
Abstract
Long bone and craniofacial bone fractures amount to an overwhelming expenditure for patients and health care systems each year. Overall, 5–10% of all bone fractures result in some form of delayed or nonunion fractures. Nonunions occur from insufficient mechanical stabilization or a compromised wound environment lacking in vasculature and progenitor cells. The current standard for treating these critical-sized fractures and defects is the use of autologous bone grafts. However, advancements in tissue engineering have cultivated a shift in scientific efforts toward harnessing the body’s own regenerative resources. As such, research on fracture healing has shifted as well. Transforming growth factor-beta 1 (TGFβ-1) has been studied in fracture healing for over 25 years, though many of these studies have been in vitro or in small animal models. The few studies in large animals have disagreement due to the heterogeneity within the experimental design. Because TGFβ-1 plays such a crucial role in the bone healing process, this systematic review investigates the application of TGFβ-1 in various carrier vehicles for repairing bone injuries in large animal and rabbit models. A systematic search was conducted in PubMed, Embase, and Web of Science (from database construction—October 2024). A total of 244 articles were screened, and 24 studies were included for review. Most large animal long bone studies used coated titanium implants, while most rabbit long bone studies used some form of degradable polymer constructs. TGFβ-1 doses in large animal long bone studies range from 0.005 to 750 µg, doses in large animal calvaria and mandible studies range from 1 to 5000 µg, and doses in rabbit long bone studies range from 0.05 to 120 µg. Nineteen out of 24 articles reviewed indicate successful use of TGFβ-1 for bone regeneration compared with experimental controls. It is clear that dose and controlled release of growth factor play a crucial role in defect closure, but outcome measures and success criteria were inconsistent across studies. More studies with consistent experimental designs are critical for understanding the therapeutic potential of TGFβ-1 in fracture repair, but overall, this review indicates that TGFβ-1 can be used alone or in conjunction with other growth factors to accelerate successful bone repair.
Impact Statement
Studies using TGFβ-1 to enhance successful long bone fracture and defect repair in large animals are limited, preventing therapeutic advancements from reaching clinical translation. Experimental design has also been heterogeneous, leading to contrasting results and a lack of insight into the potential benefits that TGFβ-1 may offer. This review summarizes much of the in vivo research utilizing TGFβ-1 in both large animal and rabbit long bone repair, as well as in large animal calvaria and mandible repair to capture the most up-to-date perspectives on its advantages and deficiencies. This review can serve as a guideline for future exploration and innovation.
Introduction
Bone injuries and healing
Normal healing of reduced fractures occurs through several stages.1,2 Immediately following a fracture, a hematoma forms that is infiltrated by immune cells which remove injured tissue and secrete factors capable of recruiting additional cells into the injury site. This yields granulation tissue in which progenitor cells proliferate and differentiate to form a cartilaginous tissue that mechanically stabilizes the site 5–10 days postfracture and supports healing. The callus undergoes mineralization 7–21 postfracture due to hypertrophy of chondrocytes (endochondral ossification) and deposition of woven bone by invading osteoblasts (intramembranous ossification). In the subsequent early remodeling phase, mineralized tissue is replaced with secondary bone (lamellar bone) through osteoclast–osteoblast coupling in the process of bone remodeling. In the late remodeling phase, the bone contour is restored, and the Haversian system is reestablished (Fig. 1).
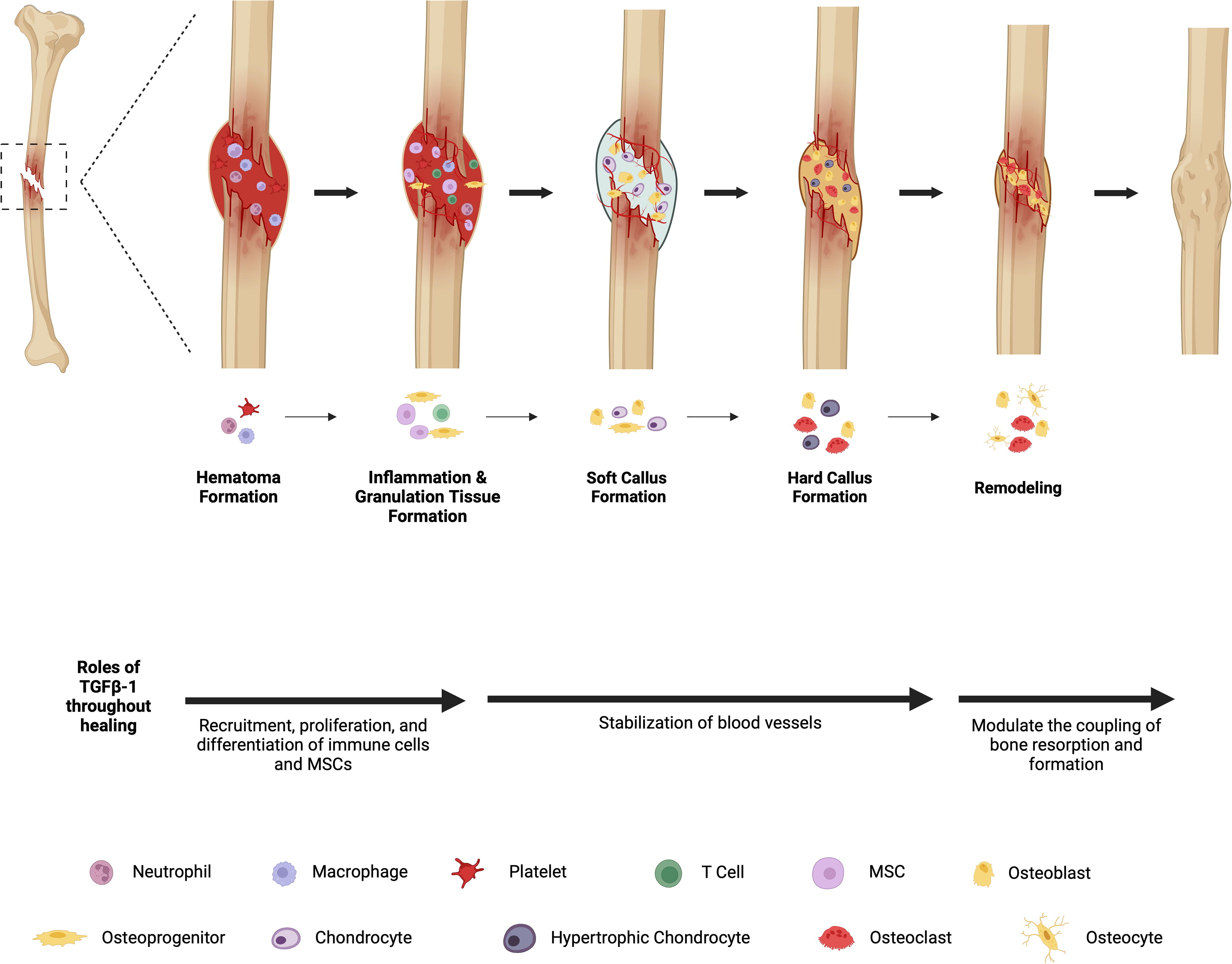
Fracture healing process with TGFβ-1’s involvement. A hematoma is formed, immune cells are recruited and granulation tissue is formed. Angiogenesis is stimulated and additional cells are recruited to the injury site, which forms a soft cartilaginous callus (primary callus). The cartilage then begins to calcify as chondrocytes undergo hypertrophy and osteoblasts deposit bone, forming a hard callus (woven bone). Calcified cartilage is removed, and bone remodeling takes place via coupled bone removal and formation via osteoblasts and osteoclast activity, resulting in lamellar bone formation with restored Haversian systems.3–5 TGFβ-1, transforming growth factor-beta 1. *Image created using BioRender.com. Created in BioRender. Trbojevic, S. (2024) https://BioRender.com/e69m620.
Most patients with bone fractures successfully undergo healing after reduction (cast or fixation implant) without further intervention (Fig. 1). However, improper fixation, infection, ischemia, and disease can lead to a delayed or nonunion fracture (DNF), sometimes requiring multiple revision surgeries. Notably, 5–10% of all bone fractures and 25% of long bone fractures result in some form of DNF. Treatment of DNFs costs a median of around $25,600 USD per fracture.4,6 The Food and Drug Administration states the diagnosis of a nonunion as “a minimum of nine months has elapsed since injury and fracture shows no visibly progressive signs of healing for three months.” 7 There are two forms of nonunion: (1) hypertrophic nonunion and (2) atrophic nonunion. Hypertrophic nonunions occur when sufficient biological factors are present, but mechanical stabilization is inadequate due to a disorganized callus structure outside of the fracture site. In contrast, atrophic nonunions occur due to a lack of regenerative potential from deficiencies in biological factors and blood supply, leading to inadequate callus formation.7,8
Treatments for DNFs include conservative therapies for delayed unions and less-severe nonunions or surgical interventions for more severe nonunions. Atrophic nonunions are typically more severe due to the lack of biological factors and the need for complex treatment, compared to the addition of mechanical stability in hypertrophic nonunions. The use of autologous bone grafts has been the primary method of atrophic nonunion treatment for decades. 7 Though these grafts typically have high success rates, donor site morbidity or insufficient graft size continue to complicate the healing process. 9 However, with modern advancements in the fields of tissue engineering and regenerative medicine, research has shifted focus to improving the inadequate biological environment by the use of stem cells, growth factors, and osteoconductive scaffolds. 7
Healing of critically sized defects is impaired compared to reduced fractures because of the relatively compromised wound environment, namely less progenitor cells and vascular supply for the large volume of tissue to form (Fig. 2). Without intervention, the injury typically results in nonunion with a fibrotic immune response and impaired osteogenesis. 16 Regenerative approaches generally recognize that large critically sized segmental defects do not fully bridge without some combination of osteogenic drugs, scaffolds, and progenitor cells, because a large volume of bone must be regenerated in a brief time window before fibrosis and nonunion ensues.16,17 Autogenous bone grafting remains the gold standard for large bone defects, with varied clinical techniques depending on the volume and available bony tissue, for example, Masquelet technique, plate-assisted bone segment transport, and free flaps transplant (Fig. 2). 18

Defect healing in long bones (top) versus calvaria (bottom). Healing of critical-sized segmental defects in long bones is compromised due to an inadequate reparative environment and must be supported by additional scaffolds, factors, or cells. Upon presentation (hematoma and inflammation), a bone spacer may be placed (Masquelet technique) during wound temporization (typically open wounds) to prevent fibrous tissue growth and to induce a biomembrane and vascularization. With soft tissue restored/uninjured, various implants are placed with autologous bone grafts the most common, leading to progenitor cell differentiation of chondrocytes and osteoblasts. Bone formation occurs via endochondral and intramembranous ossification depending on implant type, defect site, and vascular support. Over time, mineralized cartilage and woven bone are replaced with mature bone via remodeling.10–12 Calvarial segmental defect healing via intramembranous ossification. After initial hematoma formation and inflammatory response, condensation of mesenchymal stem cells enlarges and differentiates into osteoblasts. Vascular ingrowth to the defect occurs and osteoblasts deposit bone matrix. Finally, bone remodeling leads to mature bone containing osteocytes.13–15 *Image created using BioRender.com. Created in BioRender. Trbojevic, S. (2024) https://BioRender.com/u11n608.
Role of TGFβ-1 in bone healing
The TGFβ superfamily is a group of proteins that are involved in recruiting, proliferating, and differentiating multiple cell types. Bone morphogenic proteins (BMPs), part of the TGFβ superfamily, have been extensively researched due to their innate osteogenic potential; in fact, recombinant human BMP-2 and BMP-7 were approved by the FDA for specific clinical applications. However, the supraphysiological dose of commercial devices yields rapid bone formation of inferior mechanical integrity and often debilitating complications, including heterotopic ossification. Researchers continue optimizing BMP2/7 delivery to minimize dose while maintaining efficacy.8,19
TGFβ-1, a member of TGFβ superfamily, is expressed earlier in fracture healing and exhibits both osteogenic and immunomodulatory roles. TGFβ-1 is one of the most abundant cytokines in the matrix, and its expression, along with its receptor, has been shown to increase and remain elevated throughout the entire healing process. 20 TGFβ-1 in particular is involved in the proliferation of undifferentiated mesenchymal stem cells (MSCs), which are then promoted into chondrocytes and osteoblasts by BMPs (Fig. 1). Interestingly, its structure is phylogenetically conserved (unchanged in primate, bovine, avian, and porcine species), making interspecies research relatively straight-forward.21,22 TGFβ-1 has gained more attention due to its multifunctional roles and environment-based response in a variety of biological processes, making it a suitable candidate for tissue engineering studies. Thus, TGFβs have been considered promising biologics for fracture repair. 23
Animal models of bone healing
Small rodents have been popular in studying TGFβ-1 due to cost and genetic manipulation. However, ferrets, guinea pigs, rats, and mice have several limitations in the study of bone compared with larger animals, particularly regenerative approaches. The small size of tissue and defects do not model physiological impediments to healing, such as distance of biomolecule diffusion and progenitor cell migration. The bones also lack Haversian canals, which are conduits for osteoprogenitors and osteoclasts. While rabbits are also rodents, they do have Haversian systems similar to those of humans. However, functional mechanical loads are lower than large animals and thus, are reviewed separately.
Large animals (canines, sheep, goats, pigs, and nonhuman primates) better model the composition and biomechanical properties of human bones.24–26 Several models of bone healing exist in the literature. These can be classified as fractures, critically sized defects, and hardware osteointegration. Fractures may be created with mechanical overloads or surgical osteotomy, with gaps in the latter mimicking incomplete reduction. Critically sized defects are created by removing a large portion of bone, typically segmental osteotomes and cylindrically drilled holes. Segmental defects in long bones are critically sized when the defect length is 2× the bone width. Osteointegration tests the integration of bone to implanted hardware.24–26 Here, we review the limited number of studies using TGFβ-1 for bone healing in long bones of large animals and rabbits, as well as craniofacial bones of large animals to allow for a wider scope of information.
Methods
Search strategy
Literature reporting on the use of TGFβ-1 for healing fractures and segmental defects, as well as device integration in long bones, calvaria, and mandibles in large animal and rabbit models, were systematically searched in PubMed, Embase, and Web of Science (from database construction—October 2024). The following search terms were used: (transforming growth factor beta-1 OR TGFb-1) AND (regeneration OR repair) AND (mandible OR calvaria OR long bone OR femur OR tibia OR fibula OR radius OR ulna) AND (rabbit OR pig OR swine OR porcine OR dog OR canine OR sheep OR goat OR minipig). Reference lists of related articles were also screened. The protocol for this systematic review was registered on the PROSPERO database (registration number: CRD42024596528).
Eligibility criteria
Inclusion and exclusion criteria were determined using the population, intervention, comparison, and outcomes (PICO) framework. 27 Inclusion criteria were as follows: (1) original research studies; (2) used TGFβ-1 to repair bone fractures and defects in large animal models or rabbits in vivo, with no restriction of growth factor dose or combination therapy (growth factors, scaffold biomaterials carriers); (3) repaired fractures and defects, or integrated fixation device in long bones, mandibles (large animal only), or calvaria (large animal only); (4) conducted histological, radiological, and/or mechanical outcome analyses; and (5) included a study control group.
Exclusion criteria were as follows: (1) studies were not original research, (2) evaluated using in vitro methods, (3) did not use large animal models or rabbits (e.g., mice, rats), (4) involved human participants/specimen, (5) involved compromised wounds (trauma, infection), (6) did not include a control group, (7) not in English, and (8) full text not accessible.
Data extraction and analysis
Titles and abstracts from the literature search results were assessed by two independent reviewers (S.T. and J.T.) and disagreements were resolved by a third reviewer (A.A.). Studies deemed eligible for review were chosen based on our inclusion and exclusion criteria. If the abstract did not provide sufficient information, the full article was assessed. Extracted data included the following: (1) author name(s) and year of publication; (2) animal model, age, sex, weight, and experimental model used; (3) injury defect size and location; (4) TGFβ-1 dose and carrier/delivery method; (5) experiment duration; and (6) main findings: outcomes of histological, radiological, and/or mechanical analyses. The selected studies were grouped and subsequently ordered based on model type (fracture, defect, and osteointegration), animal model, bone type, and TGFβ-1 carrier used.
Quality and risk of bias assessments
SYRCLE’s risk of bias tool was used for the quality assessment of the studies. 28 Two independent reviewers (S.T. and J.T.) answered 10 questions from SYRCLE’s tool with “Yes,” “No,” or “Unsure.” Discrepancies were resolved by a third independent reviewer (A.A.). Total bias free (“Yes”) and risk of bias (“No” or “Unsure”) scores were utilized to determine the overall quality of the studies included.
Results
The initial article searches in PubMed, Embase, and Web of Science resulted in 272 results. In addition, five articles were identified via reference lists. Thirty-three replicates were removed, and 214 articles were excluded as they did not fit our inclusion criteria, primarily as they did not deliver TGFβ-1 as a therapeutic. One article could not be retrieved, and full-text screening resulted in 24 studies included for review (Fig. 3). Thirteen of these articles used large animal long bone models, one article focused on a baboon calvaria model, two focused on canine mandibular models, one focused on a minipig mandibular model, and seven studies used rabbit long bone models (Fig. 4A). Risk of bias assessment (Table 1) resulted in six articles with “no risk of bias” scores greater than the “suspected and unknown risk of bias scores.” However, 18 articles had “unknown risk of bias scores” greater than “no risk of bias” scores. Therefore, the majority of the studies did not adequately address many components of their study that would clarify potential biases, and thus, should not be assumed “bias free.”
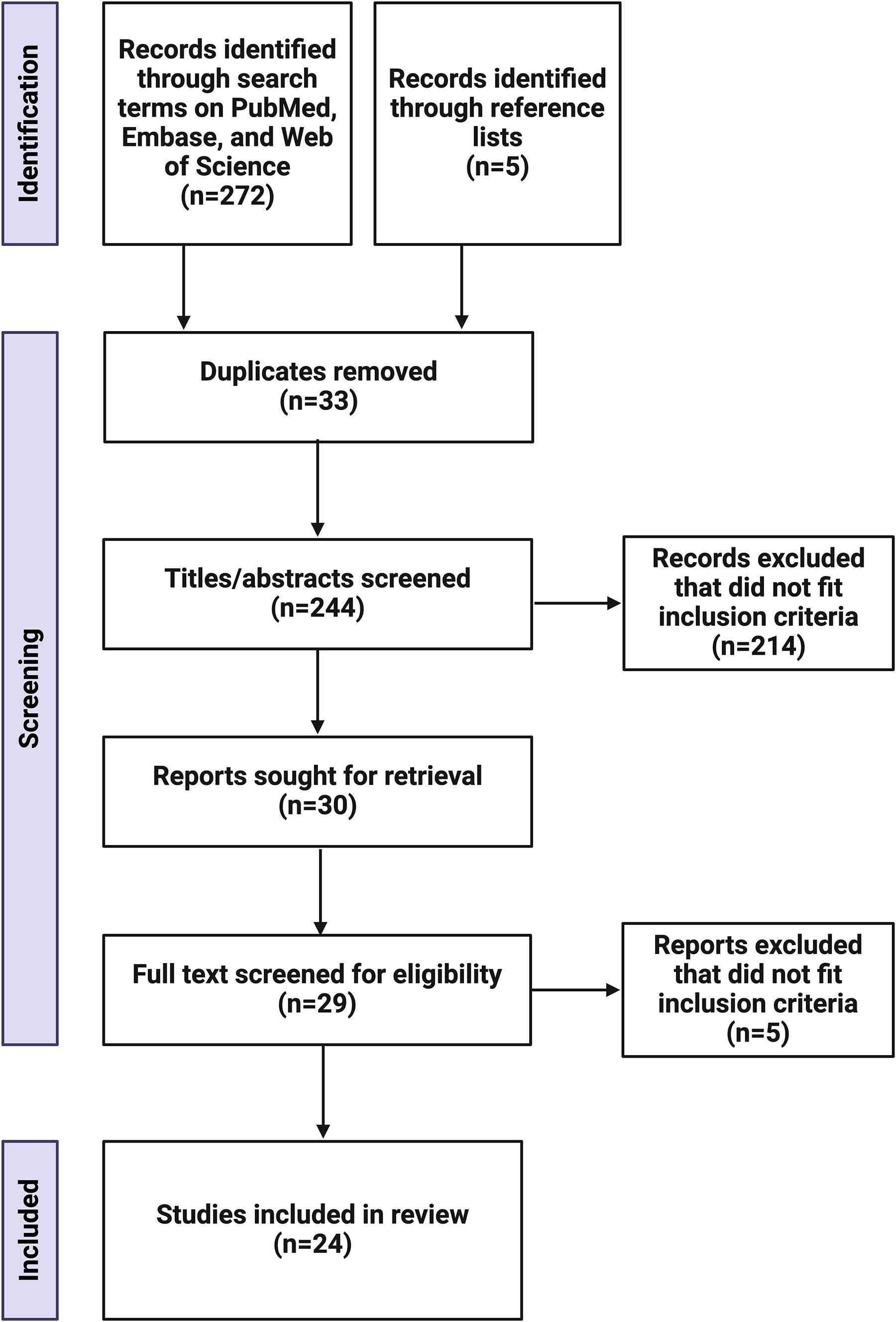
Flow diagram of study selection. *Image created using BioRender.com. Created in BioRender. Trbojevic, S. (2024) https://BioRender.com/g97u403.
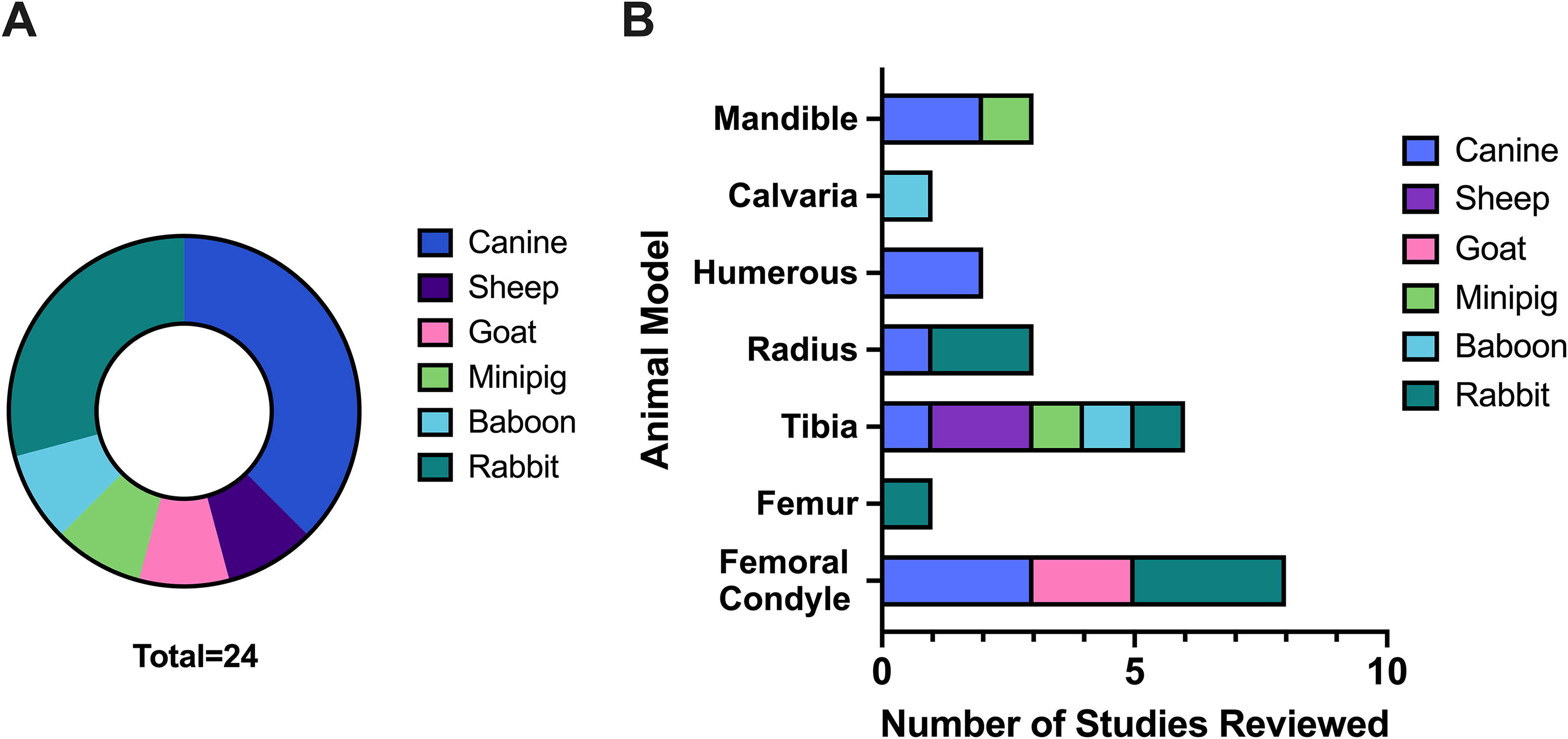
Categorization of experimental model parameters within the 24 studies reviewed.
SYRCLE’s Risk of Bias Assessment 28
Canine long bone studies
The majority of TGFβ-1 research of long bones in large animals has used canine models, in which seven studies used TGFβ-1 for fractures, bone defects, and osteointegration of implants or devices (Table 2). Regarding models of fracture nonunions using osteotomes, one study treated 3 mm radius osteotomes in canines with poly-
Large Animal Long Bone Studies
While the potential to enhance fracture healing is an exciting aspect of using TGFβ-1, another interesting application is for coatings on orthopedic hardware and devices to improve osteointegration. Calcium phosphate ceramic-coated strain gauges were treated with 0.5, 1, and 2.0 µg doses of TGFβ-1 and implanted on femoral condyles for 3, 6, or 12 weeks. The amount of new bone growth, bonding between the strain gauge and bone, and gauge accuracy were superior within the 1 µg TGFβ-treated gauge group at 6 and 12 weeks, though the other doses also had more bone formation when compared to the untreated control gauge. Notably, while the highest dose produced the most bone growth, the new bone was not present in the intended region prompting the authors to hypothesize that the TGFβ-1 diffused away from the implant. 50 In two similar studies, TCP-coated Ti alloy implants loaded with 0.3 µg TGFβ-1 were implanted with a 0.75 or 2 mm gap (between the implant and bone) in cylindrical femoral condyle defects. This resulted in a 16% bone volume increase in the group with a 0.75 mm gap and 45.5% bone volume increase in the group with a 2 mm gap when compared to untreated control implants at 6 weeks.34,51 In cylindrical humeral defects treated with either 0.3 or 3.0 µg TGFβ-1 loaded hydroxyapatite (HA)-coated Ti implants with a 2 mm gap, low dose of TGFβ-1 led to a 36% increase in bone coverage and 22% increase in gap healing when compared to the unloaded control group. However, while bone growth showed a significant increase, mechanical properties were not significantly different than the control. 52 Another study treated cylindrical humeral defects with either 120 or 335 µg TGFβ-1 coated HA/TCP-Ti implants with a 3 mm gap. The low dose resulted in a 222% increase in bone volume ingrowth compared to control implants at 4 weeks. 46 From these studies, it is apparent that the largest TGFβ-1 dose may not be the most beneficial.
Overall, these canine long bone repair studies showed that TGFβ-1 incorporated into DBM, scaffolds, and devices improved bone healing without evidence of heterotopic ossification. It is likely that the best carrier method for large bone defects needs to be developed to maintain the bioactivity of TGFβ-1 and have an effective release profile. It is important to note that the largest dose of TGFβ-1 used for large bone fracture repair in canines was only 10 ng, while the largest dose used for adsorption onto devices was 335 µg.32,51 Future studies should determine the impact of higher doses of TGFβ-1 on large bone defects.
Other large animal models with long bone studies
Aside from the canine models, six studies used other large animal models to research TGFβ-1 on long bone regeneration (Table 2). Regarding segmental defects, sheep tibial defects, 2.5 cm in size, were filled with Guanine-DBM loaded with 750 µg TGFβ-1. Twelve weeks postrepair, densely mineralized bone was observed, while no bony bridging of the defect occurred from implants of the DBM carrier alone. Four-point bending tests also supported evidence of regenerated bone with yield curves similar to native bone. 35 A study in baboons repaired a 5.3 mm cylindrical tibial defect with 1 µg or 10 µg TGFβ-1 encapsulated in gelatin within a methylcellulose carrier. After 21 days, both TGFβ-1 groups formed less bone volume than unloaded control defects. However, TGFβ-1 induced substantially more osteoblast proliferation compared to controls, and the authors believe this is due to sampling at an inadequate early time point. 29 A second study in sheep investigated the healing of a 3 cm tibial osteotomy in which some regenerative tissues were allowed to form before treatment. A porous polylactide membrane coated with 100 µg insulin-like growth factor-1 (IGF-1) and 20 µg TGFβ-1 or autologous cancellous bone supplemented with the coated membrane was compared to unloaded membranes or autologous cancellous bone. After 8 weeks, both groups treated with growth factors showed evidence of accelerated bone regeneration complete with woven bone, lamellar bone, and intact periosteum seen in histology. No evidence of heterotopic ossification was found. 30
Regarding osteointegration studies, 4 mm goat femoral condyle cylindrical defects were treated with CaP-coated, Ti alloy implants, loaded or not loaded with 1 µg TGFβ-1. Bone healing and bone contact with the implant were significantly enhanced by implants coated with CaP, but the addition of TGFβ-1 did not yield a notable difference. 42 The ineffectiveness of TGFβ-1 may be due to a burst release of the growth factor, limiting its retainment in the defect site. A study in goats aimed to accelerate bone growth on Ti dental screws with either 0.5 µg TGFβ-1 or 1 µg TGFβ-1 adsorbed to the surface and implanted in sheep femoral condyles. After 6 weeks, histology showed that control implants had the highest amount of bone-implant contact, while the implant loaded with 1 µg TGFβ-1 had the lowest amount. Both doses of TGFβ-1 resulted in fibrous tissue growth around the implant. The authors postulated that this was due to over-accelerated matrix growth during the inflammatory phase of healing. 36
Regarding fractures, a group treated Yucatan minipig 1 mm midshaft tibial osteotomies with Ti intramedullary nails coated with poly(
Across all large animal long bone studies, the majority of studies focused on osteointegration of implants or devices, highlighting the deficit of studies utilizing TGFβ-1 for improving larger bone injuries with insufficient biological environments. It should be emphasized that the TGFβ-1 doses ranged from 0.3 to 335 µg for osteointegration, 1 to 750 µg for defect repair, and 10 ng for fracture repair. Three of four studies that did not report successful results used TGFβ-1 doses under 1 µg. Therefore, while most articles report improved results from TGFβ-1, future research should focus on increasing fracture and defect studies and optimizing fracture growth factor dosage.
Large animal calvaria and mandible studies
The capacity of TGFβ-1 to enhance the healing of defects in craniofacial bones in large animals is less studied (Table 3). One group repaired critical
Large Animal Calvaria and Mandible Studies
Within the large animal calvaria and mandible repair studies, only one group used TGFβ-1 to repair a calvaria defect, while three used TGFβ-1 to repair mandibular defects. The only TGFβ-1 doses used for calvaria were 5 or 15 mg doses in combination with BMP7. TGFβ-1 doses for mandibular repair ranged from 1 to 250 µg. Alhough less studies are available for the repair of large animal calvaria and mandibles using TGFβ-1, all of the existing studies report successful defect repair. It is essential to highlight that these successful outcomes may be due to the enhanced regenerative capacity of these bones due to their surrounding environments and developmental mechanisms.13,14,53
Rabbit long bone studies
Because not many studies have used large animal models to research the influence of TGFβ-1 on long bone regeneration, it is useful to consider rabbit models from among other rodent models due to their similar bone anatomy (Table 4). One study reported filling 1.5 cm segmental radial defects with cancellous bone carriers containing 120 µg TGFβ-1. Defects were completely bridged by new, uniform bone at 20 weeks.
48
The same group then reported filling 1.5 cm segmental radial defects with cancellous bone carriers containing either 120 µg TGFβ-1, 12 mg BMP, or a composite containing both. All groups showed defects completely bridged with new bone at 16 weeks. However, the TGFβ-1 + BMP composite group showed regeneration of the most mature bone, with lamella and a recanalized medullar canal.48,49 Another group also compared TGFβ-1 to BMP-2, treating 4.5 mm femoral condyle cylindrical defects with porous poly(lactic-co-glycolic acid) (PLGA) cylinders overlaid with PLGA microspheres containing either 50 ng TGFβ-1, 2.5 µg BMP-2, or 5 µg BMP-2. Growth factor
Rabbit Long Bone Studies
All rabbit studies used TGFβ-1 to repair long bone defects, with doses ranging from 50 to 120 µg. No studies were found using a rabbit long bone model for fracture repair or device osteointegration, again emphasizing a lack of fracture repair models within TGFβ-1 research. While the majority of these studies report successful use of TGFβ-1 for defect repair, the only study that reported repair failure treated defects by local injection of TGFβ-1, which may have contributed to the unsuccessful outcomes.33,37,54
Discussion
Though varied models and devices were used, the research reviewed here largely shows positive outcomes for TGFβ-1 delivery, with 19 out of 24 articles concluding a beneficial effect of TGFβ-1 in bone repair. Some trends were identified, with 7 out of 13 large animal long bone studies using coated titanium implants for repair, while 3 of 7 rabbit long bone studies used some form of degradable polymer constructs, reflecting the maturity of the scaffold technology. TGFβ-1 doses in large animal long bone studies range from 0.005 to 750 µg with 0.3 µg the most common. Doses in large animal calvaria and mandible studies range from 1 to 5000 µg with 5 µg the most common. Doses in rabbit long bone studies range from 0.05 to 120 µg with the 0.1 and 120 µg used the most. The study duration or time point considered most often was 6 weeks but ranged between 4 weeks and one year. The most common defect model used was a femoral condyle defect, followed by tibial segmental defects (Fig. 4B).
Currently, there still is a lack of extensive research on TGFβ-1 for the repair of large animal long bone DNFs, which has an important clinical need due to the high prevalence of nonunion tibial fractures. From our review, most large animal long bone studies were done using coated metal implants or DBM. The field has expanded far beyond these methods for smaller animal models, such as the use of degradable hydrogels with or without the inclusion of cells. 55 However, large animal testing of only the most promising approaches is feasible for translation given the cost and ethical considerations. Further exploration into the compatibility and translatability of therapeutics in various defect sites is needed.
It is clear from the 22 years of research covered in this review that TGFβ-1 has the ability to enhance the bone healing process in defects in the long bones, calvaria, and mandibles of large animals and rabbits. However, no recommended carrier, dosage, or delivery profile exists. A change in carrier materials to polymeric substitutes can be seen around the end of the 20th century, when research shifted towards functional and less invasive therapeutics. As such, the use of metal alloys, DBM, and cancellous bone matrix carriers has decreased as popularity among tunable hydrogels has increased. Degradable hydrogels, barrier membranes, and other carriers that permit controlled release, such as microparticles, have shown efficacy in regenerating mature bone rather than the quick influx of immature fibrous tissues.54,56 Of note, the invasion of fibrous tissue has been the most prevalent cause of failed attempts discussed in this review. Therefore, it is crucial that a balance is found between fibrous tissue growth and new bone growth. Further, some high doses appeared to correlate with low osteogenesis and resorption around implants (e.g., strain gauges). Thus, potential immunomodulating effects of TGFβ-1 may be of concern, but addressable via control over the timing of release.
The developmental origin as well as the site of injury may impact regeneration outcomes. Long bones and parietal, temporal, and occipital calvaria are mesodermal in origin, while frontal calvaria and mandible are of neural crest origin. Long bones are developed via endochondral ossification where cartilage tissue remodels into bone (Fig. 1). Calvaria bone is developed via intramembranous ossification where bone forms directly from mesenchyme tissue (Fig. 2). The mandible bone is formed by both mechanisms depending on the location.5,14,53 Critical factors of long bone healing include both cartilage and bone-encoding genes, supply of bone marrow, and the periosteum which is a large source of chondrocytes and osteoprogenitor cells. Alternatively, critical factors of calvaria bone healing include a differing marrow microenvironment, gene expression from suture cells, and most importantly, the dura mater, which is a major source of mesenchymal cells and paracrine signaling. Critical factors of mandible remodeling include mesenchymal cells with greater osteogenic potential and high collagen content with immature crosslinks that enable a greater rate of remodeling. These specifics must be explored in terms of the cell source compatibility, mechanical requirements, growth factor dose response, and degradation and remodeling rate requirements across defect sites.14,53 Of course, these metrics may also vary between species as well, adding another level of complexity.
Limitations
Limitations of this review include an incomplete sampling of the literature, which was confined by our search terms, and an inability to make conclusive recommendations for future studies. The limitations provide further justification for the importance of additional exploration.
Conclusion
Though the osteogenic efficacy of TGFβ-1 is clear, controlled delivery is needed because it appears to have a bimodal dose response on the various physiological processes supporting bone healing. Direct comparison between studies is difficult due to differences in drug carrier, dose, delivery kinetics, defect type, and species. Overall, TGFβ-1 seemed effective when adsorbed onto metallic implants. It appears larger doses are more effective in healing the critically sized defects, for example, the 750 µg dose in the sheep tibia, rather than the 5 ng/mL doses used in the canine DBM study.35,43 It is apparent that some doses may elicit a deleterious effect such as fibrosis.50,52 Furthermore, it is evident that controlled release augments the potential of lower doses. 40 This may arise due to its pleiotropic effects on cellular response during healing. Nevertheless, supraphysiologic doses near the milligram scale showed no evidence of heterotopic ossification, unlike its family member BMP2. More large animal model testing is needed to isolate TGFβ-1’s role in the progenitor, vascular, bone, and immune cell compartments to develop therapeutics utilizing new, potentially safer molecules.
Footnotes
Authors’ Contributions
S.T., A.J.A., and J.M.T.: Conceptualization, data analysis, interpretation, and article writing. All authors revised and approved the final report.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Institute of Health (R01DE030296, F31DE031967-01A1) and the Department of Defense (W81XWH-16–1-0793).