
Abstract
Peripheral nerve injuries (PNI) can result in significant losses of motor and sensory function. Although peripheral nerves have an innate capacity for regeneration, restoration of function after severe injury remains suboptimal. The gold standard for peripheral nerve regeneration (PNR) is autologous nerve transplantation, but this method is limited by the generation of an additional surgical site, donor-site morbidity, and neuroma formation at the site of harvest. Although targeted drug compounds have the potential to influence axonal growth, there are no drugs currently approved to treat PNI. Therefore, we propose to repurpose commonly used nonsteroidal anti-inflammatory drugs (NSAIDs) to enhance PNR, facilitating easier clinical translation. Additionally, calcium signaling plays a crucial role in neuronal connectivity and regeneration, but how specific drugs modulate this process remains unclear. We developed an in vitro hollow channel collagen gel platform that successfully supports neuronal network formation. This study evaluated the effects of commonly used NSAIDs, namely ibuprofen and indomethacin, in our in vitro model of axonal growth, regeneration, and calcium signaling as potential treatments for PNI. Our results demonstrate enhanced axonal growth and regrowth with both ibuprofen and indomethacin, suggesting a positive influence on PNR. Further, these drugs showed enhanced calcium signaling dynamics, which we posit is a crucial aspect for nerve repair. Taken together, these findings highlight the potential of ibuprofen and indomethacin to be used as treatment options for PNI, given their dual capability to promote axonal growth and enhance calcium signaling.
Impact Statement
Although targeted drug compounds have the potential to influence axonal growth, there are no drugs currently approved to treat peripheral nerve injury. We developed an in vitro hollow channel collagen gel platform that successfully supports neuronal network formation. This study evaluated the effects of commonly used nonsteroidal anti-inflammatory drugs, namely ibuprofen and indomethacin, in our in vitro model of axonal growth, regeneration, and calcium signaling as potential treatments for PNI. Our results demonstrate enhanced axonal growth and regrowth with both ibuprofen and indomethacin. Further, these drugs showed enhanced calcium signaling dynamics, which we posit is a crucial aspect for nerve repair.
Introduction
Damage to the peripheral nervous system (PNS) impacts 20 million individuals yearly in the United States, leads to severe sensory and motor deficits, and reduces quality of life. 1 The current standard of care for peripheral nerve repair is autologous nerve transplantation,2,3 which has limitations including donor site morbidity, matching difficulties due to different tissue size and shape, generation of an additional injury site, and neuroma formation at the site of injury. 4 Even after transplantation, treatments often yield poor functional recovery, especially when the nerve gap is large (>5 cm). To address these concerns, tissue engineering aims to develop innovative strategies for repairing and regenerating damaged peripheral nerves. 5 This shift underscores a proactive approach in seeking advanced treatments to enhance recovery and well-being for those affected by peripheral nerve injuries (PNI). A major barrier to improving peripheral nerve regeneration (PNR) treatments is our limited understanding of the complex biological processes involved in nerve regeneration. Developing experimental platforms to study these processes at the cellular level could be leveraged to identify targets to enhance regeneration after PNI. Here, we developed an in vitro platform to screen potential PNR therapies that can explore relevant form and function of regenerative growth important for translation to the clinic.
Appropriately targeted drug compounds have the potential to maintain neuronal viability, influence axonal growth over larger gaps, and manage neuropathic pain.6,7 However, no drug treatments are currently administered after PNI to improve regeneration, despite increased understanding of the specific molecular and cellular events that occur following injury. Although some medications are prescribed to manage symptoms after PNI, such as pain reduction or reduction of inflammation, none are specifically given to directly improve regeneration of the damaged nerve. 8 Recent data indicate that only 8%–14% of drugs explored for potential application in specific neurodegenerative diseases manage to successfully navigate through human clinical trials. 9 Hence, there is an unmet clinical need for pharmacological therapies to improve outcomes.8,10
Neuronal function, including calcium signaling, plays an important role in establishing and improving neuronal connectivity during regeneration.11,12 Recent research has suggested that understanding how specific drugs modulate calcium signaling will help in developing targeted treatments for PNR. 13 Goldberg et al. investigated whether intermittent depolarization with elevated concentrations of potassium ions (K+) enhances axon outgrowth in response to brain-derived neurotrophic factor, and determined that 40 mM potassium chloride (KCl) stimulation significantly enhanced axonal growth. 14 It has also been suggested that elevated calcium concentrations near the growth cone direct axonal growth,15,16 which, in total, suggest that while calcium flux may be an important metric for nerve regeneration, it remains relatively unexplored, particularly in response to putative drug treatments.
The present study utilizes our 3D in vitro platform model, which mimics native nerve architecture in an optically transparent 3D system, to identify potential drugs that can enhance axonal regeneration after injury and also identify neurotoxins to avoid such drugs for PNR.17,18 Given the high cost and risk of failure in drug discovery, we focus on drug repurposing, specifically examining FDA-approved nonsteroidal anti-inflammatory drugs (NSAIDs), which are relatively safe, readily available, and have been shown to support axonal regrowth in vivo. 19 In this study, we first benchmarked the suitability of our platform as a drug screening model by testing ibuprofen, which has previously been shown to promote axonal growth, 20 as a positive control and suramin, known for its neurotoxic effects, 21 as a negative control. Both drugs were evaluated leveraging our clinically relevant laceration injury model. Literature indicates that another NSAID, indomethacin, enhances recovery from spinal cord injury,22,23 prompting us to identify the role of indomethacin in axonal growth and regeneration in peripheral nerves using our model platform. Finally, we determine the impacts of treatments on calcium signaling and demonstrate that ibuprofen and indomethacin stimulate elevated calcium levels, suggesting a potential mechanism to enhance PNR.
Materials and Methods
Fabrication of hollow channel collagen gels for 3D study
Hollow channel collagen gels were prepared using our previously established method.17,18 Briefly, a custom mold (1 cm × 0.75 cm) was created using black resin (Formlabs, Somerville, MA) in a Form3 SLA 3D printer (Formlabs). Polydimethylsiloxane (PDMS; Sylgard 184; Dow Corning, Midland, MI) was prepared by combining the base and curing agent in a 10:1 (w/w) ratio. The mixture was then degassed for at least 1 h, poured into the mold, and further degassed for 30 min. The PDMS was cured at 60°C for a minimum of 8 h, then carefully peeled from the mold along its inclined walls, creating two wells. Following autoclave sterilization, sterile 21G blunt-end needles (McMaster-Carr, Robbinsville, NJ) were attached to both sides of each 1 cm × 0.75 cm well in aseptic conditions.
A 25G blunt end needle (McMaster-Carr) was inserted through each pair of 21G needles. To form the main structure of the 3D tissue constructs, a 6.5 mg/mL (final concentration) collagen hydrogel was prepared on ice by mixing type I rat tail collagen (Corning, Corning, NY) with 10X phosphate buffered saline (PBS; Fisher Scientific, Hampton, NH), growth medium (medium) (Dulbecco's Modified Eagle Medium [DMEM]/F-12 (Gibco, Gaithersburg, MD) medium supplemented with 10% fetal bovine serum (Gibco) and 1% anti-anti (Gibco), and sodium hydroxide (NaOH; Sigma-Aldrich, St. Louis, MO)] according to the manufacturer’s guidelines. The mixed collagen solution was pipetted into PDMS wells, covering the 25G needles (∼400 µL) to create 2-mm high gels (Fig. 1A, A’). Following 1 h of gelation at 37°C, the 25G needles were extracted to form hollow channels. The resulting hydrogel constructs were then removed from the PDMS frames (Fig. 1B, B’) and transferred into medium-filled 24 well plates.
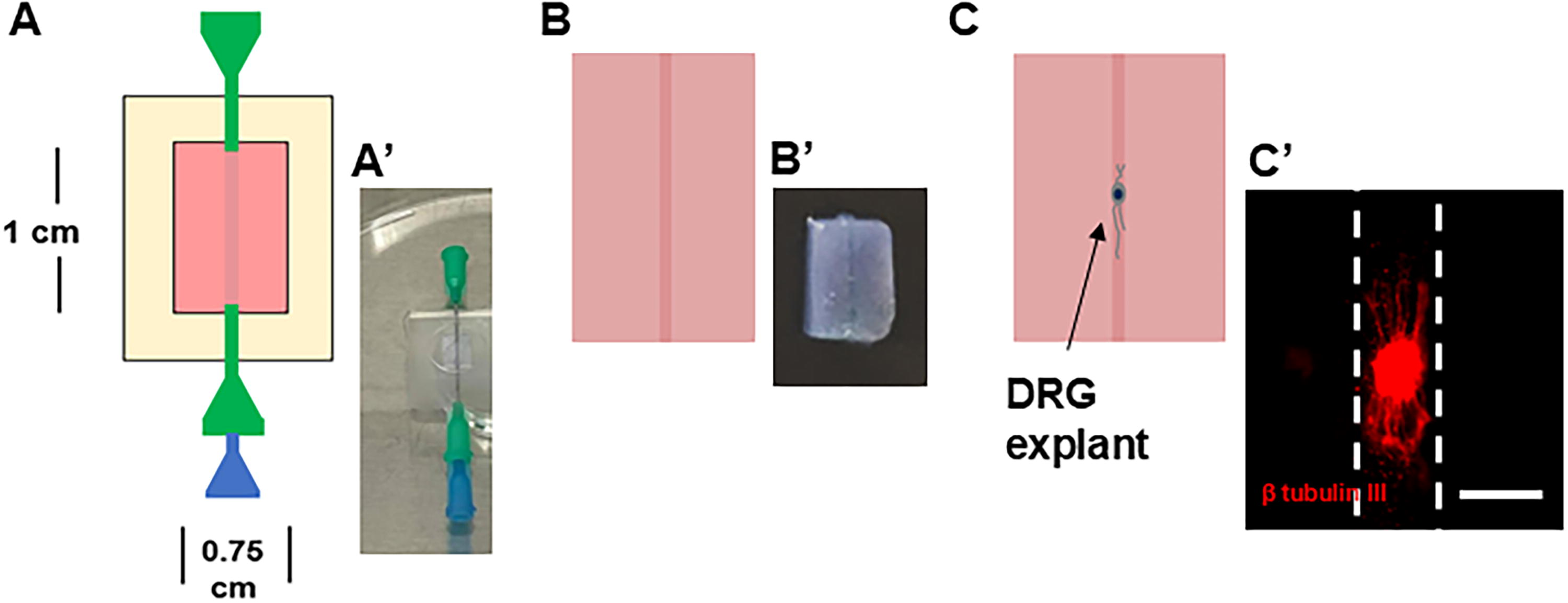
Overview of fabrication of hollow channels within 3D collagen gels.
DRG isolation and culture
All animal protocols were approved by the Rutgers-Newark Institutional Animal Care and Use Committees, adhering to institutional and Animal Research: Reporting of In Vivo Experiments guidelines. Dorsal root ganglia (DRGs) were isolated from E16 rat embryos following American Veterinary Medical Association-approved euthanasia as previously described. 18 Briefly, under aseptic conditions, the spinal cord was exposed, and lumbar DRGs were extracted using fine-pointed forceps. DRGs were transferred to a petri dish containing DMEM/F-12 supplemented with 1% anti-anti, pooling the tissue from the entire litter, trimmed to remove any surrounding fascia, and cut in half with a microscalpel. For 2D studies, DRGs were seeded on top of gels using micropipettes, while for 3D studies, they were injected into hollow channels using a 1 mL syringe (Becton Dickinson Medical, Franklin Lake, NJ) with a 25G needle (Fig. 1C, C’). Each collagen gel in 2D studies was seeded with up to three DRG explants on its surface. To study axonal growth in 3D, 2–4 DRGs were injected into hollow channels and, for the laceration model, 5–6 DRGs were injected into the central region of the channels and cultured for 5 days to allow for axon formation.
Development of injury model
We performed laceration-based injuries following our established protocol.17,18 Briefly, DRGs were grown inside hollow channels for 5 days after seeding to establish neuronal networks prior to inducing a laceration injury. A laceration injury was created by cutting the hydrogels between DRG explant bodies using surgical scissors, avoiding damage to the explants or dislodging them from the channels. An identical hollow collagen channel was created, cut, and carefully aligned with the injured construct to ensure the two channels were lined up before fusing them with several drops of 3.5 mg/mL collagen gel. Fused constructs were cultured for 2 weeks.
Drug testing
To observe the effect of drug compounds on axonal length, DRG-seeded constructs were incubated with medium supplemented with 0 or 100 µM ibuprofen (Sigma-Aldrich), reconstituted in 1X PBS; 0 or 200 µM suramin (Sigma-Aldrich), reconstituted in deionized water; or 0 or 100 µM indomethacin (Fisher Scientific), reconstituted in dimethyl sulfoxide. Drug concentrations were selected according to optimized concentrations found in the literature.20,21 After 5 days of culture, the constructs were fixed, immunostained, and both maximum and average axonal lengths were measured using ImageJ. Fused constructs in the injury model were incubated with medium supplemented with the same drug concentrations for 2 weeks.
Immunostaining
Constructs were fixed with 4% paraformaldehyde (Boston BioProducts, Ashland, MA) in PBS, rinsed with PBS, and then permeabilized using 0.1% (v/v) Triton X-100 in PBS for 20 min for 2D studies and 45 min for 3D studies. Samples were rinsed with PBS + 0.05% (v/v) Tween-20 (PBST) 2 times for 5 min each for 2D studies and 3 times for 10 mins each for 3D studies and blocked with 5% (w/v) bovine serum albumin (BSA)/PBST for 30 min for 2D studies and 90 min for 3D studies. The samples were then immediately incubated for 1 h for 2D studies and overnight for 3D studies at room temperature with a primary antibody against β-tubulin III (Sigma-Aldrich; 1:500 in 1% BSA/PBST). Following an additional round of PBST washes, samples were incubated 30 min for 2D studies and overnight for 3D studies with an appropriately species-matched Alexafluor 594 secondary antibody (Fisher Scientific; 1:500) and DAPI (1:1000) in 1% BSA/PBST. Finally, samples were rinsed three times for 2D studies and five times for 3D studies with 1X PBS before imaging.
Imaging and DRG axon length measurements
Fluorescent imaging was performed using a Keyence BZ-X810 inverted microscope (Keyence, Elmwood Park, NJ) and associated software using a 4X or 10X objective. The microscope’s “full focus” feature was used to scan through the constructs’ thickness and capture in-focus pixels throughout. Image brightness and contrast were adjusted uniformly across all images throughout the study. All the axons from each explant body were measured to obtain their lengths using ImageJ (NIH, Bethesda, MD). These lengths were averaged to obtain the average axonal length and identify the maximum axonal length from each explant body to obtain maximum axonal lengths from each explant body. Observers were blinded to treatment groups during both imaging and analysis. The diameter of each DRG was measured, and any that were smaller than 150 µm or lacked a well-defined body were excluded from further analysis. No size differences were observed among DRGs.
Functional analysis of DRG neurons
To assess neuronal function in the presence of our drugs of interest, we performed calcium imaging using Fluo-4 AM (Fisher Scientific) to measure changes in intracellular calcium concentration using time-lapse images, as previously described.24,25 Briefly, DRGs were seeded on top of collagen gels in glass-bottom 35 mm dishes (Fisher Scientific) and cultured for 5 days in growth medium supplemented with a mitotic inhibitor (20 µM 5-fluorodeoxyuridine and uridine, Sigma-Aldrich) containing either 100 µM ibuprofen, 100 µM indomethacin, vehicle, or controls with no drugs. Growth medium was then replaced with recording medium [DMEM/F-12, HEPES, no phenol red (Gibco)] supplemented with 3.2 μM of both F-127 (Fisher Scientific) and Fluo 4-AM. After 30 min of incubation, cells were washed with fresh recording medium. Live imaging was acquired for 20 min using a Nikon eclipse Ti microscope (Nikon, Melville, NY) equipped with a Hamamatsu camera (Hamamatsu Photonics, Bridgewater, NJ). The first 10 min were recorded without any stimulation, after which 40 mM KCl was added, and another 10 min were recorded with the stimulator present in the medium. On average, 30 regions of interest (ROI) per DRG were selected using ImageJ (Fig. 2A). The fluorescence intensity of each ROI was measured, and the average background intensity was subtracted from each cytoplasmic measurement; these subsequent values were normalized to the first measurement recorded for each ROI. The ratio of measured fluorescence (F) to baseline fluorescence (F0) was plotted against time for each ROI to facilitate the visualization of calcium flux both before and after adding the external stimulus, KCl (Fig. 2B). From the plot and associated measurements, fluorescence intensity and spike frequency were calculated. Spike frequency was calculated by quantifying the number of spikes per minute for each ROI.
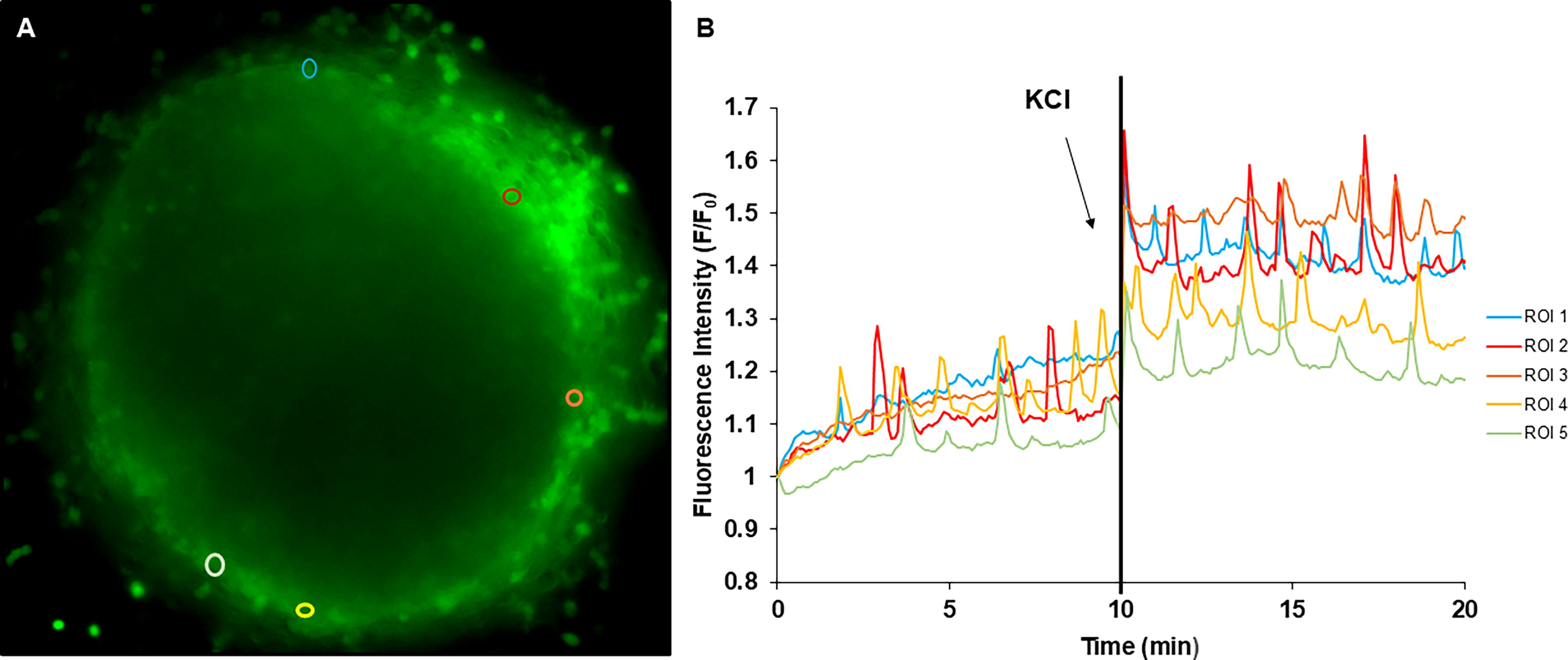
Statistics
Data are presented as mean ± standard deviation. All experiments were run in triplicate and from multiple litters to strengthen the scientific rigor of the studies, and sample sizes are reported as the total number of DRGs analyzed. Statistical analyses were conducted using IBM SPSS Statistics (IBM, Chicago, IL). One-way analysis of variance (ANOVA) with Tukey’s post hoc analysis was the primary statistical method, while the Student’s t-test was employed where specified. Statistical significance was defined as p < 0.05 for all comparisons between experimental conditions.
Results
Investigation of potential drug candidates to support axonal growth
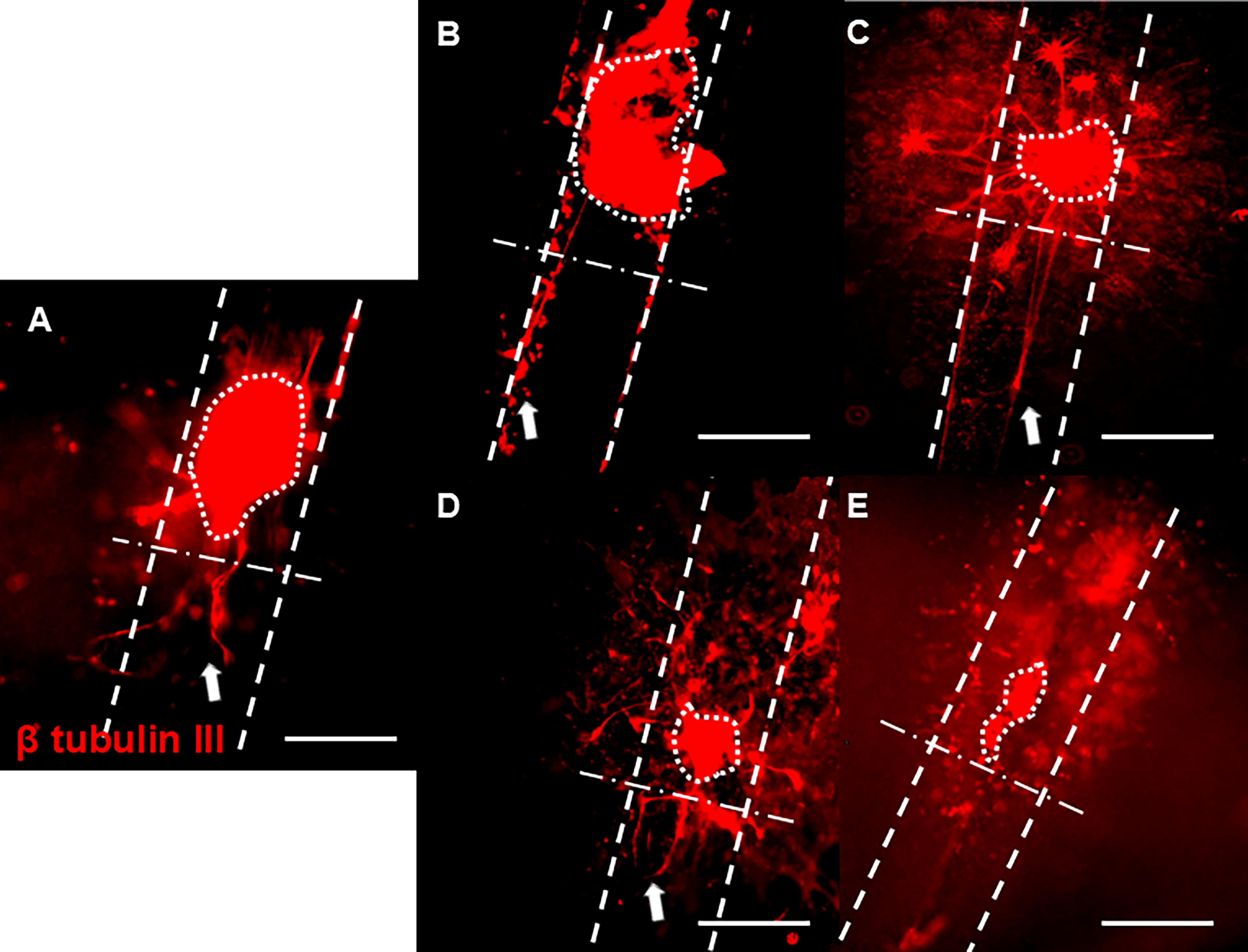
To evaluate whether our 3D hollow channel platform could identify potential drugs to treat PNI based on their ability to enhance axonal growth, DRGs were seeded inside hollow channels and cultured with either standard growth medium (control), vehicle, 100 µM ibuprofen, or 200 µM suramin for five days. Ibuprofen-supplemented cultures supported robust axonal growth compared to control groups (Fig. 3A–C). Notably, DRGs incubated with ibuprofen supported an increased number of longer axons in comparison to those incubated with no ibuprofen or vehicle. To obtain quantitative results, the average and maximum length of every axon was measured, and cultures supplemented with ibuprofen supported significantly longer average and maximum axonal lengths with respect to controls (Fig. 3D and E). Ibuprofen not only enhanced axonal length but also increased the number of axons per explant, with the average number of axons per explant increasing from 11 in the control and vehicle groups to 19 when cultured with ibuprofen. In contrast, incubation with suramin supported considerably fewer and shorter axons compared to controls (growth medium and vehicle) (Fig. 4A–C). Quantitative analysis indicated a significant reduction in axonal length with suramin-treated explants compared to controls (Fig. 4D and E). Taken together, these data reproduce results found in the literature showing that ibuprofen enhances axonal growth while suramin suppresses it in vitro. Therefore, we expect that we can identify potential drugs for PNR based on their ability to enhance axonal growth using our platform system.
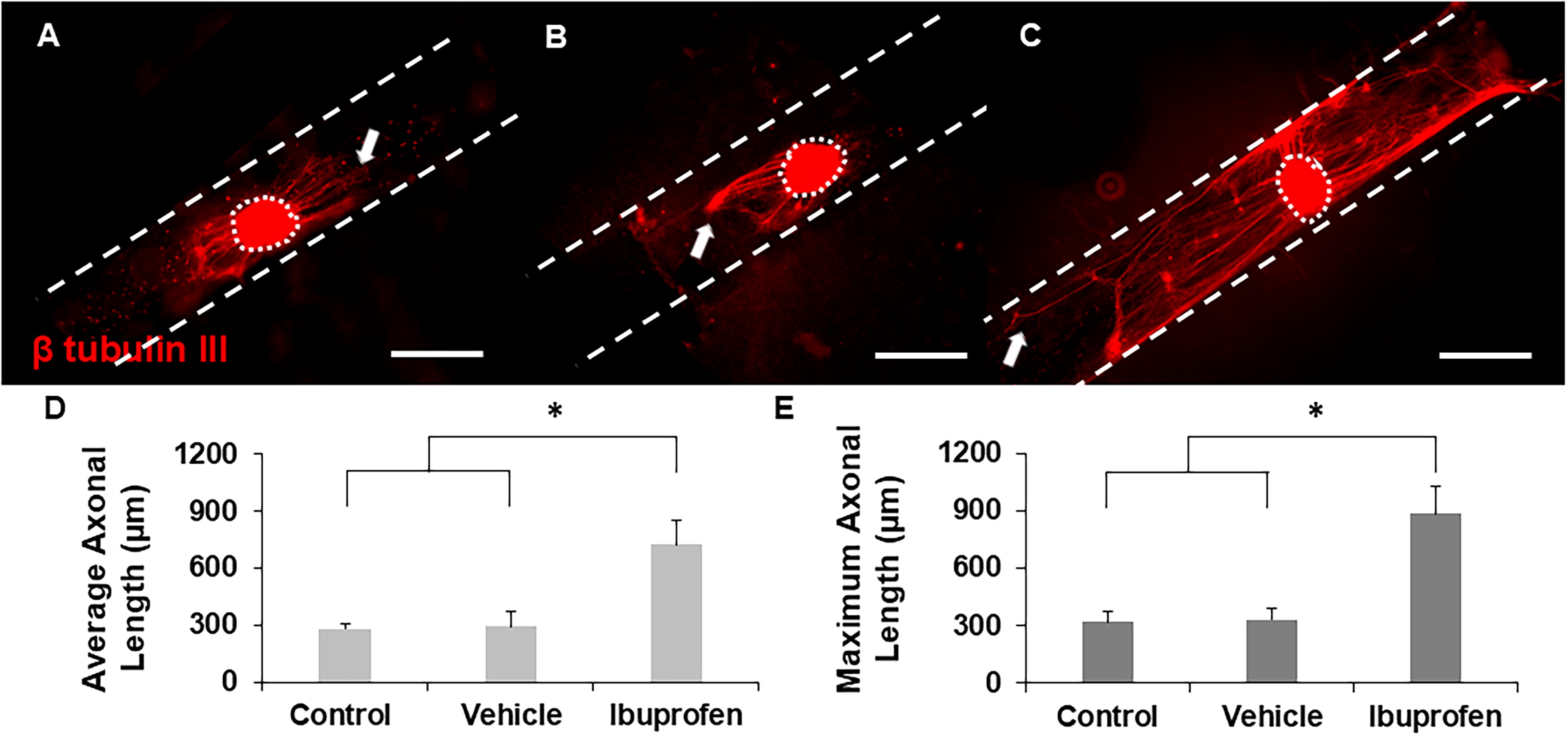
Representative images of axonal growth from DRGs cultured inside hollow channels with
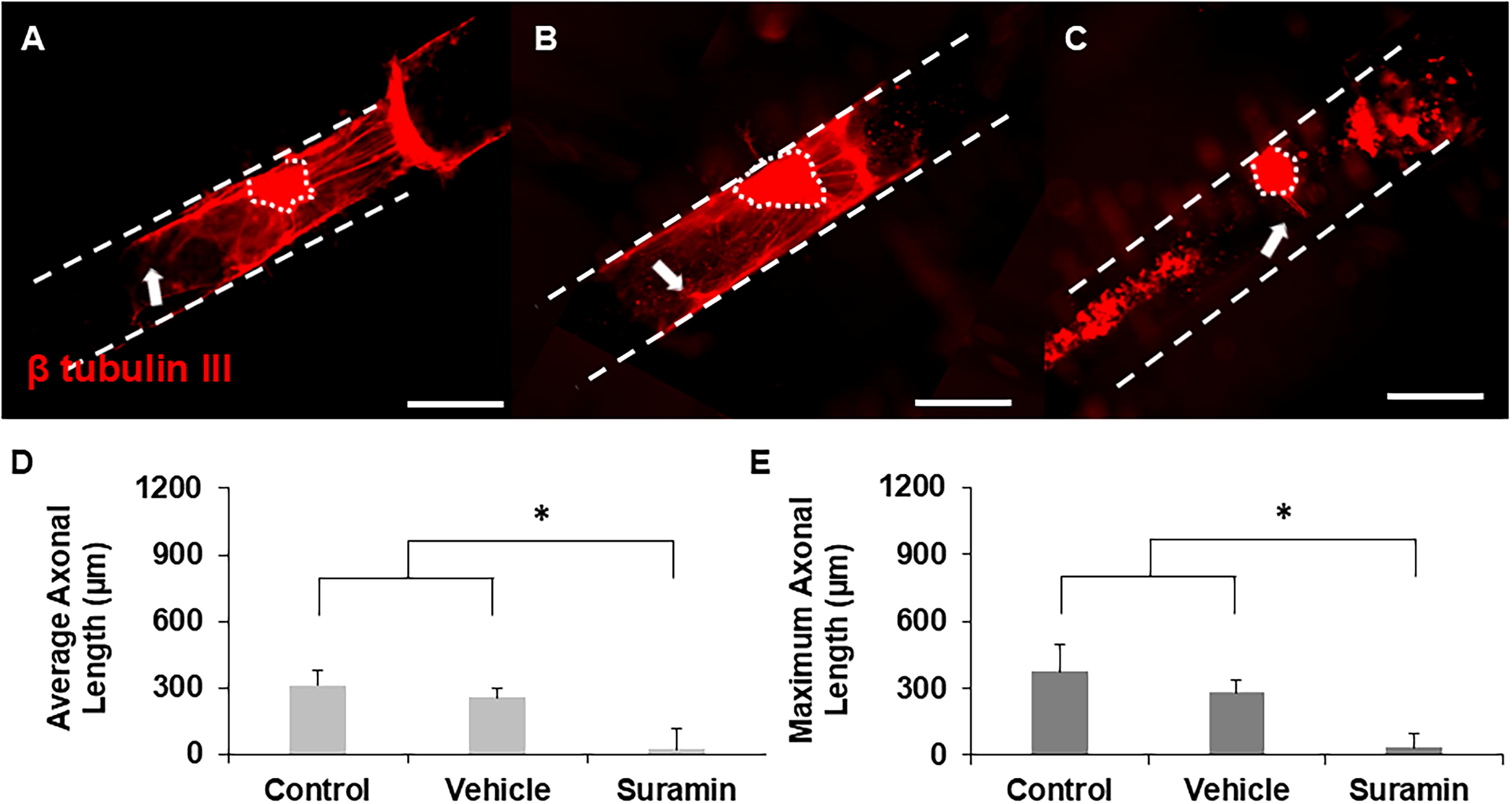
Representative images of axonal growth from DRGs cultured inside hollow channels with
Investigation of potential drug candidates to support axonal regeneration
To identify potential drugs for PNR based on the ability of the drug to enhance axonal regrowth after injury, we employed our laceration model. 17 Constructs were cultured in growth medium prior to injury to generate axons throughout the channel and after creating the injury, lacerated constructs were cultured in medium supplemented with growth medium, vehicle, 100 µM ibuprofen, or 200 µM suramin, and imaged to quantify axonal regrowth across the lesion site after 2 weeks of culture postinjury. Ibuprofen-treated constructs supported robust regeneration (Fig. 5C), while suramin-treated constructs supported no axonal growth (Fig. 5E). Control and vehicle treatment groups supported modest regeneration (Fig. 5A and B, D). Quantitative analyses measured average and maximum axon lengths from each explant into the new channel. Ibuprofen significantly enhanced average and maximum axonal regeneration (Fig. 6A and B), whereas suramin completely suppressed it (Fig. 6C and D). These results demonstrate that our laceration model can identify drugs based on their ability to enhance axonal regeneration and that ibuprofen enhances axonal regeneration after PNI.
Representative images of axonal growth from DRGs cultured inside hollow channels with
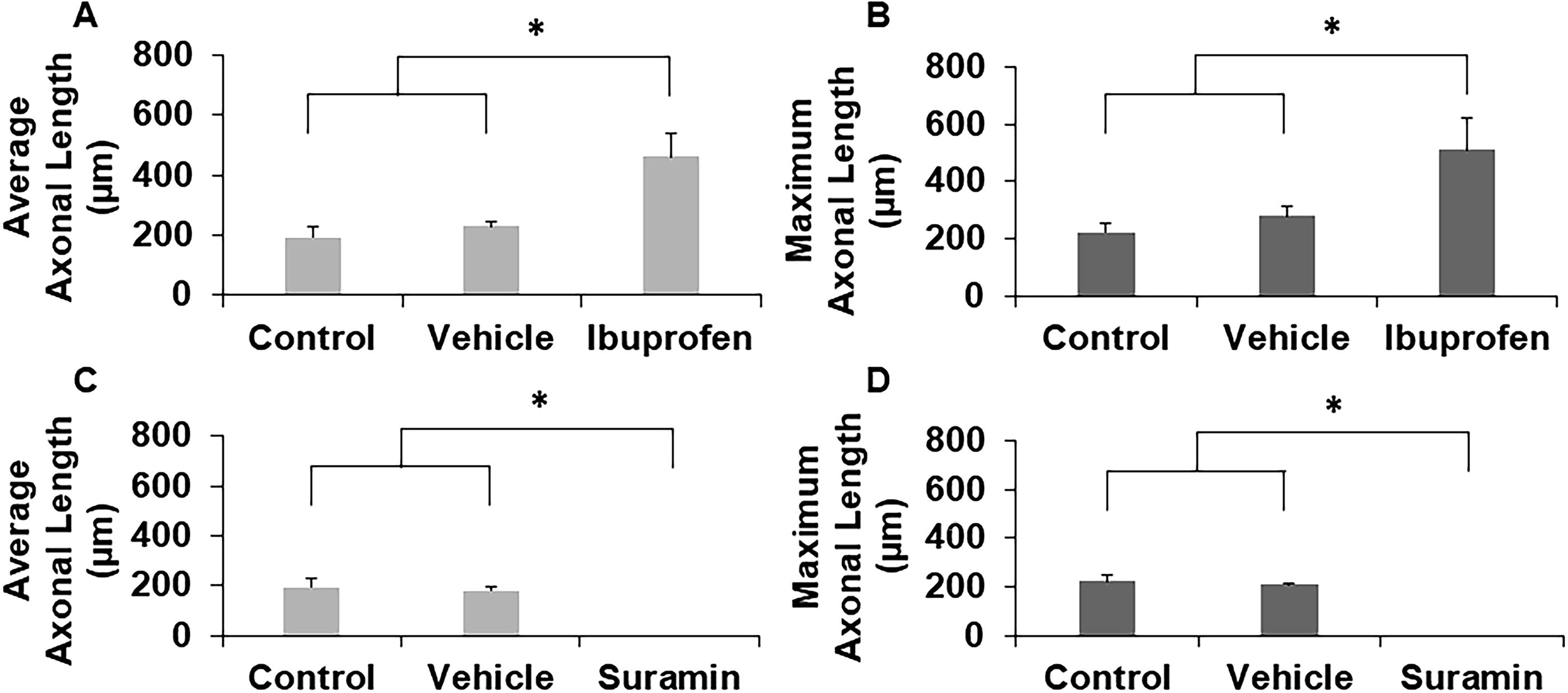
Quantification of
Optimization of the concentration of indomethacin to induce axonal growth
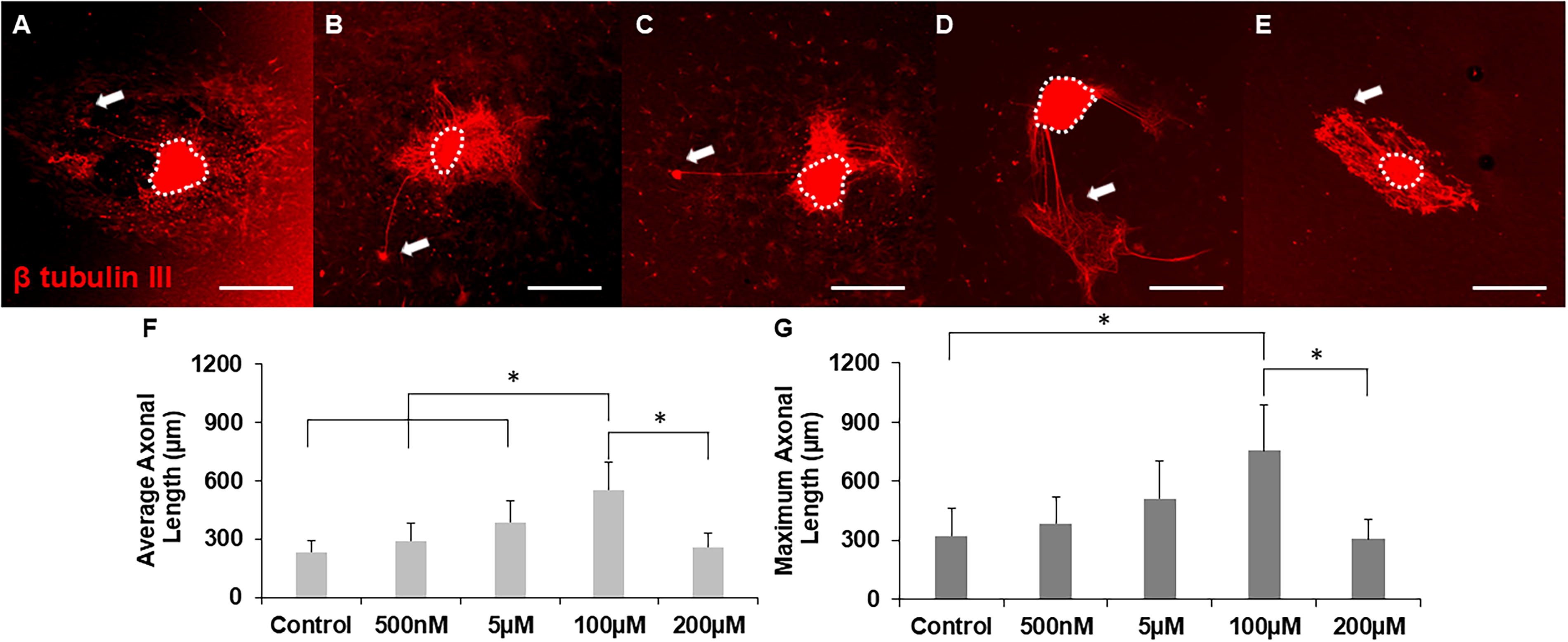
Using our model system, we tested another NSAID, indomethacin, which shares functional similarities with ibuprofen and has shown promising results in treating spinal cord injury.22,26 To optimize the indomethacin concentration to induce maximal axonal growth, DRGs were cultured on 2D collagen gels with increasing concentrations of indomethacin, and the average and maximum axonal lengths were calculated using ImageJ after 5 days of culture (Fig. 7). Visually, 500 nM and 5 µM of indomethacin supported numerous shorter axons, with several longer axons present. Supplementation with 100 µM indomethacin supported numerous axons extending from the explant that were uniformly longer than other culture conditions, whereas 200 µM supported axonal morphologies similar to those from lower concentrations of indomethacin (Fig. 7A–E). Quantitative analyses support these observations: DRGs cultured with 100 µM of indomethacin promoted significantly longer average axonal growth compared to control and all other doses of indomethacin (Fig. 7F). This concentration also significantly enhanced maximum axonal growth with respect to control and 200 µM of indomethacin (Fig. 7G). As such, 100 μM of indomethacin was able to robustly enhance axonal growth and this concentration will be utilized in our 3D model.
Representative images of axonal growth from DRGs cultured inside hollow channels with
Indomethacin significantly enhances axonal growth and regeneration
To determine the effect of indomethacin on axonal growth, DRGs were seeded inside hollow channels with indomethacin-supplemented medium. In these experiments, standard cell culture medium (without any drugs) served as the negative control, which was evaluated against medium supplemented with the optimized concentration of indomethacin. Representative images revealed significantly longer axons extending from DRGs incubated with indomethacin compared to no indomethacin (control) and vehicle (Fig. 8A–C). Moreover, similar to ibuprofen, indomethacin supported qualitatively more axons per explant than control and vehicle DRGs. Quantitative results demonstrate that indomethacin significantly enhanced both average and maximum axonal length with respect to controls (Fig. 8D and E).
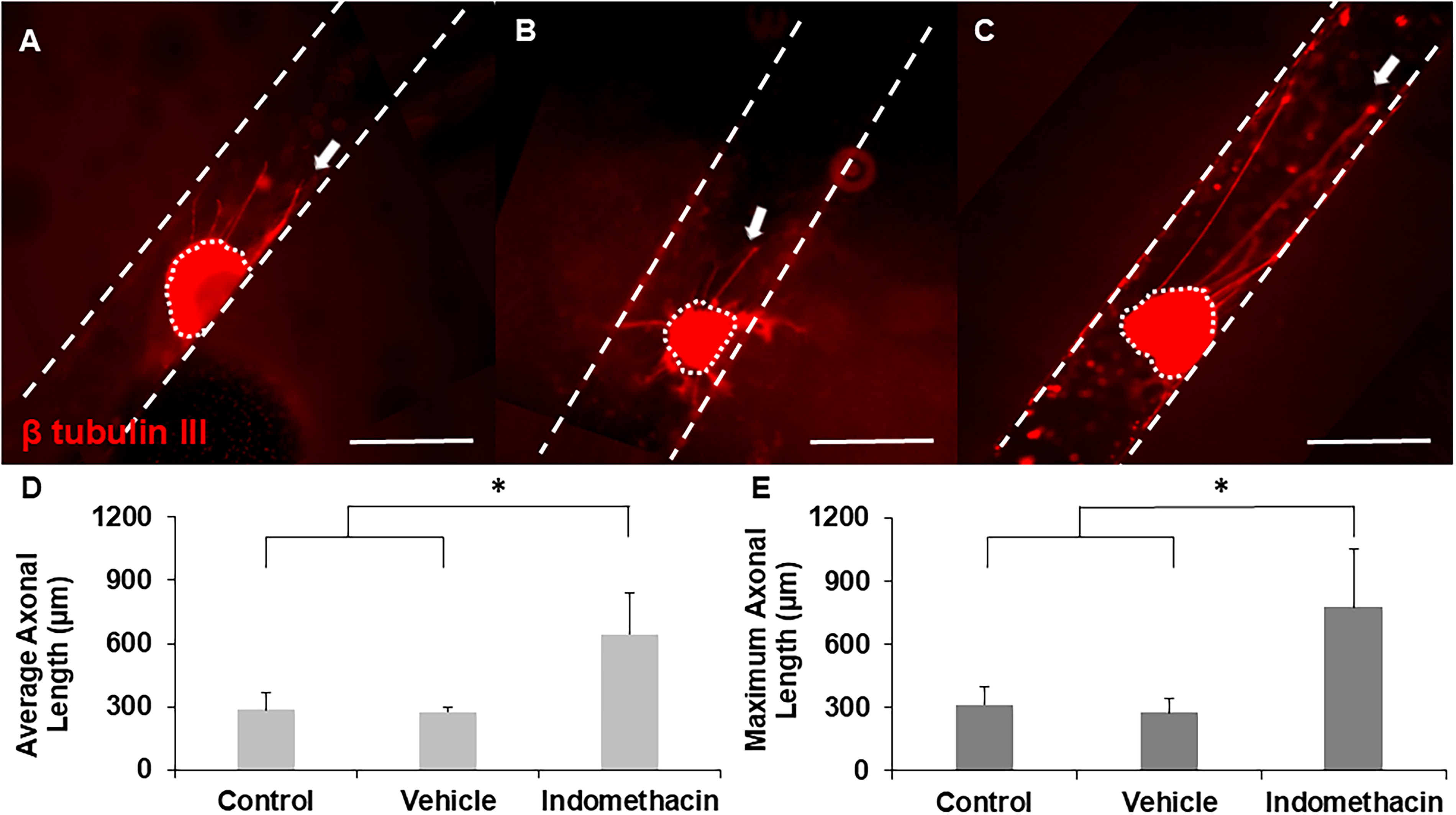
Representative images of axonal growth from DRGs cultured inside hollow channels with
To examine the effect of indomethacin on regenerating axons after injury, lacerated constructs were cultured with indomethacin for 2 weeks after injury. Representative images revealed axonal regrowth from DRGs into the new channel for all groups, and in particular, indomethacin-supplemented constructs supported longer axons across the lesion site (Fig. 9A–C). Quantitative analysis of both the average and maximum length of axons demonstrated that indomethacin supplementation supports significantly longer average and maximum axonal regrowth compared to controls (Fig. 9D and E). Hence, our results indicate that indomethacin can potentially enhance axonal regeneration after PNI.
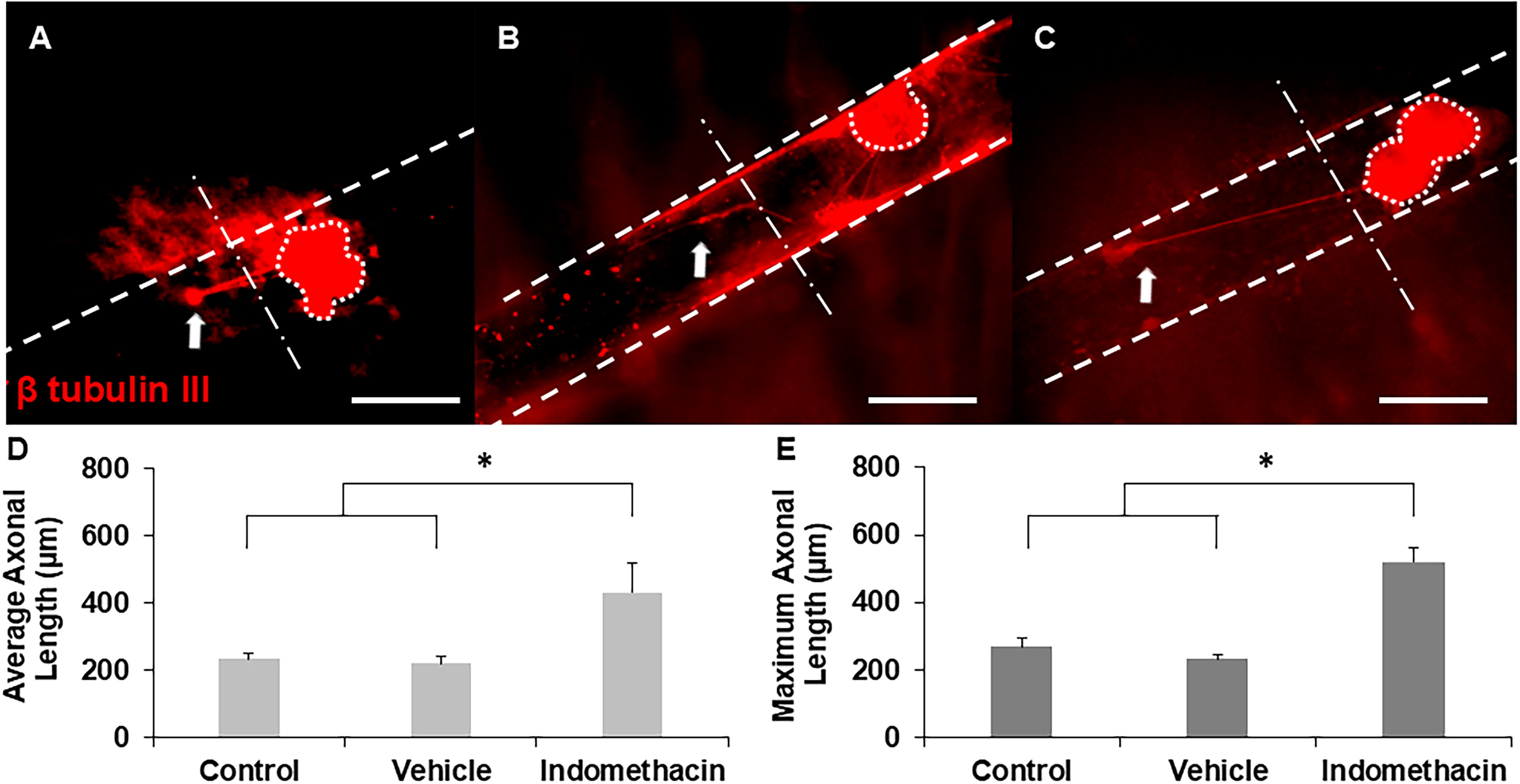
Representative images of axonal growth from DRGs cultured inside hollow channels with
Ibuprofen and indomethacin enhance calcium signaling
To identify the role of select NSAIDs on calcium signaling, we cultured DRGs in the presence of 100 μM ibuprofen or 100 μM indomethacin for five days. Fluo-4 AM was used to measure changes in intracellular calcium levels. To study calcium dynamics, cultures were recorded for 20 min and 40 mM KCl was added to the culture at the 10-min mark to depolarize the cells and activate voltage gated channels (Fig. 10A). Changes in fluorescence intensity and spike frequency were calculated for each explant.
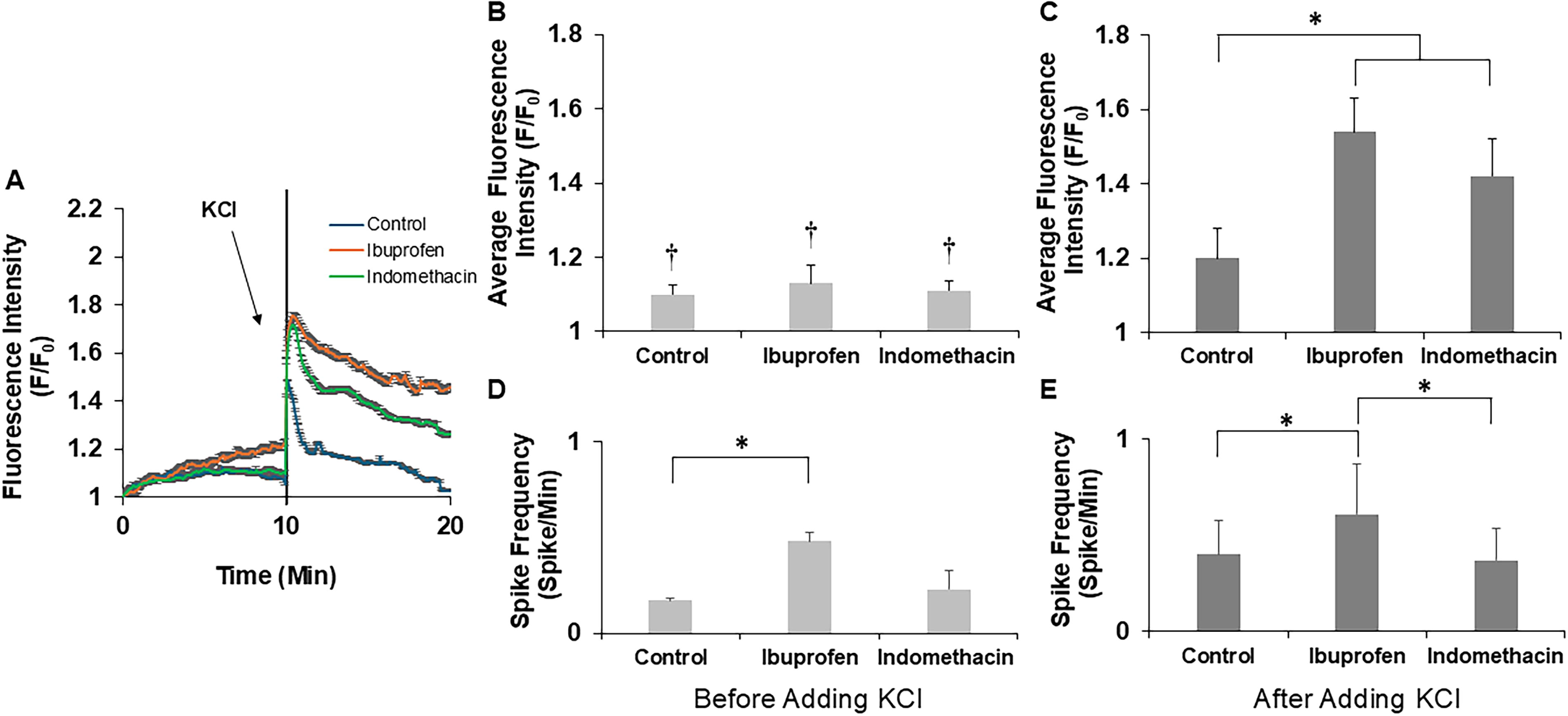
In all experimental conditions, we observed fluctuations in intracellular calcium concentrations, confirming the viability and activity of these cells in culture. Distinct patterns of changes in intracellular calcium were observed, with some cells exhibiting spontaneous oscillatory changes in intracellular calcium concentration (oscillatory cells),12,27 while others displayed a synchronized rise only following external stimulus application (40 mM KCl) (evoked cells). Qualitatively, there was a higher prevalence in the number of oscillatory cells in cultures supplemented with ibuprofen compared to indomethacin and control cultures, and indomethacin treatment also displayed qualitatively more oscillatory cells than the control. For comparative analysis and visualization of differences in calcium signaling, average fluorescence intensity was plotted against time for all conditions (Fig. 10A). Although measurements were similar prior to stimulation, there was a clear increase in average fluorescence intensity (F/F0) after the addition of KCl to the medium. Quantitative analysis revealed significant differences in average fluorescence intensity within each individual condition before and after KCl stimulation. Notably, the average baseline fluorescence intensity before stimulation was similar across all conditions (Fig. 10B). After stimulation with KCl, all analyzed fields of DRGs treated with ibuprofen or indomethacin contained cells with significantly higher average fluorescence intensities than control cultures (Fig. 10C). Spike frequency was calculated from oscillatory cells as these were the only cells to show spontaneous activity; fields of cells analyzed from cultures treated with ibuprofen supported significantly higher spike frequency both before and after addition of KCl (Fig. 10Dand E). These results suggest that ibuprofen and indomethacin not only enhance axonal growth and regeneration but also enhance calcium signaling.
Discussion
In this study, our goal was to characterize the effect of two common NSAIDs, ibuprofen and indomethacin, on axonal growth, regeneration, and calcium signaling to ultimately develop novel treatments for PNI. For that, first, we demonstrate the utility of our 3D hollow channel collagen gel model as a platform for identifying potential drugs for treating PNI by initially evaluating drugs with known effects, selecting ibuprofen and suramin as positive and negative controls, respectively. We found that ibuprofen significantly enhanced axonal growth and regeneration postinjury, while suramin inhibited axonal growth and regeneration in our in vitro platform. These results align with findings reported in existing literature,20,21 demonstrating the reliability and utility of our model for drug screening in PNI treatment. We next evaluated indomethacin, a different NSAID with structural similarities to ibuprofen, which has been shown to facilitate recovery after spinal cord injury.22,23,28 We determined that indomethacin also enhances axonal growth and regeneration with effectiveness similar to ibuprofen. Importantly, we evaluated neuronal function using calcium signaling, which to our knowledge has not yet been done in the context of a regenerative model. We observed that both ibuprofen and indomethacin, which showed enhanced axonal growth, also enhanced calcium signaling by elevating intracellular calcium concentrations. This is significant as calcium signaling plays a crucial role in neuronal connectivity and nerve regeneration.11,12,14 Thus, we identified the influence of two commonly used NSAIDs, ibuprofen and indomethacin, on key factors to address PNI: axonal growth, regrowth, and calcium signaling.
Our findings highlight the potential of repurposing NSAIDs, specifically ibuprofen and indomethacin, as regenerative treatments for PNI. The high costs and risks associated with new drug discovery make repurposing FDA-approved drugs an attractive alternative. NSAIDs are widely used as pain relievers and in particular, ibuprofen is commonly provided to reduce inflammation. Our study confirms that ibuprofen significantly enhanced axonal growth and regeneration after injury, which aligns with previous literature demonstrating its capacity to increase average neurite length in vitro, 20 and promote tibial nerve repair in vivo. 19 Ibuprofen has also been demonstrated to overcome the inhibitory role of chondroitin sulfate proteoglycan on axonal growth by activating peroxisome proliferator-activated receptor gamma (PPARγ), which ultimately suppressed RhoA and thereby stabilized the growth cone. 29 Indomethacin has been reported to activate PPARγ in other cell types such as adipocytes, 30 suggesting its potential for working in the same pathway as ibuprofen, and it has been shown to promote recovery after spinal cord injury.22,26 This instigated our exploration of the effects of indomethacin in the peripheral nervous system and our data indicate that indomethacin also enhanced both axonal growth and regeneration following injury with similar efficacy as ibuprofen.
Our study provides compelling evidence that ibuprofen and indomethacin not only promote axonal growth but also significantly affect calcium signaling in DRG neurons. Although positive results in terms of axonal growth are promising, assessing the drug’s effect on neuronal function through techniques such as calcium imaging adds depth to the understanding of its biological impact. Calcium imaging also enables a comprehensive evaluation of drug efficacy and safety as it can reveals drugs that lead to pathological calcium signaling, which can also be detrimental.31,32 Thus, we performed calcium imaging to measure changes in intracellular calcium concentration. 33 Suramin, initially used as a negative control to validate our 3D platform to mimic peripheral nerve regeneration, was excluded from these analyses because of its inhibitory effects on axonal growth and neuronal viability. Thus, we focused on ibuprofen and indomethacin, both of which showed positive results in our 3D platform system. To our knowledge, the effects of these NSAIDs on calcium signaling in this context have not been evaluated, making our investigation particularly novel. In our study, the fluorescence intensity (F/F0) was initially similar across all conditions, indicating that the overall calcium uptake was comparable regardless of the presence of NSAIDs. Most interesting, ibuprofen not only increased the number of oscillatory cells, but also increased the spontaneous spike frequency of these neurons, potentially suggesting a different mechanism of action between ibuprofen and indomethacin with regards to calcium signaling. Upon KCl stimulation, all cells in the ibuprofen and indomethacin treatment groups exhibited a significantly sustained increase in fluorescence intensity. In oscillatory cells, spike frequency was also enhanced with ibuprofen incubation, indicating some level of intrinsic ability for ibuprofen to modulate intracellular activity. Enhanced fluorescence intensity and spike frequency indicate increased calcium signaling.12,34 Ca2+ develops and improves functional connections between neuronal cells,11,12 so we hypothesize that these alterations in calcium signaling may facilitate drug-mediated axon growth-promoting effects to facilitate PNR. Our observations align with previous findings that calcium signaling can also promote axonal outgrowth rate and direction,11,14,16,35,36 which are independent of pathological calcium overloading mechanisms,31,32 suggesting a positive role in elevated calcium signaling for growth and regeneration.
Neuronal activity and calcium signaling are crucial for axon growth and guidance during development, and these mechanisms likely play a similar role in nerve regeneration by promoting axon regrowth and reconnection with afferent targets, aiding in the re-establishment of neuronal connections.37,38 Intracellular delivery of Ca2+ from the endoplasmic reticulum, which typically occurs immediately after PNI, activates unfolded protein response signaling, facilitating the promotion of axonal regeneration. 39 Extracellular Ca2+ delivery from a nerve guidance conduit also accelerated axon extension and neuronal connectivity in PNR, illustrating the importance of calcium signaling in these processes. 11 Thus, our findings demonstrate that ibuprofen and indomethacin not only promote axonal growth and regeneration, but also have a notable impact on altering calcium signaling, particularly after external stimulation. These drugs may enhance the responsiveness of neurons to depolarizing stimuli such as KCl and the enhanced activity likely contributes to axonal growth-promoting effects, although this will need to be confirmed in future studies. Taken together, these results suggest the potential use of these drugs as treatment options for PNI.
Although NSAIDs such as ibuprofen are commonly taken orally for pain relief, this route of administration can lead to significant adverse effects such as gastrointestinal complications and kidney damage, particularly with long-term use.40,41 Thus, future studies should focus on optimizing targeted drug delivery methods to the injury site to enhance efficacy while minimizing off-target effects. A promising approach could be the use of specialized conduits capable of facilitating localized drug administration directly to the injury site. This strategy could potentially overcome the limitations associated with systemic delivery, offering an innovative and more precise alternative that would enhance PNR and improve patient outcomes.
Conclusions
This study highlights the potential of repurposing ibuprofen and indomethacin, NSAIDs commonly used to suppress inflammation and pain, for enhancing PNR using a facile platform system. Using our 3D in vitro hollow channel platform, we demonstrated that both ibuprofen and indomethacin significantly increased axonal growth and regeneration as well as enhanced calcium signaling after chemical stimulation. This heightened activity of calcium signaling likely contributes to promoting axonal growth and restoring neuronal connectivity observed in our tissue model. Understanding the dual impact of these drugs on calcium signaling and axonal growth sheds light on their potential therapeutic role in nerve regeneration to ultimately improve outcomes for individuals affected by PNI. Future research should concentrate on refining targeted drug administration techniques to maximize therapeutic impact while reducing off-target effects.
Footnotes
Acknowledgments
The authors thank Dr. Ying Li and Dr. Millie Swietek for their assistance with the collection of the calcium imaging recordings, and Dr. Bonnie Firestein for helpful discussions throughout the development of this article. The authors would also like to thank Karl Chelala and Nyssa Nixon for their assistance with gel making and immunostaining.
Authors’ Contributions
J.T.: Conceived the idea, performed experiments and data analysis, and wrote this article. B.J.P.: Assisted with the experimental planning and interpretation of calcium imaging data. J.M.G.: Supervised and directed the project. All authors edited the article.
Disclosure Statement
The authors declare no competing financial or nonfinancial interests.
Funding Information
This research was funded in part by the NIH (