timepoints in the defect-only controls (Fig. 3Q, R).

JCC sheets were successfully transplanted and produced neocartilage in vivo. Stereomicroscope images of treated knees at the time of transplant (0 weeks)
Comparison of sheet in vivo regeneration and histological scoring in the rodent cartilage defect model
Histological sections from rodent defect sites harvested at different time points were compared. Safranin O staining of formalin-fixed, paraffin-embedded sections (Fig. 4A) shows that conventional JCC sheets rapidly matured within 2 weeks in vivo, developing hyaline-like morphologies and integrating well with host tissue (Supplementary Figure S1). Differentiated JCC sheet hyaline-like phenotypes from the time of transplant are maintained at two weeks posttransplant, with a high degree of integration with surrounding host tissue and defect filling (Fig. 4A). At 4-weeks posttransplantation, both differentiated and conventional JCC sheet transplants present with deeper safranin O staining and increased lacuna formation and full defect filling (Fig. 4A), indicating ongoing maturation. Subchondral bone reforms to abut the regenerated neocartilage at 4 weeks in transplant groups (Fig. 4A). No neocartilage regeneration is seen in defect-only groups (Fig. 4A) with minimal defect filling and extensive subchondral disruption.
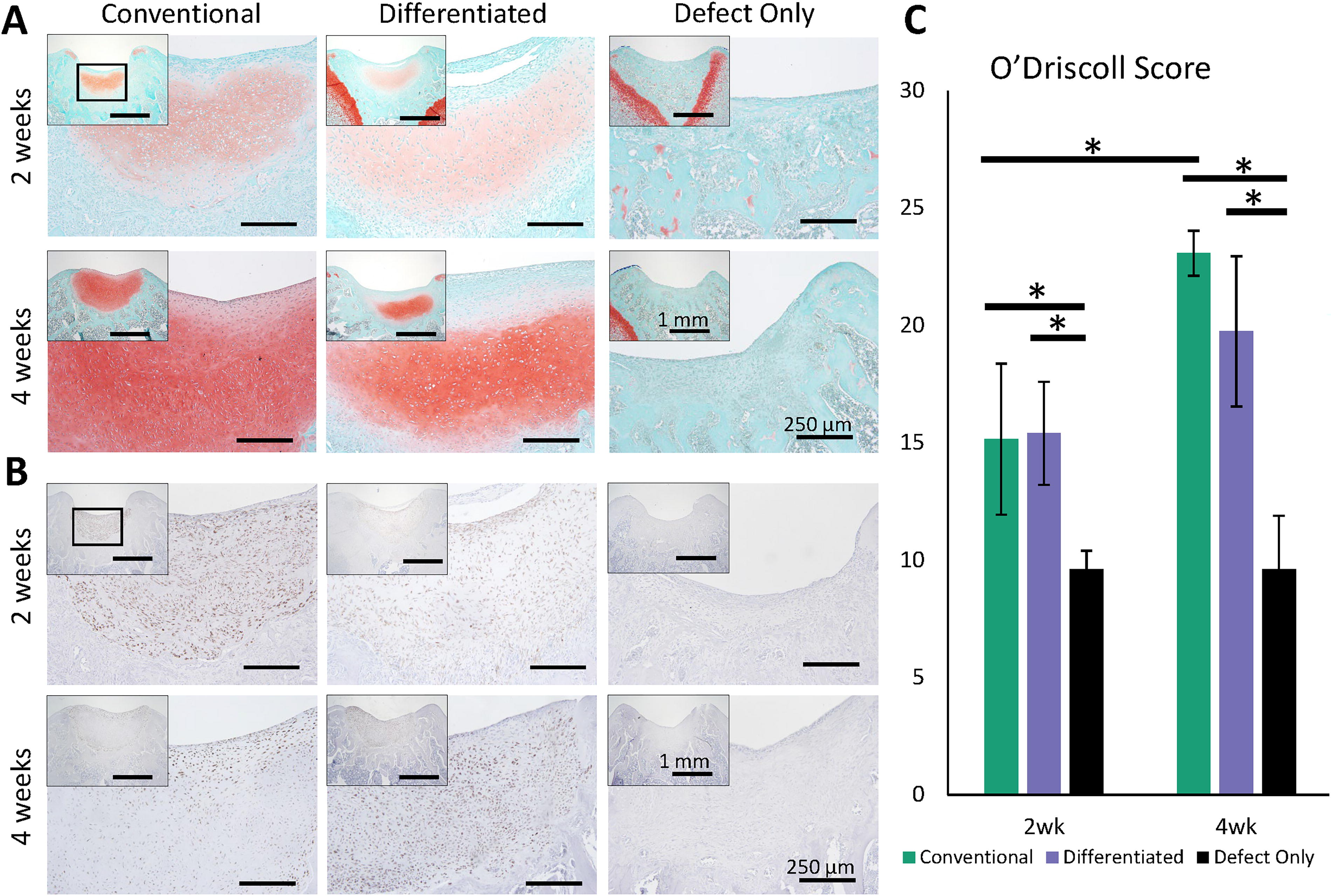
Histological analysis shows hyaline-like regenerated neocartilage of human origin.
hVim staining of sections (Fig. 4B) shows that regenerated neocartilage formation seen in safranin O staining of all transplant groups at all time points is of human origin. Nearly all cells within the neocartilage area stain for hVim, suggesting limited-to-no infiltration of rodent host cells into the regenerated area. Conversely, few cells were stained for hVim outside the sheet neocartilage tissue zone.
Modified OD scoring of cartilage regeneration (Fig. 4C) shows no significant difference between differentiated and conventional JCC sheet constructs at both 2- and 4-weeks posttransplantation (p > 0.05). In addition, transplant groups are superior to defect-only groups at all time points (differentiated vs. defect-only p = 0.005, 0.001, and conventional vs. defect-only p = 0.022, 0.001 at 2- and 4-weeks, respectively). Conventional JCC sheet-treated defects exhibit significantly improved healing at 4 weeks vs. 2 weeks (p = 0.004). Differentiated JCC sheet-treated rat defect-modified OD scores trended higher at 4 weeks than at 2 weeks (19.7 ± 3.2, 15.4 ± 2.2, respectively), but that difference was not statistically significant (p = 0.055).
Composition of the regenerated neocartilage matrix in defects from JCC sheets
IHC staining for characteristic articular cartilage matrix components, collagen type II and aggrecan, and absence of collagen type I confirm mature neocartilage composition in both JCC sheet transplant groups 4 weeks posttransplant (Fig. 5). Aggrecan was evenly expressed throughout neocartilage in all transplant groups and time points. At 2 weeks, collagen type II was only weakly expressed at the edges of the regenerated cartilage areas in undifferentiated JCC sheet-treated defects; whereas, differentiated sheet-treated defects exhibit strong, even staining.
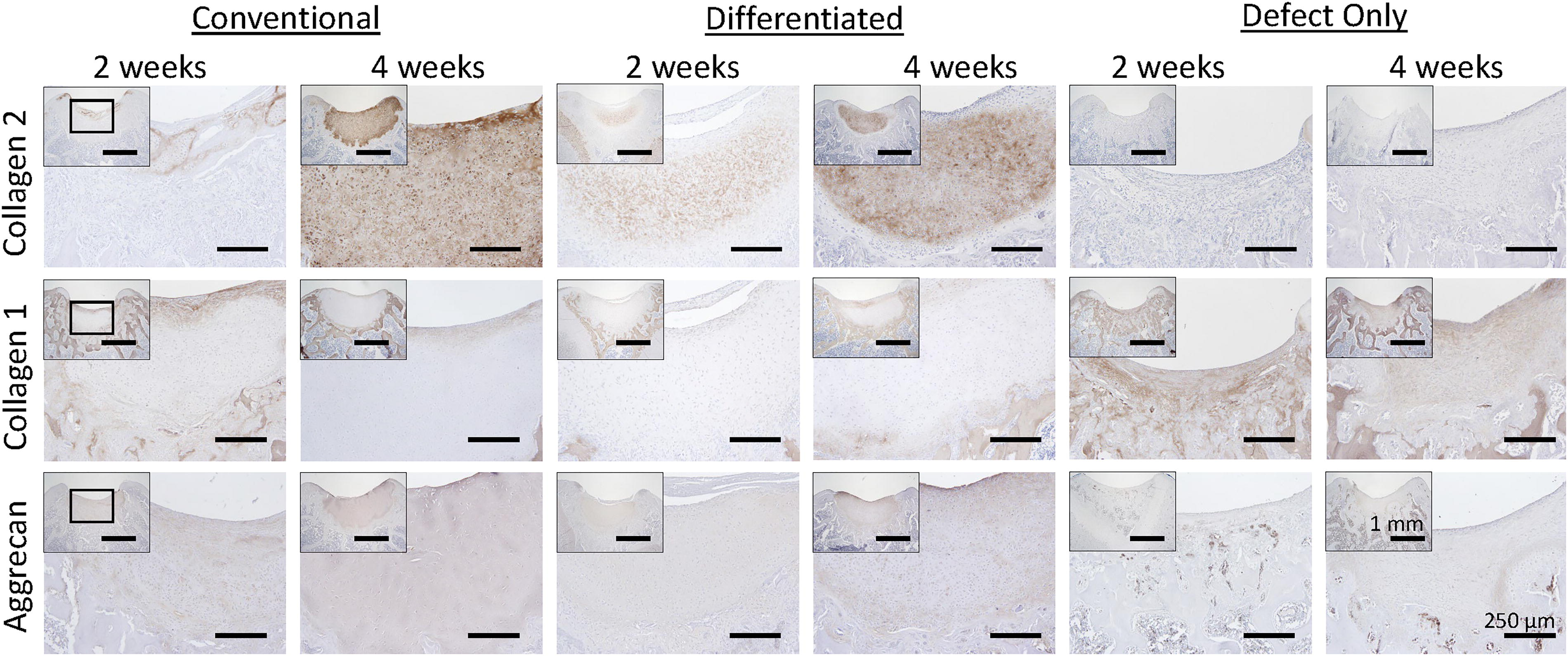
Regenerated neocartilage has a composition characteristic of hyaline cartilage. IHC for matrix proteins collagen 2 (top row), collagen 1 (middle row), and aggrecan in knees treated with undifferentiated JCC sheets (first two columns), differentiated JCC sheets (middle two columns), and no treatment (last two columns) after 2 weeks (left column for each treatment) and 4 weeks (right column). Low magnification defect site image insets in each composite image are shown at increased magnification in each image. Scale bar = 250 μm (1 mm for each inset). JCC, juvenile cartilage-derived chondrocytes.
Sheet integration with surrounding tissue and remodeling within subchondral bone
Chondrocytes throughout the neocartilage stain for PCNA at 2 weeks, but by 4 weeks, proliferating cells are limited to the peripheries of regenerated areas (Fig. 6). OCN is diffusely expressed in disrupted tissue deeper than in regenerated human-origin neocartilage (border denoted by dashed line, Fig. 6) present in the transplant groups at 2 weeks. By 4 weeks this tissue is fully replaced by subchondral bone surrounded by strong OCN staining. Within the regenerated neocartilage of JCC sheet-treated defects, chondrocytes are OCN-positive at 2 weeks; however, by 4 weeks, only deep chondrocytes stain for OCN (Fig. 6). In defect-only groups OCN is absent from the defect site and diffusely present adjacent to subchondral bone.
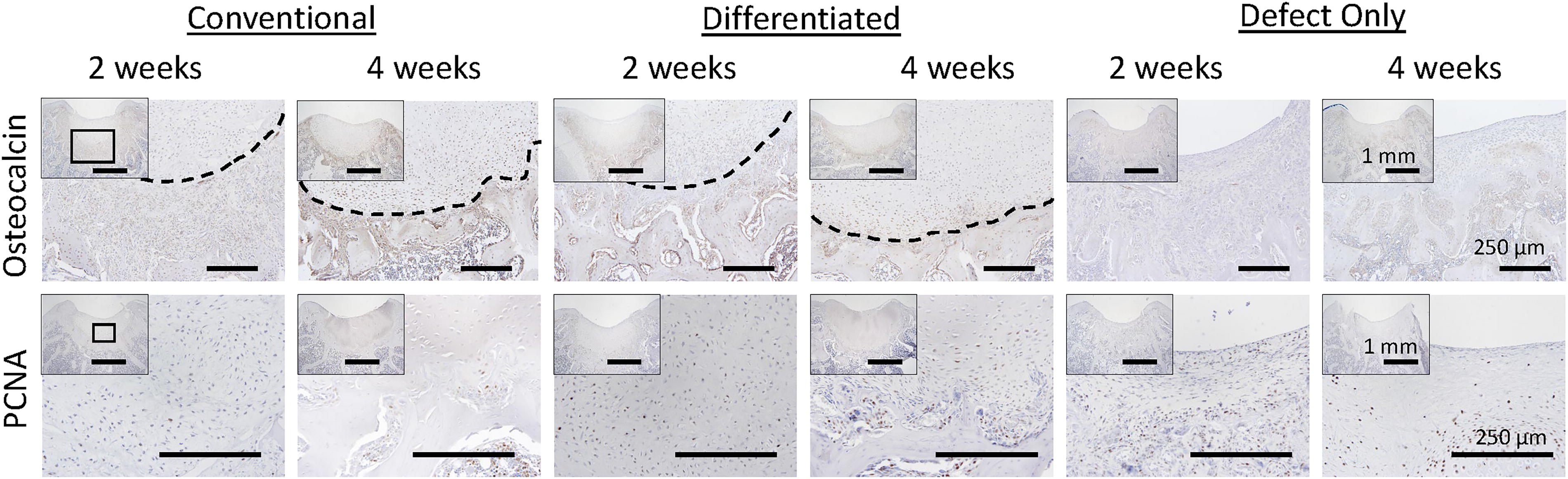
JCC transplants develop mature cartilage phenotypes by four weeks. IHC for osteocalcin (top row), dashed line denotes the edge of regenerated tissue, and proliferative marker, PCNA (bottom row) in knees treated with undifferentiated JCC sheets (first two columns), differentiated JCC sheets (middle two columns), and no treatment (last two columns) after 2 weeks (left column for each treatment) and 4 weeks (right column). Low magnification defect site image insets in each composite image are shown at increased magnification in each image. Scale bar = 250 μm (1 mm for each inset). PCNA, proliferating cell nuclear antigen; JCC, juvenile cartilage-derived chondrocytes.
Discussion
Human JCC sheets are a promising scaffold-free approach to cartilage regeneration, manifesting previous success in small animal models of cartilage defect healing including nude rats 26 and immunosuppressed rabbits. 21 In addition, a recent limited 10-patient pilot study of allogenic JCC sheet treatment with concomitant instrumented tibial osteotomy and microfracture manifested promising regeneration in individuals with OA.22 However, neocartilage formation in each study required substantial regeneration time (4 weeks or more). Long recovery times are attributed to the lack of hyaline-like phenotypes in JCC sheets at the time of transplant, as JCCs are 2D culture-adapted and undergo dedifferentiation common to all chondrocyte culture expansions.22,25,26,28,29 Human MSC sheets have been shown previously to robustly differentiate into hyaline-like cartilage in vitro in a similar fashion to conventional pellet cultures while maintaining surface proteins that enable adhesion to cartilage.12,37 Based on this evidence, we hypothesized that transplantation of JCC sheets exhibiting mature hyaline-like phenotypes established during in vitro differentiation would directly and more rapidly regenerate missing cartilage, accelerating cartilage healing over conventionally prepared JCC sheets. This study therefore compared healing of rodent focal chondral defects treated with differentiated human JCC sheets vs. conventional, that is de-differentiated, JCC sheets.
With media stimulus, JCC sheets differentiate more rapidly than reported for MSC sheet in vitro differentiation using the same chondrogenic media: 1-week producing cultured tissue thickness (345 ± 82 μm) similar to that reported at 3 weeks for differentiated MSC sheets (300–400 μm).12,31 In focal chondral defects, both conventional and differentiated JCC sheets adhere spontaneously to the subchondral bone without additional fixation (e.g., no sutures, fibrin glue, or other biomaterial support) while maintaining a readily transplantable, planar tissue geometry (Fig. 3A–D). hVim staining (Fig. 4B) confirms that JCC sheets persist within defect sites with high reliability (100% of conventional JCC sheets and 92% of differentiated JCC sheets). Preservation of ECM using thermo-responsive cultureware is important to cell sheet tissue site adhesion; other differentiated scaffold-free cell constructs with similar planar geometries require fibrin glue for in vivo tissue bed adhesion.32,38 Previous ex vivo work showed MSC sheet surface adhesive proteins (laminin, fibronectin) retained from thermal cell culture sheet release correlate to spontaneous sheet adhesion to tissue surfaces after in vitro chondrogenesis.12,18
Regardless of chondrogenic maturity, JCC sheets drive chondral defect repair observed across all sheet transplant groups at 2- and 4-weeks (Fig. 3G–J, M–O), but not in defect-only groups (Fig. 3K, L, Q, and R). At 4 weeks, defects are filled with regenerated neocartilage comprising sulfated proteoglycans (Fig. 4A), collagen II, and aggrecan, and without collagen I (Fig. 5). In addition, hVim staining (Fig. 4B) is consistent with a previous rodent sheet study, 26 supporting neocartilage formation from transplanted human cells and not host tissue stimulation and infiltration. Further, sheet-derived human cells are limited to defect area neocartilage, indicating that implanted cells do not migrate from defects and are less likely to present systemic toxicity risks or other possible ectopic adverse events.
Notably, accelerated healing predicted for differentiated JCC sheets is not supported histologically. While differentiated JCC sheet-treated defects are visually better filled with opaque, glassy new tissue (Fig. 3G–J) than conventional JCC sheet-filled defects at 2 weeks, histological OD scoring (Fig. 4C) supports healing equivalence between transplant groups at both 2- and 4-week timepoints. Differences in matrix components, namely collagen II, not considered by OD scoring, may explain this disparity. Clear collagen II differences are observed at 2-week timepoints. Physical barriers to host cytokines and chondrocyte quiescence within differentiated JCC sheets (Fig. 2L) may delay sheet responses to in vivo stimuli, equalizing conventional and differentiated JCC sheets in vivo despite differences in their in vitro maturity.
JCC sheet transplant integration with host tissue and continued maturation is evident: differentiated JCC sheets change notably upon implantation to produce sheet integration. Differentiated JCC sheets change from nonproliferative to proliferative within 2 weeks in vivo (compare Figs. 2L and 6), filling defects before reverting to nonproliferative mature cartilage at 4 weeks. Normal cartilage morphology relies on the initiation and cessation of chondrocyte proliferation and subsequent maturation, processes controlled in vivo by mechanical stimuli. 39 Thus, JCC sheet transplants proliferation and phenotype information (i.e, both visual, Fig. 3M–P, and histological, Fig. 4B) showing smooth regenerated neocartilage surfaces without outgrowth into surrounding tissue, support successful transplant integration into host joints. OD scoring (Fig. 4C) documents sheet transplant maturation in vivo: conventional JCC sheet treatment scores significantly increase between 2- and 4-weeks and those for differentiated JCC sheet sites trend toward increased healing. Furthermore, differentiated JCC sheets, rich in collagen I at the time of transplant (Fig. 2J), completely eliminate collagen I within neocartilage (Fig. 5B) within 2 weeks.
OCN expression within neocartilage follows PCNA proliferative dynamics: initially absent in cell sheets prior to transplant, evenly expressed within all chondrocytes in neocartilage at 2 weeks, and finally limited to chondrocytes adjacent to subchondral bone (Fig. 6). Typically an osteoblast marker, OCN is also expressed in hypertrophic chondrocytes and chondrocytes during late-stage OA. 40 Given stable JCC sheet transplant neocartilage formation reported to 28 weeks in rats, 26 and over 1 year in humans, 22 OCN-positive chondrocytes in bone-adjacent neocartilage likely do not represent the onset of hypertrophy and transplant calcification. OCN observed in subchondral spaces supports bone reformation, transitioning from diffuse within disorganized tissue at 2 weeks, to highly localized signal adjacent to newly reformed subchondral bone at 4 weeks (see Fig. 6) consistent with producing a stable osteochondral interface critical to the health and stability of articular cartilage.
Cell sheet treatment is observed to disrupt subchondral bone at 2 weeks before restoration at 4 weeks; defect-only groups did not yield deep subchondral bone disruption (Fig. 4A). This results in supraphysiological regenerated neocartilage thickness in both transplant groups (∼1 mm) at 4 weeks. All previous animal studies of human chondrocyte sheet transplants at similar time points demonstrated similar increased neocartilage thickness and encroachment into subchondral bone.21,26,27,41 A clinical JCC sheet study also reported thicker-than-normal regenerated condylar cartilage (3.5 ± 0.6 mm) at 1 year compared with normal 1.5–2.5 mm thickness,22,42,43 despite not being designed to assert such comparisons. Other scaffold-free techniques result in similar implant encroachment into subchondral bone.44–47 The impact of supraphysiological neocartilage thickness on joint function appears minimal, as animal studies demonstrate a return to equal weight bearing,21,26 and significant improvement in knee injury and OA outcome score (KOOS) and lysholm knee score in the human study. 22
OCN expression observed at 2 weeks (Fig. 6) suggests that defect creation initiates an early inflammatory response similar to that of OA, but that JCC sheets resolve this inflammation. JCCs are reported to exert potent immunomodulatory and chondrogenic paracrine and contact-dependent signaling.25,27,48,49 Tissue inhibitors of metalloproteinase 1 (TIMP1), TGF-βs, 25 and indoleamine-2,3-dioxygenase (IDO)48,49 have been identified in vitro as key factors based on their known activity directly inhibiting further ECM degradation,50–52 and promoting regenerative phenotypes of innate leukocytes (e.g., macrophages and NK cells).48,53–55 Though nude rat models lack T-cells, TGF-βs and IDO also exert potent anti-inflammatory and tolerogenic effects on T-cells and are involved in immune tolerance observed in human allogeneic transplants in synovial compartments of articulating joints.48,53–55
Donor-derived variations affecting JCC sheets’ overall regenerative efficacy are reported.22,26,27,30,33 JCC donors used both here and in a pilot study of early-stage JCC sheet-mediated neocartilage regeneration dynamics 30 produced equivalent results in a nude rat chondral defect model at 4 weeks. 26 Thus, donor-donor variations in cartilage regeneration observed at 2 weeks were unexpected. 30 Early-stage JCC sheet-mediated healing assessment across donors is therefore important to determine how JCC sheet in vitro differentiation may be relevant to certain JCC donor subsets, for example, accelerating healing from delayed neocartilage formation, or augmenting otherwise poor overall regenerative efficacy. However, as differentiated JCC sheet preparation increases culture time and uses large amounts of costly recombinant cytokines, JCC sheet cultured differentiation benefits in cartilage regeneration and healing should be balanced against increased costs over conventional sheets.
In summary, differentiated human JCC sheets bearing mature hyaline-like cartilage hallmarks adhere, integrate, and fill rodent cartilage defects with new tissue, continuing to mature as cartilage when transplanted into nude rat cartilage defects. While our hypothesized accelerated defect healing over conventional JCC sheet treatment was not observed, differentiated JCC sheet treatment produces healing and neocartilage regeneration equivalent to that of conventional JCC sheet treatment.
Footnotes
Authors’ Contributions
N.F.M., D.W.G., M.K., and T.O.: Conceptualized the study. N.F.M. and M.K.: designed, performed, and interpreted in vitro experiments and analysis. K.M.: Performed the preclinical animal study. Histological analyses were done by N.F.M. and A.J.F. using methods validated by M.K., K.M., and N.F.M.: Wrote the draft article. M.K., A.J.F., D.W.G., and T.O.: Reviewed and edited the article. D.W.G. and T.O.: Supervised the technical teams and coordinated resources. All authors approve of the final article content.
Disclosure Statement
T.O. is a shareholder of CellSeed, Inc. and is an inventor/developer designated on the patent for CellSeed’s commercialized thermo-responsive cultureware. All other authors declare that they have no competing interests.
Funding Information
This research was supported in part by a University Technology Acceleration Grant (UTAG, Utah Science, Technology, and Research-USTAR program, State of Utah, USA) to T.O. and D.W.G., and
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.